
Emerging Role of AP-1 Transcription Factor JunB in Angiogenesis and Vascular Development


Abstract
:1. Vascular Endothelial Cells and Activator Protein 1 (AP-1) Transcription Factors
1.1. Endothelial Cell Heterogeneities and Gene Expression
1.2. AP-1 Transcription Factors in Endothelial Cells
2. AP-1 Transcription Factor JunB in Angiogenesis
2.1. JunB Expression in Endothelial Cells
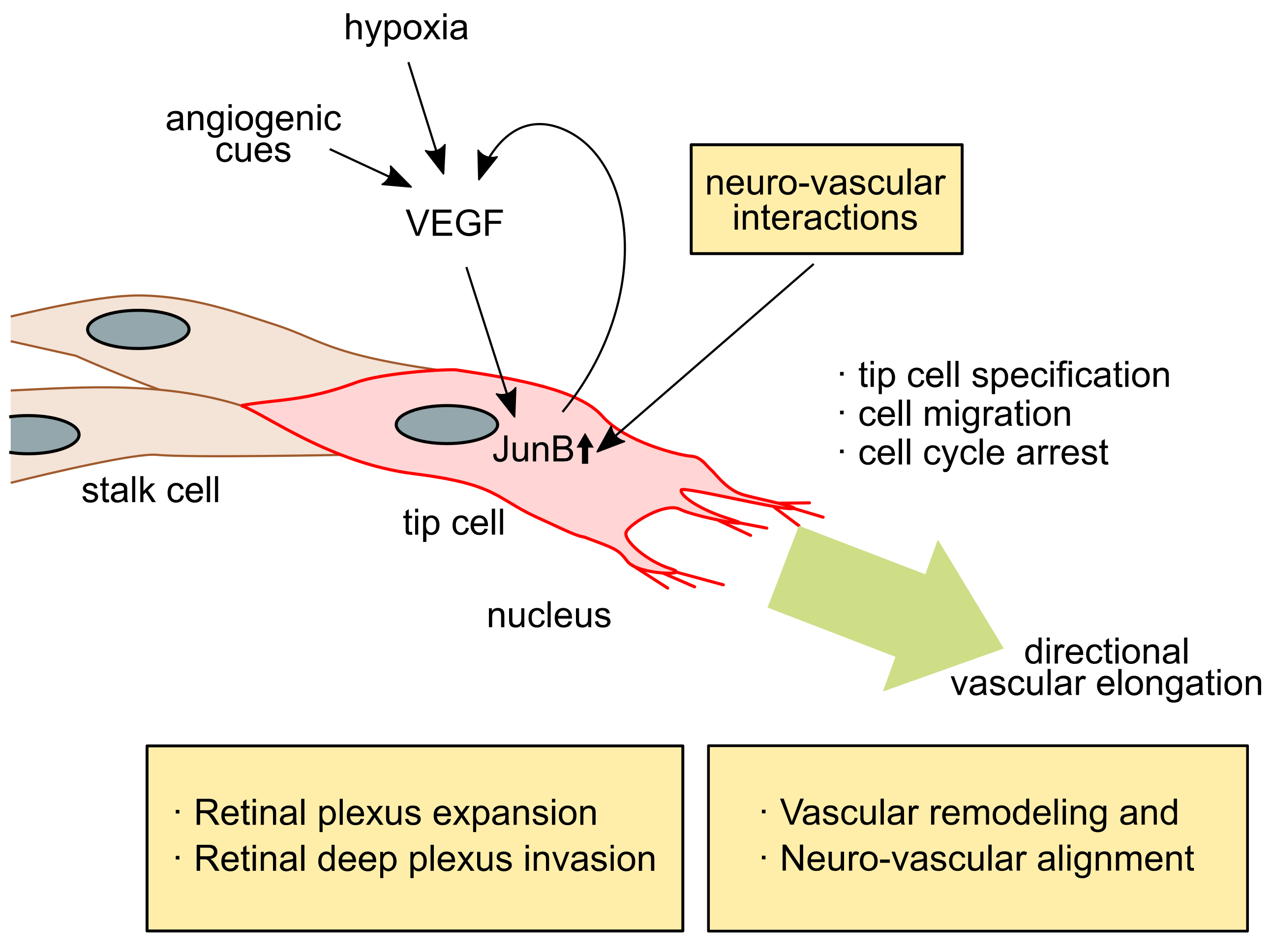
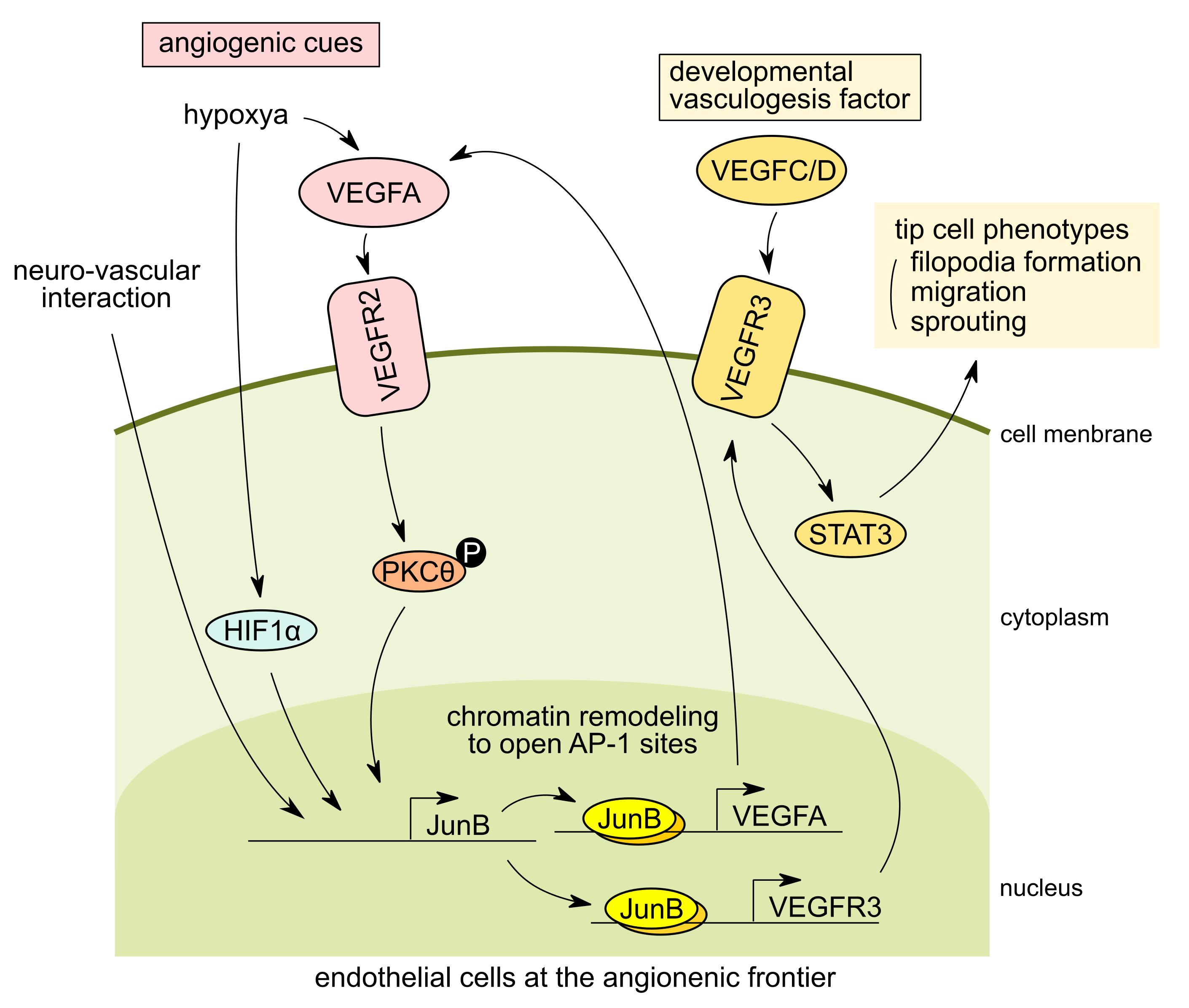
2.2. JunB Is a Tip Cell Factor in Response to VEGF Signaling
3. Endothelial JunB Functions in Vascular Development In Vivo
3.1. JunB Is Required for Placentation and Heart Vascular Development in Mice
3.2. JunB Is Involved in Retinal Vascular Outgrowth and Retinal Special Vascular Differentiation
3.3. JunB Regulates Neurovascular Parallel Alignment in Developing Skin Vasculture in Mice
3.4. JunB also Controls Lymphangiogenesis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AP-1 | Activator protein 1 |
| HMVEC | Human microvascular endothelial cells |
| S1P | Sphingosine 1-phosphate |
| VEGF | Vascular endothelial growth factor |
| VEGFR | Vascular endothelial growth factor receptor |
References
- Sabbagh, M.F.; Heng, J.S.; Luo, C.; Castanon, R.G.; Nery, J.R.; Rattner, A.; Goff, L.A.; Ecker, J.R.; Nathans, J. Transcriptional and Epigenomic Landscapes of CNS and Non-CNS Vascular Endothelial Cells. eLife 2018, 7, e36187. [Google Scholar] [CrossRef] [PubMed]
- Géraud, C.; Koch, P.-S.; Zierow, J.; Klapproth, K.; Busch, K.; Olsavszky, V.; Leibing, T.; Demory, A.; Ulbrich, F.; Diett, M.; et al. GATA4-Dependent Organ-Specific Endothelial Differentiation Controls Liver Development and Embryonic Hematopoiesis. J. Clin. Investig. 2017, 127, 1099–1114. [Google Scholar] [CrossRef]
- Nolan, D.J.; Ginsberg, M.; Israely, E.; Palikuqi, B.; Poulos, M.G.; James, D.; Ding, B.-S.; Schachterle, W.; Liu, Y.; Rosenwaks, Z.; et al. Molecular Signatures of Tissue-Specific Microvascular Endothelial Cell Heterogeneity in Organ Maintenance and Regeneration. Dev. Cell 2013, 26, 204–219. [Google Scholar] [CrossRef] [Green Version]
- Paik, D.T.; Tian, L.; Williams, I.M.; Rhee, S.; Zhang, H.; Liu, C.; Mishra, R.; Wu, S.M.; Red-Horse, K.; Wu, J.C. Single-Cell RNA Sequencing Unveils Unique Transcriptomic Signatures of Organ-Specific Endothelial Cells. Circulation 2020, 142, 1848–1862. [Google Scholar] [CrossRef]
- Kalucka, J.; de Rooij, L.P.M.H.; Goveia, J.; Rohlenova, K.; Dumas, S.J.; Meta, E.; Conchinha, N.V.; Taverna, F.; Teuwen, L.-A.; Veys, K.; et al. Single-Cell Transcriptome Atlas of Murine Endothelial Cells. Cell 2020, 180, 764–779.e20. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Taverna, F.; Rohlenova, K.; Treps, L.; Geldhof, V.; de Rooij, L.; Sokol, L.; Pircher, A.; Conradi, L.-C.; Kalucka, J.; et al. EndoDB: A Database of Endothelial Cell Transcriptomics Data. Nucleic Acids Res. 2019, 47, D736–D744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakato, R.; Wada, Y.; Nakaki, R.; Nagae, G.; Katou, Y.; Tsutsumi, S.; Nakajima, N.; Fukuhara, H.; Iguchi, A.; Kohro, T.; et al. Comprehensive Epigenome Characterization Reveals Diverse Transcriptional Regulation across Human Vascular Endothelial Cells. Epigenetics Chromatin 2019, 12, 77. [Google Scholar] [CrossRef] [Green Version]
- Gerhardt, H.; Golding, M.; Fruttiger, M.; Ruhrberg, C.; Lundkvist, A.; Abramsson, A.; Jeltsch, M.; Mitchell, C.; Alitalo, K.; Shima, D.; et al. VEGF Guides Angiogenic Sprouting Utilizing Endothelial Tip Cell Filopodia. J. Cell Biol. 2003, 161, 1163–1177. [Google Scholar] [CrossRef]
- Hellström, M.; Phng, L.-K.; Hofmann, J.J.; Wallgard, E.; Coultas, L.; Lindblom, P.; Alva, J.; Nilsson, A.-K.; Karlsson, L.; Gaiano, N.; et al. Dll4 Signalling through Notch1 Regulates Formation of Tip Cells during Angiogenesis. Nature 2007, 445, 776–780. [Google Scholar] [CrossRef]
- Zundel, W.; Schindler, C.; Haas-Kogan, D.; Koong, A.; Kaper, F.; Chen, E.; Gottschalk, A.R.; Ryan, H.E.; Johnson, R.S.; Jefferson, A.B.; et al. Loss of PTEN Facilitates HIF-1-Mediated Gene Expression. Genes Dev. 2000, 14, 391–396. [Google Scholar] [CrossRef]
- Gray, M.J.; Zhang, J.; Ellis, L.M.; Semenza, G.L.; Evans, D.B.; Watowich, S.S.; Gallick, G.E. HIF-1alpha, STAT3, CBP/P300 and Ref-1/APE Are Components of a Transcriptional Complex That Regulates Src-Dependent Hypoxia-Induced Expression of VEGF in Pancreatic and Prostate Carcinomas. Oncogene 2005, 24, 3110–3120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chintharlapalli, S.; Papineni, S.; Ramaiah, S.K.; Safe, S. Betulinic Acid Inhibits Prostate Cancer Growth through Inhibition of Specificity Protein Transcription Factors. Cancer Res. 2007, 67, 2816–2823. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Shen, G.; Chen, C.; Gélinas, C.; Kong, A.-N.T. Suppression of NF- κ B and NF- κ B-Regulated Gene Expression by Sulforaphane and PEITC through I κ B α, IKK Pathway in Human Prostate Cancer PC-3 Cells. Oncogene 2005, 24, 4486–4495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.; Jha, S.; Ivovic, A.; Fratzl-Zelman, N.; Deng, Z.; Mitra, A.; Cabral, W.A.; Hanson, E.P.; Lange, E.; Cowen, E.W.; et al. Somatic SMAD3-Activating Mutations Cause Melorheostosis by up-Regulating the TGF-β/SMAD Pathway. J. Exp. Med. 2020, 217. [Google Scholar] [CrossRef]
- Hattori, T.; Müller, C.; Gebhard, S.; Bauer, E.; Pausch, F.; Schlund, B.; Bösl, M.R.; Hess, A.; Surmann-Schmitt, C.; von der Mark, H.; et al. SOX9 Is a Major Negative Regulator of Cartilage Vascularization, Bone Marrow Formation and Endochondral Ossification. Dev. Camb. Engl. 2010, 137, 901–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renault, V.M.; Rafalski, V.A.; Morgan, A.A.; Salih, D.A.M.; Brett, J.O.; Webb, A.E.; Villeda, S.A.; Thekkat, P.U.; Guillerey, C.; Denko, N.C.; et al. FoxO3 Regulates Neural Stem Cell Homeostasis. Cell Stem Cell 2009, 5, 527–539. [Google Scholar] [CrossRef] [Green Version]
- Potente, M.; Urbich, C.; Sasaki, K.; Hofmann, W.K.; Heeschen, C.; Aicher, A.; Kollipara, R.; DePinho, R.A.; Zeiher, A.M.; Dimmeler, S. Involvement of Foxo Transcription Factors in Angiogenesis and Postnatal Neovascularization. J. Clin. Investig. 2005, 115, 2382–2392. [Google Scholar] [CrossRef] [Green Version]
- Balic, J.J.; Garama, D.J.; Saad, M.I.; Yu, L.; West, A.C.; West, A.J.; Livis, T.; Bhathal, P.S.; Gough, D.J.; Jenkins, B.J. Serine-Phosphorylated STAT3 Promotes Tumorigenesis via Modulation of RNA Polymerase Transcriptional Activity. Cancer Res. 2019, 79, 5272–5287. [Google Scholar] [CrossRef] [Green Version]
- Kostromina, E.; Gustavsson, N.; Wang, X.; Lim, C.-Y.; Radda, G.K.; Li, C.; Han, W. Glucose Intolerance and Impaired Insulin Secretion in Pancreas-Specific Signal Transducer and Activator of Transcription-3 Knockout Mice Are Associated with Microvascular Alterations in the Pancreas. Endocrinology 2010, 151, 2050–2059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhee, S.H.; Ma, E.L.; Lee, Y.; Taché, Y.; Pothoulakis, C.; Im, E. Corticotropin Releasing Hormone and Urocortin 3 Stimulate Vascular Endothelial Growth Factor Expression through the CAMP/CREB Pathway. J. Biol. Chem. 2015, 290, 26194–26203. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Zhou, J.; Liu, D.; Dong, F.; Cheng, H.; Wang, W.; Pang, Y.; Wang, Y.; Mu, X.; Ni, Y.; et al. ATF4 Plays a Pivotal Role in the Development of Functional Hematopoietic Stem Cells in Mouse Fetal Liver. Blood 2015, 126, 2383–2391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.S.; Qiao, L.; Su, Z.Z.; Hinman, D.; Willoughby, K.; McKinstry, R.; Yacoub, A.; Duigou, G.J.; Young, C.S.; Grant, S.; et al. Ionizing Radiation Modulates Vascular Endothelial Growth Factor (VEGF) Expression through Multiple Mitogen Activated Protein Kinase Dependent Pathways. Oncogene 2001, 20, 3266–3280. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.L.; Tee, M.K.; Goetzl, E.J.; Auersperg, N.; Mills, G.B.; Ferrara, N.; Jaffe, R.B. Lysophosphatidic Acid Induction of Vascular Endothelial Growth Factor Expression in Human Ovarian Cancer Cells. J. Natl. Cancer Inst. 2001, 93, 762–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, J.; Li, J.; Chen, J.; Chen, H.; Ouyang, W.; Zhang, R.; Xue, C.; Zhang, D.; Amin, S.; Desai, D.; et al. Effects of Polycyclic Aromatic Hydrocarbons (PAHs) on Vascular Endothelial Growth Factor Induction through Phosphatidylinositol 3-Kinase/AP-1-Dependent, HIF-1alpha-Independent Pathway. J. Biol. Chem. 2006, 281, 9093–9100. [Google Scholar] [CrossRef] [Green Version]
- Monje, P.; Hernández-Losa, J.; Lyons, R.J.; Castellone, M.D.; Gutkind, J.S. Regulation of the Transcriptional Activity of C-Fos by ERK. A Novel Role for the Prolyl Isomerase PIN1. J. Biol. Chem. 2005, 280, 35081–35084. [Google Scholar] [CrossRef] [Green Version]
- Yoshitomi, Y.; Ikeda, T.; Saito, H.; Yoshitake, Y.; Ishigaki, Y.; Hatta, T.; Kato, N.; Yonekura, H. JunB Regulates Angiogenesis and Neurovascular Parallel Alignment in Mouse Embryonic Skin. J. Cell Sci. 2017, 130, 916. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Cao, L.; Zhang, X.; Zhuang, Y.; Zhang, Y.; Wang, Q.; Chen, Y.; Xu, L.; Sun, G. MiR-3133 Inhibits Proliferation and Angiogenesis by Targeting the JUNB/VEGF Pathway in Human Umbilical Vein Endothelial Cells. Oncol. Rep. 2020, 44, 1699–1708. [Google Scholar] [CrossRef]
- Gerald, D.; Berra, E.; Frapart, Y.M.; Chan, D.A.; Giaccia, A.J.; Mansuy, D.; Pouysségur, J.; Yaniv, M.; Mechta-Grigoriou, F. JunD Reduces Tumor Angiogenesis by Protecting Cells from Oxidative Stress. Cell 2004, 118, 781–794. [Google Scholar] [CrossRef]
- Angel, P.; Karin, M. The Role of Jun, Fos and the AP-1 Complex in Cell-Proliferation and Transformation. Biochim. Biophys. Acta BBA Rev. Cancer 1991, 1072, 129–157. [Google Scholar] [CrossRef]
- Eferl, R.; Wagner, E.F. AP-1: A Double-Edged Sword in Tumorigenesis. Nat. Rev. Cancer 2003, 3, 859–868. [Google Scholar] [CrossRef]
- Hess, J.; Angel, P.; Schorpp-Kistner, M. AP-1 Subunits: Quarrel and Harmony among Siblings. J. Cell Sci. 2004, 117, 5965–5973. [Google Scholar] [CrossRef] [Green Version]
- Hai, T.W.; Liu, F.; Coukos, W.J.; Green, M.R. Transcription Factor ATF CDNA Clones: An Extensive Family of Leucine Zipper Proteins Able to Selectively Form DNA-Binding Heterodimers. Genes Dev. 1989, 3, 2083–2090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, J.; Ye, T.; Cui, P.; Hua, Q.; Zeng, H.; Zhao, D. AP-1 Transcription Factor Mediates VEGF-Induced Endothelial Cell Migration and Proliferation. Microvasc. Res. 2016, 105, 103–108. [Google Scholar] [CrossRef] [Green Version]
- Chien, M.-H.; Ku, C.-C.; Johansson, G.; Chen, M.-W.; Hsiao, M.; Su, J.-L.; Inoue, H.; Hua, K.-T.; Wei, L.-H.; Kuo, M.-L. Vascular Endothelial Growth Factor-C (VEGF-C) Promotes Angiogenesis by Induction of COX-2 in Leukemic Cells via the VEGF-R3/JNK/AP-1 Pathway. Carcinogenesis 2009, 30, 2005–2013. [Google Scholar] [CrossRef] [Green Version]
- Shih, S.C.; Claffey, K.P. Role of AP-1 and HIF-1 Transcription Factors in TGF-Beta Activation of VEGF Expression. Growth Factors Chur Switz. 2001, 19, 19–34. [Google Scholar] [CrossRef]
- Wang, S.; Lu, J.; You, Q.; Huang, H.; Chen, Y.; Liu, K. The MTOR/AP-1/VEGF Signaling Pathway Regulates Vascular Endothelial Cell Growth. Oncotarget 2016, 7, 53269–53276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolch, W.; Martiny-Baron, G.; Kieser, A.; Marmé, D. Regulation of the Expression of the VEGF/VPS and Its Receptors: Role in Tumor Angiogenesis. Breast Cancer Res. Treat. 1995, 36, 139–155. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.K.; Quyen, D.V.; Kundumani-Sridharan, V.; Brooks, P.C.; Rao, G.N. AP-1 (Fra-1/c-Jun)-Mediated Induction of Expression of Matrix Metalloproteinase-2 Is Required for 15S-Hydroxyeicosatetraenoic Acid-Induced Angiogenesis. J. Biol. Chem. 2010, 285, 16830–16843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galvagni, F.; Orlandini, M.; Oliviero, S. Role of the AP-1 Transcription Factor FOSL1 in Endothelial Cells Adhesion and Migration. Cell Adhes. Migr. 2013, 7, 408–411. [Google Scholar] [CrossRef] [Green Version]
- Marconcini, L.; Marchio, S.; Morbidelli, L.; Cartocci, E.; Albini, A.; Ziche, M.; Bussolino, F.; Oliviero, S. C-Fos-Induced Growth Factor/Vascular Endothelial Growth Factor D Induces Angiogenesis in Vivo and in Vitro. Proc. Natl. Acad. Sci. USA 1999, 96, 9671–9676. [Google Scholar] [CrossRef] [Green Version]
- Toft, D.J.; Rosenberg, S.B.; Bergers, G.; Volpert, O.; Linzer, D.I. Reactivation of Proliferin Gene Expression Is Associated with Increased Angiogenesis in a Cell Culture Model of Fibrosarcoma Tumor Progression. Proc. Natl. Acad. Sci. USA 2001, 98, 13055–13059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Licht, A.H.; Pein, O.T.; Florin, L.; Hartenstein, B.; Reuter, H.; Arnold, B.; Lichter, P.; Angel, P.; Schorpp-Kistner, M. JunB Is Required for Endothelial Cell Morphogenesis by Regulating Core-Binding Factor Beta. J. Cell Biol. 2006, 175, 981–991. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Dass, C.R.; Sumithran, E.; Di Girolamo, N.; Sun, L.-Q.; Khachigian, L.M. Effect of Deoxyribozymes Targeting C-Jun on Solid Tumor Growth and Angiogenesis in Rodents. J. Natl. Cancer Inst. 2004, 96, 683–696. [Google Scholar] [CrossRef] [Green Version]
- Jacobs-Helber, S.M.; Abutin, R.M.; Tian, C.; Bondurant, M.; Wickrema, A.; Sawyer, S.T. Role of JunB in Erythroid Differentiation. J. Biol. Chem. 2002, 277, 4859–4866. [Google Scholar] [CrossRef] [Green Version]
- Katagiri, T.; Kameda, H.; Nakano, H.; Yamazaki, S. Regulation of T Cell Differentiation by the AP-1 Transcription Factor JunB. Immunol. Med. 2021, 1–12. [Google Scholar] [CrossRef]
- Nakamura, M.; Yoshida, H.; Takahashi, E.; Wlizla, M.; Takebayashi-Suzuki, K.; Horb, M.E.; Suzuki, A. The AP-1 Transcription Factor JunB Functions in Xenopus Tail Regeneration by Positively Regulating Cell Proliferation. Biochem. Biophys. Res. Commun. 2020, 522, 990–995. [Google Scholar] [CrossRef]
- Andrecht, S.; Kolbus, A.; Hartenstein, B.; Angel, P.; Schorpp-Kistner, M. Cell Cycle Promoting Activity of JunB through Cyclin A Activation. J. Biol. Chem. 2002, 277, 35961–35968. [Google Scholar] [CrossRef] [Green Version]
- Bakiri, L.; Lallemand, D.; Bossy-Wetzel, E.; Yaniv, M. Cell Cycle-Dependent Variations in c-Jun and JunB Phosphorylation: A Role in the Control of Cyclin D1 Expression. EMBO J. 2000, 19, 2056–2068. [Google Scholar] [CrossRef]
- Nuzzo, A.M.; Giuffrida, D.; Zenerino, C.; Piazzese, A.; Olearo, E.; Todros, T.; Rolfo, A. JunB/Cyclin-D1 Imbalance in Placental Mesenchymal Stromal Cells Derived from Preeclamptic Pregnancies with Fetal-Placental Compromise. Placenta 2014, 35, 483–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, X.; Orchard, G.; Lillington, D.M.; Russell-Jones, R.; Young, B.D.; Whittaker, S.J. Amplification and Overexpression of JUNB Is Associated with Primary Cutaneous T-Cell Lymphomas. Blood 2003, 101, 1513–1519. [Google Scholar] [CrossRef] [Green Version]
- Carr, T.M.; Wheaton, J.D.; Houtz, G.M.; Ciofani, M. JunB Promotes Th17 Cell Identity and Restrains Alternative CD4 + T-Cell Programs during Inflammation. Nat. Commun. 2017, 8, 301. [Google Scholar] [CrossRef] [Green Version]
- Hasan, Z.; Koizumi, S.; Sasaki, D.; Yamada, H.; Arakaki, N.; Fujihara, Y.; Okitsu, S.; Shirahata, H.; Ishikawa, H. JunB Is Essential for IL-23-Dependent Pathogenicity of Th17 Cells. Nat. Commun. 2017, 8, 15628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamazaki, S.; Tanaka, Y.; Araki, H.; Kohda, A.; Sanematsu, F.; Arasaki, T.; Duan, X.; Miura, F.; Katagiri, T.; Shindo, R.; et al. The AP-1 Transcription Factor JunB Is Required for Th17 Cell Differentiation. Sci. Rep. 2017, 7, 17402. [Google Scholar] [CrossRef] [Green Version]
- Hoshino, K.; Quintás-Cardama, A.; Radich, J.; Dai, H.; Yang, H.; Garcia-Manero, G. Downregulation of JUNB MRNA Expression in Advanced Phase Chronic Myelogenous Leukemia. Leuk. Res. 2009, 33, 1361–1366. [Google Scholar] [CrossRef] [Green Version]
- Gong, C.; Shen, J.; Fang, Z.; Qiao, L.; Feng, R.; Lin, X.; Li, S. Abnormally Expressed JunB Transactivated by IL-6/STAT3 Signaling Promotes Uveal Melanoma Aggressiveness via Epithelial–Mesenchymal Transition. Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef] [Green Version]
- Zenz, R.; Eferl, R.; Kenner, L.; Florin, L.; Hummerich, L.; Mehic, D.; Scheuch, H.; Angel, P.; Tschachler, E.; Wagner, E.F. Psoriasis-like Skin Disease and Arthritis Caused by Inducible Epidermal Deletion of Jun Proteins. Nature 2005, 437, 369–375. [Google Scholar] [CrossRef]
- Kobayashi, S.; Nagafuchi, Y.; Okubo, M.; Sugimori, Y.; Shirai, H.; Hatano, H.; Junko, M.; Yanaoka, H.; Takeshima, Y.; Ota, M.; et al. Integrated Bulk and Single-Cell RNA-Sequencing Identified Disease-Relevant Monocytes and a Gene Network Module Underlying Systemic Sclerosis. J. Autoimmun. 2021, 116, 102547. [Google Scholar] [CrossRef]
- Ponticos, M.; Papaioannou, I.; Xu, S.; Holmes, A.M.; Khan, K.; Denton, C.P.; Bou-Gharios, G.; Abraham, D.J. Failed Degradation of JunB Contributes to Overproduction of Type I Collagen and Development of Dermal Fibrosis in Patients With Systemic Sclerosis. Arthritis Rheumatol. 2015, 67, 243–253. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, D.; Textor, B.; Pein, O.T.; Licht, A.H.; Andrecht, S.; Sator-Schmitt, M.; Fusenig, N.E.; Angel, P.; Schorpp-Kistner, M. Critical Role for NF-KappaB-Induced JunB in VEGF Regulation and Tumor Angiogenesis. EMBO J. 2007, 26, 710–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Tournier, C.; Davis, R.J.; Flavell, R.A. Regulation of IL-4 Expression by the Transcription Factor JunB during T Helper Cell Differentiation. EMBO J. 1999, 18, 420–432. [Google Scholar] [CrossRef] [Green Version]
- He, H.; Luo, Y. Brg1 Regulates the Transcription of Human Papillomavirus Type 18 E6 and E7 Genes. Cell Cycle 2012, 11, 617–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henderson, A.; Holloway, A.; Reeves, R.; Tremethick, D.J. Recruitment of SWI/SNF to the Human Immunodeficiency Virus Type 1 Promoter. Mol. Cell. Biol. 2004, 24, 389–397. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Yamauchi, M.; Nishina, M.; Yamamichi, N.; Mizutani, T.; Ui, M.; Murakami, M.; Iba, H. Identification of SWI·SNF Complex Subunit BAF60a as a Determinant of the Transactivation Potential of Fos/Jun Dimers. J. Biol. Chem. 2001, 276, 2852–2857. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.-F.; Li, R. JunB Potentiates Function of BRCA1 Activation Domain 1 (AD1) through a Coiled-Coil-Mediated Interaction. Genes Dev. 2002, 16, 1509–1517. [Google Scholar] [CrossRef] [Green Version]
- Moynahan, M.E.; Chiu, J.W.; Koller, B.H.; Jasin, M. Brca1 Controls Homology-Directed DNA Repair. Mol. Cell 1999, 4, 511–518. [Google Scholar] [CrossRef]
- Scully, R.; Chen, J.; Plug, A.; Xiao, Y.; Weaver, D.; Feunteun, J.; Ashley, T.; Livingston, D.M. Association of BRCA1 with Rad51 in Mitotic and Meiotic Cells. Cell 1997, 88, 265–275. [Google Scholar] [CrossRef] [Green Version]
- Bochar, D.A.; Wang, L.; Beniya, H.; Kinev, A.; Xue, Y.; Lane, W.S.; Wang, W.; Kashanchi, F.; Shiekhattar, R. BRCA1 Is Associated with a Human SWI/SNF-Related Complex: Linking Chromatin Remodeling to Breast Cancer. Cell 2000, 102, 257–265. [Google Scholar] [CrossRef] [Green Version]
- Yanagida, K.; Engelbrecht, E.; Niaudet, C.; Jung, B.; Gaengel, K.; Holton, K.; Swendeman, S.; Liu, C.H.; Levesque, M.V.; Kuo, A.; et al. Sphingosine 1-Phosphate Receptor Signaling Establishes AP-1 Gradients to Allow for Retinal Endothelial Cell Specialization. Dev. Cell 2020, 52, 779–793.e7. [Google Scholar] [CrossRef]
- Carmeliet, P.; Tessier-Lavigne, M. Common Mechanisms of Nerve and Blood Vessel Wiring. Nature 2005, 436, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Schorpp-Kistner, M.; Wang, Z.Q.; Angel, P.; Wagner, E.F. JunB Is Essential for Mammalian Placentation. EMBO J. 1999, 18, 934–948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Covassin, L.D.; Villefranc, J.A.; Kacergis, M.C.; Weinstein, B.M.; Lawson, N.D. Distinct Genetic Interactions between Multiple Vegf Receptors Are Required for Development of Different Blood Vessel Types in Zebrafish. Proc. Natl. Acad. Sci. USA 2006, 103, 6554–6559. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, I.; Bahram, F.; Li, X.; Gualandi, L.; Koch, S.; Jarvius, M.; Söderberg, O.; Anisimov, A.; Kholová, I.; Pytowski, B.; et al. VEGF Receptor 2/-3 Heterodimers Detected in Situ by Proximity Ligation on Angiogenic Sprouts. EMBO J. 2010, 29, 1377–1388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benedito, R.; Rocha, S.F.; Woeste, M.; Zamykal, M.; Radtke, F.; Casanovas, O.; Duarte, A.; Pytowski, B.; Adams, R.H. Notch-Dependent VEGFR3 Upregulation Allows Angiogenesis without VEGF-VEGFR2 Signalling. Nature 2012, 484, 110–114. [Google Scholar] [CrossRef]
- Achen, M.G.; Jeltsch, M.; Kukk, E.; Mäkinen, T.; Vitali, A.; Wilks, A.F.; Alitalo, K.; Stacker, S.A. Vascular Endothelial Growth Factor D (VEGF-D) Is a Ligand for the Tyrosine Kinases VEGF Receptor 2 (Flk1) and VEGF Receptor 3 (Flt4). Proc. Natl. Acad. Sci. USA 1998, 95, 548–553. [Google Scholar] [CrossRef] [Green Version]
- Joukov, V.; Pajusola, K.; Kaipainen, A.; Chilov, D.; Lahtinen, I.; Kukk, E.; Saksela, O.; Kalkkinen, N.; Alitalo, K. A Novel Vascular Endothelial Growth Factor, VEGF-C, Is a Ligand for the Flt4 (VEGFR-3) and KDR (VEGFR-2) Receptor Tyrosine Kinases. EMBO J. 1996, 15, 290–298. [Google Scholar] [CrossRef]
- Dumont, D.J.; Jussila, L.; Taipale, J.; Lymboussaki, A.; Mustonen, T.; Pajusola, K.; Breitman, M.; Alitalo, K. Cardiovascular Failure in Mouse Embryos Deficient in VEGF Receptor-3. Science 1998, 282, 946–949. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Mani, A.M.; Singh, N.K.; Rao, G.N. PKCθ-JunB Axis via Upregulation of VEGFR3 Expression Mediates Hypoxia-Induced Pathological Retinal Neovascularization. Cell Death Dis. 2020, 11, 325. [Google Scholar] [CrossRef]
- Mukouyama, Y.; Shin, D.; Britsch, S.; Taniguchi, M.; Anderson, D.J. Sensory Nerves Determine the Pattern of Arterial Differentiation and Blood Vessel Branching in the Skin. Cell 2002, 109, 693–705. [Google Scholar] [CrossRef] [Green Version]
- Saito, D.; Takase, Y.; Murai, H.; Takahashi, Y. The Dorsal Aorta Initiates a Molecular Cascade That Instructs Sympatho-Adrenal Specification. Science 2012, 336, 1578–1581. [Google Scholar] [CrossRef]
- Kwon, H.-B.; Fukuhara, S.; Asakawa, K.; Ando, K.; Kashiwada, T.; Kawakami, K.; Hibi, M.; Kwon, Y.-G.; Kim, K.-W.; Alitalo, K.; et al. The Parallel Growth of Motoneuron Axons with the Dorsal Aorta Depends on Vegfc/Vegfr3 Signaling in Zebrafish. Development 2013, 140, 4081–4090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Kohara, H.; Uchida, Y.; James, J.M.; Soneji, K.; Cronshaw, D.G.; Zou, Y.-R.; Nagasawa, T.; Mukouyama, Y. Peripheral Nerve-Derived CXCL12 and VEGF-A Regulate the Patterning of Arterial Vessel Branching in Developing Limb Skin. Dev. Cell 2013, 24, 359–371. [Google Scholar] [CrossRef] [Green Version]
- Wigle, J.T.; Oliver, G. Prox1 Function Is Required for the Development of the Murine Lymphatic System. Cell 1999, 98, 769–778. [Google Scholar] [CrossRef] [Green Version]
- Wigle, J.T.; Harvey, N.; Detmar, M.; Lagutina, I.; Grosveld, G.; Gunn, M.D.; Jackson, D.G.; Oliver, G. An Essential Role for Prox1 in the Induction of the Lymphatic Endothelial Cell Phenotype. EMBO J. 2002, 21, 1505–1513. [Google Scholar] [CrossRef]
- Niimi, K.; Kohara, M.; Sedoh, E.; Fukumoto, M.; Shibata, S.; Sawano, T.; Tashiro, F.; Miyazaki, S.; Kubota, Y.; Miyazaki, J.-I.; et al. FOXO1 Regulates Developmental Lymphangiogenesis by Upregulating CXCR4 in the Mouse-Tail Dermis. Dev. Camb. Engl. 2020, 147. [Google Scholar] [CrossRef]
- Kiesow, K.; Bennewitz, K.; Miranda, L.G.; Stoll, S.J.; Hartenstein, B.; Angel, P.; Kroll, J.; Schorpp-Kistner, M. Junb Controls Lymphatic Vascular Development in Zebrafish via MiR-182. Sci. Rep. 2015, 5, 15007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furuyama, T.; Kitayama, K.; Shimoda, Y.; Ogawa, M.; Sone, K.; Yoshida-Araki, K.; Hisatsune, H.; Nishikawa, S.; Nakayama, K.; Nakayama, K.; et al. Abnormal Angiogenesis in Foxo1 (Fkhr)-Deficient Mice. J. Biol. Chem. 2004, 279, 34741–34749. [Google Scholar] [CrossRef] [Green Version]
- Sengupta, A.; Chakraborty, S.; Paik, J.; Yutzey, K.E.; Evans-Anderson, H.J. FoxO1 Is Required in Endothelial but Not Myocardial Cell Lineages during Cardiovascular Development. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2012, 241, 803–813. [Google Scholar] [CrossRef] [PubMed]
- Mäkinen, T.; Veikkola, T.; Mustjoki, S.; Karpanen, T.; Catimel, B.; Nice, E.C.; Wise, L.; Mercer, A.; Kowalski, H.; Kerjaschki, D.; et al. Isolated Lymphatic Endothelial Cells Transduce Growth, Survival and Migratory Signals via the VEGF-C/D Receptor VEGFR-3. EMBO J. 2001, 20, 4762–4773. [Google Scholar] [CrossRef] [Green Version]
- Salameh, A.; Galvagni, F.; Bardelli, M.; Bussolino, F.; Oliviero, S. Direct Recruitment of CRK and GRB2 to VEGFR-3 Induces Proliferation, Migration, and Survival of Endothelial Cells through the Activation of ERK, AKT, and JNK Pathways. Blood 2005, 106, 3423–3431. [Google Scholar] [CrossRef]
- Adams, R.H.; Wilkinson, G.A.; Weiss, C.; Diella, F.; Gale, N.W.; Deutsch, U.; Risau, W.; Klein, R. Roles of EphrinB Ligands and EphB Receptors in Cardiovascular Development: Demarcation of Arterial/Venous Domains, Vascular Morphogenesis, and Sprouting Angiogenesis. Genes Dev. 1999, 13, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Gerety, S.S.; Wang, H.U.; Chen, Z.F.; Anderson, D.J. Symmetrical Mutant Phenotypes of the Receptor EphB4 and Its Specific Transmembrane Ligand Ephrin-B2 in Cardiovascular Development. Mol. Cell 1999, 4, 403–414. [Google Scholar] [CrossRef]
- Kidoya, H.; Naito, H.; Muramatsu, F.; Yamakawa, D.; Jia, W.; Ikawa, M.; Sonobe, T.; Tsuchimochi, H.; Shirai, M.; Adams, R.H.; et al. APJ Regulates Parallel Alignment of Arteries and Veins in the Skin. Dev. Cell 2015, 33, 247–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| JunB Functions in Endothelial Cells | Reference |
|---|---|
| Angiogenesis | [26,27,42,59,68,70,77] |
| Neurovascular parallel alignment | [26] |
| Filopodia formation and tip cell specification | [26,68,77] |
| Retinal vascular development | [68,77] |
| Lymphangiogenesis | [85] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoshitomi, Y.; Ikeda, T.; Saito-Takatsuji, H.; Yonekura, H. Emerging Role of AP-1 Transcription Factor JunB in Angiogenesis and Vascular Development. Int. J. Mol. Sci. 2021, 22, 2804. https://doi.org/10.3390/ijms22062804
Yoshitomi Y, Ikeda T, Saito-Takatsuji H, Yonekura H. Emerging Role of AP-1 Transcription Factor JunB in Angiogenesis and Vascular Development. International Journal of Molecular Sciences. 2021; 22(6):2804. https://doi.org/10.3390/ijms22062804
Chicago/Turabian StyleYoshitomi, Yasuo, Takayuki Ikeda, Hidehito Saito-Takatsuji, and Hideto Yonekura. 2021. "Emerging Role of AP-1 Transcription Factor JunB in Angiogenesis and Vascular Development" International Journal of Molecular Sciences 22, no. 6: 2804. https://doi.org/10.3390/ijms22062804
APA StyleYoshitomi, Y., Ikeda, T., Saito-Takatsuji, H., & Yonekura, H. (2021). Emerging Role of AP-1 Transcription Factor JunB in Angiogenesis and Vascular Development. International Journal of Molecular Sciences, 22(6), 2804. https://doi.org/10.3390/ijms22062804