
Stress Impairs Skin Barrier Function and Induces α2-3 Linked N-Acetylneuraminic Acid and Core 1 O-Glycans on Skin Mucins in Atlantic Salmon, Salmo salar

 ,
,
Abstract
:
1. Introduction
2. Methods
2.1. Experimental Design
2.2. Sampling
2.3. Blood Physiology
2.4. Plasma Cortisol
2.5. Skin Barrier Function and Morphology
2.6. Mucus Sampling and Mucin Isolation
2.7. O-Glycan Analysis by LC-MS
2.8. Statistical Analyses
3. Results
3.1. Effects of Chronic and Acute Stress on Plasma Parameters
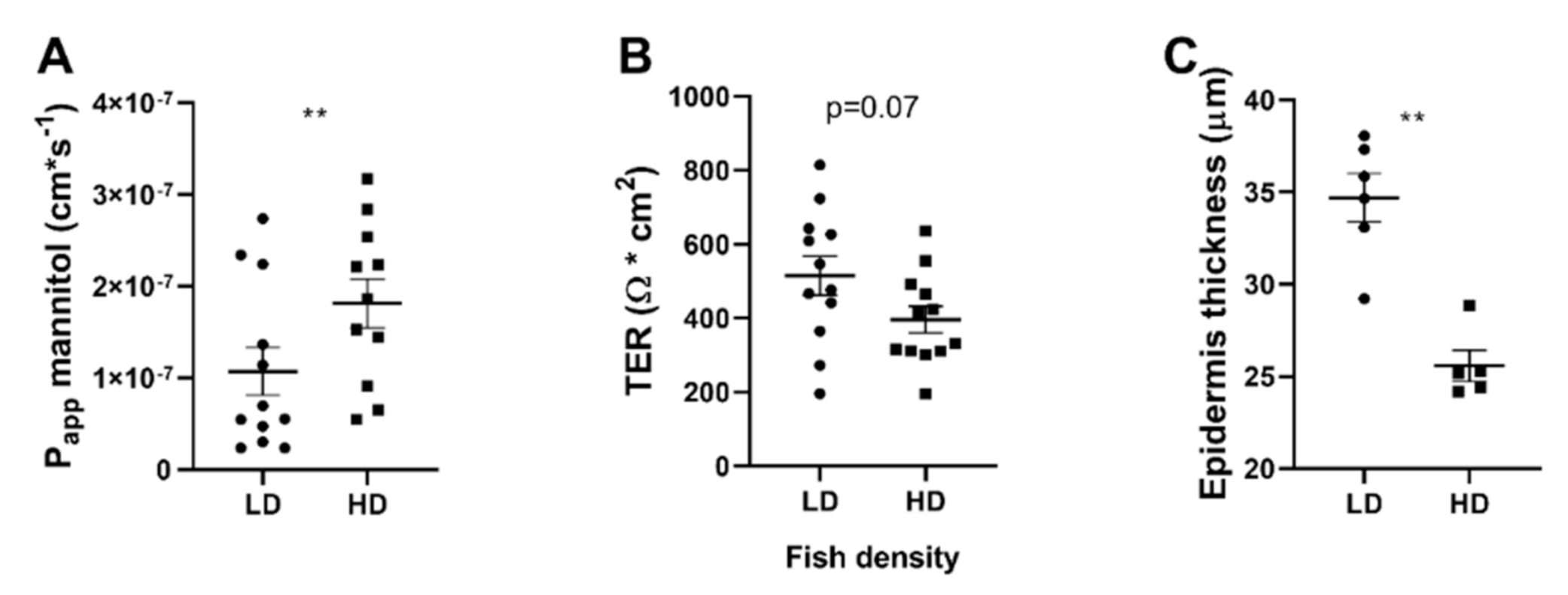
3.2. Effects of Chronic Stress on Skin Barrier Morphology and Function
3.3. LC-MS of Atlantic Salmon Skin O-Glycans Detected 81 Structures
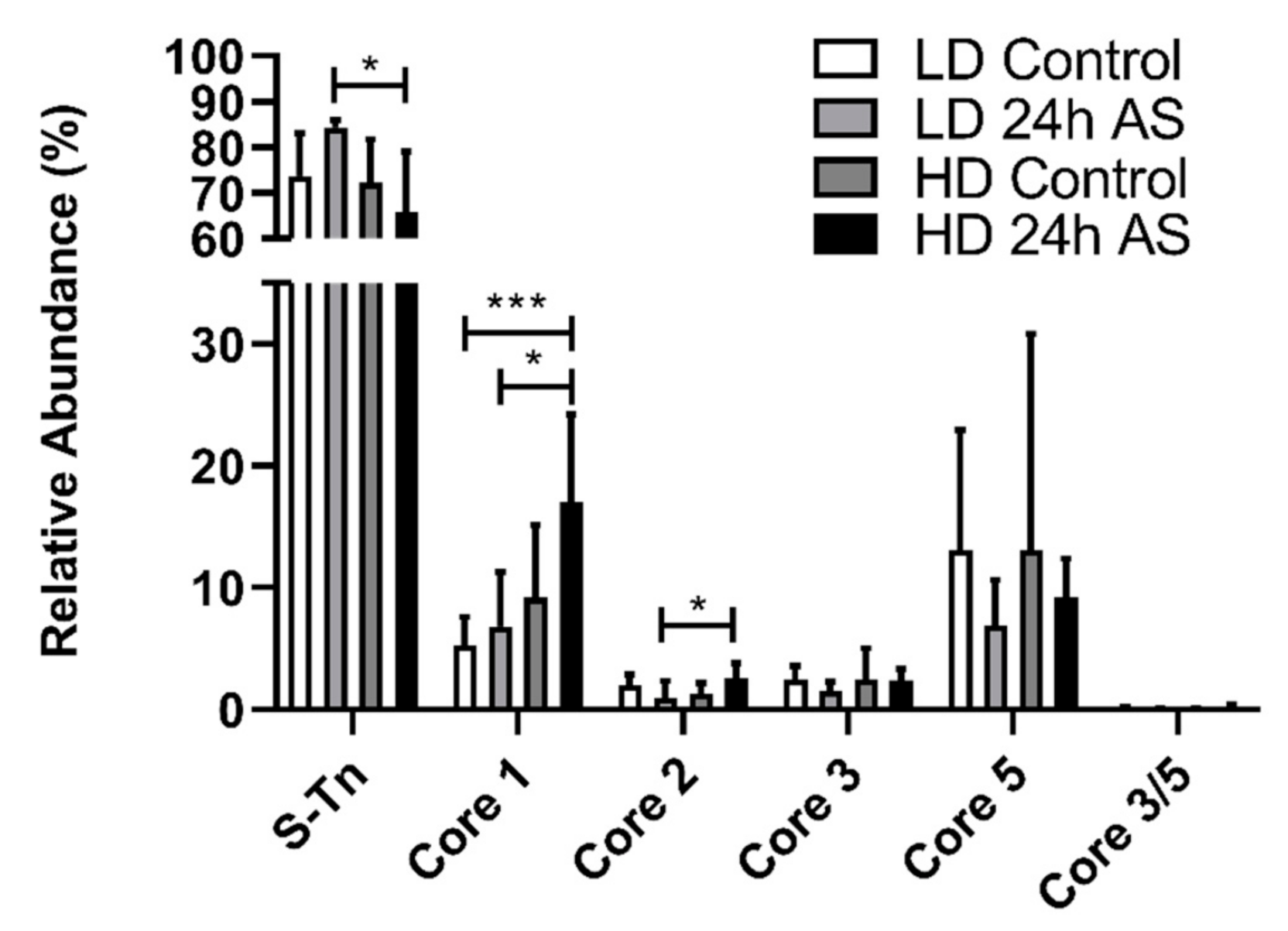
3.4. Core 1, 2, 3, and 5 Were Detected and Skin Mucins Express More Core 1 Structures When Atlantic Salmon Are Subjected to Both Chronic and Acute Stress
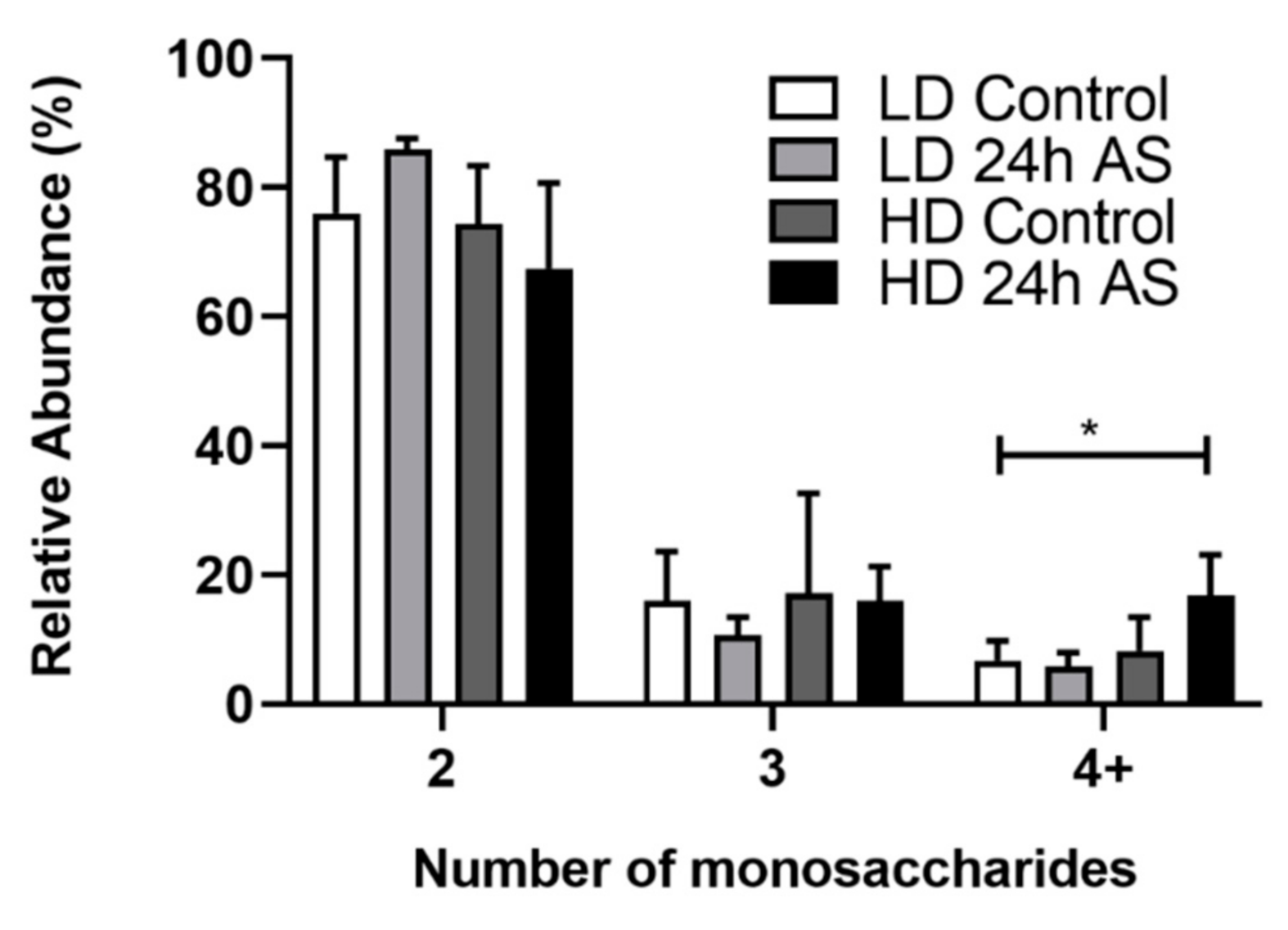
3.5. Fish Subjected to Both Chronic and Acute Stress Have a Higher Proportion of Large O-Glycan Structures
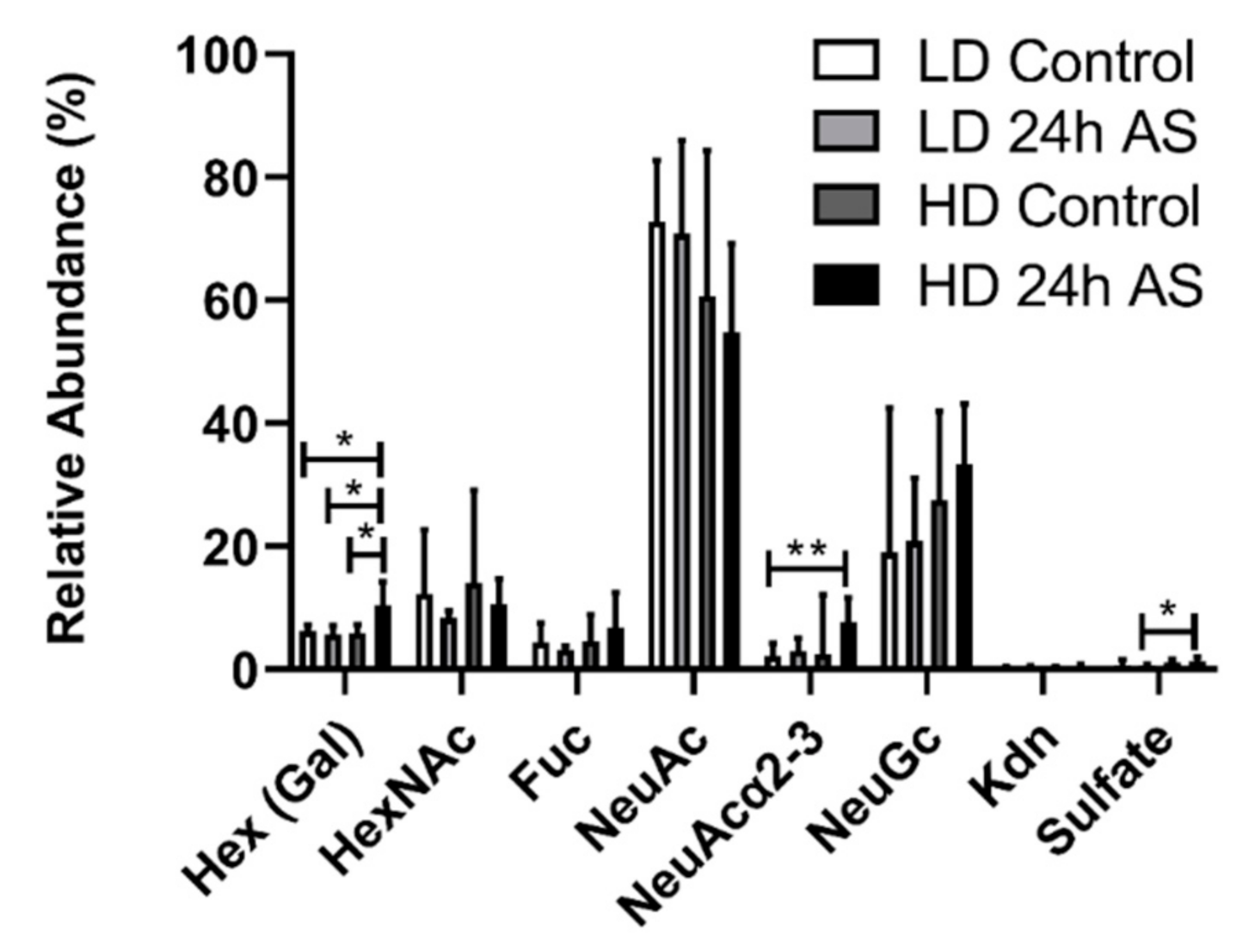
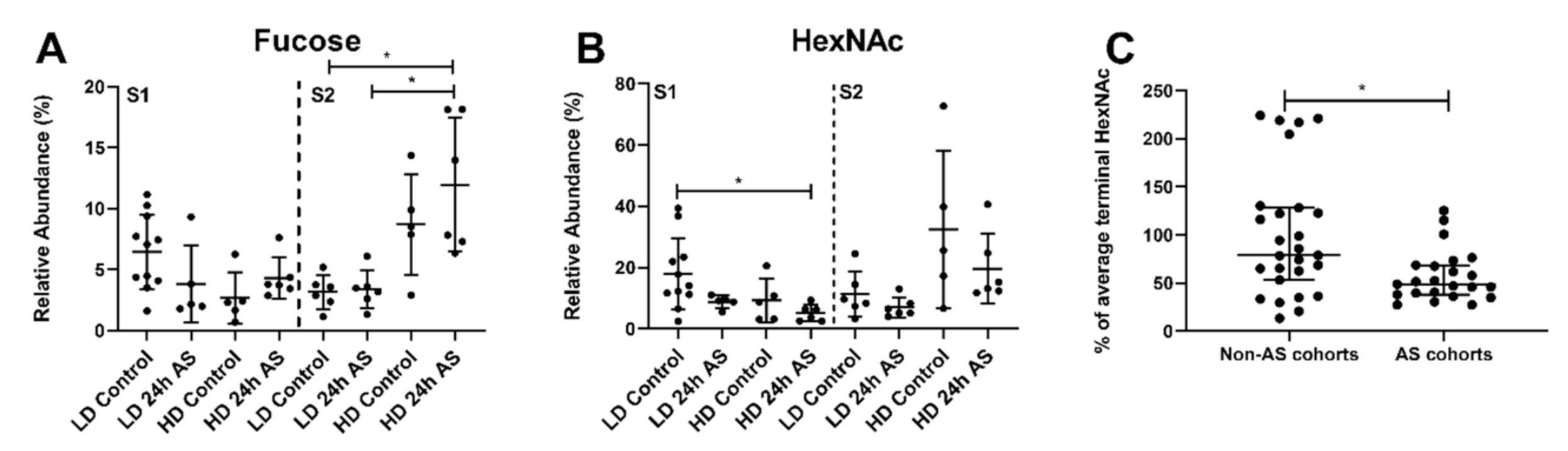
3.6. Fish Subjected to Both Chronic and Acute Stress Have an Increased Relative Abundance of Structures with Terminal Hexoses and α2-3 Linked NeuAc
3.7. Some Glycans Changed Differentially Depending on Sampling Time Point
3.8. Some O-Glycan Structures Have Potential as Stress Biomarkers in Atlantic Salmon
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2020, in The State of World Fisheries and Aquaculture (SOFIA); FAO: Rome, Italy, 2020; p. 244. [Google Scholar]
- Adams, C.E.; Turnbull, J.F.; Bell, A.S.; Bron, J.E.; Huntingford, F.A. Multiple determinants of welfare in farmed fish: Stocking density, disturbance, and aggression in Atlantic salmon (Salmo salar). Can. J. Fish. Aquat. Sci. 2007, 64, 336–344. [Google Scholar] [CrossRef]
- Ellis, T.; North, B.; Scott, A.P.; Bromage, N.R.; Porter, M.; Gadd, D. The relationships between stocking density and welfare in farmed rainbow trout. J. Fish Biol. 2002, 61, 493–531. [Google Scholar] [CrossRef]
- Calabrese, S.; Nilsen, T.; Kolarevic, J.; Ebbesson, L.; Pedrosa, C.; Fivelstad, S.; Hosfeld, C.; Stefansson, S.; Terjesen, B.; Takle, H.; et al. Stocking density limits for post-smolt Atlantic salmon (Salmo salar L.) with emphasis on production performance and welfare. Aquaculture 2017, 468, 363–370. [Google Scholar] [CrossRef] [Green Version]
- Pickering, A.D.; Stewart, A. Acclimation of the interrenal tissue of the brown trout, Salmo trutta L., to chronic crowding stress. J. Fish Biol. 1984, 24, 731–740. [Google Scholar] [CrossRef]
- Turnbull, J.; Bell, A.; Adams, C.; Bron, J.; Huntingford, F. Stocking density and welfare of cage farmed Atlantic salmon: Application of a multivariate analysis. Aquaculture 2005, 243, 121–132. [Google Scholar] [CrossRef]
- Sundh, H.; Kvamme, B.O.; Fridell, F.; E Olsen, R.; Ellis, T.; Taranger, G.L.; Sundell, K. Intestinal barrier function of Atlantic salmon (Salmo salar L.) post smolts is reduced by common sea cage environments and suggested as a possible physiological welfare indicator. BMC Physiol. 2010, 10, 22. [Google Scholar] [CrossRef] [Green Version]
- Sundh, H.; Finne-Fridell, F.; Ellis, T.; Taranger, G.; Niklasson, L.; Pettersen, E.; Wergeland, H.; Sundell, K. Reduced water quality associated with higher stocking density disturbs the intestinal barrier functions of Atlantic salmon (Salmo salar L.). Aquaculture 2019, 512, 734356. [Google Scholar] [CrossRef]
- Bonga, S.E.W. The stress response in fish. Physiol. Rev. 1997, 77, 591–625. [Google Scholar] [CrossRef]
- Barton, B.A.; Iwama, G.K. Physiological changes in fish from stress in aquaculture with emphasis on the response and effects of corticosteroids. Annu. Rev. Fish Dis. 1991, 1, 3–26. [Google Scholar] [CrossRef]
- Donaldson, E.M. The pituitary-interrenal axis as an indicator of stress in fish. Stress Fish 1981, 1981, 11–47. [Google Scholar]
- Schreck, C.B. Accumulation and long-term effects of stress in fish. Biol. Anim. Stress Basic Princ. Implic. Anim. Welf. 2009, 1, 147–158. [Google Scholar]
- Madaro, A.; E Olsen, R.; Kristiansen, T.S.; Ebbesson, L.O.; Flik, G.; Gorissen, M. A comparative study of the response to repeated chasing stress in Atlantic salmon (Salmo salar L.) parr and post-smolts. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2016, 192, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Madaro, A.; Olsen, R.E.; Kristiansen, T.S.; Ebbesson, L.O.E.; Nilsen, T.O.; Flik, G.; Gorissen, M. Stress in Atlantic salmon: Response to unpredictable chronic stress. J. Exp. Biol. 2015, 218, 2538–2550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Porchas, M.; Martinez-Cordova, L.R.; Ramos-Enriquez, R. Cortisol and Glucose: Reliable indicators of fish stress? Pan-Am. J. Aquat. Sci. 2009, 14, 158–178. [Google Scholar]
- Sundh, H.; Sundell, K.S. 7—Environmental impacts on fish mucosa A2—Beck, Benjamin H. In Mucosal Health in Aquaculture; Peatman, E., Ed.; Academic Press: San Diego, CA, USA, 2015; pp. 171–197. [Google Scholar]
- Elliott, D.G. THE SKIN|Functional Morphology of the Integumentary System in Fishes. In Encyclopedia of Fish Physiology; Farrell, A.P., Ed.; Academic Press: San Diego, CA, USA, 2011; pp. 476–488. [Google Scholar]
- Sveen, L.; Karlsen, C.; Ytteborg, E. Mechanical induced wounds in fish—A review on models and healing mechanisms. Rev. Aquac. 2020, 12, 2446–2465. [Google Scholar] [CrossRef]
- Micallef, G.; Bickerdike, R.; Reiff, C.; Fernandes, J.M.O.; Bowman, A.S.; Martin, S. Exploring the Transcriptome of Atlantic Salmon (Salmo salar) Skin, a Major Defense Organ. Mar. Biotechnol. 2012, 14, 559–569. [Google Scholar] [CrossRef]
- Hattrup, C.L.; Gendler, S.J. Structure and Function of the Cell Surface (Tethered) Mucins. Annu. Rev. Physiol. 2008, 70, 431–457. [Google Scholar] [CrossRef]
- Linden, S.K.; Sutton, P.; Karlsson, N.G.; Korolik, V.; McGuckin, M.A. Mucins in the mucosal barrier to infection. Mucosal Immunol. 2008, 1, 183–197. [Google Scholar] [CrossRef] [Green Version]
- Venkatakrishnan, V.; Padra, J.T.; Sundh, H.; Sundell, K.; Jin, C.; Langeland, M.; Carlberg, H.; Vidakovic, A.; Lundh, T.; Karlsson, N.G.; et al. Exploring the Arctic Charr Intestinal Glycome: Evidence of Increased N-Glycolylneuraminic Acid Levels and Changed Host–Pathogen Interactions in Response to Inflammation. J. Proteome Res. 2019, 18, 1760–1773. [Google Scholar] [CrossRef] [Green Version]
- Venkatakrishnan, V.; Quintana-Hayashi, M.P.; Mahu, M.; Haesebrouck, F.; Pasmans, F.; Lindén, S.K. Brachyspira hyodysenteriae Infection Regulates Mucin Glycosylation Synthesis Inducing an Increased Expression of Core-2 O-Glycans in Porcine Colon. J. Proteome Res. 2017, 16, 1728–1742. [Google Scholar] [CrossRef]
- Linden, S.; Mahdavi, J.; Semino-Mora, C.; Olsen, C.; Carlstedt, I.; Borén, T.; Dubois, A. Role of ABO Secretor Status in Mucosal Innate Immunity and H. pylori Infection. PLoS Pathog. 2008, 4, e2. [Google Scholar] [CrossRef] [PubMed]
- Padra, M.; Adamczyk, B.; Flahou, B.; Erhardsson, M.; Chahal, G.; Smet, A.; Jin, C.; Thorell, A.; Ducatelle, R.; Haesebrouck, F.; et al. Helicobacter suis infection alters glycosylation and decreases the pathogen growth inhibiting effect and binding avidity of gastric mucins. Mucosal Immunol. 2019, 12, 784–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quintana-Hayashi, M.P.; Venkatakrishnan, V.; Haesebrouck, F.; Lindén, S.K. Role of Sialic Acid inBrachyspira hyodysenteriaeAdhesion to Pig Colonic Mucins. Infect. Immun. 2019, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dabelic, S.; Flögel, M.; Maravić, G.; Lauc, G. Stress Causes Tissue-Specific Changes in the Sialyltransferase Activity. Z. Für Nat. C 2004, 59, 276–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, C.; Padra, J.T.; Sundell, K.; Sundh, H.; Karlsson, N.G.; Lindén, S.K. Atlantic Salmon Carries a Range of Novel O-Glycan Structures Differentially Localized on Skin and Intestinal Mucins. J. Proteome Res. 2015, 14, 3239–3251. [Google Scholar] [CrossRef] [PubMed]
- Benktander, J.; Venkatakrishnan, V.; Padra, J.T.; Sundh, H.; Sundell, K.; Murugan, A.V.M.; Maynard, B.; Lindén, S.K. Effects of Size and Geographical Origin on Atlantic salmon, Salmo salar, Mucin O-Glycan Repertoire. Mol. Cell. Proteom. 2019, 18, 1183–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padra, J.T.; Sundh, H.; Sundell, K.; Venkatakrishnan, V.; Jin, C.; Samuelsson, T.; Karlsson, N.G.; Linden, S.K. Aeromonas salmonicida Growth in Response to Atlantic Salmon Mucins Differs between Epithelial Sites, Is Governed by Sialylated and N-Acetylhexosamine-Containing O-Glycans, and Is Affected by Ca2+. Infect. Immun. 2017, 85. [Google Scholar] [CrossRef] [Green Version]
- Padra, J.T.; Sundh, H.; Jin, C.; Karlsson, N.G.; Sundell, K.; Linden, S.K. Aeromonas salmonicida Binds Differentially to Mucins Isolated from Skin and Intestinal Regions of Atlantic Salmon in anN-Acetylneuraminic Acid-Dependent Manner. Infect. Immun. 2014, 82, 5235–5245. [Google Scholar] [CrossRef] [Green Version]
- Padra, J.T.; Murugan, A.V.M.; Sundell, K.; Sundh, H.; Benktander, J.; Linden, S.K. Fish pathogen binding to mucins from Atlantic salmon and Arctic char differs in avidity and specificity and is modulated by fluid velocity. PLoS ONE 2019, 14, e0215583. [Google Scholar] [CrossRef]
- Skonhoft, A. National Aquaculture Legislation Overview (NALO) Fact Sheets. In National Aquaculture Legislation Overview; FAO Fisheries Division [online]: Rome, Italy, 2010. [Google Scholar]
- RSPCA. RSPCA Welfare Standards for Farmed Atlantic Salmon; Royal Society for the Prevention of Cruelty to Animals: West Sussex, UK, 2018. [Google Scholar]
- Sveen, L.R.; Timmerhaus, G.; Torgersen, J.S.; Ytteborg, E.; Jørgensen, S.M.; Handeland, S.; Stefansson, S.O.; Nilsen, T.O.; Calabrese, S.; Ebbesson, L.; et al. Impact of fish density and specific water flow on skin properties in Atlantic salmon (Salmo salar L.) post-smolts. Aquaculture 2016, 464, 629–637. [Google Scholar] [CrossRef] [Green Version]
- Terjesen, B.F.; Summerfelt, S.T.; Nerland, S.; Ulgenes, Y.; Fjæra, S.O.; Reiten, B.K.; Selset, R.; Kolarevic, J.; Brunsvik, P.; Bæverfjord, G.; et al. Design, dimensioning, and performance of a research facility for studies on the requirements of fish in RAS envi-ronments. Aquac. Eng. 2013, 54, 49–63. [Google Scholar] [CrossRef] [Green Version]
- Kolarevic, J.; Baeverfjord, G.; Takle, H.; Ytteborg, E.; Reiten, B.K.M.; Nergård, S.; Terjesen, B.F. Performance and welfare of Atlantic salmon smolt reared in recirculating or flow through aquaculture systems. Aquaculture 2014, 432, 15–25. [Google Scholar] [CrossRef]
- Van den Bos, R.; Althuizen, J.; Tschigg, K.; Bomert, M.; Zethof, J.; Filk, G.; Gorissen, M. Early life exposure to cortisol in zebrafish (Danio rerio): Similarities and differences in behaviour and physiology between larvae of the AB and TL strains. Behav. Pharmacol. 2019, 30, 260–271. [Google Scholar] [CrossRef] [PubMed]
- Grass, G.M.; Sweetana, S.A. In Vitro Measurement of Gastrointestinal Tissue Permeability Using a New Diffusion Cell. Pharm. Res. 1988, 5, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Sundell, K.; Jutfelt, F.; Ágústsson, T.; Olsen, R.-E.; Sandblom, E.; Hansen, T.; Björnsson, B.T. Intestinal transport mechanisms and plasma cortisol levels during normal and out-of-season parr–smolt transformation of Atlantic salmon, Salmo salar. Aquaculture 2003, 222, 265–285. [Google Scholar] [CrossRef]
- Sundell, K.; Sundh, H. Intestinal fluid absorption in anadromous salmonids: Importance of tight junctions and aquaporins. Front. Physiol. 2012, 3, 388. [Google Scholar] [CrossRef] [Green Version]
- Bjarnason, I.; MacPherson, A.; Hollander, D. Intestinal permeability: An overview. Gastroenterology 1995, 108, 1566–1581. [Google Scholar] [CrossRef] [PubMed]
- Schulz, B.L.; Packer, N.H.; Karlsson, N.G. Small-Scale Analysis of O-Linked Oligosaccharides from Glycoproteins and Mucins Separated by Gel Electrophoresis. Anal. Chem. 2002, 74, 6088–6097. [Google Scholar] [CrossRef]
- Hayes, C.A.; Karlsson, N.G.; Struwe, W.B.; Lisacek, F.; Rudd, P.M.; Packer, N.H.; Campbell, M.P. UniCarb-DB: A database resource for glycomic discovery. Bioinformatics 2011, 27, 1343–1344. [Google Scholar] [CrossRef] [Green Version]
- Varki, A.; Cummings, R.D.; Aebi, M.; Packer, N.H.; Seeberger, P.H.; Esko, J.D.; Stanley, P.; Hart, G.W.; Darvill, A.G.; Kinoshita, T.; et al. Symbol Nomenclature for Graphical Representations of Glycans. Glycobiology 2015, 25, 1323–1324. [Google Scholar] [CrossRef] [Green Version]
- Sharba, S.; Venkatakrishnan, V.; Padra, M.; Winther, M.; Gabl, M.; Sundqvist, M.; Wang, J.; Forsman, H.; Linden, S.K. Formyl peptide receptor 2 orchestrates mucosal protection against Citrobacter rodentium infection. Virulence 2019, 10, 610–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mota, V.C.; Nilsen, T.O.; Gerwins, J.; Gallo, M.; Kolarevic, J.; Krasnov, A.; Terjesen, B.F. Molecular and physiological responses to long-term carbon dioxide exposure in Atlantic salmon (Salmo salar). Aquaculture 2020, 519, 734715. [Google Scholar] [CrossRef]
- Brauner, C.; Thorarensen, H.; Gallaugher, P.; Farrell, A.P.; Randall, D.J. CO2 transport and excretion in rainbow trout (Oncorhynchus mykiss) during graded sustained exercise. Respir. Physiol. 2000, 119, 69–82. [Google Scholar] [CrossRef]
- Wood, C.M.; Turner, J.D.; Graham, M.S. Why do fish die after severe exercise? J. Fish Biol. 1983, 22, 189–201. [Google Scholar] [CrossRef]
- Quintana-Hayashi, M.P.; Padra, M.; Padra, J.T.; Benktander, J.; Lindén, S.K. Mucus-Pathogen Interactions in the Gastrointestinal Tract of Farmed Animals. Microorganisms 2018, 6, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skoog, E.C.; Lindberg, M.; Linden, S.K. Strain-Dependent Proliferation in Response to Human Gastric Mucin and Adhesion Properties of Helicobacter pylori are not Affected by Co-isolated Lactobacillus sp. Helicobacter 2011, 16, 9–19. [Google Scholar] [CrossRef]
- Skoog, E.C.; Sjöling, Å.; Navabi, N.; Holgersson, J.; Lundin, S.B.; Lindén, S.K. Human Gastric Mucins Differently Regulate Helicobacter pylori Proliferation, Gene Expression and Interactions with Host Cells. PLoS ONE 2012, 7, e36378. [Google Scholar] [CrossRef] [Green Version]
- Minniti, G.; Hagen, L.H.; Porcellato, D.; Jørgensen, S.M.; Pope, P.B.; Vaaje-Kolstad, G. The Skin-Mucus Microbial Community of Farmed Atlantic Salmon (Salmo salar). Front. Microbiol. 2017, 8, 2043. [Google Scholar] [CrossRef] [Green Version]
- Navabi, N.; Johansson, M.E.V.; Raghavan, S.; Linden, S.K. Helicobacter pylori Infection Impairs the Mucin Production Rate and Turnover in the Murine Gastric Mucosa. Infect. Immun. 2013, 81, 829–837. [Google Scholar] [CrossRef] [Green Version]
- Sharba, S.; Navabi, N.; Padra, M.; Persson, J.A.; Quintana-Hayashi, M.P.; Gustafsson, J.K.; Szeponik, L.; Venkatakrishnan, V.; Sjöling, Å.; Nilsson, S.; et al. Interleukin 4 induces rapid mucin transport, increases mucus thickness and quality and decreases colitis and Citrobacter rodentium in contact with epithelial cells. Virulence 2019, 10, 97–117. [Google Scholar] [CrossRef] [Green Version]
- Karlsen, C.; Ytteborg, E.; Timmerhaus, G.; Høst, V.; Handeland, S.; Jørgensen, S.M.; Krasnov, A. Atlantic salmon skin barrier functions gradually enhance after seawater transfer. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Vindas, M.A.; Gorissen, M.; Höglund, E.; Flik, G.; Tronci, V.; Damsgård, B.; Thörnqvist, P.-O.; Nilsen, T.O.; Winberg, S.; Øverli, Ø.; et al. How do individuals cope with stress? Behavioural, physiological and neuronal differences between proactive and reactive coping styles in fish. J. Exp. Biol. 2017, 220, 1524–1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pottinger, T.G.; Carrick, T. Stress Responsiveness Affects Dominant–Subordinate Relationships in Rainbow Trout. Horm. Behav. 2001, 40, 419–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day of Experiment | Treatment | Temp (°C) | O2 (% sat) | Conductivity (mS/cm) | Salinity (ppt) | pH | CO2 (mg/L) | TAN (mg/L) | Turbidity (NTU) |
|---|---|---|---|---|---|---|---|---|---|
| Day 82 | LD | 14.0 ± 0.0 | 91.7 ± 6.4 | 52.3 ± 0.1 | 33.8 ± 0.0 | 7.5 ± 0.1 * | 12.8 ± 0.9 * | 0.4 ± 0.1 | 0.2 ± 0.0 |
| HD | 14.0 ± 0.1 | 93.0 ± 7.0 | 52.3 ± 0.0 | 33.8 ± 0.0 | 7.1 ± 0.2 * | 22.1 ± 2.8 * | 0.6 ± 0.1 | 0.3 ± 0.1 | |
| Day 116 | LD | 13.0 ± 0.0 | 90.0 ± 1.0 | 49.8 ± 0.0 | 31.9 ± 0.0 | 7.5 ± 0.2 | 10.7 ± 1.0 * | 0.4 ± 0.1 | 0.2 ± 0.0 |
| HD | 13.0 ± 0.1 | 92.7 ± 1.5 | 49.8 ± 0.0 | 31.9 ± 0.0 | 7.2 ± 0.2 | 16.8 ± 3.5 * | 0.5 ± 0.1 | 0.2 ± 0.0 | |
| Day 172 | LD | 13.1 ± 0.0 | 94.0 ± 1.0 | 49.4 ± 0.0 | 31.7 ± 0.1 | 7.6 ± 0.2 | 12.3 ± 1.6 | 0.5 ± 0.0 | 0.2 ± 0.0 |
| HD | 13.0 ± 0.1 | 97.0 ± 2.0 | 49.4 ± 0.0 | 31.7 ± 0.0 | 7.5 ± 0.1 | 14.0 ± 0.4 | 0.5 ± 0.0 | 0.2 ± 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benktander, J.; Sundh, H.; Sundell, K.; Murugan, A.V.M.; Venkatakrishnan, V.; Padra, J.T.; Kolarevic, J.; Terjesen, B.F.; Gorissen, M.; Lindén, S.K. Stress Impairs Skin Barrier Function and Induces α2-3 Linked N-Acetylneuraminic Acid and Core 1 O-Glycans on Skin Mucins in Atlantic Salmon, Salmo salar. Int. J. Mol. Sci. 2021, 22, 1488. https://doi.org/10.3390/ijms22031488
Benktander J, Sundh H, Sundell K, Murugan AVM, Venkatakrishnan V, Padra JT, Kolarevic J, Terjesen BF, Gorissen M, Lindén SK. Stress Impairs Skin Barrier Function and Induces α2-3 Linked N-Acetylneuraminic Acid and Core 1 O-Glycans on Skin Mucins in Atlantic Salmon, Salmo salar. International Journal of Molecular Sciences. 2021; 22(3):1488. https://doi.org/10.3390/ijms22031488
Chicago/Turabian StyleBenktander, John, Henrik Sundh, Kristina Sundell, Abarna V. M. Murugan, Vignesh Venkatakrishnan, János Tamás Padra, Jelena Kolarevic, Bendik Fyhn Terjesen, Marnix Gorissen, and Sara K. Lindén. 2021. "Stress Impairs Skin Barrier Function and Induces α2-3 Linked N-Acetylneuraminic Acid and Core 1 O-Glycans on Skin Mucins in Atlantic Salmon, Salmo salar" International Journal of Molecular Sciences 22, no. 3: 1488. https://doi.org/10.3390/ijms22031488
APA StyleBenktander, J., Sundh, H., Sundell, K., Murugan, A. V. M., Venkatakrishnan, V., Padra, J. T., Kolarevic, J., Terjesen, B. F., Gorissen, M., & Lindén, S. K. (2021). Stress Impairs Skin Barrier Function and Induces α2-3 Linked N-Acetylneuraminic Acid and Core 1 O-Glycans on Skin Mucins in Atlantic Salmon, Salmo salar. International Journal of Molecular Sciences, 22(3), 1488. https://doi.org/10.3390/ijms22031488