
Development of Atropa belladonna L. Plants with High-Yield Hyoscyamine and without Its Derivatives Using the CRISPR/Cas9 System


 and
and
Abstract
:1. Introduction
2. Results and Discussion
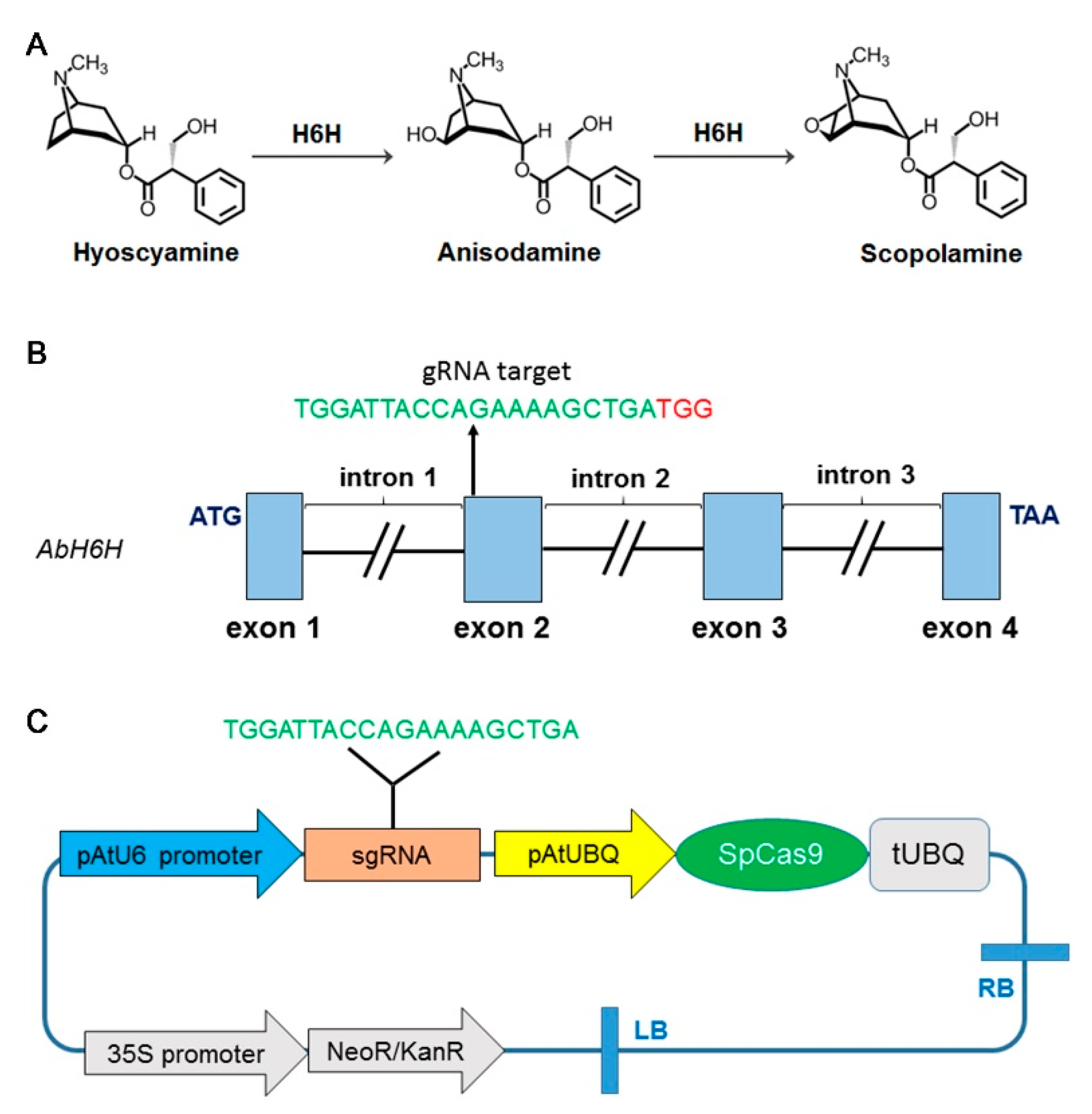
2.1. Cloning and Analysis of the Genomic Sequence of A. belladonna H6H Gene
2.2. sgRNA Design and Vector Construction
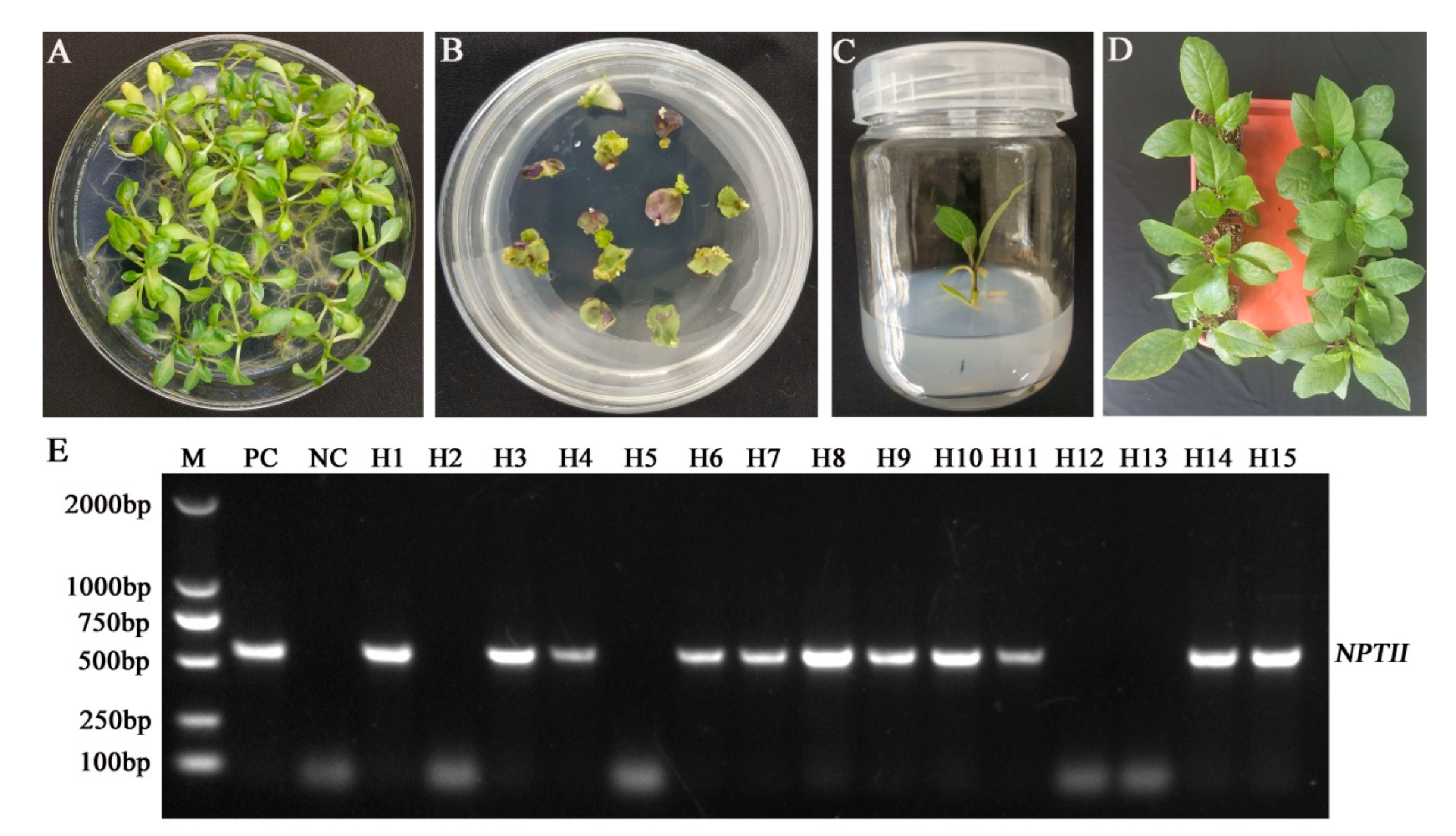
2.3. Generation of Transgenic Atropa belladonna Plants
2.4. Detection of CRISPR/Cas9-Mediated Mutagenesis in AbH6H
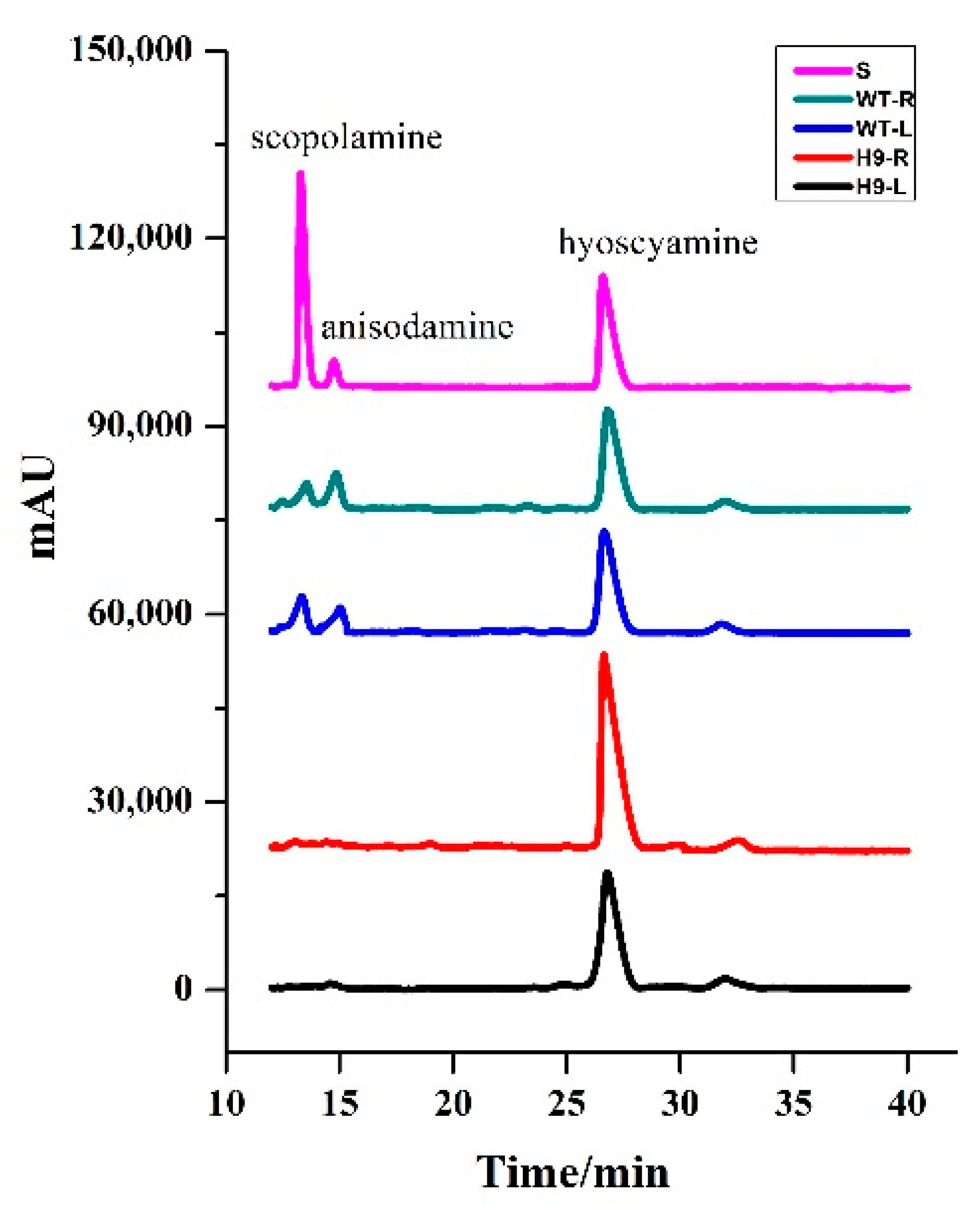
2.5. Analysis of Tropane Alkaloids
3. Material and Methods
3.1. Cloning and Analysis of the Genomic Sequence of the H6H Gene of A. belladonna
3.2. Design of sgRNA and Construction of CRISPR/Cas9 Vector
3.3. Establishment of Transgenic Plants of Atropa belladonna
3.4. Molecular Analysis of Transformants
3.5. Alkaloid Detection
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| PAM | protospacer-adjacent motif |
| NPTII | neomycin phosphotransferase II |
References
- Rita, P.; Animesh, D.K. An Updated Overview on Atropa belladonna L. Int. Res. J. Pharm. 2011, 2, 11–17. [Google Scholar]
- Geiger. Hesse Darstellung des Atropins. Ann. Pharm. 1833, 5, 43–81. [Google Scholar] [CrossRef] [Green Version]
- Lossen, W.; Lossen, W.; Lossen, W.; Lossen, W. Ueber das Atropin. Ann. Chem. Pharm. 1864, 131, 43–49. [Google Scholar] [CrossRef]
- Wolffenstein, R. Die Pflanzenalkaloide; Springer Berlin Heidelberg: Berlin/Heidelberg, Germany, 1922; ISBN 978-3-642-90592-6. [Google Scholar]
- Sweta, V.R.; Lakshmi, T. Mini Review Article Pharmacological profile of tropane alkaloids. J. Chem. Pharm. Res. 2015, 7, 117–119. [Google Scholar]
- Yun, D.; Hashimoto, T.; Yamada, Y. Metabolic engineering of medicinal plants: Transgenic Atropa belladonna with an improved alkaloid composition. Proc. Natl. Acad. Sci. USA 1992, 89, 11799–11803. [Google Scholar] [CrossRef] [Green Version]
- Poupko, J.M.; Baskin, S.I.; Moore, E. The pharmacological properties of anisodamine. J. Appl. Toxicol. 2007, 27, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, T.; Yamada, Y. Hyoscyamine 6β-Hydroxylase, a 2-Oxoglutarate-Dependent Dioxygenase, in Alkaloid-Producing Root Cultures. Plant Physiol. 1986, 81, 619–625. [Google Scholar] [CrossRef]
- Zhang, L.; Ding, R.; Chai, Y.; Bonfill, M.; Moyano, E.; Oksman-Caldentey, K.M.; Xu, T.; Pi, Y.; Wang, Z.; Zhang, H.; et al. Engineering tropane biosynthetic pathway in Hyoscyamus niger hairy root cultures. Proc. Natl. Acad. Sci. USA 2004, 101, 6786–6791. [Google Scholar] [CrossRef] [Green Version]
- Lan, X.; Zeng, J.; Liu, K.; Zhang, F.; Bai, G.; Chen, M.; Liao, Z.; Huang, L. Comparison of two hyoscyamine 6β-hydroxylases in engineering scopolamine biosynthesis in root cultures of Scopolia lurida. Biochem. Biophys. Res. Commun. 2018, 497, 25–31. [Google Scholar] [CrossRef]
- Xia, K.; Liu, X.; Zhang, Q.; Qiang, W.; Guo, J.; Lan, X.; Chen, M.; Liao, Z. Promoting scopolamine biosynthesis in transgenic Atropa belladonna plants with pmt and h6h overexpression under field conditions. Plant Physiol. Biochem. 2016, 106, 46–53. [Google Scholar] [CrossRef]
- Manghwar, H.; Lindsey, K.; Zhang, X.; Jin, S. CRISPR/Cas System: Recent Advances and Future Prospects for Genome Editing. Trends Plant Sci. 2019, 24, 1102–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalil, A.M. The genome editing revolution: Review. J. Genet. Eng. Biotechnol. 2020, 18, 68. [Google Scholar] [CrossRef] [PubMed]
- Bartsevich, V.V.; Morris, J.; Tomberlin, G.; Turner, C.; Sharer, W.; Nicholson, M.G.; Smith, J.; Kollu, N.; James, R.; Armstrong, R.; et al. Meganucleases as an Efficient Tool for Genome Editing. Mol. Ther. 2016, 24, S230. [Google Scholar] [CrossRef]
- Sun, N.; Zhao, H. Transcription activator-like effector nucleases (TALENs): A highly efficient and versatile tool for genome editing. Biotechnol. Bioeng. 2013, 110, 1811–1821. [Google Scholar] [CrossRef]
- Carroll, D. Genome Engineering With Zinc-Finger Nucleases. Genetics 2011, 188, 773–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Tan, H.; Li, Q.; Chen, J.; Gao, S.; Wang, Y.; Chen, W.; Zhang, L. CRISPR/Cas9-mediated efficient targeted mutagenesis of RAS in Salvia miltiorrhiza. Phytochemistry 2018, 148, 63–70. [Google Scholar] [CrossRef]
- Kim, D.; Alptekin, B.; Budak, H. CRISPR/Cas9 genome editing in wheat. Funct. Integr. Genom. 2018, 18, 31–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Zhang, H.; Botella, J.R.; Zhu, J.-K. Generation of new glutinous rice by CRISPR/Cas9-targeted mutagenesis of the Waxy gene in elite rice varieties. J. Integr. Plant Biol. 2018, 60, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Long, L.; Guo, D.D.; Gao, W.; Yang, W.W.; Hou, L.P.; Ma, X.N.; Miao, Y.C.; Botella, J.R.; Song, C.P. Optimization of CRISPR/Cas9 genome editing in cotton by improved sgRNA expression. Plant Methods 2018, 14, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, K.I.; Yun, D.J.; Chen, X.Y.; Yamada, Y.; Hashimoto, T. An Atropa belladonna hyoscyamine 6β-hydroxylase gene is differentially expressed in the root pericycle and anthers. Plant Mol. Biol. 1999, 40, 141–152. [Google Scholar] [CrossRef]
- Hsu, P.D.; Scott, D.A.; Weinstein, J.A.; Ran, F.A.; Konermann, S.; Agarwala, V.; Li, Y.; Fine, E.J.; Wu, X.; Shalem, O.; et al. DNA targeting specificity of RNA-guided Cas9 nucleases. Nat. Biotechnol. 2013, 31, 827–832. [Google Scholar] [CrossRef]
- Hood, E.E.; Gelvin, S.B.; Melchers, L.S.; Hoekema, A. New Agrobacterium helper plasmids for gene transfer to plants. Transgenic Res. 1993, 2, 208–218. [Google Scholar] [CrossRef]
- An, G.; Ebert, P.R.; Mitra, A.; Ha, S.B. Binary vectors. In Plant Molecular Biology Manual; Springer: Dordrecht, The Netherlands, 1989; pp. 29–47. [Google Scholar]
- Ma, X.; Chen, L.; Zhu, Q.; Chen, Y.; Liu, Y.-G. Rapid Decoding of Sequence-Specific Nuclease-Induced Heterozygous and Biallelic Mutations by Direct Sequencing of PCR Products. Mol. Plant 2015, 8, 1285–1287. [Google Scholar] [CrossRef] [Green Version]
- Zhao, T.; Li, S.; Wang, J.; Zhou, Q.; Yang, C.; Bai, F.; Lan, X.; Chen, M.; Liao, Z. Engineering tropane alkaloid production based on metabolic characterization of ornithine decarboxylase in atropa belladonna. ACS Synth. Biol. 2020, 9, 437–448. [Google Scholar] [CrossRef]
- Bedewitz, M.A.; Jones, A.D.; Dauria, J.C.; Barry, C.S. Tropinone synthesis via an atypical polyketide synthase and P450-mediated cyclization. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, F.; Yang, C.; Yuan, L.; Xiang, D.; Lan, X.; Chen, M.; Liao, Z. A Phenylpyruvic Acid Reductase Is Required for Biosynthesis of Tropane Alkaloids. Org. Lett. 2018, 20, 7807–7810. [Google Scholar] [CrossRef] [PubMed]
- Qiu, F.; Zeng, J.; Wang, J.; Huang, J.P.; Zhou, W.; Yang, C.; Lan, X.; Chen, M.; Huang, S.X.; Kai, G.; et al. Functional genomics analysis reveals two novel genes required for littorine biosynthesis. N. Phytol. 2020, 225, 1906–1914. [Google Scholar] [CrossRef] [PubMed]
- Porebski, S.; Bailey, L.G.; Baum, B.R. Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components. Plant Mol. Biol. Rep. 1997, 15, 8–15. [Google Scholar] [CrossRef]
- Mao, Y.; Zhang, Z.; Feng, Z.; Wei, P.; Zhang, H.; Botella, J.R.; Zhu, J.-K. Development of germ-line-specific CRISPR-Cas9 systems to improve the production of heritable gene modifications in Arabidopsis. Plant Biotechnol. J. 2016, 14, 519–532. [Google Scholar] [CrossRef] [Green Version]
- Song, G.Q.; Walworth, A.E. Agrobacterium tumefaciens-mediated transformation of Atropa belladonna. Plant Cell Tissue Organ Cult. 2013, 115, 107–113. [Google Scholar] [CrossRef]
- Youchang, Z.; Changhui, F.; Shu, B.; Xiaojiao, W.; Xianda, Y.; Cheng, Z.; Hongde, Q. An Improved TPS Method for Rapid DNA Extraction from Cotton Leaves. Cotton Sci. 2016, 28, 413–417. [Google Scholar] [CrossRef]
- Zarate, R.; Hermosin, B.; Cantos, M.; Troncoso, A. Tropane Alkaloid Distribution in Atropa baetica Plants. J. Chem. Ecol. 1997, 23, 2059–2066. [Google Scholar] [CrossRef]
- Bai, F.; Li, S.; Yang, C.; Zhao, T.; Zhang, T.; Lan, X.; Chen, M.; Liao, Z. Overexpression of the AbSAUR1 gene enhanced biomass production and alkaloid yield in Atropa belladonna. Ind. Crops Prod. 2019, 140, 111705. [Google Scholar] [CrossRef]
- Zeng, J.; Wang, J.; Liu, X.; Qin, J.; Lan, X.; Chen, M.; Liao, Z. An auxin-responsive transcription factor AbLBD1 promotes the development of lateral roots and reduces the biosynthesis of tropane alkaloids in Atropa belladonna. Plant Cell. Tissue Organ Cult. 2020, 142, 559–569. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Purpose | Primer Sequence (5′-3′) |
|---|---|---|
| FH6H | Gene cloning | ATGGCTACTCTTGTCTCAAATTG |
| RH6H | TTAGGCATTAATTTTATATGGCTTAAC | |
| NPTII-F (XhoI) | Vector construction | CGCCTCGAGATGATTGAACAAGATGGATTG |
| NPTII-R (XhoI) | CGCCTCGAGTCAGAAGAACTCGTCAAGAAG | |
| AbH6H-F | Vector construction | GATTGTGGATTACCAGAAAAGCTGA |
| AbH6H-R | AAACTCAGCTTTTCTGGTAATCCAC | |
| AbH6H-knock-F | PCR detection | CATGCGCATGATATGTGAAC |
| AbH6H-knock-R | CCGGATGAAGGCGATTCAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, L.; Zhang, Q.; Jiang, C.; Zheng, Y.; Zuo, Y.; Qin, J.; Liao, Z.; Deng, H. Development of Atropa belladonna L. Plants with High-Yield Hyoscyamine and without Its Derivatives Using the CRISPR/Cas9 System. Int. J. Mol. Sci. 2021, 22, 1731. https://doi.org/10.3390/ijms22041731
Zeng L, Zhang Q, Jiang C, Zheng Y, Zuo Y, Qin J, Liao Z, Deng H. Development of Atropa belladonna L. Plants with High-Yield Hyoscyamine and without Its Derivatives Using the CRISPR/Cas9 System. International Journal of Molecular Sciences. 2021; 22(4):1731. https://doi.org/10.3390/ijms22041731
Chicago/Turabian StyleZeng, Lingjiang, Qiaozhuo Zhang, Chunxue Jiang, Yueyue Zheng, Youwei Zuo, Jianbo Qin, Zhihua Liao, and Hongping Deng. 2021. "Development of Atropa belladonna L. Plants with High-Yield Hyoscyamine and without Its Derivatives Using the CRISPR/Cas9 System" International Journal of Molecular Sciences 22, no. 4: 1731. https://doi.org/10.3390/ijms22041731
APA StyleZeng, L., Zhang, Q., Jiang, C., Zheng, Y., Zuo, Y., Qin, J., Liao, Z., & Deng, H. (2021). Development of Atropa belladonna L. Plants with High-Yield Hyoscyamine and without Its Derivatives Using the CRISPR/Cas9 System. International Journal of Molecular Sciences, 22(4), 1731. https://doi.org/10.3390/ijms22041731