
Calprotectin in Lung Diseases

 ,
, 

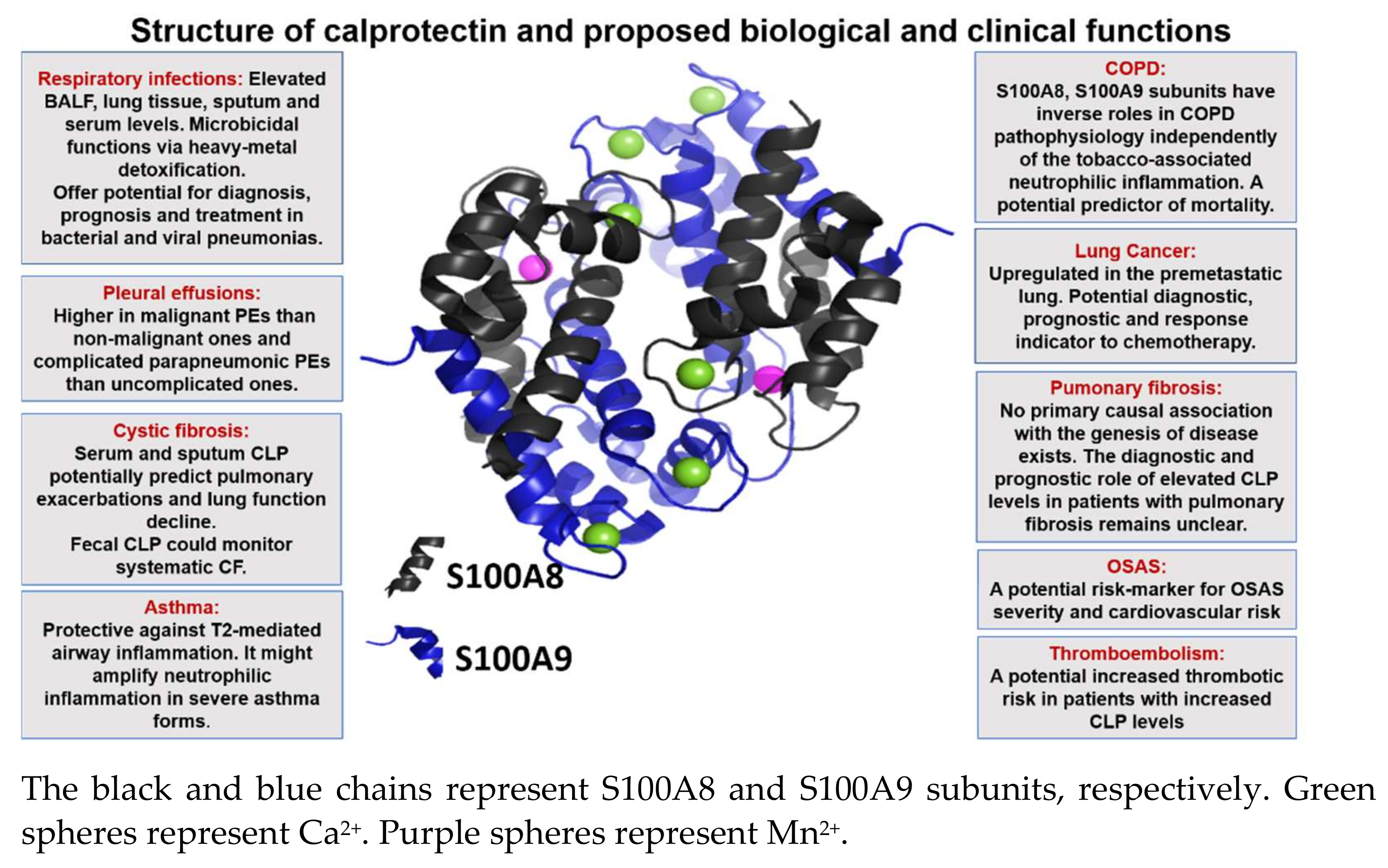{kind=link}
Abstract
:1. Introduction
2. The Expression of CLP in Lung Tissue and Potential Mechanisms of Extracellular Release of Human Phagocyte CLP
3. CLP’s Concentration Variations
4. CLP in Respiratory Infections
5. CLP in Serous Effusions
6. CLP in Cystic Fibrosis and Non-Cystic Fibrosis Bronchiectasis
7. CLP in Asthma
8. CLP in Chronic Obstructive Pulmonary Disease
9. CLP in Lung Cancer
10. CLP and Metastatic Cancers in the Lung
11. CLP in Idiopathic Pulmonary Fibrosis and Other Fibrotic Diseases
12. CLP in Obstructive Sleep Apnea Syndrome
13. CLP in Pulmonary Embolism and Pulmonary Hypertension
14. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Goebeler, M.; Roth, J.; van den Bos, C.; Ader, G.; Sorg, C. Increase of calcium levels in epithelial cells induces translocation of calcium-binding proteins migration inhibitory factor-related protein 8 (MRP8) and MRP14 to keratin intermediate filaments. Biochem. J. 1995, 309, 419–424. [Google Scholar] [CrossRef] [Green Version]
- Edgeworth, J.; Gorman, M.; Bennett, R.; Freemont, P.; Hogg, N. Identification of p8,14 as a highly abundant heterodimeric calcium binding protein complex of myeloid cells. J. Biol. Chem. 1991, 266, 7706–7713. [Google Scholar] [CrossRef]
- Ometto, F.; Friso, L.; Astorri, D.; Botsios, C.; Raffeiner, B.; Punzi, L.; Doria, A. Calprotectin in rheumatic diseases. Exp. Biol. Med. 2016, 242, 859–873. [Google Scholar] [CrossRef] [PubMed]
- Dale, I.; Brandtzaeg, P.; Fagerhol, M.K.; Scott, H. Distribution of a new myelomonocytic antigen (L1) in human peripheral blood leukocytes: Immunofluorescence and immunoperoxidase staining features in comparison with lysozyme and lactoferrin. Am. J. Clin. Pathol. 1985, 84, 24–34. [Google Scholar] [CrossRef]
- Shabani, F.; Farasat, A.; Mahdavi, M.; Gheibi, N. Calprotectin (S100A8/S100A9): A key protein between inflammation and cancer. Inflamm. Res. 2018, 67, 801–812. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Miller, A.L.; Rebelatto, M.; Brewah, Y.; Rowe, D.C.; Clarke, L.; Czapiga, M.; Rosenthal, K.; Imamichi, T.; Chen, Y.; et al. S100A9 induced in-flammatory responses are mediated by distinct damage associated molecular patterns (DAMP) receptors in vitro and in vivo. PLoS ONE 2015, 10, e0115828. [Google Scholar]
- Sun, Y.; Lu, Y.; Engeland, C.G.; Gordon, S.C.; Sroussi, H. The anti-oxidative, anti-inflammatory, and protective effect of S100A8 in endotoxemic mice. Mol. Immunol. 2013, 53, 443–449. [Google Scholar] [CrossRef] [Green Version]
- Manni, M.L.; Alcorn, J.F. Calprotectin-g the lung during type 2 allergic airway inflammation. Am. J. Respir. Cell Mol. Biol. 2019, 61, 405–407. [Google Scholar] [CrossRef]
- Palmer, L.D.; Maloney, K.N.; Boyd, K.L.; Goleniewska, A.K.; Toki, S.; Maxwell, C.N.; Chazin, W.J.; Peebles, R.S., Jr.; Newcomb, D.C.; Skaar, E.P. The innate immune protein S100A9 protects from T-helper cell type 2-mediated allergic airway inflam-mation. Am. J. Respir. Cell Mol. Biol. 2019, 61, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Vogl, T.J.; Tenbrock, K.; Ludwig, S.; Leukert, N.; Ehrhardt, C.; Van Zoelen, M.A.D.; Nacken, W.; Foell, D.; Van Der Poll, T.; Sorg, C.; et al. Mrp8 and Mrp14 are endogenous activators of Toll-like receptor 4, promoting lethal, endotoxin-induced shock. Nat. Med. 2007, 13, 1042–1049. [Google Scholar] [CrossRef] [PubMed]
- Røseth, A.G.; Fagerhol, M.K.; Aadland, E.; Schjønsby, H. Assessment of the neutrophil dominating protein calprotectin in feces: A methodologic study. Scand. J. Gastroenterol. 1992, 27, 793–798. [Google Scholar] [CrossRef] [PubMed]
- Berntzen, H.B.; Ölmez, Ü.; Fagerhol, M.K.; Munthe, E. The leukocyte protein L1 in plasma and synovial fluid from patients with rheumatoid arthritis and osteoarthritis. Scand. J. Rheumatol. 1991, 20, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Bartakova, E.; Stefan, M.; Stranikova, A.; Pospíšilová, L.; Arientová, S.; Beran, O.; Blahutová, M.; Máca, J.; Holub, M. Calprotectin and calgranulin C serum levels in bacterial sepsis. Diagn. Microbiol. Infect. Dis. 2019, 93, 219–226. [Google Scholar] [CrossRef]
- Sipponen, T.; Kolho, K.-L. Fecal calprotectin in diagnosis and clinical assessment of inflammatory bowel disease. Scand. J. Gastroenterol. 2015, 50, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Tyden, H.; Lood, C.; Gullstrand, B.; Jönsen, A.; Ivars, F.; Leanderson, T.; Bengtsson, A.A. Pro-inflammatory S100 proteins are associated with glomerulonephritis and anti-dsDNA antibodies in systemic lupus erythematosus. Lupus 2017, 26, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Zha, X.; Li, C.; Jia, Y.; Zhu, L.; Guo, J.; Su, Y. Serum calprotectin–a promising diagnostic marker for adult-onset Still’s disease. Clin. Rheumatol. 2016, 35, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Morrow, D.A.; Wang, Y.; Croce, K.; Sakuma, M.; Sabatine, M.S.; Gao, H.; Pradhan, A.D.; Healy, A.M.; Buros, J.; McCabe, C.H.; et al. Myeloid-related protein 8/14 and the risk of cardiovascular death or my-ocardial infarction after an acute coronary syndrome in the pravastatin or atorvastatin evaluation and infection therapy: Thrombolysis in myocardial infarction (PROVE ITTIMI 22) trial. Am. Heart J. 2008, 155, 49–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Song, R.; Wang, Z.; Jing, Z.; Wang, S.; Ma, J. S100A8/A9 in inflammation. Front. Immunol. 2018, 9, 1298. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Li, F.; Wang, J.; Ma, J.; Sheng, X. Upregulation of calprotectin in mild IgE-mediated ovalbumin hypersensitivity. Oncotarget 2017, 8, 7342–37354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henke, M.O.; Renner, A.; Rubin, B.K.; Gyves, J.I.; Lorenz, E.; Koo, J.S. UP-regulation of S100A8 and S100A9 protein in bronchial epithelial cells by lipopolysaccharide. Exp. Lung Res. 2006, 32, 331–347. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.E.; Stockfelt, M.; Tengvall, S.; Bergman, P.; Linden, A.; Qvarfordt, I. Endotoxin exposure increases LL-37—But not calprotectin—In healthy human airways. J. Innate Immun. 2017, 9, 475–482. [Google Scholar] [CrossRef] [PubMed]
- De Jong, H.K.; Achouiti, A.; Koh, G.C.; Parry, C.M.; Baker, S.; Faiz, M.A.; van Dissel, J.T.; Vollaard, A.M.; van Leeuwen, E.M.; Roelofs, J.J.; et al. Expression and function of S100A8/A9 (calprotectin) in human typhoid fever and the murine Salmonella model. PLoS Negl Trop Dis. 2015, 9, e0003663. [Google Scholar] [CrossRef] [Green Version]
- Gutjahr, E.; Madea, B. Inflammatory reaction patterns of the lung as a response to alveolar hypoxia and their significance for the diagnosis of asphyxiation. Forensic Sci. Int. 2019, 297, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Van Zoelen, M.A.D.; Vogl, T.; Foell, D.; Van Veen, S.Q.; Van Till, J.W.O.; Florquin, S.; Tanck, M.W.; Wittebole, X.; Laterre, P.-F.; Boermeester, M.A.; et al. Expression and role of myeloid-related protein-14 in clinical and experimental sepsis. Am. J. Respir. Crit. Care Med. 2009, 180, 1098–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipcsey, M.; Hanslin, K.; Stålberg, J.; Smekal, D.; Larsson, A. The time course of calprotectin liberation from human neutrophil granulocytes after Escherichia coli and endotoxin challenge. Innate Immun. 2019, 25, 369–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwadlo, G.; Brüggen, J.; Gerhards, G.; Schlegel, R.; Sorg, C. Two calcium-binding proteins associated with specific stages of myeloid cell differentiation are expressed by subsets of macrophages in inflammatory tissues. Clin. Exp. Immunol. 1988, 72, 510–515. [Google Scholar] [PubMed]
- Hirvonen, J.; Kortelainen, M.L.; Huttunen, P. Pulmonary and serum surfactant phospholipids and serum catecholamines in strangulation. An experimental study on rats. Forensic Sci. Int. 1997, 90, 17–24. [Google Scholar] [CrossRef]
- Ikeno, K.; Koike, K.; Fukuromoto, T.; Shimizu, T.; Nagatomo, M.; Korniyama, A. Increased macrophage-colony stimulating factor levels in neonates with perinatal complications. Early Hum. Dev. 1996, 46, 229–237. [Google Scholar] [CrossRef]
- Croce, K.; Gao, H.; Wang, Y.; Mooroka, T.; Sakuma, M.; Shi, C.; Sukhova, G.K.; Packard, R.R.; Hogg, N.; Libby, P.; et al. Myeloid-related protein-8/14 is critical for the biological response to vascular injury. Circulation 2009, 120, 427–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niemelä, M.; Niemelä, O.; Bloigu, R.; Bloigu, A.; Kangastupa, P.; Juvonen, T. Serum calprotectin, a marker of neutrophil activation, and other mediators of inflammation in response to various types of extreme physical exertion in healthy volunteers. J. Inflamm. Res. 2020, 13, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Voganatsi, A.; Panyutich, A.; Miyasaki, K.T.; Murthy, R.K. Mechanism of extracellular release of human neutrophil calprotectin complex. J. Leukoc. Biol. 2001, 70, 130–134. [Google Scholar] [PubMed]
- Tardif, M.R.; Chapeton-Montes, J.A.; Posvandzic, A.; Pagé, N.; Gilbert, C.; Tessier, P.A. Secretion of S100A8, S100A9, and S100A12 by neutrophils involves reactive oxygen species and potassium efflux. J. Immunol. Res. 2015, 2015, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stroncek, D.F.; Shankar, R.A.; Skubitz, K.M. The subcellular distribution of myeloid-related protein 8 (MRP8) and MRP14 in human neutrophils. J. Transl. Med. 2005, 3, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hetland, G.; Talgo, G.J.; Fagerhol, M.K. Chemotaxins C5a and fMLP induce release of calprotectin (leucocyte L1 protein) from polymorphonuclear cells in vitro. Mol. Pathol. 1998, 51, 143–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lominadze, G.; Rane, M.J.; Merchant, M.; Cai, J.; Ward, R.A.; McLeish, K.R. Myeloid-related protein-14 is a p38 MAPK sub-strate in human neutrophils. J. Immunol. 2005, 17, 7257–7267. [Google Scholar] [CrossRef] [Green Version]
- Guignard, F.; Mauel, J.; Markert, M. Phosphorylation of myeloid-related proteins MRP-14 and MRP-8 during human neutrophil activation. Eur. J. Biochem. 1996, 241, 265–271. [Google Scholar] [CrossRef]
- Seelenmeyer, C.; Stegmayer, C.; Nickel, W. Unconventional secretion of fibroblast growth factor 2 and galectin-1 does not require shedding of plasma membrane-derived vesicles. FEBS Lett. 2008, 58, 1362–1368. [Google Scholar] [CrossRef] [Green Version]
- Urban, C.F.; Ermert, D.; Schmid, M.; Abu-Abed, U.; Goosmann, C.; Nacken, W.; Brinkmann, V.; Jungblut, P.R.; Zychlinsky, A. Neutrophil extracellular traps contain calprotectin, a cytosolic protein complex involved in host defense against Candida al-bicans. PLoS Pathog. 2009, 5, e1000639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lügering, N.; Kucharzik, T.; Lügering, A.; Winde, G.; Sorg, C.; Domschke, W.; Stoll, R. Importance of combined treatment with IL-10 and IL-4, but not IL-13, for inhibition of monocyte release of the Ca(2+)-binding protein MRP8/14. Immunology 1997, 91, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Suryono Kido, J.-I.; Hayashi, N.; Kataoka, M.; Shinohara, Y.; Nagata, T. Norepinephrine stimulates calprotectin expression in human monocytic cells. J. Periodontal Res. 2006, 41, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Rammes, A.; Roth, J.; Goebeler, M.; Klempt, M.; Hartmann, M.; Sorg, C. Myeloid-related protein (MRP) 8 and MRP14, calcium-binding proteins of the S100 family, are secreted by activated monocytes via a novel, tubulin-dependent pathway. J. Biol. Chem. 1997, 272, 9496–9502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, S.-Y.; Segovia, J.A.; Chang, T.-H.; Morris, I.R.; Berton, M.T.; Tessier, P.A.; Tardif, M.R.; Cesaro, A.; Bose, S. DAMP molecule S100A9 acts as a molecular pattern to enhance inflammation during Influenza A virus infection: Role of DDX21-TRIF-TLR4-MyD88 pathway. PLOS Pathog. 2014, 10, e1003848. [Google Scholar] [CrossRef]
- Frosch, M.; Strey, A.; Vogl, T.; Wulffraat, N.M.; Kuis, W.; Sunderkötter, C.; Harms, E.; Sorg, C.; Roth, J. Myeloid-related proteins 8 and 14 are specifically secreted during interaction of phagocytes and activated endothelium and are useful markers for monitoring disease activity in pauciarticular-onset juvenile rheumatoid arthritis. Arthritis Rheum. 2000, 43, 628–637. [Google Scholar] [CrossRef]
- Mahnke, K.; Bhardwaj, R.; Sorg, C. Heterodimers of the calcium-binding proteins MRP8 and MRP14 are expressed on the surface of human monocytes upon adherence to fibronectin and collagen. Relation to TNF-α, IL-6, and superoxide production. J. Leukoc. Biol. 1995, 53, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Nilsen, T.; Sundström, J.; Lind, L.; Larsson, A. Serum calprotectin levels in elderly males and females without bacterial or viral infections. Clin. Biochem. 2014, 47, 1065–1068. [Google Scholar] [CrossRef]
- Garg, M.; Leach, S.T.; Coffey, M.J.; Katz, T.; Strachan, R.; Pang, T.; Needham, B.; Lui, K.; Ali, F.; Day, A.S.; et al. Age-dependent variation of fecal calprotectin in cystic fibrosis and healthy children. J. Cyst. Fibros. 2017, 16, 631–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, S.H.; Kim, D.W.; Lee, S.H.; Kolliputi, N.; Hong, S.J.; Suh, L.; Norton, J.; Hulse, K.E.; Seshadri, S.; Conley, D.B.; et al. Age-Related increased prevalence of asthma and nasal polyps in chronic rhinosinusitis and its association with altered IL-6 trans-signaling. Am. J. Respir. Cell Mol. Biol. 2015, 53, 601–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurnakova, J.; Zavada, J.; Hanova, P.; Hulejova, H.; Klein, M.; Mann, H.; Sleglova, O.; Olejarova, M.; Forejtova, S.; Ruzickova, O.; et al. Serum calprotectin (S100A8/9): An independent predictor of ultrasound synovitis in patients with rheumatoid arthritis. Arthritis Res. Ther. 2015, 17, 252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehrchen, J.M.; Sunderkötter, C.; Foell, D.; Vogl, T.; Roth, J. The endogenous Toll-like receptor 4 agonist S100A8/S100A9 (cal-protectin) as innate amplifier of infection, autoimmunity, and cancer. J. Leukoc. Biol. 2009, 86, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Calcaterra, V.; De Amici, M.; Leonard, M.M.; De Silvestri, A.; Pelizzo, G.; Buttari, N.; Michev, A.; Leggio, M.; Larizza, D.; Cena, H. Serum calprotectin level in children: Marker of obesity and its metabolic complications. Ann. Nutr. Metab. 2018, 73, 177–183. [Google Scholar] [CrossRef]
- Zhu, Q.; Li, F.; Wang, J.; Shen, L.; Sheng, X. Fecal calprotectin in healthy children aged 1-4 years. PLoS ONE 2016, 11, e0150725. [Google Scholar] [CrossRef] [Green Version]
- Joshi, S.; Lewis, S.J.; Creanor, S.; Ayling, R.M. Age-related faecal calprotectin, lactoferrin and tumour M2-PK concentrations in healthy volunteers. Ann. Clin. Biochem. 2010, 47, 259–263. [Google Scholar] [CrossRef]
- Nakashige, T.G.; Zygiel, E.M.; Drennan, C.L.; Nolan, E.M. Nickel sequestration by the host-defense protein human calprotectin. J. Am. Chem. Soc. 2017, 139, 8828–8836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Achouiti, A.; Vogl, T.; Endeman, H.; Mortensen, B.L.; Laterre, P.F.; Wittebole, X.; van Zoelen, M.A.; Zhang, Y.; Hoogerwerf, J.J.; Florquin, S.; et al. My-eloid-related protein-8/14 facilitates bacterial growth during pneumococcal pneumonia. Thorax 2014, 69, 1034–1042. [Google Scholar]
- Longbottom, D.; Sallenave, J.-M.; Van Heyningen, V. Subunit structure of calgranulins A and B obtained from sputum, plasma, granulocytes and cultured epithelial cells. Biochim. Biophys. Acta 1992, 1120, 215–222. [Google Scholar] [CrossRef]
- Huang, S.; Feng, C.; Chen, L.; Huang, Z.; Zhou, X.; Li, B.; Wang, L.-L.; Chen, W.; Lv, F.-Q.; Li, T.-S. Molecular mechanisms of mild and severe pneumonia: Insights from RNA sequencing. Med. Sci. Monit. 2017, 23, 1662–1673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raquil, M.-A.; Anceriz, N.; Rouleau, P.; Tessier, P.A. Blockade of antimicrobial proteins S100A8 and S100A9 inhibits phagocyte migration to the alveoli in streptococcal pneumonia. J. Immunol. 2008, 180, 3366–3374. [Google Scholar] [CrossRef] [Green Version]
- Corbin, B.D.; Seeley, E.H.; Raab, A.; Feldmann, J.; Miller, M.R.; Torres, V.J.; Anderson, K.L.; Dattilo, B.M.; Dunman, P.M.; Gerads, R.; et al. Metal chelation and inhibition of bacterial growth in tissue abscesses. Science 2008, 319, 962–965. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Ochoa, V.E.; Lam, D.; Lee, C.S.; Klaus, S.; Behnsen, J.; Liu, J.Z.; Chim, N.; Nuccio, S.P.; Rathi, S.G.; Mastroianni, J.R.; et al. Salmonella mitigates oxidative stress and thrives in the inflamed gut by evading calprotectin-mediated manganese sequestration. Cell Host Microbe 2016, 19, 814–825. [Google Scholar] [CrossRef]
- Hadley, R.C.; Gagnon, D.M.; Brophy, M.B.; Gu, Y.; Nakashige, T.G.; Britt, R.D.; Nolan, E.M. Biochemical and spectroscopic observation of Mn(II) sequestration from bacterial Mn(II) transport machinery by calprotectin. J. Am. Chem. Soc. 2017, 140, 110–113. [Google Scholar] [CrossRef]
- Hood, M.I.; Mortensen, B.L.; Moore, J.L.; Zhang, Y.; Kehl-Fie, T.E.; Sugitani, N.; Chazin, W.J.; Caprioli, R.M.; Skaar, E.P. Identification of an Acinetobacter baumannii zinc acquisition system that facilitates resistance to calprotectin-mediated zinc sequestration. PLoS Pathog. 2012, 8, e1003068. [Google Scholar] [CrossRef] [Green Version]
- De Filippo, K.; Neill, D.R.; Mathies, M.; Bangert, M.; McNeill, E.; Kadioglu, A.; Hogg, N. A new protective role for S100A9 in regulation of neutrophil recruitment during invasive pneumococcal pneumonia. FASEB J. 2014, 28, 3600–3608. [Google Scholar] [CrossRef] [PubMed]
- Bühling, F.; Ittenson, A.; Kaiser, D.; Thölert, G.; Hoffmann, B.; Reinhold, D.; Ansorge, S.; Welte, T. MRP8/MRP14, CD11b and HLA-DR expression of alveolar macrophages in pneumonia. Immunol. Lett. 2000, 71, 185–190. [Google Scholar] [CrossRef]
- Moore, J.L.; Becker, K.W.; Nicklay, J.J.; Boyd, K.L.; Skaar, E.P.; Caprioli, R.M. Imaging mass spectrometry for assessing temporal proteomics: Analysis of calprotectin in Acinetobacter baumannii pulmonary infection. Proteomics 2014, 14, 820–828. [Google Scholar] [CrossRef] [Green Version]
- Glaros, T.G.; Blancett, C.D.; Bell, T.M.; Natesan, M.; Ulrich, R.G. Serum biomarkers of Burkholderia mallei infection elucidated by proteomic imaging of skin and lung abscesses. Clin. Proteom. 2015, 12, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikacenic, C.; Moore, R.; Dmyterko, V.; West, T.E.; Altemeier, W.A.; Liles, W.C.; Lood, C. Neutrophil extracellular traps (NETs) are increased in the alveolar spaces of patients with ventilator-associated pneumonia. Crit. Care 2018, 22, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuipers, M.T.; Vogl, T.; Aslami, H.; Jongsma, G.; van den Berg, E.; Vlaar, A.P.J.; Roelofs, J.J.T.H.; Juffermans, N.P.; Schultz, M.J.; van der Poll, T.; et al. High levels of S100A8/A9 proteins aggravate ventilator-induced lung injury via TLR4 signaling. PLoS ONE 2013, 8, e68694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiroshima, Y.; Hsu, K.; Tedla, N.; Chung, Y.M.; Chow, S.; Herbert, C.; Geczy, C.L. S100A8 Induces IL-10 and protects against acute lung injury. J. Immunol. 2014, 192, 2800–2811. [Google Scholar] [CrossRef] [PubMed]
- Jerkic, S.P.; Michel, F.; Donath, H.; Herrmann, E.; Schubert, R.; Rosewich, M.; Zielen, S. Calprotectin as a new sensitive marker of neutrophilic inflammation in patients with bronchiolitis obliterans. Mediat. Inflamm. 2020, 2020, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Karadjian, G.; Fercoq, F.; Pionnier, N.; Vallarino-Lhermitte, N.; Lefoulon, E.; Nieguitsila, A.; Specht, S.; Carlin, L.M.; Martin, C. Migratory phase of Litomosoides sigmodontis filarial infective larvae is associated with pathology and transient increase of S100A9 expressing neutrophils in the lung. PLoS Negl. Trop. Dis. 2017, 11, e0005596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contreras, I.; Shio, M.T.; Cesaro, A.; Tessier, P.A.; Olivier, M. Impact of neutrophil-secreted myeloid related proteins 8 and 14 (MRP 8/14) on leishmaniasis progression. PLoS Negl. Trop. Dis. 2013, 7, e2461. [Google Scholar] [CrossRef] [PubMed]
- Pechkovsky, D.V.; Zalutskaya, O.M.; Ivanov, G.I.; Misuno, N.I. Calprotectin (MRP8/14 protein complex) release during my-cobacterial infection in vitro and in vivo. FEMS Immunol. Med. Microbiol. 2000, 29, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J. Early diagnosis of pulmonary tuberculosis using serum biomarkers. Proteomics 2015, 15, 6–7. [Google Scholar] [CrossRef]
- Xu, D.; Li, Y.; Li, X.; Wei, L.L.; Pan, Z.; Jiang, T.T.; Chen, Z.L.; Wang, C.; Cao, W.M.; Zhang, X.; et al. Serum protein S100A9, SOD3, and MMP9 as new diagnostic biomarkers for pulmonary tuberculosis by iTRAQ-coupled two-dimensional LC-MS/MS. Proteomics 2015, 15, 58–67. [Google Scholar] [CrossRef]
- Gopal, R.; Monin, L.; Torres, D.; Slight, S.; Mehra, S.; McKenna, K.C.; Fallert Junecko, B.A.; Reinhart, T.A.; Kolls, J.; Báez-Saldaña, R.; et al. S100A8/A9 Proteins mediate neutrophilic inflammation and lung pathology during tuberculosis. Am. J. Respir. Crit. Care Med. 2013, 188, 1137–1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Pan, L.; Han, F.; Luo, B.; Jia, H.; Xing, A.; Li, Q.; Zhang, Z. Proteomic profiling for plasma biomarkers of tuberculosis progression. Mol. Med. Rep. 2018, 18, 1551–1559. [Google Scholar] [CrossRef] [Green Version]
- Delabie, J.; de Wolf-Peeters, C.; van den Oord, J.J.; Desmet, V.J. Differential expression of the calcium-binding proteins MRP8 and MRP14 in granulomatous conditions: An immunohistochemical study. Clin. Exp. Immunol. 1990, 81, 123–126. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Huang, C.; Wu, M.; Zhong, Q.; Yang, K.; Li, M.; Zhan, X.; Wen, J.; Zhou, L.; Huang, X. MRP8/14 induces autophagy to eliminate intracellular Mycobacterium bovis BCG. J. Infect. 2015, 70, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Larsson, G.; Shenoy, K.T.; Ramasubramanian, R.; Thayumanavan, L.; Balakumaran, L.K.; Bjune, G.A.; Moum, B.A. High faecal calprotectin levels in intestinal tuberculosis are associated with granulomas in intestinal biopsies. Infect. Dis. 2015, 47, 137–143. [Google Scholar] [CrossRef]
- Larsson, G.; Shenoy, K.T.; Ramasubramanian, R.; Thayumanavan, L.; Balakumaran, L.K.; Bjune, G.A.; Moum, B.A. Faecal calprotectin levels differentiate intestinal from pulmonary tuberculosis: An observational study from Southern India. United Eur. Gastroenterol. J. 2014, 2, 397–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Zhao, F.; Su, W.; Li, Q.; Li, J.; Ji, J.; Deng, Y.; Zhou, Y.; Wang, X.; Yang, H.-M.; et al. Zinc finger and interferon-stimulated genes play a vital role in TB-IRIS following HAART in AIDS. Pers. Med. 2018, 15, 251–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García de Guadiana, R.L.; Mulero, M.D.R.; Olivo, M.H.; Rojas, C.R.; Arenas, V.R.; Morales, M.G.; Abellán, A.B.; Conesa-Zamora, P.; García-García, J.; Hernández, A.C.; et al. Circulating levels of GDF-15 and calprotectin for prediction of in-hospital mortality in COVID-19 patients: A case series. J. Infect. 2020, S0163–4453(20)30543-0. [Google Scholar] [CrossRef]
- Silvin, A.; Chapuis, N.; Dunsmore, G.; Goubet, A.-G.; Dubuisson, A.; DeRosa, L.; Almire, C.; Hénon, C.; Kosmider, O.; Droin, N.; et al. Elevated calprotectin and abnormal myeloid cell subsets discriminate severe from mild COVID-19. Cell 2020, 182, 1401–1418. [Google Scholar] [CrossRef]
- Shi, H.; Zuo, Y.; Yalavarthi, S.; Gockman, K.; Zuo, M.; Madison, J.A.; Blair, C.; Woodward, W.; Lezak, S.P.; Lugogo, N.L.; et al. Neutrophil calprotectin identifies severe pulmonary disease in COVID-19. J. Leukoc. Biol. 2020, 109, 67–72. [Google Scholar] [CrossRef]
- Petito, E.; Falcinelli, E.; Paliani, U.; Cesari, E.; Vaudo, G.; Sebastiano, M.; Cerotto, V.; Guglielmini, G.; Gori, F.; Malvestiti, M.; et al. COVIR study investigators. Neutrophil more than platelet activation associates with thrombotic complications in COVID-19 patients. J. Infect. Dis 2020, jiaa756. [Google Scholar] [CrossRef]
- Effenberger, M.; Grabherr, F.; Mayr, L.; Schwaerzler, J.; Nairz, M.; Seifert, M.; Hilbe, R.; Seiwald, S.; Scholl-Buergi, S.; Fritsche, G.; et al. Faecal calprotectin indicates intestinal inflammation in COVID-19. Gut 2020, 69, 1543–1544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ojetti, V.; Saviano, A.; Covino, M.; Acampora, N.; Troiani, E.; Franceschi, F.; GEMELLI AGAINST COVID-19 Group. COVID-19 and intestinal inflammation: Role of fecal calprotectin. Dig. Liver Dis. 2020, 52, 1231–1233. [Google Scholar] [CrossRef] [PubMed]
- Mago, S.; Vaziri, H.; Tadros, M. The utility of fecal calprotectin in the era of COVID-19 pandemic. Gastroenterology 2020, S0016–5085(20)30678-8. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Otero, N.; Blanco-Prieto, S.; Páez de la Cadena, M.; Vázquez-Iglesias, L.; Fernández-Villar, A.; Botana-Rial, M.I.; Rodríguez-Berrocal, F.J. Calprotectin: A novel biomarker for the diagnosis of pleural effusion. Br. J. Cancer 2012, 107, 1876–1882. [Google Scholar] [CrossRef] [Green Version]
- Vázquez-Iglesias, L.; Casado-Rey, P.; Botana-Rial, M.; Andrade-Olivié, M.A.; Rodriguez-Berrocal, F.J.; Páez de La Cadena, M.; Fernández-Villar, A. Evaluation of an automated commercial ELISA method for calprotectin determination in pleural fluid. Clin. Chem. Lab. Med. 2016, 55, e172–e174. [Google Scholar] [CrossRef]
- Luo, J.; Wang, M.; Li, C.; Liang, B.; Liu, D.; Shi, C.; Jiang, F.; Wang, T.; Li, P.; Liang, Z. A Novel combination of calprotectin and CXCL12 for predicting malignancy in patients with exudative pleural effusion. Medicine 2015, 94, e2105. [Google Scholar] [CrossRef]
- Botana-Rial, M.; Vázquez-Iglesias, L.; Casado-Rey, P.; Cadena, M.P.; Andrade-Olivié, M.A.; Abal-Arca, J.; García-Nimo, L.; Ferreiro-Fernández, L.; Valdés-Cuadrado, L.; San-José, M.E.; et al. Validation of calprotectin as a novel biomarker for the diagnosis of pleural effusion: A multicentre trial. Sci. Rep. 2020, 10, 5679. [Google Scholar] [CrossRef]
- Davidson, B.; Stavnes, H.T.; Førsund, M.; Berner, A.; Staff, A.C. CD105 (Endoglin) expression in breast carcinoma effusions is a marker of poor survival. Breast 2010, 19, 493–498. [Google Scholar] [CrossRef]
- Ødegaard, E.; Davidson, B.; Engh, V.; Onsrud, M.; Staff, A.C. Assessment of endoglin and calprotectin as potential biomarkers in ovarian carcinoma and borderline tumors of the ovary. Am. J. Obstet. Gynecol. 2008, 199, 533–538. [Google Scholar] [CrossRef]
- Wu, K.-A.; Wu, C.-C.; Chen, C.-D.; Chu, C.-M.; Shih, L.-J.; Liu, Y.-C.; Wang, C.-L.; Lin, H.-H.; Yang, C.-Y. Proteome profiling reveals novel biomarkers to identify complicated parapneumonic effusions. Sci. Rep. 2017, 7, 4026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutz, P.; Pfarr, K.; Nischalke, H.D.; Kramer, B.; Goeser, F.; Glässner, A.; Wolter, F.; Kokordelis, P.; Nattermann, J.; Sauerbruch, T.; et al. The ratio of calprotectin to total protein as a diagnostic and prognostic marker for spontaneous bacterial peritonitis in patients with liver cirrhosis and ascites. Clin. Chem. Lab. Med. 2015, 53, 2031–2039. [Google Scholar] [CrossRef] [PubMed]
- Reid, P.A.; McAllister, D.; Boyd, A.C.; Innes, J.A.; Porteous, D.; Greening, A.P.; Gray, R.D. Measurement of serum calprotectin in stable patients predicts exacerbation and lung function decline in cystic fibrosis. Am. J. Respir. Crit. Care Med. 2015, 191, 233–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnapp, Z.; Hartman, C.; Livnat, G.; Shteinberg, M.; Elenberg, Y. Decreased fecal calprotectin levels in cystic fibrosis patients after antibiotic treatment for respiratory exacerbation. J. Pediatr. Gastroenterol. Nutr. 2019, 68, 282–284. [Google Scholar] [CrossRef] [PubMed]
- Muhlebach, M.S.; Clancy, J.P.; Heltshe, S.L.; Ziady, A.; Kelley, T.; Accurso, F.; Pilewski, J.; Mayer-Hamblett, N.; Joseloff, E.; Sagel, S.D. Biomarkers for cystic fibrosis drug development. J. Cyst. Fibros. 2016, 15, 714–723. [Google Scholar] [CrossRef] [Green Version]
- Parisi, G.; Papale, M.; Rotolo, N.; Aloisio, D.; Tardino, L.; Scuderi, M.G.; Di Benedetto, V.; Nenna, R.; Midulla, F.; Leonardi, S. Severe disease in cystic fibrosis and fecal calprotectin levels. Immunobiology 2017, 222, 582–586. [Google Scholar] [CrossRef] [PubMed]
- Coffey, M.J.; Garg, M.; Homaira, N.; Jaffe, A.; Ooi, C.Y. Probiotics for people with cystic fibrosis. Cochrane Database Syst. Rev. 2020, 1, CD012949. [Google Scholar] [CrossRef] [PubMed]
- Sagel, S.D.; Thompson, V.; Chmiel, J.F.; Montgomery, G.S.; Nasr, S.Z.; Perkett, E.; Saavedra, M.T.; Slovis, B.; Anthony, M.M.; Emmett, P.; et al. Effect of treatment of cystic fibrosis pulmonary exacerbations on systemic inflammation. Ann. Am. Thorac. Soc. 2015, 12, 708–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, R.D.; MacGregor, G.; Noble, D.; Imrie, M.; Dewar, M.; Boyd, A.C.; Innes, J.A.; Porteous, D.J.; Greening, A.P. Sputum proteomics in inflammatory and suppurative respiratory diseases. Am. J. Respir. Crit. Care Med. 2008, 178, 444–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, R.D.; Imrie, M.; Boyd, A.C.; Porteous, D.; Innes, J.A.; Greening, A.P. Sputum and serum calprotectin are useful biomarkers during CF exacerbation. J. Cyst. Fibros. 2010, 9, 193–198. [Google Scholar] [CrossRef] [Green Version]
- Sagel, S.D.; Wagner, B.D.; Ziady, A.; Kelley, T.; Clancy, J.P.; Narvaez-Rivas, M.; Pilewski, J.; Joseloff, E.; Sha, W.; Zelnick, L.; et al. Utilizing centralized biorepository samples for biomarkers of cystic fibrosis lung disease severity. J. Cyst. Fibros. 2020, 19, 632–640. [Google Scholar] [CrossRef] [PubMed]
- Jung, D.; Dong, K.; Jang, J.; Lam, G.Y.; Wilcox, P.G.; Quon, B.S. Circulating CRP and calprotectin to diagnose CF pulmonary exacerbations. J. Cyst. Fibros. 2020, 46–49. [Google Scholar] [CrossRef] [PubMed]
- Dong, K.; Singh, A.; Ng, R.T.; Sin, N.D.; Tebbutt, S.J.; Ratjen, F.; Quon, B.S. Proteomic profiling to identify blood biomarkers predictive of response to azithromycin in children and adolescents with cystic fibrosis. Chest 2019, 156, 667–673. [Google Scholar] [CrossRef]
- Wakeman, C.A.; Moore, J.L.; Noto, M.J.; Zhang, Y.; Singleton, M.D.; Prentice, B.M.; Gilston, B.A.; Doster, R.S.; Gaddy, J.A.; Chazin, W.J.; et al. The innate immune protein calprotectin promotes Pseudomonas aeruginosa and Staphylococcus aureus interaction. Nat. Commun. 2016, 7, 11951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.H.; Hwang, S.M.; Chung, I.Y. S100A8, S100A9 and S100A12 activate airway epithelial cells to produce MUC5AC via extracellular signal-regulated kinase and nuclear factor-κB pathways. Immunology 2015, 144, 79–90. [Google Scholar] [CrossRef] [Green Version]
- Oczypok, E.A.; Milutinovic, P.S.; Alcorn, J.F.; Khare, A.; Crum, L.T.; Manni, M.L.; Epperly, M.W.; Pawluk, A.M.; Ray, A.; Oury, T.D. Pulmonary receptor for advanced glycation end-products promotes asthma pathogenesis through IL-33 and accumulation of group 2 innate lymphoid cells. J. Allergy Clin. Immunol. 2015, 136, 747–756.e4. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.-D.; Wang, Y.; Yin, L.-M.; Park, G.-H.; Ulloa, L.; Yang, Y.-Q. S100A8 protein attenuates airway hyperresponsiveness by suppressing the contraction of airway smooth muscle. Biochem. Biophys. Res. Commun. 2017, 484, 184–188. [Google Scholar] [CrossRef]
- Hammad, H.; Chieppa, M.; Perros, F.; Willart, M.A.; Germain, R.N.; Lambrecht, B.N. House dust mite allergen induces asthma via Toll-like receptor 4 triggering of airway structural cells. Nat. Med. 2009, 15, 410–416. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Endoh, I.; Hsu, K.; Tedla, N.; Endoh, Y.; Geczy, C.L. S100A8 modulates mast cell function and suppresses eosinophil migration in acute asthma. Antioxid. Redox. Signal. 2011, 14, 1589–1600. [Google Scholar] [CrossRef]
- Yin, L.-M.; Li, H.-Y.; Zhang, Q.-H.; Xu, Y.-D.; Wang, Y.; Jiang, Y.-L.; Wei, Y.; Liu, Y.-Y.; Yang, Y.-Q. Effects of S100A9 in a rat model of asthma and in isolated tracheal spirals. Biochem. Biophys. Res. Commun. 2010, 398, 547–552. [Google Scholar] [CrossRef]
- Cobanoglu, N.; Galip, N.; Dalkan, C.; Bahceciler, N.N. Leptin, ghrelin and calprotectin: Inflammatory markers in childhood asthma? Multidiscip. Respir. Med. 2013, 8, 62. [Google Scholar] [CrossRef] [Green Version]
- Aoki, T.; Matsumoto, Y.; Hirata, K.; Ochiai, K.; Okada, M.; Ichikawa, K.; Shibasaki, M.; Arinami, T.; Sumazaki, R.; Noguchi, E. Expression profiling of genes related to asthma exacerbations. Clin. Exp. Allergy 2009, 39, 213–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, T.-H.; Jang, A.-S.; Park, J.-S.; Kim, T.-H.; Choi, Y.S.; Shin, H.-R.; Park, S.-W.; Uh, S.-T.; Choi, J.-S.; Kim, Y.H.; et al. Elevation of S100 calcium binding protein A9 in sputum of neutrophilic inflammation in severe uncontrolled asthma. Ann. Allergy Asthma Immunol. 2013, 111, 268–275.e1. [Google Scholar] [CrossRef]
- Lee, T.-H.; Chang, H.S.; Bae, D.-J.; Song, H.J.; Kim, M.-S.; Park, J.S.; Jun, J.A.; Lee, S.Y.; Uh, S.T.; Kim, S.H.; et al. Role of S100A9 in the development of neutrophilic inflammation in asthmatics and in a murine model. Clin. Immunol. 2017, 183, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.G.; Hong, J.; Lee, P.H.; Lee, J.; Park, S.W.; Kim, D.; Jang, A.S. Serum Calprotectin Is a Potential Marker in Patients with Asthma. J. Korean Med. Sci. 2020, 35, e362. (In Chinese) [Google Scholar] [CrossRef] [PubMed]
- Wilkerson, E.M.; Johansson, M.W.; Hebert, A.S.; Westphall, M.S.; Mathur, S.K.; Jarjour, N.N.; Schwantes, E.A.; Mosher, D.F.; Coon, J.J. The peripheral blood eosinophil proteome. J. Proteome Res. 2016, 15, 1524–1533. [Google Scholar] [CrossRef] [Green Version]
- Greenlee, K.J.; Corry, D.B.; Engler, D.A.; Matsunami, R.K.; Tessier, P.; Cook, R.G.; Werb, Z.; Kheradmand, F. Proteomic iden-tification of in vivo substrates for matrix metalloproteinases 2 and 9 reveals a mechanism for resolution of inflammation. J. Immunol. 2006, 177, 7312–7321. [Google Scholar] [CrossRef] [Green Version]
- Orivuori, L.; Mustonen, K.; De Goffau, M.C.; Hakala, S.; Paasela, M.; Roduit, C.; Dalphin, J.; Genuneit, J.; Lauener, R.; Riedler, J.; et al. High level of fecal calprotectin at age 2 months as a marker of intestinal inflammation predicts atopic dermatitis and asthma by age 6. Clin. Exp. Allergy 2015, 45, 928–939. [Google Scholar] [CrossRef]
- Foong, R.E.; Bosco, A.; Jones, A.C.; Gout, A.; Gorman, S.; Hart, P.H.; Zosky, G.R. The Effects ofIn UteroVitamin D Deficiency on airway smooth muscle mass and lung function. Am. J. Respir. Cell Mol. Biol. 2015, 53, 664–675. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, E.; Muhlebach, M.S.; Tessier, P.A.; Alexis, N.E.; Duncan Hite, R.; Seeds, M.C.; Peden, D.B.; Meredith, W. Different expression ratio of S100A8/A9 and S100A12 in acute and chronic lung diseases. Respir. Med. 2008, 102, 567–573. [Google Scholar] [CrossRef] [Green Version]
- Railwah, C.; Lora, A.; Zahid, K.; Goldenberg, H.; Campos, M.A.; Wyman, A.; Jundi, B.; Ploszaj, M.; Rivas, M.; Dabo, A.J.; et al. Cigarette smoke induction of S100A9 contributes to chronic obstructive pulmonary disease. Am. J. Physiol. Lung Cell. Mol. Physiol. 2020, 319, L1021–L1035. [Google Scholar] [CrossRef]
- Lin, C.R.; Bahmed, K.; Criner, G.J.; Marchetti, N.; Tuder, R.M.; Kelsen, S.; Bolla, S.; Mandapati, C.; Kosmider, B. S100A8 Protects Human Primary Alveolar Type II Cells against Injury and Emphysema. Am. J. Respir. Cell Mol. Biol. 2019, 60, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-R.; Bahmed, K.; Tomar, D.; Marchetti, N.; Criner, G.J.; Bolla, S.; Wilson, M.A.; Madesh, M.; Kosmider, B. The relationship between DJ-1 and S100A8 in human primary alveolar type II cells in emphysema. Am. J. Physiol. Lung Cell. Mol. Physiol. 2019, 317, L791–L804. [Google Scholar] [CrossRef]
- Gomes, L.H.; Raftery, M.J.; Yan, W.X.; Goyette, J.D.; Thomas, P.S.; Geczy, C.L. S100A8 and S100A9—Oxidant scavengers in inflammation. Free Radic Biol. Med. 2013, 58, 170–186. [Google Scholar] [CrossRef]
- Foronjy, R.F.; Ochieng, P.O.; Salathe, M.A.; Dabo, A.J.; Eden, E.; Baumlin, N.; Cummins, N.; Barik, S.; Campos, M.; Thorp, E.B.; et al. Protein tyrosine phosphatase 1B negatively regulates S100A9-mediated lung damage during respiratory syncytial virus exacerbations. Mucosal Immunol. 2016, 9, 1317–1329. [Google Scholar] [CrossRef]
- Chen, M.; Wang, T.; Shen, Y.; Xu, D.; Li, X.; An, J.; Dong, J.; Li, D.; Wen, F.-Q.; Chen, L. Knockout of RAGE ameliorates mainstream cigarette smoke-induced airway inflammation in mice. Int. Immunopharmacol. 2017, 50, 230–235. [Google Scholar] [CrossRef]
- Li, M.; Guo, L.; Wang, H.; Wang, T.; Shen, Y.; Liao, Z.; Wen, F.; Chen, L. RAGE-ligands axis: A new ‘driving force’ for cigarette smoke-induced airway inflammation in COPD? Respirology 2015, 20, 998–999. [Google Scholar] [CrossRef]
- Sallenave, J.M.; Ryle, A.P. Purification and characterization of elastase-specific inhibitor. Sequence homology with mucus proteinase inhibitor. Biol. Chem. Hoppe-Seyler 1991, 372, 13–21. [Google Scholar] [CrossRef]
- Holmgaard, D.B.; Mygind, L.H.; Titlestad, I.L.; Madsen, H.; Pedersen, S.S.; Mortensen, O.H.; Pedersen, C. Calprotectin—A marker of mortality in COPD? Results from a prospective cohort study. COPD 2013, 10, 581–587. [Google Scholar] [CrossRef]
- Pouwels, S.D.; Nawijn, M.C.; Bathoorn, E.; Riezebos-Brilman, A.; van Oosterhout, A.J.; Kerstjens, H.A.; Heijink, I.H. Increased serum levels of LL37, HMGB1 and S100A9 during exacerbation in COPD patients. Eur. Respir. J. 2015, 45, 1482–1485. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, A.K.; Holmgaard, D.B.; Mygind, L.H.; Johansen, J.S.; Pedersen, C. Neutrophil-to-lymphocyte ratio, calprotectin and YKL-40 in patients with chronic obstructive pulmonary disease: Correlations and 5-year mortality—a cohort study. J. Inflamm. 2015, 12, 20. [Google Scholar] [CrossRef] [Green Version]
- Gebhardt, C.; Breitenbach, U.; Tuckermann, J.P.; Dittrich, B.T.; Richter, K.H.; Angel, P. Calgranulins S100A8 and S100A9 are negatively regulated by glucocorticoids in a c-Fos-dependent manner and overexpressed throughout skin carcinogenesis. Oncogene 2002, 21, 4266–4276. [Google Scholar] [CrossRef] [Green Version]
- Bozinovski, S.; Cross, M.; Vlahos, R.; Jones, J.E.; Hsuu, K.; Tessier, P.A.; Reynolds, E.C.; Hume, D.A.; Hamilton, J.A.; Geczy, C.L.; et al. S100A8 chemotactic protein is abundantly increased, but only a minor contributor to LPS-induced, steroid resistant neutrophilic lung inflammation in vivo. J. Proteome Res. 2004, 4, 136–145. [Google Scholar] [CrossRef]
- Balkwill, F.; Mantovani, A. Inflammation and cancer: Back to Virchow? Lancet 2001, 357, 539–545. [Google Scholar] [CrossRef]
- Munn, L.L. Cancer and inflammation. Wiley Interdiscip. Rev. Syst. Biol. Med. 2017, 9, e1370. [Google Scholar] [CrossRef] [Green Version]
- Silva, E.J.; Argyris, P.P.; Zou, X.; Ross, K.F.; Herzberg, M.C. S100A8/A9 regulates MMP-2 expression and invasion and migration by carcinoma cells. Int. J. Biochem. Cell Biol. 2014, 55, 279–287. [Google Scholar] [CrossRef]
- Ghavami, S.; Tehrani, F.K.; Hashemi, M.; Zarif, M.N. Possible involvement of a specific cell surface receptor for calprotectin-induced apoptosis in colon adenocarcinoma and carcinam cell lines (SW742 and HT29/219). J. Sci. Islamic Repub. Iran. 2004, 15, 3–12. [Google Scholar]
- Zheng, Y.; Hou, J.; Peng, L.; Zhang, X.; Jia, L.; Wang, X.; Wei, S.; Meng, H. The pro-apoptotic and pro-inflammatory effects of calprotectin on human periodontal ligament cells. PLoS ONE 2014, 9, e110421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghavami, S.; Kerkhoff, C.; Chazin, W.J.; Kadkhoda, K.; Xiao, W.; Zuse, A.; Hashemi, M.; Eshraghi, M.; Schulze-Osthoff, K.; Klonisch, T.; et al. S100A8/9 induces cell death via a novel, RAGE-independent pathway that involves selective release of Smac/DIABLO and Omi/HtrA2. Biochim. Biophys. Acta 2008, 1783, 297–311. [Google Scholar] [CrossRef] [Green Version]
- Rajendran, P.; Chen, Y.-F.; Chen, Y.-F.; Chung, L.-C.; Tamilselvi, S.; Shen, C.-Y.; Day, C.H.; Chen, R.-J.; Padma, V.V.; Kuo, W.-W.; et al. The multifaceted link between inflammation and human diseases. J. Cell. Physiol. 2018, 233, 6458–6471. [Google Scholar] [CrossRef]
- Nishikawa, Y.; Kajiura, Y.; Lew, J.H.; Kido, J.-I.; Nagata, T.; Naruishi, K. Calprotectin induces IL-6 and MCP-1 production via toll-like receptor 4 signaling in human gingival fibroblasts. J. Cell. Physiol. 2017, 232, 1862–1871. [Google Scholar] [CrossRef]
- Heizmann, C.; Ackermann, G.; Galichet, A. Pathologies involving the S100 proteins and RAGE. Alzheimer’s Disease 2007, 45, 93–138. [Google Scholar] [CrossRef]
- Leclerc, E.F.G.; Weibel, M.; Heizmann, C.W.; Galichet, A. S100B and S100A6 differentially modulate cell survival by interacting with distinct RAGE (Receptor for Advanced Glycation End Products) immunoglobulin domains. J. Biol. Chem. 2007, 282, 31317–31331. [Google Scholar] [CrossRef] [Green Version]
- Ichikawa, M.; Williams, R.; Wang, L.; Vogl, T.; Srikrishna, G. S100A8/ A9 activate key genes and pathways in colon tumor progression. Mol. Cancer Res. 2011, 9, 133–148. [Google Scholar] [CrossRef] [Green Version]
- Zhong, J.M.; Li, J.; Kang, A.D.; Huang, S.Q.; Liu, W.B.; Zhang, Y.; Liul, Z.H.; Zeng, L. Protein S100-A8: A potential metastasis-associated protein for breast cancer determined via iTRAQ quantitative proteomic and clinicopathological analysis. Oncol. Lett. 2018, 15, 5285–5293. [Google Scholar] [CrossRef] [PubMed]
- Deguchi, A.; Tomita, T.; Ohto, U.; Takemura, K.; Kitao, A.; Akashitakamura, S.; Miyake, K.; Maru, Y. Eritoran inhibits S100A8-mediated TLR4/MD-2 activation and tumor growth by changing the immune microenvironment. Oncogene 2016, 35, 1445–1456. [Google Scholar] [CrossRef]
- Sumardika, I.W.; Chen, Y.; Tomonobu, N.; Kinoshita, R.; Ruma, I.M.W.; Sato, H.; Kondo, E.; Inoue, Y.; Yamauchi, A.; Murata, H.; et al. Neuroplastin-β mediates S100A8/A9-induced lung cancer disseminative progression. Mol. Carcinog. 2019, 58, 980–995. [Google Scholar] [CrossRef]
- Srikrishna, G. S100A8 and S100A9: New insights into their roles in malignancy. J. Innate Immun. 2012, 4, 31–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Sumardika, I.W.; Tomonobu, N.; Kinoshita, R.; Inoue, Y.; Iioka, H.; Mitsui, Y.; Saito, K.; Ruma, I.M.W.; Sato, H.; et al. Critical role of the MCAM-ETV4 axis triggered by extracellular S100A8/A9 in breast cancer aggressiveness. Neoplasia. 2019, 21, 627–640. [Google Scholar] [CrossRef]
- Núñez-Naveira, L.; Mariñas-Pardo, L.; Montero-Martínez, C. Mass spectrometry analysis of the exhaled breath condensate and proposal of dermcidin and S100A9 as possible markers for lung cancer prognosis. Lung 2019, 197, 523–531. [Google Scholar] [CrossRef]
- Kawai, H.; Minamiya, Y.; Takahashi, N. Prognostic impact of S100A9 overexpression in non-small cell lung cancer. Tumor Biol. 2011, 32, 641–646. [Google Scholar] [CrossRef]
- Liu, Y.; Cui, J.; Tang, Y.-L.; Huang, L.; Zhou, C.-Y.; Xu, J. Prognostic roles of mRNA expression of S100 in non-small-cell lung cancer. BioMed Res. Int. 2018, 2018, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Feng, P.-H.; Lee, K.-Y.; Chang, Y.-L.; Chan, Y.-F.; Kuo, L.-W.; Lin, T.-Y.; Chung, F.-T.; Kuo, C.-S.; Yu, C.-T.; Lin, S.-M.; et al. CD14(+)S100A9(+) monocytic myeloid-derived suppressor cells and their clinical relevance in non-small cell lung cancer. Am. J. Respir. Crit. Care Med. 2012, 186, 1025–1036. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Huang, Q.; Tang, T.; Gu, L.; Du, J.; Li, Z.; Lu, X.; Zhou, X. Clinical significance of calcium-binding protein S100A8 and S100A9 expression in non-small cell lung cancer. Thorac. Cancer 2018, 9, 800–804. [Google Scholar] [CrossRef]
- Blanco-Prieto, S.; De Chiara, L.; Rodríguez-Girondo, M.; Vázquez-Iglesias, L.; Rodríguez-Berrocal, F.J.; Fernández-Villar, A.; Botana-Rial, M.I.; De La Cadena, M.P. Highly sensitive marker panel for guidance in lung cancer rapid diagnostic units. Sci. Rep. 2017, 7, 41151. [Google Scholar] [CrossRef] [PubMed]
- Eisenblaetter, M.; Flores-Borja, F.; Lee, J.J.; Wefers, C.; Smith, H.; Hueting, R.; Cooper, M.S.; Blower, P.J.; Patel, D.; Rodríguez-Justo, M.; et al. Visualization of tumor-immune interaction—Target-specific imaging of S100A8/A9 reveals pre-metastatic niche establishment. Theranostics 2017, 7, 2392–2401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, P.-H.; Chen, K.-Y.; Huang, Y.-C.; Luo, C.-S.; Wu, S.M.; Chen, T.-T.; Lee, C.-N.; Yeh, C.-T.; Chuang, H.-C.; Han, C.-L.; et al. Bevacizumab reduces S100A9-positive MDSCs linked to intracranial control in patients with EGFR-mutant lung adenocarcinoma. J. Thorac. Oncol. 2018, 13, 958–967. [Google Scholar] [CrossRef]
- Tian, T.; Li, X.; Hua, Z.; Ma, J.; Liu, Z.; Chen, H.; Cui, Z. S100A1 promotes cell proliferation and migration and is associated with lymph node metastasis in ovarian cancer. Discov. Med. 2017, 23, 235–245. [Google Scholar] [PubMed]
- Kinoshita, R.; Sato, H.; Yamauchi, A.; Takahashi, Y.; Inoue, Y.; Sumardika, I.W.; Chen, Y.; Tomonobu, N.; Araki, K.; Shien, K.; et al. Newly developed an-ti-S100A8/A9 monoclonal antibody efficiently prevents lung tropic cancer metastasis. Int. J. Cancer 2019, 145, 569–575. [Google Scholar] [CrossRef]
- Grebhardt, S.; Müller-Decker, K.; Bestvater, F.; Hershfinkel, M.; Mayer, D. Impact of S100A8/A9 expression on prostate cancer progression in vitro and in vivo. J. Cell. Physiol. 2014, 229, 661–671. [Google Scholar] [CrossRef]
- Vrakas, C.N.; O’Sullivan, R.M.; Evans, S.E.; Ingram, D.A.; Jones, C.B.; Phuong, T.; Kurt, R.A. The Measure of DAMPs and a role for S100A8 in recruiting suppressor cells in breast cancer lung metastasis. Immunol. Investig. 2015, 44, 174–188. [Google Scholar] [CrossRef]
- Boyd, J.H.; Kan, B.; Roberts, H.; Wang, Y.; Walley, K.R. S100A8 and S100A9 mediate endotoxin-induced cardiomyocyte dys-function via the receptor for advanced glycation end products. Circ. Res. 2008, 102, 1239–1246. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Liu, S.; Wu, C.; Sun, Z. TGFβ modulates inflammatory cytokines and growth factors to create premetastatic microen-vironment and stimulate lung metastasis. J. Mol. Histol. 2015, 46, 365–375. [Google Scholar] [CrossRef]
- Tanriover, G.; Eyinc, M.B.; Aliyev, E.; Dilmac, S.; Erin, N. Presence of S100A8/Gr1-positive myeloid-derived suppressor cells in primary tumors and visceral organs invaded by breast carcinoma cells. Clin. Breast Cancer 2018, 18, e1067–e1076. [Google Scholar] [CrossRef]
- Yin, C.; Li, H.; Zhang, B.; Liu, Y.; Lu, G.; Lu, S.; Sun, L.; Qi, Y.; Li, X.; Chen, W. RAGE-binding S100A8/A9 promotes the mi-gration and invasion of human breast cancer cells through actin polymerization and epithelial-mesenchymal transition. Breast Cancer Res. Treat. 2013, 142, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Ghavami, S.; Rashedi, I.; Dattilo, B.M.; Eshraghi, M.; Chazin, W.J.; Hashemi, M.; Wesselborg, S.; Kerkhoff, C.; Los, M.J. S100A8/A9 at low concentration promotes tumor cell growth via RAGE ligation and MAP kinase-dependent pathway. J. Leukoc. Biol. 2008, 83, 1484–1492. [Google Scholar] [CrossRef]
- Hiratsuka, S.; Watanabe, A.; Sakurai, Y.; Akashi-Takamura, S.; Ishibashi, S.; Miyake, K.; Shibuya, M.; Akira, S.; Aburatani, H.; Maru, Y. The S100A8–serum amyloid A3–TLR4 paracrine cascade establishes a pre-metastatic phase. Nat. Cell Biol. 2008, 10, 1349–1355. [Google Scholar] [CrossRef]
- Marelli, G.; Sica, A.; Vannucci, L.; Allavena, P. Inflammation as target in cancer therapy. Curr. Opin. Pharmacol. 2017, 35, 57–65. [Google Scholar] [CrossRef]
- Nukui, T.; Ehama, R.; Sakaguchi, M.; Sonegawa, H.; Katagiri, C.; Hibino, T.; Huh, N.-H. S100A8/A9, a key mediator for positive feedback growth stimulation of normal human keratinocytes. J. Cell. Biochem. 2008, 104, 453–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, S.Y.; Yuzhalin, A.E.; Gordon-Weeks, A.N.; Muschel, R.J. Tumor-infiltrating monocytes/macrophages promote tumor in-vasion and migration by upregulating S100A8 and S100A9 expression in cancer cells. Oncogene 2016, 35, 5735–5745. [Google Scholar] [CrossRef] [Green Version]
- Xin, X.; Zeng, X.; Gu, H.; Li, M.; Tan, H.; Jin, Z.; Hua, T.; Shi, R.; Wang, H. CD147/EMMPRIN overexpression and prognosis in cancer: A systematic review and meta-analysis. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Hibino, T.; Sakaguchi, M.; Miyamoto, S.; Yamamoto, M.; Motoyama, A.; Hosoi, J.; Shimokata, T.; Ito, T.; Tsuboi, R.; Huh, N.-H. S100A9 is a novel ligand of EMMPRIN that promotes melanoma metastasis. Cancer Res. 2013, 73, 172–183. [Google Scholar] [CrossRef] [Green Version]
- Yurchenko, V.; Constant, S.; Eisenmesser, E.; Bukrinsky, M. Cyclophilin-CD147 interactions: A new target for anti-inflammatory therapeutics. Clin. Exp. Immunol. 2010, 160, 305–317. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Chen, H.; Zhu, X.; Ma, Y.; Liu, Q.; Xue, Y.; Chu, H.; Wu, W.; Wang, J.; Zou, H. S100A9 promotes human lung fibroblast cells activation through receptor for advanced glycation end-product-mediated extracellular-regulated kinase 1/2, mitogen-activated protein-kinase and nuclear factor-κB-dependent pathways. Clin. Exp. Immunol. 2013, 173, 523–535. [Google Scholar] [CrossRef]
- Bennett, D.; Salvini, M.; Fui, A.; Cillis, G.; Cameli, P.; Mazzei, M.A.; Fossi, A.; Refini, R.M.; Rottoli, P. Calgranulin B and KL-6 in bronchoalveolar lavage of patients with IPF and NSIP. Inflammation 2019, 42, 463–470. [Google Scholar] [CrossRef]
- Korthagen, N.M.; Nagtegaal, M.M.; van Moorsel, C.H.M.; Kazemier, K.M.; van den Bosch, J.M.; Grutters, J.C. MRP14 is elevated in the bronchoalveolar lavage fluid of fibrosing interstitial lung diseases. Clin. Exp. Immunol. 2010, 161, 342–347. [Google Scholar] [CrossRef]
- Machahua, C.; Guler, S.A.; Horn, M.P.; Planas-Cerezales, L.; Montes-Worboys, A.; Geiser, T.K.; Molina-Molina, M.; Funke-Chambour, M. Serum calprotectin as new biomarker for disease severity in idiopathic pulmonary fibrosis: A cross-sectional study in two independent cohorts. BMJ Open Respir. Res. 2021, 8, e000827. [Google Scholar] [CrossRef]
- Maver, A.; Medica, I.; Peterlin, B. Search for sarcoidosis candidate genes by integration of data from genomic, transcriptomic and proteomic studies. Med. Sci. Monit. 2009, 15, SR22–SR28. [Google Scholar] [PubMed]
- Terasaki, F.; Fujita, M.; Shimomura, H.; Tsukada, B.; Otsuka, K.; Otsuka, K.; Katashima, T.; Ikemoto, M.; Kitaura, Y. Enhanced expression of myeloid-related protein complex (MRP8/14) in macrophages and multinucleated giant cells in granulomas of pa-tients with active cardiac sarcoidosis. Circ. J. 2007, 71, 1545–1550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Wu, W.-Y.; Tu, W.-Z.; Chu, H.-Y.; Zhu, X.-X.; Liang, M.-R.; Xue, Y.; Wang, J.-C.; Zou, H.-J. Increased expression of S100A8 and S100A9 in patients with diffuse cutaneous systemic sclerosis. A correlation with organ involvement and immunological abnormalities. Clin. Rheumatol. 2013, 32, 1501–1510. [Google Scholar] [CrossRef] [PubMed]
- Van Bon, L.; Cossu, M.; Loof, A.; Gohar, F.; Wittkowski, H.; Vonk, M.; Roth, J.; van den Berg, W.; van Heerde, W.; Broen, J.C.A.; et al. Proteomic analysis of plasma identifies the Toll-like receptor agonists S100A8/A9 as a novel possible marker for systemic sclerosis phenotype. Ann. Rheum. Dis. 2014, 73, 1585–1589. [Google Scholar] [CrossRef]
- Fietta, A.; Bardoni, A.; Salvini, R.; Passadore, I.; Morosini, M.; Cavagna, L.; Codullo, V.; Pozzi, E.; Meloni, F.; Montecucco, C. Analysis of bronchoalveolar lavage fluid proteome from systemic sclerosis patients with or without functional, clinical and radiological signs of lung fibrosis. Arthritis Res. Ther. 2006, 8, R160. [Google Scholar] [CrossRef] [Green Version]
- Hesselstrand, R.; Wildt, M.; Bozovic, G.; Andersson-Sjöland, A.; Andréasson, K.; Scheja, A.; Westergren-Thorsson, G.; Bjermer, L.; Wuttge, D.M. Biomarkers from bronchoalveolar lavage fluid in systemic sclerosis patients with interstitial lung disease relate to severity of lung fibrosis. Respir. Med. 2013, 107, 1079–1086. [Google Scholar] [CrossRef] [Green Version]
- Caimmi, C.; Bertoldo, E.; Venturini, A.; Caramaschi, P.; Frulloni, L.; Ciccocioppo, R.; Brunelli, S.; Idolazzi, L.; Gatti, D.; Rossini, M.; et al. relationship between increased fecal calprotectin levels and interstitial lung disease in systemic sclerosis. J. Rheumatol. 2018, 46, 274–278. [Google Scholar] [CrossRef]
- He, M.; Kubo, H.; Ishizawa, K.; Hegab, A.E.; Yamamoto, Y.; Yamamoto, H.; Yamaya, M. The role of the receptor for advanced glycation end-products in lung fibrosis. Am. J. Physiol. Lung Cell. Mol. Physiol. 2007, 293, L1427–L1436. [Google Scholar] [CrossRef] [Green Version]
- Yoshizaki, A.; Komura, K.; Iwata, Y.; Ogawa, F.; Hara, T.; Muroi, E.; Takenaka, M.; Shimizu, K.; Hasegawa, M.; Fujimoto, M.; et al. Clinical significance of serum HMGB-1 and sRAGE levels in systemic sclerosis: Association with disease severity. J. Clin. Immunol. 2009, 29, 180–189. [Google Scholar] [CrossRef] [Green Version]
- Andréasson, K.; Alrawi, Z.; Persson, A.; Jönsson, G.; Marsal, J. Intestinal dysbiosis is common in systemic sclerosis and associated with gastrointestinal and extraintestinal features of disease. Arthritis Res. Ther. 2016, 18, 278. [Google Scholar] [CrossRef] [Green Version]
- Marie, I.; Leroi, A.-M.; Menard, J.-F.; Levesque, H.; Quillard, M.; Ducrotte, P. Fecal calprotectin in systemic sclerosis and review of the literature. Autoimmun. Rev. 2015, 14, 547–554. [Google Scholar] [CrossRef]
- Torun, M.T.; Ünver, E.; Yalçın, Y.; Kanmaz, L.; Seçkin, E. The importance of calprotectin levels in obstructive sleep apnea syndrome severity. Curr. Med. Res. Opin. 2018, 34, 401–405. [Google Scholar] [CrossRef]
- Kum, N.Y.; Kum, R.O.; Candar, T.; Baklaci, D.; Guler, I.; Kuzucu, I.; Ozcan, K.M.; Ozcan, M.; Dere, H. Elevated serum calprotectin as an inflammatory marker in obstructive sleep apnea. Cranio 2020, 1–7. [Google Scholar] [CrossRef]
- Bhattacharjee, R.; Kim, J.; Alotaibi, W.H.; Kheirandish-Gozal, L.; Capdevila, O.S.; Gozal, D. Endothelial dysfunction in children without hypertension: Potential contributions of obesity and obstructive sleep apnea. Chest 2012, 141, 682–691. [Google Scholar] [CrossRef] [Green Version]
- Cholidou, K.G.; Kostakis, I.D.; Manali, E.D.; Perrea, D.; Margeli, A.; Vougas, K.; Markozannes, E.; Koulouris, N.; Alchanatis, M. Calprotectin: A protein related to cardiovascular risk in adult patients with obstructive sleep apnea. Cytokine 2013, 61, 917–923. [Google Scholar] [CrossRef]
- Kim, J.; Lee, S.; Bhattacharjee, R.; Khalyfa, A.; Kheirandish-Gozal, L.; Gozal, D. Leukocyte telomere length and plasma catestatin and myeloid-related protein 8/14 concentrations in children with obstructive sleep apnea. Chest 2010, 138, 91–99. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Bhattacharjee, R.; Snow, A.B.; Capdevila, O.S.; Kheirandish-Gozal, L.; Gozal, D. Myeloid-related protein 8/14 levels in children with obstructive sleep apnoea. Eur. Respir. J. 2010, 35, 843–850. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Bhattacharjee, R.; Kheirandish-Gozal, L.; Spruyt, K.; Gozal, D. Circulating microparticles in children with sleep dis-ordered breathing. Chest 2011, 140, 408–417. [Google Scholar] [CrossRef] [PubMed]
- De Luca Canto, G.; Pachêco-Pereira, C.; Aydinoz, S.; Major, P.W.; Flores-Mir, C.; Gozal, D. Biomarkers associated with obstructive sleep apnea and morbidities: A scoping review. Sleep Med. 2015, 16, 347–357. [Google Scholar] [CrossRef]
- Oto, J.; Plana, E.; Solmoirago, M.J.; Fernández-Pardo, Á.; Hervás-Marín, D.; Cana, F.; España, F.; Artoni, A.; Bucciarelli, P.; Carrabba, G.; et al. microRNAs and markers of neutrophil activation as predictors of early incidental post-surgical pulmonary embolism in patients with intracranial tumors. Cancers 2020, 12, 1536. [Google Scholar] [CrossRef]
- Zuo, Y.; Zuo, M.; Yalavarthi, S.; Gockman, K.; Madison, J.A.; Shi, H.; Woodard, W.; Lezak, S.P.; Lugogo, N.L.; Knight, J.S.; et al. Neutrophil extracellular traps and thrombosis in COVID-19. J. Thromb. Thrombolysis 2020, 1–8. [Google Scholar] [CrossRef]
- Martos, L.; Oto, J.; Fernández-Pardo, Á.; Plana, E.; Solmoirago, M.J.; Cana, F.; Hervás-Marín, D.; Bonanad, S.; Ferrando, F.; España, F.; et al. Increase of neutrophil activation markers in venous thrombosis—Contribution of circulating activated protein C. Int. J. Mol. Sci. 2020, 21, 5651. [Google Scholar] [CrossRef]
- Oto, J.; Navarro, S.; Larsen, A.C.; Solmoirago, M.J.; Plana, E.; Hervás-Marín, D.; Fernández-Pardo, Á.; España, F.; Kristensen, S.R.; Thorlacius-Ussing, O.; et al. MicroRNAs and neutrophil activation markers predict venous thrombosis in pancreatic ductal adenocarcinoma and distal extrahepatic cholangiocarcinoma. Int. J. Mol. Sci. 2020, 21, 840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, K.; Sakaguchi, M.; Matsubara, H.; Akagi, S.; Sarashina, T.; Ejiri, K.; Akazawa, K.; Kondo, M.; Nakagawa, K.; Yoshida, M.; et al. Crucial role of RAGE in inappropriate increase of smooth muscle cells from patients with pulmonary arterial hypertension. PLoS ONE 2018, 13, e0203046. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kotsiou, O.S.; Papagiannis, D.; Papadopoulou, R.; Gourgoulianis, K.I. Calprotectin in Lung Diseases. Int. J. Mol. Sci. 2021, 22, 1706. https://doi.org/10.3390/ijms22041706
Kotsiou OS, Papagiannis D, Papadopoulou R, Gourgoulianis KI. Calprotectin in Lung Diseases. International Journal of Molecular Sciences. 2021; 22(4):1706. https://doi.org/10.3390/ijms22041706
Chicago/Turabian StyleKotsiou, Ourania S., Dimitrios Papagiannis, Rodanthi Papadopoulou, and Konstantinos I. Gourgoulianis. 2021. "Calprotectin in Lung Diseases" International Journal of Molecular Sciences 22, no. 4: 1706. https://doi.org/10.3390/ijms22041706
APA StyleKotsiou, O. S., Papagiannis, D., Papadopoulou, R., & Gourgoulianis, K. I. (2021). Calprotectin in Lung Diseases. International Journal of Molecular Sciences, 22(4), 1706. https://doi.org/10.3390/ijms22041706