
The Serine Biosynthesis of Paenibacillus polymyxa WLY78 Is Regulated by the T-Box Riboswitch


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
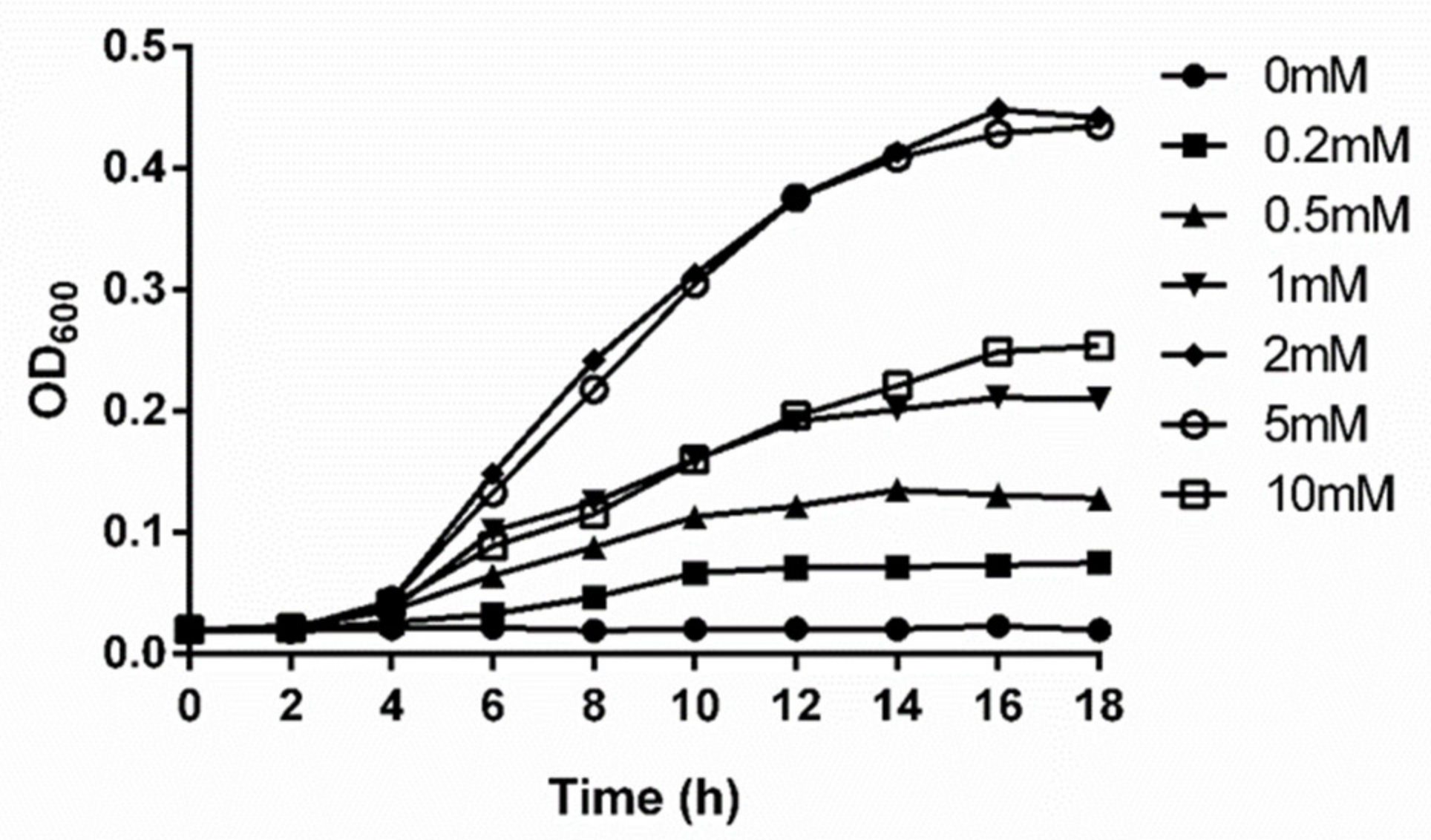
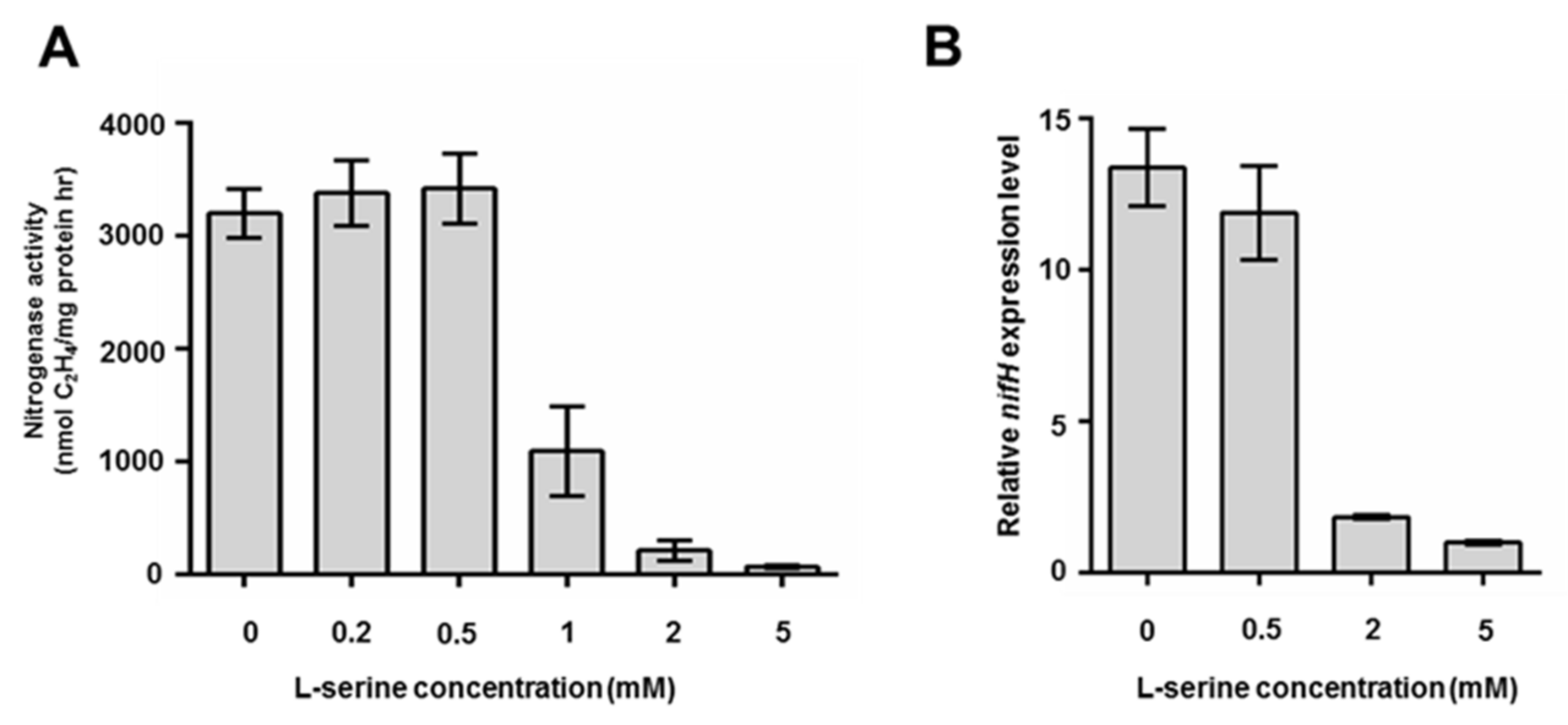
2.1. Impacts of Serine Concentration on Growth, Nitrogenase Activity, and nifH Expression of P. polymyxa WLY78
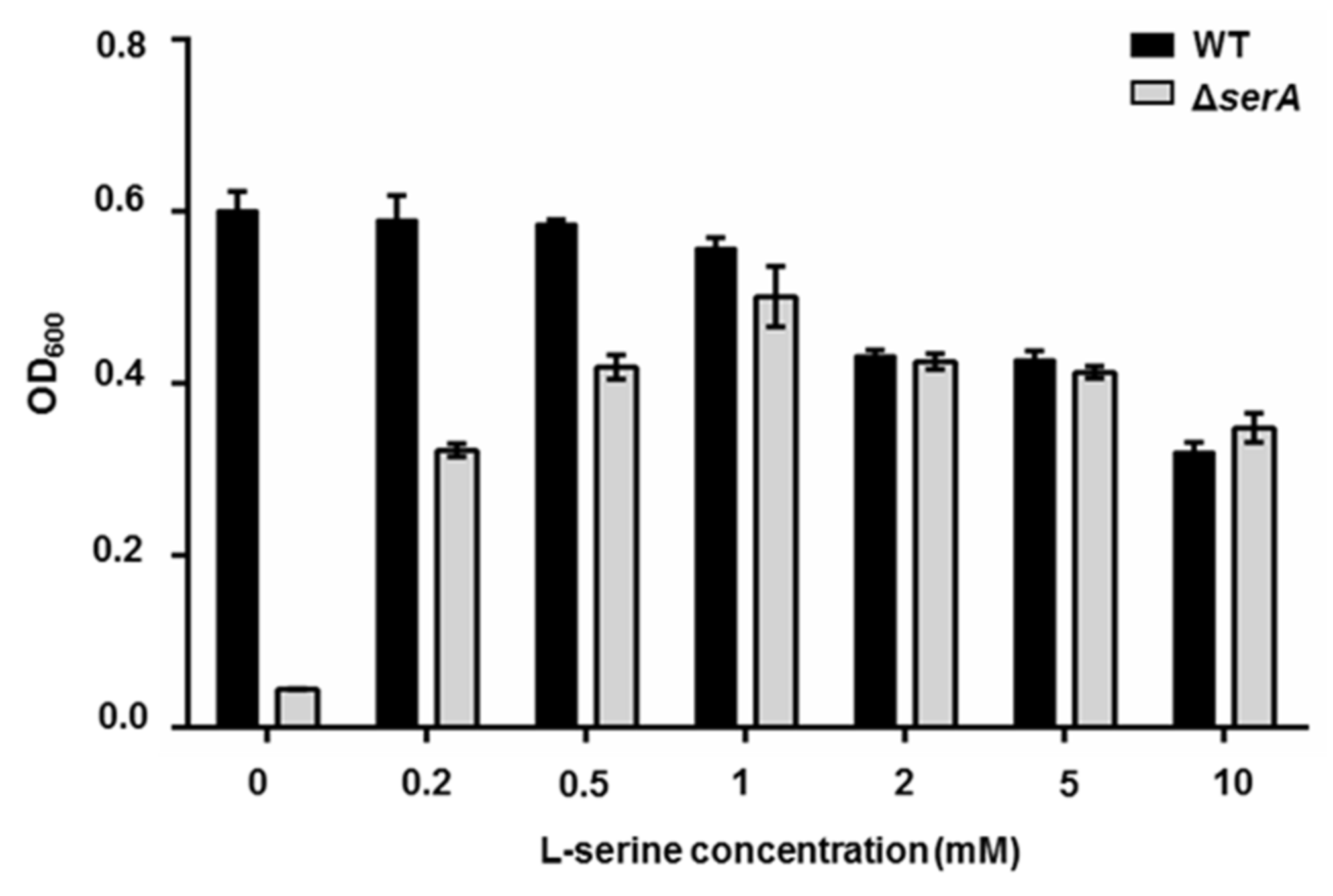
2.2. The serA Gene Is Essential for Growth of P. polymyxa WLY78 in Absence of Serine
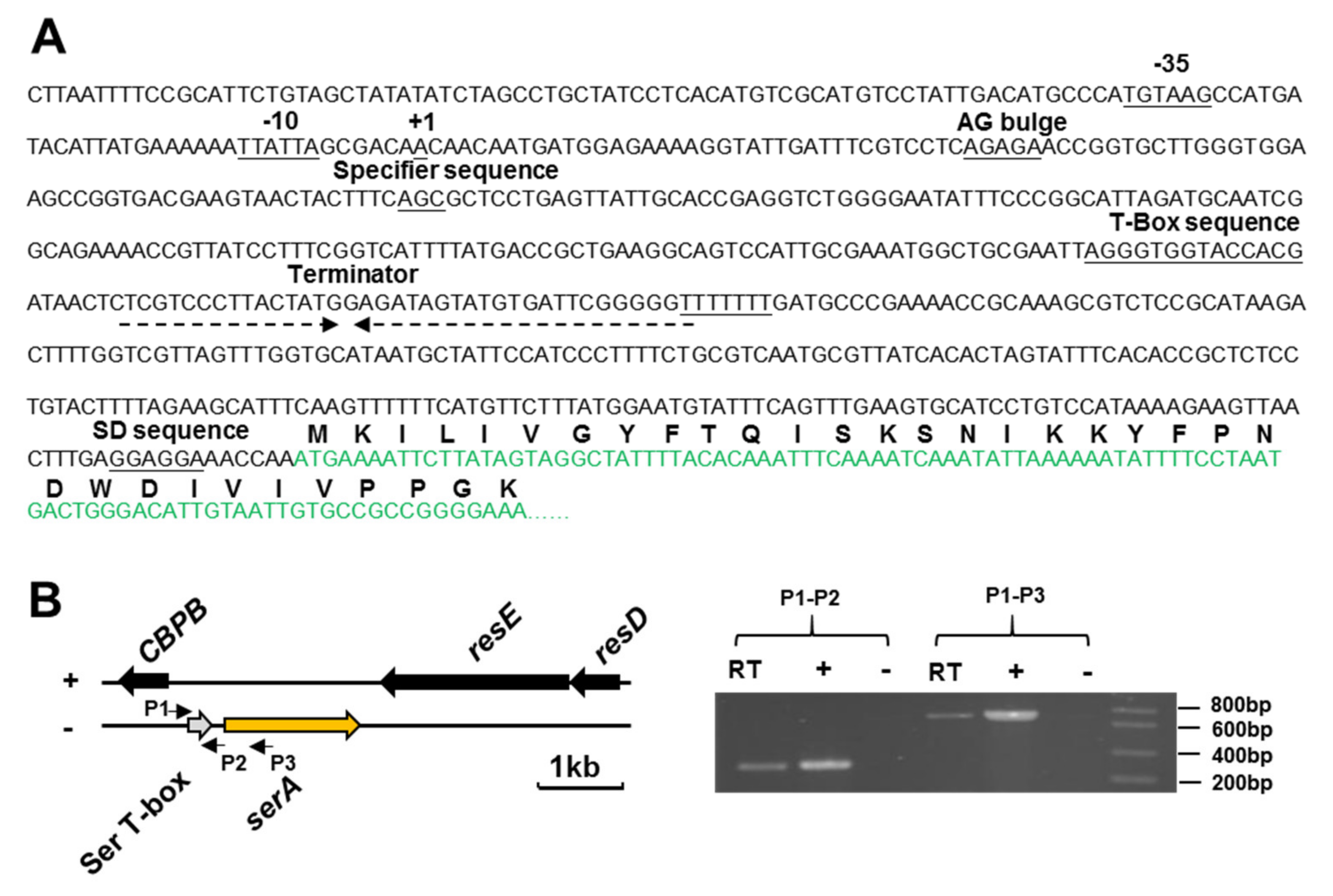
2.3. Characterization of the T-box Riboswitch of the serA Gene
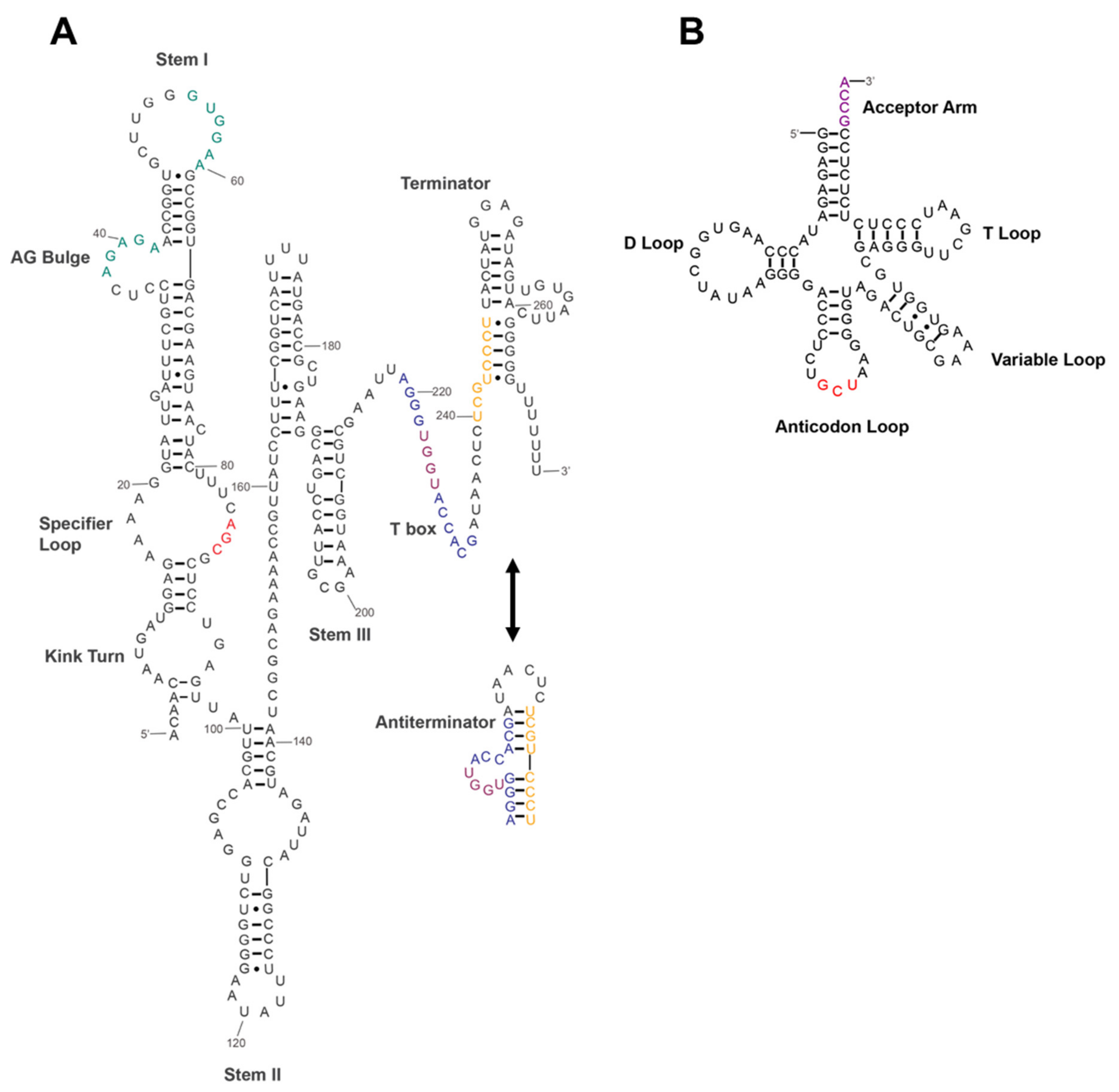
2.4. The Secondary Structure of Ser T-box RNA
2.5. Identification of the Specifier Codon for Ser in the T-box Riboswitch
2.6. Mutations of the Conserved Motifs in T-box RNA Affect the serA Expression
3. Discussion
4. Materials and Methods
4.1. Bacteria Strains, Plasmids, and Growth Conditions
4.2. Construction of Ser T-boxΔ216, Ser T-boxΔ70, Ser T-boxΔ280, and ΔserA Strains
4.3. Construction of Ser T-boxΔ70-BamHI Strain
4.4. Construction of Promoters-lacZ Fusion
4.5. RNA Preparation, RT-PCR, and qRT-PCR Analysis
4.6. Acetylene Reduction Assays of Nitrogenase Activity
4.7. Identification of Transcription Start Site
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Winkler, W.; Nahvi, A.; Breaker, R.R. Thiamine derivatives bind messenger RNAs directly to regulate bacterial gene ex-pression. Nature 2002, 419, 952–956. [Google Scholar] [CrossRef] [PubMed]
- Mandal, M.; Boese, B.; E Barrick, J.; Winkler, W.C.; Breaker, R.R. Riboswitches Control Fundamental Biochemical Pathways in Bacillus subtilis and Other Bacteria. Cell 2003, 113, 577–586. [Google Scholar] [CrossRef] [Green Version]
- Breaker, R.R. Riboswitches and the RNA World. Cold Spring Harb. Perspect. Biol. 2010, 4, a003566. [Google Scholar] [CrossRef] [Green Version]
- Serganov, A.; Nudler, E. A Decade of Riboswitches. Cell 2013, 152, 17–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCown, P.J.; Corbino, K.A.; Stav, S.; Sherlock, M.E.; Breaker, R.R. Riboswitch diversity and distribution. RNA 2017, 23, 995–1011. [Google Scholar] [CrossRef] [PubMed]
- Mandal, M.; Lee, M.; Barrick, J.E.; Weinberg, Z.; Emilsson, G.M.; Ruzzo, W.L.; Breaker, R.R. A glycine-dependent ri-boswitch that uses cooperative binding to control gene expression. Science 2004, 306, 275–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodionov, D.A.; Vitreschak, A.G.; Mironov, A.A.; Gelfand, M.S. Regulation of lysine biosynthesis and transport genes in bacteria: Yet another RNA riboswitch? Nucleic Acids Res. 2003, 31, 6748–6757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batey, R.T.; Gilbert, S.D.; Montange, R.K. Structure of a natural guanine-responsive riboswitch complexed with the me-tabolite hypoxanthine. Nature 2004, 432, 411–415. [Google Scholar] [CrossRef]
- Ames, T.D.; Rodionov, D.A.; Weinberg, Z.; Breaker, R.R. A eubacterial riboswitch class that senses the coenzyme tetrahy-drofolate. Chem. Biol. 2010, 17, 681–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudarsan, N.; Lee, E.R.; Weinberg, Z.; Moy, R.H.; Kim, J.N.; Link, K.H.; Breaker, R.R. Riboswitches in Eubacteria Sense the Second Messenger Cyclic Di-GMP. Science 2008, 321, 411–413. [Google Scholar] [CrossRef] [Green Version]
- Montange, R.K.; Batey, R.T. Riboswitches: Emerging Themes in RNA Structure and Function. Annu. Rev. Biophys. 2008, 37, 117–133. [Google Scholar] [CrossRef] [PubMed]
- Sherwood, A.V.; Henkin, T.M. Riboswitch-Mediated Gene Regulation: Novel RNA Architectures Dictate Gene Expression Responses. Annu. Rev. Microbiol. 2016, 70, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Grundy, F.J.; Henkin, T.M. The S box regulon: A new global transcription termination control system for methionine and cysteine biosynthesis genes in Gram-positive bacteria. Mol. Microbiol. 1998, 30, 737–749. [Google Scholar] [CrossRef]
- McDaniel, B.A.; Grundy, F.J.; Henkin, T.M. A tertiary structural element in S box leader RNAs is required for S-adenosylmethionine-directed transcription termination. Mol. Microbiol. 2005, 57, 1008–1021. [Google Scholar] [CrossRef] [PubMed]
- Green, N.J.; Grundy, F.J.; Henkin, T.M. The T box mechanism: tRNA as a regulatory molecule. FEBS Lett. 2009, 584, 318–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreuzer, K.D.; Henkin, T.M. The T-Box Riboswitch: tRNA as an effector to modulate gene regulation. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef]
- Grundy, F.J.; Rollins, S.M.; Henkin, T.M. Interaction between the acceptor end of tRNA and the T box stimulates antiter-mination in the Bacillus subtilis tyrS gene: A new role for the discriminator base. J. Bacterial. 1994, 176, 4518–4526. [Google Scholar] [CrossRef] [Green Version]
- Chopin, A.; Biaudet, V.; Ehrlich, S.D. Analysis of the Bacillus subtilis genome sequence reveals nine new T-box leaders. Mol. Microbiol. 1998, 29, 662–664. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Preciado, A.; Henkin, T.M.; Grundy, F.J.; Yanofsky, C.; Merino, E. Biochemical features and functional implica-tions of the RNA-based T-box regulatory mechanism. Microbiol. Mol. Biol. Rev. 2009, 73, 36–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weaver, J.W.; Serganov, A. T-box RNA gets boxed. Nat. Struct. Mol. Biol. 2019, 26, 1081–1083. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J. Unboxing the T-box riboswitches-A glimpse into multivalent and multimodal RNA-RNA interactions. Wiley In-terdiscip. Rev. RNA 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Henkin, T.M.; Glass, B.L.; Grundy, F.J. Analysis of the Bacillus subtilis tyrS gene: Conservation of a regulatory sequence in multiple tRNA synthetase genes. J. Bacteriol. 1992, 174, 1299–1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grigg, J.C.; Ke, A. Structural Determinants for Geometry and Information Decoding of tRNA by T Box Leader RNA. Struct. 2013, 21, 2025–2032. [Google Scholar] [CrossRef] [Green Version]
- Klein, D.; Schmeing, T.; Moore, P.; Steitz, T. The kink-turn: A new RNA secondary structure motif. In The Excitement of Discovery: Selected Papers of Alexander Rich; World Scientific Publishing: Singapore, 2020; Volume 20, pp. 517–524. [Google Scholar] [CrossRef]
- Zhang, J.; Ferré-D’Amaré, A.R. Co-crystal structure of a T-box riboswitch stem I domain in complex with its cognate tRNA. Nat. Cell Biol. 2013, 500, 363–366. [Google Scholar] [CrossRef]
- Grundy, F.J.; Winkler, W.C.; Henkin, T.M. tRNA-mediated transcription antitermination in vitro: Codon-anticodon pairing independent of the ribosome. Proc. Natl. Acad. Sci. USA 2002, 99, 11121–11126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haitani, Y.; Awano, N.; Yamazaki, M.; Wada, M.; Nakamori, S.; Takagi, H. Functional analysis of l-serineO-acetyltransferase fromCorynebacterium glutamicum. FEMS Microbiol. Lett. 2006, 255, 156–163. [Google Scholar] [CrossRef] [Green Version]
- Netzer, R.; Peters-Wendisch, P.; Eggeling, L.; Sahm, H. Cometabolism of a Nongrowth Substrate: L-Serine Utilization by Corynebacterium glutamicum. Appl. Environ. Microbiol. 2004, 70, 7148–7155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Locasale, J.W. Serine, glycine and one-carbon units: Cancer metabolism in full circle. Nat. Rev. Cancer 2013, 13, 572–583. [Google Scholar] [CrossRef] [Green Version]
- Schaechter, M. Escherichia coli and Salmonella 2000: The view from here. Microbiol. Mol. Biol. Rev. 2001, 65, 119–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Shang, X.; Lai, S.; Zhang, Y.; Hu, Q.; Chai, X.; Wang, B.; Liu, S.; Wen, T. Reprogramming one-carbon metabolic pathways to decouple L-Serine catabolism from cell growth in Corynebacterium glutamicum. ACS Synth. Biol. 2018, 7, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Mundhada, H.; Schneider, K.; Christensen, H.B.; Nielsen, A.T. Engineering of high yield production of L-serine in Esche-richia coli. Biotechnol. Bioeng. 2016, 113, 807–816. [Google Scholar] [CrossRef] [PubMed]
- Hama, H.; Sumita, Y.; Kakutani, Y.; Tsuda, M.; Tsuchiya, T. Target of serine inhibition in Escherichia coli. Biochem. Biophys. Res. Commun. 1990, 168, 1211–1216. [Google Scholar] [CrossRef]
- Sun, E.I.; Leyn, S.A.; Kazanov, M.D.; Saier, M.J.; Novichkov, P.S.; Rodionov, D.A. Comparative genomics of metabolic ca-pacities of regulons controlled by cis-regulatory RNA motifs in bacteria. BMC Genomics 2013, 14, 597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonenshein, A.L. Control of key metabolic intersections in Bacillus subtilis. Nat. Rev. Genet. 2007, 5, 917–927. [Google Scholar] [CrossRef] [PubMed]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef] [PubMed]
- Marchand, J.A.; Pierson Smela, M.D.; Jordan, T.H.H.; Narasimhan, K.; Church, G.M. TBDB: A database of structurally an-notated T-box riboswitch: tRNA pairs. Nucleic Acids Res. 2020, 49, D229–D235. [Google Scholar] [CrossRef]
- Klewing, A.; Koo, B.M.; Krüger, L.; Poehlein, A.; Reuß, D.; Daniel, R.; Gross, C.A.; Stülke, J. Resistance to serine in Bacillus subtilis: Identification of the serine transporter YbeC and of a metabolic network that links serine and threonine metabo-lism. Environ. Microbiol. 2020, 22, 3937–3949. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, A.; Fu, H.A.; Burris, R.H. Influence of amino acids on nitrogen fixation ability and growth of Azospirillum spp. Appl. Environ. Microbiol. 1988, 54, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhang, L.; Liu, Z.; Zhao, D.; Liu, X.; Zhang, B.; Xie, J.; Hong, Y.; Li, P.; Chen, S.; et al. A minimal nitro-gen fixation gene cluster from Paenibacillus sp. WLY78 enables expression of active nitrogenase in Escherichia coli. PLoS Genet. 2013, 9, e1003865. [Google Scholar] [CrossRef]
- Koo, B.; Kritikos, G.; Farelli, J.D.; Todor, H.; Tong, K.; Kimsey, H.; Wapinski, I.; Galardini, M.; Cabal, A.; Peters, J.M.; et al. Construction and analysis of two genome-scale deletion librar-ies for Bacillus subtilis. Cell Syst. 2017, 4, 291–305. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Li, Q.; Li, Y.; Chen, S. The Serine Biosynthesis of Paenibacillus polymyxa WLY78 Is Regulated by the T-Box Riboswitch. Int. J. Mol. Sci. 2021, 22, 3033. https://doi.org/10.3390/ijms22063033
Zhang H, Li Q, Li Y, Chen S. The Serine Biosynthesis of Paenibacillus polymyxa WLY78 Is Regulated by the T-Box Riboswitch. International Journal of Molecular Sciences. 2021; 22(6):3033. https://doi.org/10.3390/ijms22063033
Chicago/Turabian StyleZhang, Haowei, Qin Li, Yongbin Li, and Sanfeng Chen. 2021. "The Serine Biosynthesis of Paenibacillus polymyxa WLY78 Is Regulated by the T-Box Riboswitch" International Journal of Molecular Sciences 22, no. 6: 3033. https://doi.org/10.3390/ijms22063033
APA StyleZhang, H., Li, Q., Li, Y., & Chen, S. (2021). The Serine Biosynthesis of Paenibacillus polymyxa WLY78 Is Regulated by the T-Box Riboswitch. International Journal of Molecular Sciences, 22(6), 3033. https://doi.org/10.3390/ijms22063033