
Dynamics of Central Remyelination and Treatment Evolution in a Model of Multiple Sclerosis with Optic Coherence Tomography

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
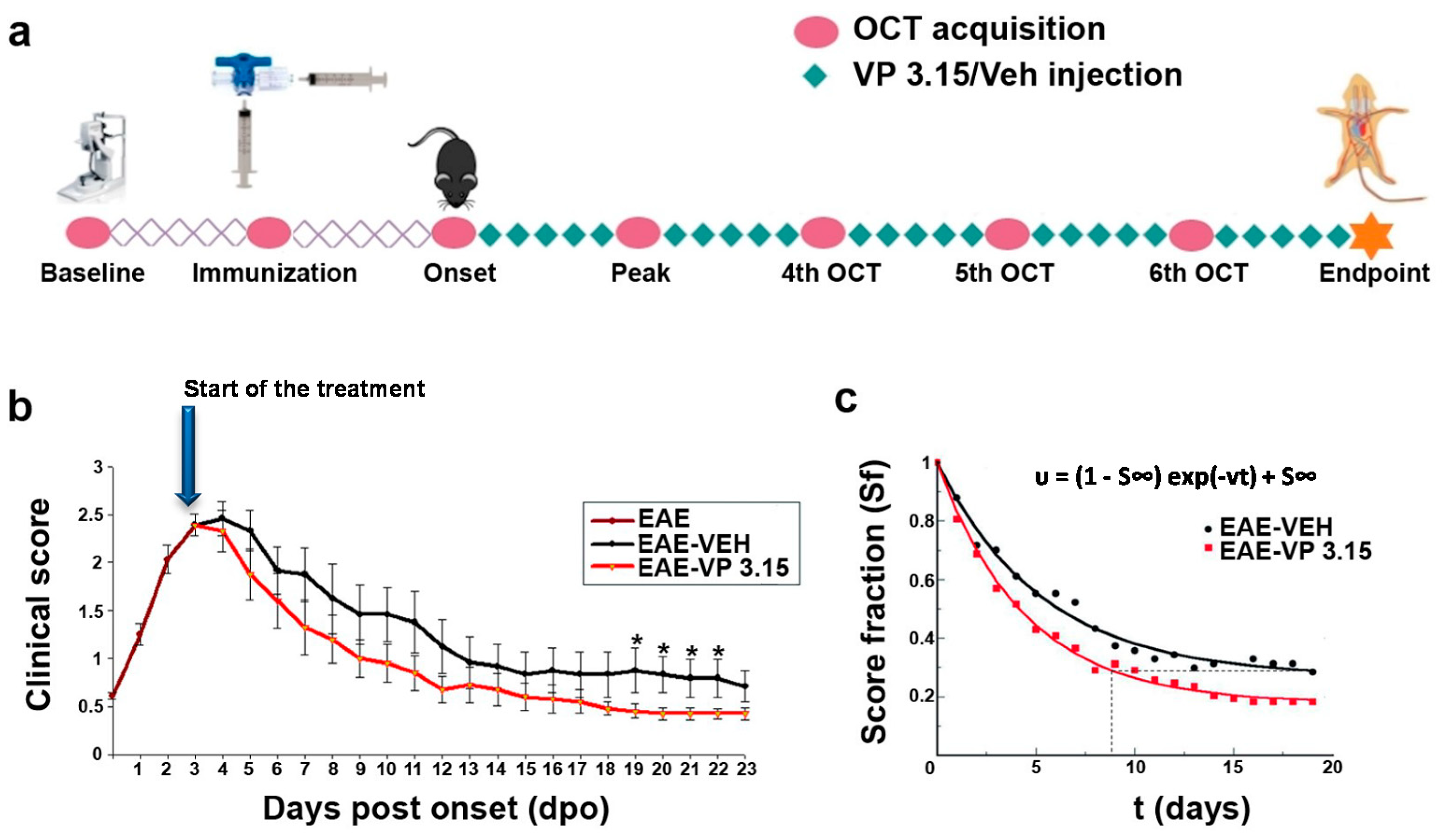
2.1. The PDE7/GSK3 Dual Inhibitor VP3.15 Ameliorates Clinical Course of Experimental Autoimmune Encephalomyelitis (EAE)
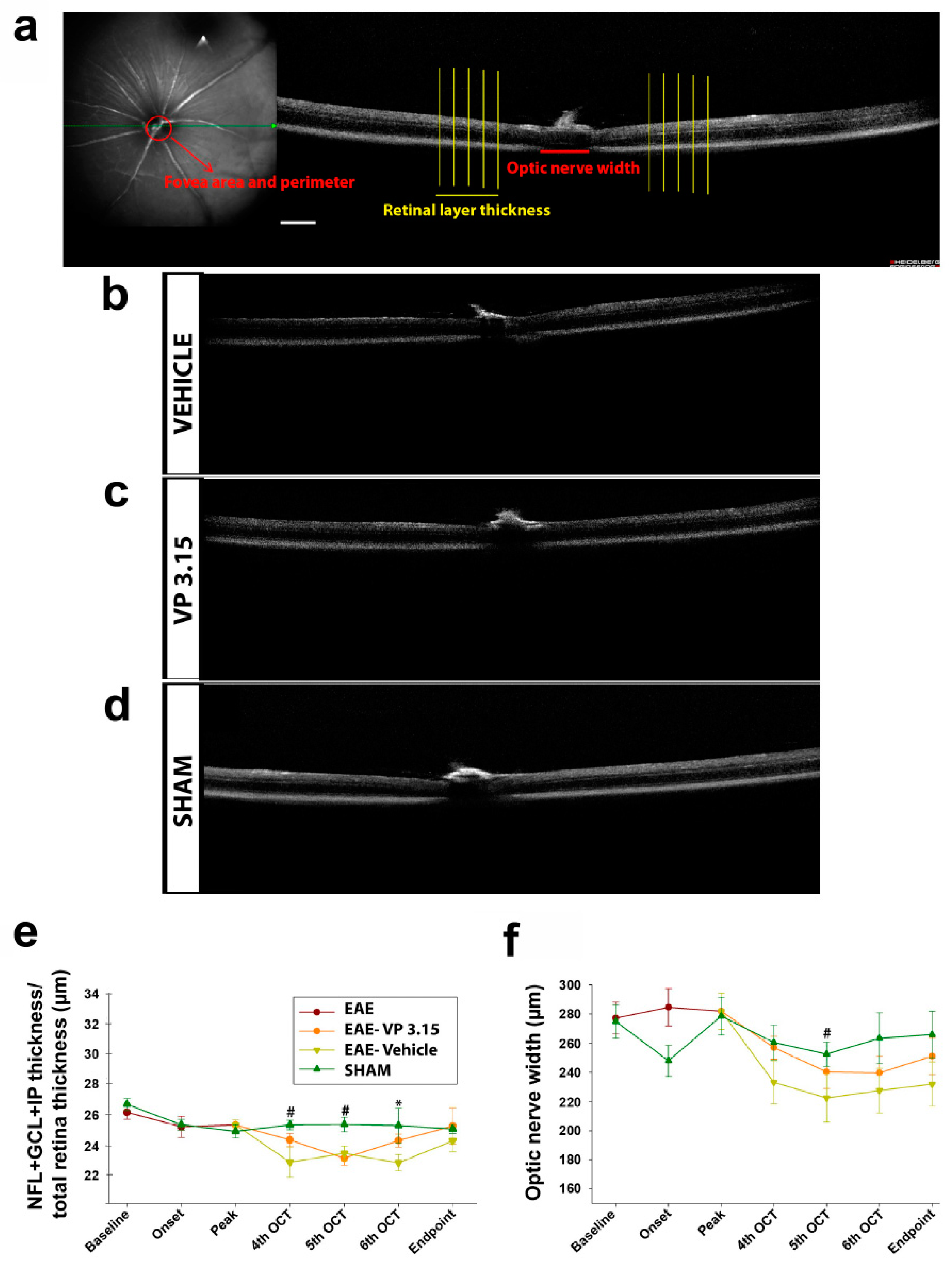
2.2. Retinal and Optic Nerve Changes Can Be Monitored Using Optic Coherence Tomography (OCT) in the EAE Model
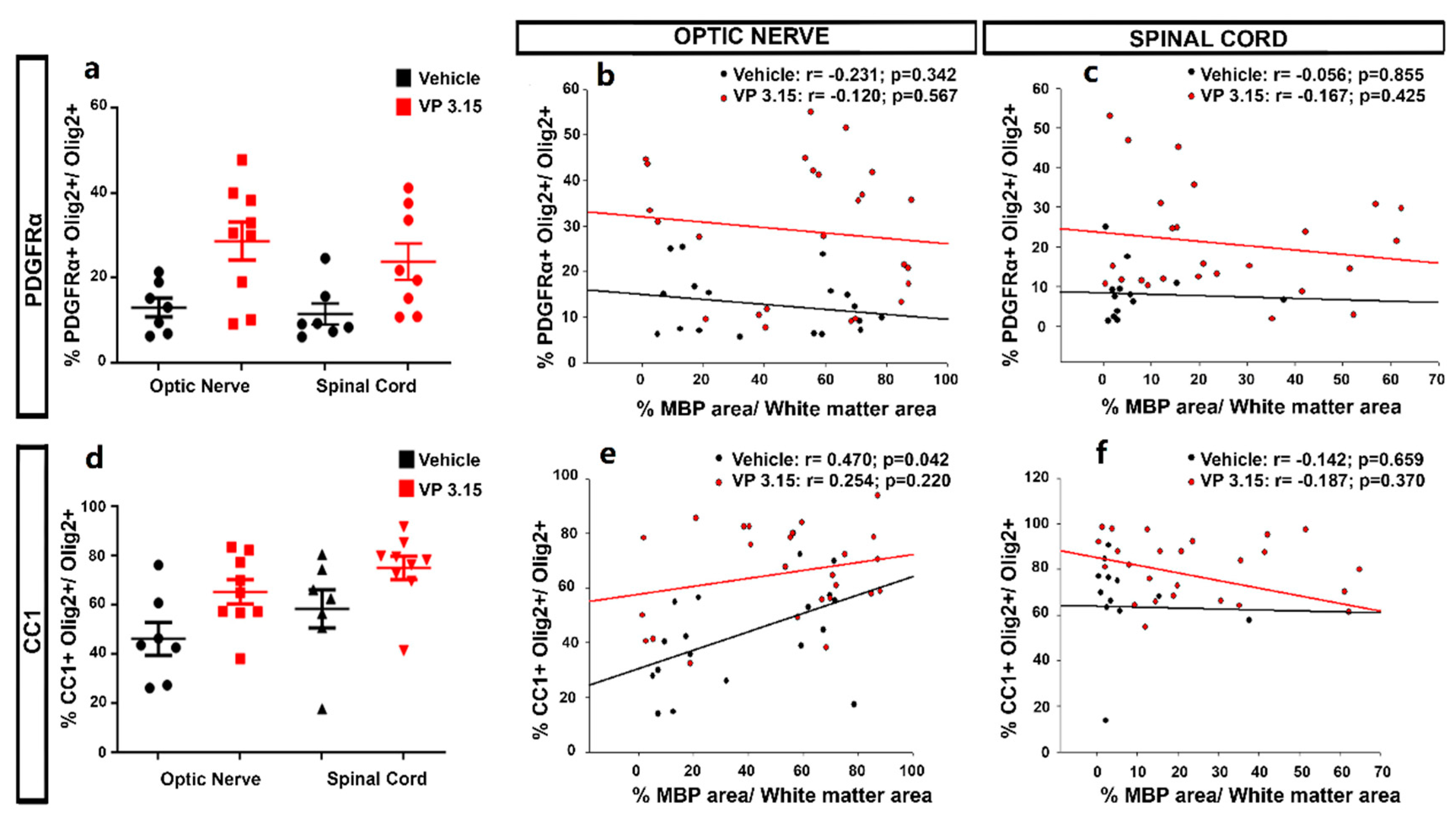
2.3. The Remyelinating Role and the Effect in the Oligodendrocyte Lineage of VP3.15
2.4. VP3.15 Treatment Modifies Microglial Activation State
3. Discussion
4. Material and Methods
4.1. Induction of EAE and Treatment
4.2. OCT Data Acquisition and Analysis
4.3. Tissue Section
4.4. Eriochrome Cyanine Staining
4.5. Immunohistochemistry
4.6. Tissue Analysis and Cell Quantification
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Browne, P.; Chandraratna, D.; Angood, C.; Tremlett, H.; Baker, C.; Taylor, B.V.; Thompson, A.J. Atlas of Multiple Sclerosis 2013: A growing global problem with widespread inequity. Neurology 2014, 83, 1022–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sospedra, M.; Martin, R. Immunology of multiple sclerosis. Annu. Rev. Immunol. 2005, 23, 683–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, D.Y.S.; Vereyken, E.J.F.; Glim, J.E.; Heijnen, P.D.A.M.; Moeton, M.; van der Valk, P.; Amor, S.; Teunissen, C.E.; van Horssen, J.; Dijkstra, C.D. Macrophages in inflammatory multiple sclerosis lesions have an intermediate activation status. J. Neuroinflammation 2013, 10, 809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clemente, D.; Ortega, M.C.; Melero-Jerez, C.; de Castro, F. The effect of glia-glia interactions on oligodendrocyte precursor cell biology during development and in demyelinating diseases. Front. Cell. Neurosci. 2013, 7, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miron, V.E.; Boyd, A.; Zhao, J.-W.; Yuen, T.J.; Ruckh, J.M.; Shadrach, J.L.; Van Wijngaarden, P.; Wagers, A.J.; Williams, A.; van Wijngaarden, P.; et al. Europe PMC Funders Group M2 microglia / macrophages drive oligodendrocyte differentiation during CNS remyelination. Nat. Neurosci. 2013, 16, 1211–1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, C.; Jian, C.; Liao, Y.; Huang, Q.; Wu, Y.; Liu, X.; Zou, D.; Wu, Y. The role of microglia in multiple sclerosis. Neuropsychiatr. Dis. Treat. 2017, 13, 1661–1667. [Google Scholar] [CrossRef] [Green Version]
- Hart, F.M.; Bainbridge, J. Current and emerging treatment of multiple sclerosis. Am. J. Manag. Care 2016, 22, s159–s170. [Google Scholar]
- Green, A.J.; Gelfand, J.M.; Cree, B.A.; Bevan, C.; Boscardin, W.J.; Mei, F.; Inman, J.; Arnow, S.; Devereux, M.; Abounasr, A.; et al. Clemastine fumarate as a remyelinating therapy for multiple sclerosis (ReBUILD): A randomised, controlled, double-blind, crossover trial. Lancet 2017, 390, 2481–2489. [Google Scholar] [CrossRef] [Green Version]
- de Castro, F.; Josa-Prado, F. Regulation of oligodendrocyte differentiation: New targets for drug discovery in remyelination. Emerg. Drugs Targets Mult. Scler. 2019, 10, 222–240. [Google Scholar]
- Zhang, Y.; Argaw, A.T.; Gurfein, B.T.; Zameer, A.; Snyder, B.J.; Ge, C.; Lu, Q.R.; Rowitch, D.H.; Raine, C.S.; Brosnan, C.F.; et al. Notch1 signaling plays a role in regulating precursor differentiation during CNS remyelination. Proc. Natl. Acad. Sci. USA 2009, 106, 19162–19167. [Google Scholar] [CrossRef] [Green Version]
- Jensen, S.K.; Yong, V.W. Activity-Dependent and Experience-Driven Myelination Provide New Directions for the Management of Multiple Sclerosis. Trends Neurosci. 2016, 39, 356–365. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.K.; Jarjour, A.A.; Nait Oumesmar, B.; Kerninon, C.; Williams, A.; Krezel, W.; Kagechika, H.; Bauer, J.; Zhao, C.; Baron-Van Evercooren, A.; et al. Retinoid X receptor gamma signaling accelerates CNS remyelination. Nat. Neurosci. 2011, 14, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Ge, H.; Liu, W.; Zhu, H.; Chen, Y.; Zhang, X.; Yang, Y.; Yin, Y.; Chen, W.; Wu, W.; et al. M2 microglia promotes neurogenesis and oligodendrogenesis from neural stem/progenitor cells via the PPARgamma signaling pathway. Oncotarget 2017, 8, 19855–19865. [Google Scholar] [CrossRef] [Green Version]
- De La Fuente, A.G.; Lange, S.; Silva, M.E.; Gonzalez, G.A.; Tempfer, H.; van Wijngaarden, P.; Zhao, C.; Di Canio, L.; Trost, A.; Bieler, L.; et al. Pericytes Stimulate Oligodendrocyte Progenitor Cell Differentiation during CNS Remyelination. Cell Rep. 2017, 20, 1755–1764. [Google Scholar] [CrossRef] [PubMed]
- Behrangi, N.; Namvar, N.; Ataei, M.; Dizaji, S.; Javdani, G.; Sanati, M.H. MMP9 Gene Expression Variation by Ingesting Tart Cherry and P-Coumaric Acid During Remyelination in the Cuprizone Mouse Model. Acta Med. Iran. 2017, 55, 539–549. [Google Scholar]
- Kuboyama, K.; Tanga, N.; Suzuki, R.; Fujikawa, A.; Noda, M. Protamine neutralizes chondroitin sulfate proteoglycan-mediated inhibition of oligodendrocyte differentiation. PLoS ONE 2017, 12, e0189164. [Google Scholar] [CrossRef] [PubMed]
- De Castro, F.; Ortega, M.C.; Bribián, A. Regulation of oligodendrocyte precursor migration during development, in adulthood and in pathology. Cell Mol. Life Sci. 2013, 70, 4355–4368. [Google Scholar] [CrossRef]
- Clemente, D.; Ortega, M.C.; Arenzana, F.J.; de Castro, F. FGF-2 and anosmin-1 are selectively expressed in different types of multiple sclerosis lesions. J. Neurosci. 2011, 31, 14899–14909. [Google Scholar] [CrossRef] [Green Version]
- Absinta, M.; Sati, P.; Reich, D.S. Advanced MRI and staging of multiple sclerosis lesions. Nat. Rev. Neurol. 2016, 12, 358–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franklin, R.J.M.; ffrench-Constant, C. Regenerating CNS myelin—From mechanisms to experimental medicines. Nat. Rev. Neurosci. 2017, 18, 753–769. [Google Scholar] [CrossRef]
- Hein, K.; Gadjanski, I.; Kretzschmar, B.; Lange, K.; Diem, R.; Sa, M.B. Immunology and Microbiology An Optical Coherence Tomography Study on Degeneration of Retinal Nerve Fiber Layer in Rats with Autoimmune Optic Neuritis. Investig. Ophthalmol. Vis. Sci. 2012, 53, 157–163. [Google Scholar] [CrossRef] [Green Version]
- Oertel, F.C.; Outteryck, O.; Knier, B.; Zimmermann, H.; Borisow, N.; Bellmann-strobl, J.; Blaschek, A.; Jarius, S.; Reindl, M.; Ruprecht, K.; et al. Optical coherence tomography in myelin- seropositive patients: A longitudinal study. J. Neuroiflamm. 2019, 5, 1–9. [Google Scholar]
- London, F.; Zéphir, H.; Drumez, E.; Labreuche, J.; Hadhoum, N.; Lannoy, J.; Hodel, J.; Vermersch, P.; Pruvo, J.-P.; Leclerc, X.; et al. Optical coherence tomography: A window to the optic nerve in clinically isolated syndrome. Brain 2019, 142, 903–915. [Google Scholar] [CrossRef]
- Ratchford, J.N.; Quigg, M.E.; Conger, A.; Frohman, T.; Frohman, E.; Balcer, L.J.; Calabresi, P.A.; Kerr, D.A. Optical coherence tomography helps differentiate neuromyelitis optica and MS optic neuropathies. Neurology 2009, 73, 302–308. [Google Scholar] [CrossRef] [Green Version]
- Tugcu, B.; Soysal, A.; Murat, K.; Yuksel, B.; Arpaci, B. Assessment of structural and functıonal vısual outcomes ın relapsıng remıttıng multıple sclerosıs wıth vısual evoked potentıals and optical coherence tomography. J. Neurol. Sci. 2013, 335, 182–185. [Google Scholar] [CrossRef] [PubMed]
- Di Maggio, G.; Santangelo, R.; Guerrieri, S.; Bianco, M.; Ferrari, L.; Medaglini, S.; Rodegher, M.; Colombo, B.; Moiola, L.; Chieffo, R.; et al. Optical coherence tomography and visual evoked potentials: Which is more sensitive in multiple sclerosis? Mult. Scler. 2014, 20, 1342–1347. [Google Scholar] [CrossRef] [PubMed]
- Ashtari, F.; Emami, P.; Akbari, M. Association between retinal nerve fiber layer thickness and magnetic resonance imaging findings and intelligence in patients with multiple sclerosis. Adv. Biomed. Res. 2015, 4, 223. [Google Scholar]
- Manogaran, P.; Hanson, J.V.M.; Olbert, E.D.; Egger, C.; Wicki, C.; Gerth-Kahlert, C.; Landau, K.; Schippling, S. Optical coherence tomography and magnetic resonance imaging in multiple sclerosis and neuromyelitis optica spectrum disorder. Int. J. Mol. Sci. 2016, 17, 1894. [Google Scholar] [CrossRef] [Green Version]
- Puthenparampil, M.; Federle, L.; Poggiali, D.; Miante, S.; Signori, A.; Pilotto, E.; Rinaldi, F.; Perini, P.; Sormani, P.; Midena, E.; et al. Trans-synaptic degeneration in the optic pathway. A study in clinically isolated syndrome and early relapsing-remitting multiple sclerosis with or without optic neuritis. PLoS ONE 2017, 12, e0183957. [Google Scholar]
- Talebi, M.; Nikanfar, M.; Sorkhabi, R.; Sharifipour, E.; Bahrebar, M.; Kiavar, A.; Andalib, S.; Khanli, H.M. Optic coherence tomography findings in relapsing-remitting multiple sclerosis patients of the northwest of Iran. Iran. J. Neurol. 2013, 12, 81–86. [Google Scholar]
- Nolan-kenney, R.C.; Liu, M.; Akhand, O.; Calabresi, P.A.; Paul, F.; Petzold, A.; Balk, L.; Brandt, A.U.; Martínez-lapiscina, E.H.; Saidha, S.; et al. Optimal Intereye Difference Thresholds by Optical Coherence Tomography in Multiple Sclerosis: An International Study. Ann. Neurol. 2019, 85, 618–629. [Google Scholar] [CrossRef] [PubMed]
- Gordon-Lipkin, E.; Calabresi, P.A. Optical coherence tomography: A quantitative tool to measure neurodegeneration and facilitate testing of novel treatments for tissue protection in multiple sclerosis. J. Neuroimmunol. 2017, 304, 93–96. [Google Scholar] [CrossRef]
- Pulicken, M.; Gordon-Lipkin, E.; Balcer, L.J.; Frohman, E.; Cutter, G.; Calabresi, P.A. Optical coherence tomography and disease subtype in multiple sclerosis. Neurology 2007, 69, 2085–2092. [Google Scholar] [CrossRef] [PubMed]
- Palomo, V.; Pérez, D.I.; Pérez, C.; Morales-García, J.A.; Soteras, I.; Alonso-Gil, S.; Encinas, A.; Castro, A.; Campillo, N.E.; Pérez-Castillo, A.; et al. 5-Imino-1,2,4-thiadiazoles: First small molecules As substrate competitive inhibitors of glycogen synthase kinase 3. J. Med. Chem. 2012, 55, 1645–1661. [Google Scholar] [CrossRef] [PubMed]
- Medina-Rodriguez, E.M.; Bribián, A.; Boyd, A.; Palomo, V.; Pastor, J.; Lagares, A.; Gil, C.; Martínez, A.; Williams, A.; de Castro, F. Promoting in vivo remyelination with small molecules: A neuroreparative pharmacological treatment for Multiple Sclerosis. Sci. Rep. 2017, 7, 1–14. [Google Scholar]
- Medina-Rodríguez, E.M.; Arenzana, F.J.; Pastor, J.; Redondo, M.; Palomo, V.; García De Sola, R.; Gil, C.; Martínez, A.; Bribián, A.; de Castro, F. Inhibition of endogenous phosphodiesterase 7 promotes oligodendrocyte precursor differentiation and survival. Cell. Mol. Life Sci. 2013, 70, 3449–3462. [Google Scholar] [CrossRef]
- Sánchez-Cruz, A.; Villarejo-Zori, B.; Marchena, M.; Zaldivar-Díez, J.; Palomo, V.; Gil, C.; Lizasoain, I.; De La Villa, P.; Martínez, A.; De La Rosa, E.J.; et al. Modulation of GSK-3 provides cellular and functional neuroprotection in the rd10 mouse model of retinitis pigmentosa. Mol. Neurodegener. 2018, 13, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Martín-Álvarez, R.; Paúl-Fernández, N.; Palomo, V.; Gil, C.; Martínez, A.; Mengod, G. A preliminary investigation of phoshodiesterase 7 inhibitor VP3.15 as therapeutic agent for the treatment of experimental autoimmune encephalomyelitis mice. J. Chem. Neuroanat. 2017, 80, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Ellsworth, S.G.; Yalamanchali, A.; Zhang, H.; Grossman, S.A.; Hobbs, R.; Jin, J.-Y. Comprehensive Analysis of the Kinetics of Radiation-Induced Lymphocyte Loss in Patients Treated with External Beam Radiation Therapy. Radiat. Res. 2020, 193, 73–81. [Google Scholar] [CrossRef]
- Petzold, A.; Balcer, L.J.; Calabresi, P.A.; Costello, F.; Frohman, T.C.; Frohman, E.M.; Martinez-lapiscina, E.H.; Green, A.J. Retinal layer segmentation in multiple sclerosis: A systematic review and meta-analysis. Lancet Neurol. 2017, 16, 797–812. [Google Scholar] [CrossRef] [Green Version]
- Mecha, M.; Carrillo-Salinas, F.J.; Feliú, A.; Mestre, L.; Guaza, C. Microglia activation states and cannabinoid system: Therapeutic implications. Pharmacol. Ther. 2016, 166, 40–55. [Google Scholar] [CrossRef]
- Mecha, M.; Feliú, A.; Machín, I.; Cordero, C.; Carrillo-Salinas, F.; Mestre, L.; Hernández-Torres, G.; Ortega-Gutiérrez, S.; López-Rodríguez, M.; de Castro, F.; et al. 2-AG limits Theiler’s virus induced acute neuroinflammation by modulating microglia and promoting MDSCs. Glia 2018, 66, 1447–1463. [Google Scholar] [CrossRef]
- Silbermann, E.; Wooliscroft, L.; Bourdette, D. Using the Anterior Visual System to Assess Neuroprotection and Remyelination in Multiple Sclerosis Trials. Curr. Neurol. Neurosci. Rep. 2017, 18, 1–9. [Google Scholar] [CrossRef]
- Martinez-Lapiscina, E.H.; Arnow, S.; Wilson, J.A.; Saidha, S.; Preiningerova, J.L.; Oberwahrenbrock, T.; Brandt, A.U.; Pablo, L.E.; Guerrieri, S.; Gonzalez, I.; et al. Retinal thickness measured with optical coherence tomography and risk of disability worsening in multiple sclerosis: A cohort study. Lancet Neurol. 2016, 15, 574–584. [Google Scholar] [CrossRef] [Green Version]
- Villoslada, P.; Cuneo, A.; Gelfand, J.; Hauser, S.L.; Green, A. Color vision is strongly associated with retinal thinning in multiple sclerosis. Mult. Scler. 2012, 18, 991–999. [Google Scholar] [CrossRef]
- Green, A.J.; McQuaid, S.; Hauser, S.L.; Allen, I.V.; Lyness, R. Ocular pathology in multiple sclerosis: Retinal atrophy and inflammation irrespective of disease duration. Brain 2010, 133, 1591–1601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthews, P.M.; Datta, G. Positron-emission tomography molecular imaging of glia and myelin in drug discovery for multiple sclerosis. Expert Opin. Drug Discov. 2015, 10, 557–570. [Google Scholar] [CrossRef] [PubMed]
- Bove, R.M.; Green, A.J. Remyelinating Pharmacotherapies in Multiple Sclerosis. Neurotherapeutics 2017, 14, 894–904. [Google Scholar] [CrossRef] [Green Version]
- Leferink, P.S.; Heine, V.M. The Healthy and Diseased Microenvironments Regulate Oligodendrocyte Properties Implications for Regenerative Medicine. Am. J. Pathol. 2018, 188, 39–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falcão, A.M.; Van Bruggen, D.; Marques, S.; Meijer, M.; Jäkel, S.; Agirre, E.; Floriddia, E.M.; Vanichkina, D.P.; Williams, A.; Guerreiro-Cacais, A.O.; et al. Disease-specific oligodendrocyte lineage cells arise in multiple sclerosis. Nat. Med. 2018, 24, 1837–1844. [Google Scholar] [CrossRef]
- Zeis, T.; Howell, O.W.; Reynolds, R.; Schaeren-wiemers, N. Molecular pathology of Multiple Sclerosis lesions reveals a heterogeneous expression pattern of genes involved in oligodendrogliogenesis IdL. Exp. Neurol. 2018, 305, 76–88. [Google Scholar] [CrossRef]
- Jäkel, S.; Agirre, E.; Falcão, A.M.; Bruggen, D.; Lee, K.W.; Knuesel, I.; Malhotra, D.; Williams, A.; Castelo-branco, G. Altered human oligodendrocyte heterogeneity in multiple sclerosis. Nature 2019, 566, 543–547. [Google Scholar] [CrossRef]
- Yeung, M.S.Y.; Djelloul, M.; Steiner, E.; Bernard, S.; Possnert, G.; Brundin, L.; Frisén, J. Europe PMC Funders Group Oligodendrocyte generation dynamics in multiple sclerosis. Nature 2019, 566, 538–542. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, M.Ě.; Lowery, R.L.; Majewska, A.K. Microglial interactions with synapses are modulated by visual experience. PLoS Biol. 2010, 8, e1000527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zabala, A.; Vazquez-villoldo, N.; Rissiek, B.; Gejo, J.; Martin, A.; Palomino, A.; Perez-samartín, A.; Pulagam, K.R.; Lukowiak, M.; Capetillo-zarate, E.; et al. P 2 X 4 receptor controls microglia activation and favors remyelination in autoimmune encephalitis. EMBO Mol. Med. 2018, 10, 1–20. [Google Scholar] [CrossRef]
- Butovsky, O.; Ziv, Y.; Schwartz, A.; Landa, G.; Talpalar, A.E.; Pluchino, S.; Martino, G.; Schwartz, M. Microglia activated by IL-4 or IFN-γ differentially induce neurogenesis and oligodendrogenesis from adult stem/progenitor cells. Mol. Cell. Neurosci. 2006, 31, 149–160. [Google Scholar] [CrossRef]
- Lloyd, A.F.; Davies, C.L.; Miron, V.E. ScienceDirect Microglia: Origins, homeostasis, and roles in myelin repair. Curr. Opin. Neurobiol. 2017, 47, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Plemel, J.R.; Stratton, J.A.; Michaels, N.J.; Rawji, K.S.; Zhang, E.; Sinha, S.; Baaklini, C.S.; Dong, Y.; Ho, M.; Thorburn, K.; et al. Microglia response following acute demyelination is heterogeneous and limits infiltrating macrophage dispersion. Sci. Adv. 2020, 6, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Kalincik, T.; Manouchehrinia, A.; Sobisek, L.; Jokubaitis, V.; Spelman, T.; Horakova, D.; Havrdova, E.; Trojano, M.; Izquierdo, G.; Lugaresi, A.; et al. Towards personalized therapy for multiple sclerosis: Prediction of individual treatment response. Brain 2017, 140, 2426–2443. [Google Scholar] [CrossRef] [Green Version]
- Plemel, J.R.; Liu, W.; Yong, V.W. Remyelination therapies: A new direction and challenge in multiple sclerosis. Brain 2017, 140, 2426–2443. [Google Scholar] [CrossRef] [PubMed]
- Schepers, M.; Tiane, A.; Paes, D.; Sanchez, S.; Rombaut, B.; Vanmierlo, T. Targeting Phosphodiesterases—Towards a Tailor-Made Approach in Multiple Sclerosis Treatment. Front. Immunol. 2019, 10, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Beurel, E.; Kaidanovich-Beilin, O.; Yeh, W.-I.; Song, L.; Palomo, V.; Michalek, S.M.; Woodgett, J.R.; Harrington, L.E.; Eldar-Finkelman, H.; Martinez, A.; et al. Regulation of Th1 Cells and Experimental Autoimmune Encephalomyelitis by Glycogen Synthase Kinase-3. J. Immunol. 2013, 190, 5000–5011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barkhof, F. The optic nerve should be included as one of the typical CNS regions for establishing dissemination in space when diagnosing MS—Commentary. Mult. Scler. 2018, 24, 125–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tintoré, M.; Montalban, X. The optic nerve should be included as one of the typical CNS regions for establishing dissemination in space when diagnosing MS—No. Mult. Scler. 2018, 24, 123–125. [Google Scholar] [CrossRef]
- Filippi, M.; Preziosa, P.; Meani, A.; Ciccarelli, O.; Mesaros, S.; Rovira, A.; Frederiksen, J.; Enzinger, C.; Barkhof, F.; Gasperini, C.; et al. Prediction of a multiple sclerosis diagnosis in patients with clinically isolated syndrome using the 2016 MAGNIMS and 2010 McDonald criteria: A retrospective study. Lancet. Neurol. 2018, 17, 133–142. [Google Scholar] [CrossRef]
- Aktas, O.; Hartung, H.-P. CSI: Multiple sclerosis. Tracing optic nerve involvement by standardized optical coherence tomography. Ann. Neurol. 2019, 85, 615–617. [Google Scholar] [CrossRef]
- Davion, J.-B.; Lopes, R.; Drumez, É.; Labreuche, J.; Hadhoum, N.; Lannoy, J.; Vermersch, P.; Pruvo, J.-P.; Leclerc, X.; Zéphir, H.; et al. Asymptomatic optic nerve lesions. Neurology 2020, 94, 2468–2478. [Google Scholar] [CrossRef]
- Heidari, M.; Radcliff, A.B.; Mclellan, G.J.; Ver, J.N.; Chan, K.; Kiland, J.A. Evoked potentials as a biomarker of remyelination. Proc. Nat. Acad. Sci. USA 2019, 116, 27074–27083. [Google Scholar] [CrossRef]
- Moliné-Velázquez, V.; Cuervo, H.; Vila del Sol, V.; Ortega, M.C.; Clemente, D.; de Castro, F. Myeloid-Derived Suppressor Cells Limit the Inflammation by Promoting T Lymphocyte Apoptosis in the Spinal Cord of a Murine Model of Multiple Sclerosis. Brain Pathol. 2011, 21, 678–691. [Google Scholar] [CrossRef]
- Moliné-Velázquez, V.; Ortega, M.C.; Vila, V.; Melero-Jerez, C.; de Castro, F.; Clemente, D. Neurobiology of Disease The synthetic retinoid Am80 delays recovery in a model of multiple sclerosis by modulating myeloid-derived suppressor cell fate and viability. Neurobiol. Dis. 2014, 67, 149–164. [Google Scholar] [CrossRef]
- Marín-Bañasco, C.; Benabdellah, K.; Melero-Jerez, C.; Oliver, B.; Pinto-Medel, M.J.; Hurtado-Guerrero, I.; de Castro, F.; Clemente, D.; Fernández, O.; Martín, F.; et al. Gene therapy with mesenchymal stem cells expressing IFN-ß ameliorates neuroinflammation in experimental models of multiple sclerosis. Br. J. Pharmacol. 2017, 174, 238–253. [Google Scholar] [CrossRef] [PubMed]
- Melero-jerez, C.; Alonso-Gómez, A.; Moñivas, E.; Lebrón-Galán, R.; Machín-Díaz, I.; De Castro, F.; Clemente, D. Neurobiology of Disease The proportion of myeloid-derived suppressor cells in the spleen is related to the severity of the clinical course and tissue damage extent in a murine model of multiple sclerosis. Neurobiol. Dis. 2020, 140, 104869. [Google Scholar] [CrossRef]
- Torres-Platas, G.S.; Comeau, S.; Rachalski, A.; Bo, D.G.; Cruceanu, C.; Turecki, G.; Giros, B.; Mechawar, N. Morphometric characterization of microglial phenotypes in human cerebral cortex. J. Neuroinflamm. 2014, 11, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.W.; Gajavelli, S.; Spurlock, M.S.; Andreoni, C.; De Rivero Vaccari, J.P.; Bullock, M.R.; Keane, R.W.; Dietrich, W.D. Microglial Inflammasome Activation in Penetrating Ballistic-Like Brain Injury. J. Neurotrauma 2018, 35, 1681–1693. [Google Scholar] [CrossRef] [PubMed]
- Parra, I.; Martínez, I.; Ramírez-García, G.; Tizabi, Y.; Mendieta, L. Differential Effects of LPS and 6-OHDA on Microglia’s Morphology in Rats: Implications for Inflammatory Model of Parkinson’s Disease. Neurotox. Res. 2020, 37, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Leavitt, V.M.; Brandstadter, R.; Fabian, M.; Katz Sand, I.; Klineova, S.; Krieger, S.; Lewis, C.; Lublin, F.; Miller, A.; Pelle, G.; et al. Dissociable cognitive patterns related to depression and anxiety in multiple sclerosis. Mult. Scler. 2020, 26, 1247–1255. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Veh (n = 7) | VP3.15 (n = 10) | |||
|---|---|---|---|---|
| Spinal cord | Pearson’s coefficient (r) | MBP area—NFH area | 0.933 | 0.803 |
| MBP area—optic nerve width at fifth OCT | −0.212 | −0.024 | ||
| MBP area—optic nerve width at endpoint OCT | −0.413 | −0.186 | ||
| NFH area—optic nerve width at fifth OCT | −0.364 | −0.228 | ||
| NFH area—optic nerve width at endpoint OCT | −0.533 | −0.362 | ||
| Multiple correlation at fifth OCT (two-tailed multicomparison test) | 5.52246 | 3.78265 | ||
| p (fifth OCT) | <0.01 | <0.05 | ||
| Multiple correlation at endpoint OCT (two-tailed multicomparison test) | 4.87270 | 3.56740 | ||
| p (endpoint OCT) | <0.01 | <0.05 | ||
| Optic Nerve | Pearson’s coefficient (r) | MBP area—NFH area | 0.559 | 0.546 |
| MBP area—optic nerve width at fifth OCT | 0.579 | −0.385 | ||
| MBP area—optic nerve width at endpoint OCT | 0.120 | 0.222 | ||
| NFH area—optic nerve width at fifth OCT | 0.579 | 0.390 | ||
| NFH area—optic nerve width at endpoint OCT | 0.077 | 0.202 | ||
| Multiple correlation at fifth OCT (two-tailed multicomparison test) | 0.71492 | 1.39569 | ||
| p (fifth OCT) | NS | NS | ||
| Multiple correlation at endpoint OCT (two-tailed multicomparison test) | 1.33580 | 1.63117 | ||
| p (endpoint OCT) | NS | NS |
| Antibody | Target | Cellular Location | Dilution | Host Species | Class | Manufacturer | Antibody ID |
|---|---|---|---|---|---|---|---|
| MBP | Myelin | Plasma Membrane | 1:500 | Rat | Monoclonal clone 12 | Biorad | aa 82–87 |
| NFH | Neurons/ Axons | Cell Body | 1:1000 | Rabbit | Polyclonal | Abcam | Ab 8135 |
| Iba-1 | Microglia | Plasma membrane | 1:500 | Guinea pig | Polyclonal | Synaptic Systems | 234 004 |
| PDGFRα | OPCs | Plasm membrane | 1:200 | Goat | Polyclonal | RD Systems | AF 1062 |
| CC1 | Mature oligodendrocytes | Cell body | 1:200 | Mouse | Monoclonal clone CC1 | Merck Millipore | OP 80 |
| Olig2 | Oligodendrocyte lineage | nucleus | 1:200 | Rabbit | Polyclonal | Merck Millipore | AB 9610 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benítez-Fernández, R.; Melero-Jerez, C.; Gil, C.; de la Rosa, E.J.; Martínez, A.; de Castro, F. Dynamics of Central Remyelination and Treatment Evolution in a Model of Multiple Sclerosis with Optic Coherence Tomography. Int. J. Mol. Sci. 2021, 22, 2440. https://doi.org/10.3390/ijms22052440
Benítez-Fernández R, Melero-Jerez C, Gil C, de la Rosa EJ, Martínez A, de Castro F. Dynamics of Central Remyelination and Treatment Evolution in a Model of Multiple Sclerosis with Optic Coherence Tomography. International Journal of Molecular Sciences. 2021; 22(5):2440. https://doi.org/10.3390/ijms22052440
Chicago/Turabian StyleBenítez-Fernández, Rocío, Carolina Melero-Jerez, Carmen Gil, Enrique J. de la Rosa, Ana Martínez, and Fernando de Castro. 2021. "Dynamics of Central Remyelination and Treatment Evolution in a Model of Multiple Sclerosis with Optic Coherence Tomography" International Journal of Molecular Sciences 22, no. 5: 2440. https://doi.org/10.3390/ijms22052440
APA StyleBenítez-Fernández, R., Melero-Jerez, C., Gil, C., de la Rosa, E. J., Martínez, A., & de Castro, F. (2021). Dynamics of Central Remyelination and Treatment Evolution in a Model of Multiple Sclerosis with Optic Coherence Tomography. International Journal of Molecular Sciences, 22(5), 2440. https://doi.org/10.3390/ijms22052440