
The Role of Endoglin in Hepatocellular Carcinoma


Abstract
:1. Introduction
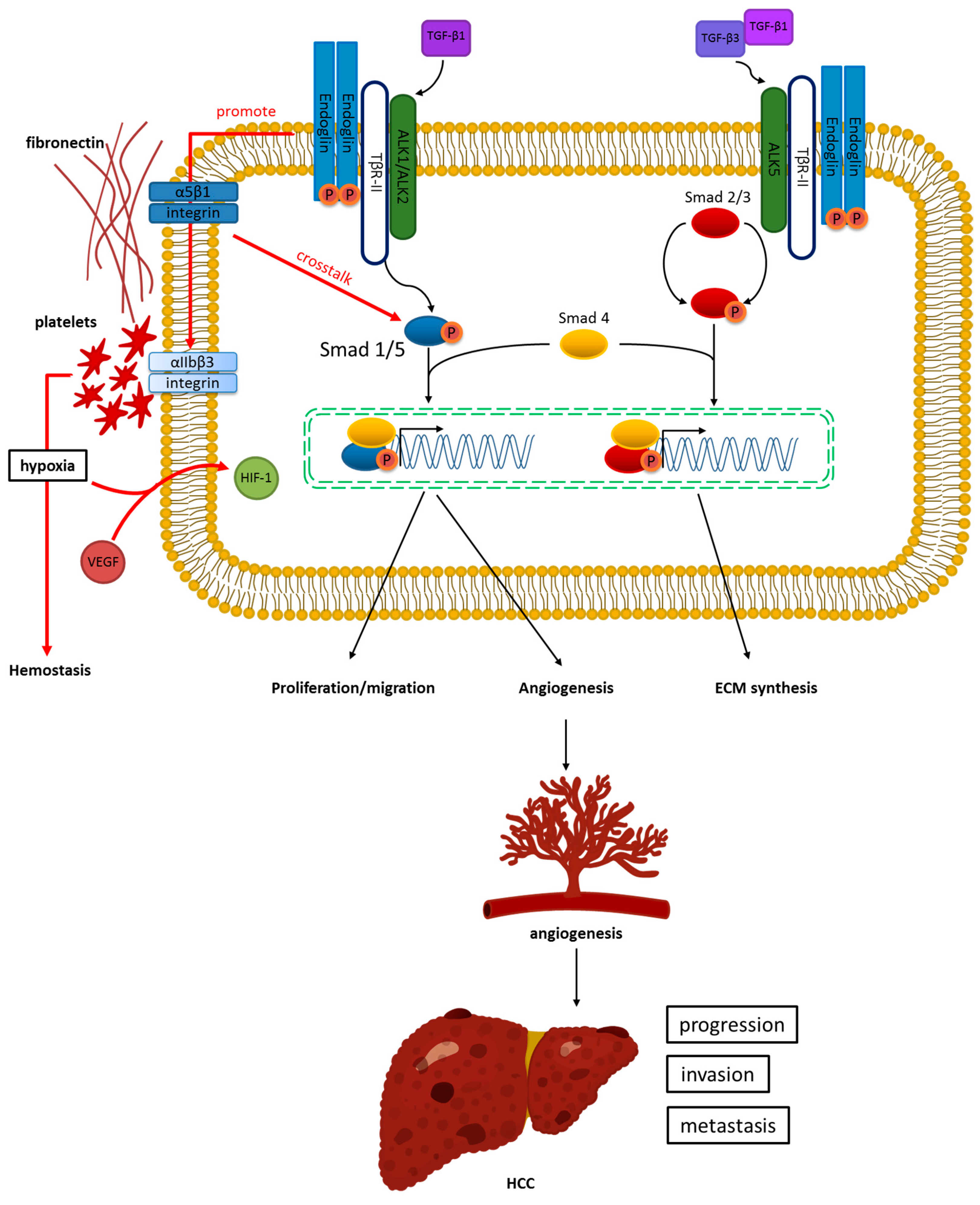
2. Structure, Function, and Signaling Pathway of Endoglin
3. Angiogenesis of HCC and Its Relation to Endoglin
4. Circulating Endoglin in HCC Patients
5. Hepatitis C Virus Core Protein Modulates the Endoglin Signaling Pathway and the Role of Endoglin in Cancer Stem Cells, Hepatic Stellate Cells and Cancer Associated Fibroblasts
6. Endoglin in Tumor-Derived Endothelial Cells and HCC
7. Endoglin Detection and the Inhibition of Endoglin in HCC Therapy
8. Endoglin as a Biomarker and Controversial Issues in Endoglin-Targeted Therapy
9. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sayiner, M.; Golabi, P.; Younossi, Z.M. Disease Burden of Hepatocellular Carcinoma: A Global Perspective. Dig. Dis. Sci. 2019, 64, 910–917. [Google Scholar] [CrossRef]
- Mossenta, M.; Busato, D.; Baboci, L.; Di Cintio, F.; Toffoli, G.; Bo, M.D. New Insight into Therapies Targeting Angiogenesis in Hepatocellular Carcinoma. Cancers 2019, 11, 1086. [Google Scholar] [CrossRef] [Green Version]
- Jamshidi-Parsian, A.; Griffin, R.J.; Kore, R.A.; Todorova, V.K.; Makhoul, I. Tumor-Endothelial Cell Interaction in an Experimental Model of Human Hepatocellular Carcinoma. Exp. Cell Res. 2018, 372, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Paschoal, J.P.; Bernardo, V.; Canedo, N.H.S.; Ribeiro, O.D.; Caroli-Bottino, A.; Pannain, V.L. Microvascular Density of Regenerative Nodule to Small Hepatocellular Carcinoma by Automated Analysis Using CD105 and CD34 Immunoexpression. BMC Cancer 2014, 14, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Bergers, G.; Benjamin, L.E. Tumorigenesis and the Angiogenic Switch. Nat. Rev. Cancer 2003, 3, 401–410. [Google Scholar] [CrossRef]
- Kerbel, R.S. Tumor Angiogenesis. N. Engl. J. Med. 2008, 358, 2039–2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, C.; Shao, S.; Yan, Y.; Yang, S.; Bai, S.; Wu, Y.; Zhang, J.; Liu, R.; Ma, H.; Chai, L.; et al. Association between G-Protein Coupled Receptor 4 Expression and Microvessel Density, Clinicopathological Characteristics and Survival in Hepatocellular Carcinoma. Oncol. Lett. 2020, 19, 2609–2620. [Google Scholar] [CrossRef]
- Yang, L.-Y.; Lu, W.-Q.; Huang, G.-W.; Wang, W. Correlation between CD105 Expression and Postoperative Recurrence and Metastasis of Hepatocellular Carcinoma. BMC Cancer 2006, 6, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gougos, A.; Letarte, M. Identification of a Human Endothelial Cell Antigen with Monoclonal Antibody 44G4 Produced against a Pre-B Leukemic Cell Line. J. Immunol. 1988, 141, 1925–1933. [Google Scholar]
- Fernández-Ruiz, E.; St-Jacques, S.; Bellón, T.; Letarte, M.; Bernabéu, C. Assignment of the Human Endoglin Gene (END) to 9q34→qter. Cytogenet. Genome Res. 1993, 64, 204–207. [Google Scholar] [CrossRef] [PubMed]
- Gougos, A.; Letarte, M. Primary Structure of Endoglin, an RGD-Containing Glycoprotein of Human Endothelial Cells. J. Biol. Chem. 1990, 265, 8361–8364. [Google Scholar] [CrossRef]
- López-Novoa, J.M.; Bernabeu, C. The Physiological Role of Endoglin in the Cardiovascular System. Am. J. Physiol. Circ. Physiol. 2010, 299, H959–H974. [Google Scholar] [CrossRef] [Green Version]
- Maring, J.A.; Trojanowska, M.; Dijke, P.T. Role of Endoglin in Fibrosis and Scleroderma. Int. Rev. Cell Mol. Biol. 2012, 297, 295–308. [Google Scholar] [CrossRef] [Green Version]
- Kwon, Y.-C.; Sasaki, R.; Meyer, K.; Ray, R. Hepatitis C Virus Core Protein Modulates Endoglin (CD105) Signaling Pathway for Liver Pathogenesis. J. Virol. 2017, 91, e01235-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasprzak, A.; Adamek, A. Role of Endoglin (CD105) in the Progression of Hepatocellular Carcinoma and Anti-Angiogenic Therapy. Int. J. Mol. Sci. 2018, 19, 3887. [Google Scholar] [CrossRef] [Green Version]
- Ray, B.N.; Lee, N.Y.; How, T.; Blobe, G.C. ALK5 Phosphorylation of the Endoglin Cytoplasmic Domain Regulates Smad1/5/8 Signaling and Endothelial Cell Migration. Carcinogenesis 2009, 31, 435–441. [Google Scholar] [CrossRef] [Green Version]
- Bernabeu, C.; Lopez-Novoa, J.M.; Quintanilla, M. The Emerging Role of TGF-β Superfamily Coreceptors in Cancer. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2009, 1792, 954–973. [Google Scholar] [CrossRef] [PubMed]
- Finnson, K.W.; Philip, A. Endoglin in Liver Fibrosis. J. Cell Commun. Signal. 2011, 6, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellón, T.; Corbi, A.; Lastres, P.; Calés, C.; Cebrián, M.; Vera, S.; Cheifetz, S.; Massagué, J.; Letarte, M.; Bernabeu, C. Identification and Expression of Two Forms of the Human Transforming Growth Factor-β-Binding Protein Endoglin with Distinct Cytoplasmic Regions. Eur. J. Immunol. 1993, 23, 2340–2345. [Google Scholar] [CrossRef]
- Blanco, F.J.; Bernabeu, C. Alternative Splicing Factor or Splicing Factor-2 Plays a Key Role in Intron Retention of the Endoglin Gene during Endothelial Senescence. Aging Cell 2011, 10, 896–907. [Google Scholar] [CrossRef] [PubMed]
- Alsamman, M.; Sterzer, V.; Meurer, S.K.; Sahin, H.; Schaeper, U.; Kuscuoglu, D.; Strnad, P.; Weiskirchen, R.; Trautwein, C.; Scholten, D. Endoglin in Human Liver Disease and Murine Models of Liver Fibrosis—A Protective Factor against Liver Fibrosis. Liver Int. 2017, 38, 858–867. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Gómez, E.; Eleno, N.; López-Novoa, J.M.; Ramirez, J.R.; Velasco, B.; Letarte, M.; Bernabeu, C.; Quintanilla, M. Characterization of Murine S-endoglin Isoform and Its Effects on Tumor Development. Oncogene 2005, 24, 4450–4461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meurer, S.K.; Tihaa, L.; Borkham-Kamphorst, E.; Weiskirchen, R. Expression and Functional Analysis of Endoglin in Isolated Liver Cells and Its Involvement in Fibrogenic Smad Signalling. Cell. Signal. 2011, 23, 683–699. [Google Scholar] [CrossRef]
- Velasco, S.; Pericacho, M.; Alvarez-Muñoz, P.; Rodríguez-Barbero, A.; Dijke, P.T.; Bernabeu, C.; López-Novoa, J.M. L-and S-endoglin Differentially Modulate TGF 1 Signaling Mediated by ALK1 and ALK5 in L6E9 Myoblasts. J. Cell Sci. 2008, 121, 913–919. [Google Scholar] [CrossRef] [Green Version]
- Lebrin, F.; Deckers, M.; Bertolino, P.; Dijke, P.T. TGF-β Receptor Function in the Endothelium. Cardiovasc. Res. 2005, 65, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Dallas, N.A.; Samuel, S.; Xia, L.; Fan, F.; Gray, M.J.; Lim, S.J.; Ellis, L.M. Endoglin (CD105): A Marker of Tumor Vasculature and Potential Target for Therapy. Clin. Cancer Res. 2008, 14, 1931–1937. [Google Scholar] [CrossRef] [Green Version]
- Cheifetz, S.; Bellon, T.; Cales, C.; Vera, S.; Bernabeu, C.; Massague, J.; Letarte, M. Endoglin Is a Component of the Trans-Forming Growth Factor-Beta Receptor System in Human Endothelial Cells. J. Biol. Chem. 1992, 267, 19027–19030. [Google Scholar] [CrossRef]
- Fonsatti, E.; Del Vecchio, L.; Altomonte, M.; Sigalotti, L.; Nicotra, M.R.; Coral, S.; Natali, P.G.; Maio, M. Endoglin: An Accessory Component of the TGF-β-Binding Receptor-Complex with Diagnostic, Prognostic, and Bioimmunotherapeutic Potential in Human Malignancies. J. Cell. Physiol. 2001, 188, 1–7. [Google Scholar] [CrossRef]
- Guerrero-Esteo, M.; Sánchez-Elsner, T.; Letamendia, A.; Bernabéu, C. Extracellular and Cytoplasmic Domains of Endoglin Interact with the Transforming Growth Factor-β Receptors I and II. J. Biol. Chem. 2002, 277, 29197–29209. [Google Scholar] [CrossRef] [Green Version]
- Blanco, F.J.; Santibanez, J.F.; Guerrero-Esteo, M.; Langa, C.; Vary, C.P.H.; Bernabeu, C. Interaction and Functional Interplay between Endoglin and ALK-1, Two Components of the Endothelial Transforming Growth Factor-β Receptor Complex. J. Cell. Physiol. 2005, 204, 574–584. [Google Scholar] [CrossRef]
- Pomeraniec, L.; Hector-Greene, M.; Ehrlich, M.; Blobe, G.C.; Henis, Y.I. Regulation of TGF-β Receptor Hetero-Oligomerization and Signaling by Endoglin. Mol. Biol. Cell 2015, 26, 3117–3127. [Google Scholar] [CrossRef] [PubMed]
- Rius, C.; Smith, J.D.; Almendro, N.; Langa, C.; Botella, L.M.; Marchuk, D.A.; Vary, C.P.; Bernabeu, C. Cloning of the Promoter Region of Human Endoglin, the Target Gene for Hereditary Hemorrhagic Telangiectasia Type 1. Blood 1998, 92, 4677–4690. [Google Scholar] [CrossRef]
- Nassiri, F.; Cusimano, M.D.; Scheithauer, B.W.; Rotondo, F.; Fazio, A.; Yousef, G.M.; Syro, L.V.; Kovacs, K.; Lloyd, R.V. Endoglin (CD105): A Review of Its Role in Angiogenesis and Tumor Diagnosis, Progression and Therapy. Anticancer. Res. 2011, 31, 2283–2290. [Google Scholar]
- Lebrin, F.; Goumans, M.-J.; Jonker, L.; Carvalho, R.L.C.; Valdimarsdottir, G.; Thorikay, M.; Mummery, C.; Arthur, H.M.; Dijke, P.T. Endoglin Promotes Endothelial Cell Proliferation and TGF-β/ALK1 Signal Transduction. EMBO J. 2004, 23, 4018–4028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapur, N.K.; Morine, K.J.; Letarte, M. Endoglin: A Critical Mediator of Cardiovascular Health. Vasc. Health Risk Manag. 2013, 9, 195–206. [Google Scholar] [CrossRef] [Green Version]
- Rossi, E.; Sanz-Rodriguez, F.; Eleno, N.; Düwell, A.; Blanco, F.J.; Langa, C.; Botella, L.M.; Cabañas, C.; Lopez-Novoa, J.M.; Bernabeu, C. Endothelial Endoglin Is Involved in Inflammation: Role in Leukocyte Adhesion and Transmigration. Blood 2013, 121, 403–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Barbero, A.; Obreo, J.; Álvarez-Muñoz, P.; Pandiella, A.; Bernabéu, C.; López-Novoa, J.M. Endoglin Modulation of TGF-ß1-Induced Collagen Synthesis is Dependent on ERK1/2 MAPK Activation. Cell. Physiol. Biochem. 2006, 18, 135–142. [Google Scholar] [CrossRef]
- Lee, N.Y.; Blobe, G.C. The Interaction of Endoglin with β-Arrestin2 Regulates Transforming Growth Factor-β-mediated ERK Activation and Migration in Endothelial Cells. J. Biol. Chem. 2007, 282, 21507–21517. [Google Scholar] [CrossRef] [Green Version]
- Scherner, O.; Meurer, S.K.; Tihaa, L.; Gressner, A.M.; Weiskirchen, R. Endoglin Differentially Modulates Antagonistic Transforming Growth Factor-β1 and BMP-7 Signaling*. J. Biol. Chem. 2007, 282, 13934–13943. [Google Scholar] [CrossRef] [Green Version]
- Tian, H.; Mythreye, K.; Golzio, C.; Katsanis, N.; Blobe, G.C. Endoglin Mediates Fibronectin/α5β1 Integrin and TGF-β Pathway Crosstalk in Endothelial Cells. EMBO J. 2012, 31, 3885–3900. [Google Scholar] [CrossRef] [Green Version]
- Rossi, E.; Pericacho, M.; Bachelot-Loza, C.; Pidard, D.; Gaussem, P.; Poirault-Chassac, S.; Blanco, F.J.; Langa, C.; González-Manchón, C.; Novoa, J.M.L.; et al. Human Endoglin as a Potential New Partner Involved in Platelet–Endothelium Interactions. Cell. Mol. Life Sci. 2018, 75, 1269–1284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onoe, T.; Ohdan, H.; Tokita, D.; Hara, H.; Tanaka, Y.; Ishiyama, K.; Asahara, T. Liver Sinusoidal Endothelial Cells Have a Capacity for Inducing Nonresponsiveness of T Cells across Major Histocompatibility Complex Barriers. Transpl. Int. 2005, 18, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Zhu, A.X.; Duda, D.G.; Sahani, D.V.; Jain, R.K. HCC and Angiogenesis: Possible Targets and Future Directions. Nat. Rev. Clin. Oncol. 2011, 8, 292–301. [Google Scholar] [CrossRef]
- Honda, H.; Tajima, T.; Kajiyama, K.; Kuroiwa, T.; Yoshimitsu, K.; Irie, H.; Aibe, H.; Shimada, M.; Masuda, K. Vascular Changes in Hepatocellular Carcinoma: Correlation of Radiologic and Pathologic Findings. Am. J. Roentgenol. 1999, 173, 1213–1217. [Google Scholar] [CrossRef] [Green Version]
- OsamuMatsui, O.; Kobayashi, S.; Sanada, J.; Kouda, W.; Ryu, Y.; Kozaka, K.; Kitao, A.; Nakamura, K.; Gabata, T. Hepatocelluar Nodules in Liver Cirrhosis: Hemodynamic Evaluation (Angiography-Assisted CT) with Special Reference to Multi-Step Hepatocarcinogenesis. Abdom. Imaging 2011, 36, 264–272. [Google Scholar] [CrossRef] [Green Version]
- Poisson, J.; Lemoinne, S.; Boulanger, C.; Durand, F.; Moreau, R.; Valla, D.; Rautou, P.-E. Liver Sinusoidal Endothelial Cells: Physiology and Role in Liver Diseases. J. Hepatol. 2017, 66, 212–227. [Google Scholar] [CrossRef] [Green Version]
- Muto, J.; Shirabe, K.; Sugimachi, K.; Maehara, Y. Review of Angiogenesis in Hepatocellular Carcinoma. Hepatol. Res. 2014, 45, 1–9. [Google Scholar] [CrossRef]
- Sanz-Cameno, P.; Trapero-Marugán, M.; Chaparro, M.; Jones, E.A.; Moreno-Otero, R. Angiogenesis: From Chronic Liver Inflammation to Hepatocellular Carcinoma. J. Oncol. 2010, 2010, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.F.; Poon, R.T. Vascular Changes in Hepatocellular Carcinoma. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2008, 291, 721–734. [Google Scholar] [CrossRef]
- Sun, B.; Zhang, S.; Zhang, D.; Du, J.; Guo, H.; Zhao, X.; Zhang, W.; Hao, X. Vasculogenic Mimicry Is Associated with High Tumor Grade, Invasion and Metastasis, and Short Survival in Patients with Hepatocellular Carcinoma. Oncol. Rep. 2006, 16, 693–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Zhao, X.; Zhu, D.; Liu, T.; Liang, X.; Liu, F.; Zhang, Y.; Dong, X.; Sun, B. HIF-1α Promoted Vasculogenic Mimicry Formation in Hepatocellular Carcinoma through LOXL2 up-Regulation in Hypoxic Tumor Microenvironment. J. Exp. Clin. Cancer Res. 2017, 36, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segatelli, V.; De Oliveira, E.C.; Boin, I.F.S.F.; Ataide, E.C.; Escanhoela, C.A.F. Evaluation and Comparison of Microvessel Density Using the Markers CD34 and CD105 in Regenerative Nodules, Dysplastic Nodules and Hepatocellular Carcinoma. Hepatol. Int. 2014, 8, 260–265. [Google Scholar] [CrossRef]
- Yu, D.; Sun, X.; Qiu, Y.; Zhou, J.; Wu, Y.; Zhuang, L.; Chen, J.; Ding, Y. Identification and Clinical Significance of Mobilized Endothelial Progenitor Cells in Tumor Vasculogenesis of Hepatocellular Carcinoma. Clin. Cancer Res. 2007, 13, 3814–3824. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.-C.; Chen, J.; Ding, Y.-T. Hypoxic and Highly Angiogenic Non-Tumor Tissues Surrounding Hepatocellular Carcinoma: The ‘Niche’ of Endothelial Progenitor Cells. Int. J. Mol. Sci. 2010, 11, 2901–2909. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.N.; Kim, Y.B.; Yang, K.M.; Park, C. Increased Expression of Vascular Endothelial Growth Factor and Angiogenesis in the Early Stage of Multistep Hepatocarcinogenesis. Arch. Pathol. Lab. Med. 2000, 124, 1061–1065. [Google Scholar] [CrossRef]
- Teixeira, A.C.; Brasil, I.R.; Torres, A.F.; Tavora, F. The Evaluation of Angiogenesis Markers in Hepatocellular Carcinoma and Precursor Lesions in Liver Explants from a Single Institution. Appl. Immunohistochem. Mol. Morphol. 2018, 26, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Kin, M.; Torimura, T.; Ueno, T.; Inuzuka, S.; Tanikawa, K. Sinusoidal Capillarization in Small Hepatocellular Carcinoma. Pathol. Int. 1994, 44, 771–778. [Google Scholar] [CrossRef]
- Poon, R.T.-P.; Ng, I.O.-L.; Lau, C.; Yu, W.-C.; Yang, Z.-F.; Fan, S.-T.; Wong, J. Tumor Microvessel Density as a Predictor of Recurrence After Resection of Hepatocellular Carcinoma: A Prospective Study. J. Clin. Oncol. 2002, 20, 1775–1785. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Ghellal, A.; Li, C.; Byrne, G.; Haboubi, N.; Wang, J.M.; Bundred, N. Breast Carcinoma: Vascular Density Determined Using CD105 Antibody Correlates with Tumor Prognosis. Cancer Res. 1999, 59, 856–861. [Google Scholar] [PubMed]
- Fonsatti, E.; Altomonte, M.; Nicotra, M.R.; Natali, P.G.; Maio, M. Endoglin (CD105): A Powerful Therapeutic Target on Tumor-Associated Angiogenetic Blood Vessels. Oncogene 2003, 22, 6557–6563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, S.; Muro, H.; Suzuki, S.; Sakaguchi, T.; Konno, H.; Baba, S.; Syed, A.S. Immunohistochemical Studies on Endothelial Cell Phenotype in Hepatocellular Carcinoma. Hepatology 1997, 26, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Pan, Y.; Chen, J.; Sun, X.; Qiu, Y.; Ding, Y. Endoglin (CD105) Expression in Angiogenesis of Primary Hepatocellular Carcinomas: Analysis Using Tissue Microarrays and Comparisons with CD34 and VEGF. Ann. Clin. Lab. Sci. 2007, 37, 39–48. [Google Scholar]
- Ho, J.W.; Poon, R.T.; Sun, C.K.; Xue, W.-C.; Fan, S.-T. Clinicopathological and Prognostic Implications of Endoglin (CD105) Expression in Hepatocellular Carcinoma and Its Adjacent Non-tumorous Liver. World J. Gastroenterol. 2005, 11, 176–181. [Google Scholar] [CrossRef] [Green Version]
- Benetti, A.; Berenzi, A.; Gambarotti, M.; Garrafa, E.; Gelati, M.; Dessy, E.; Portolani, N.; Piardi, T.; Giulini, S.M.; Caruso, A.; et al. Transforming Growth Factor-β1 and CD105 Promote the Migration of Hepatocellular Carcinoma–Derived Endothelium. Cancer Res. 2008, 68, 8626–8634. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Yang, L.; Chen, X.; Qian, H.; Zhang, S.; Chen, Y.; Luo, R.; Shao, J.; Liu, H.; Chen, J. Phenotypic and Functional Characterization of Tumor-Derived Endothelial Cells Isolated from Primary Human Hepatocellular Carcinoma. Hepatol. Res. 2018, 48, 1149–1162. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, X.-H.; Guo, P.; Yan, L.-N.; He, D. Tumor Microvascular Density Detected by Anti-CD105 and Anti-CD34 in Hepatocellular Carcinoma Patients and Its Predictive Value of Tumor Recurrence after Liver Transplantation. Sichuan Da Xue Xue Bao. Yi Xue Ban/J. Sichuan Univ. Med Sci. Ed. 2010, 41, 818–821. [Google Scholar]
- Xiong, Y.-Q.; Sun, H.-C.; Zhang, W.; Zhu, X.-D.; Zhuang, P.-Y.; Zhang, J.-B.; Wang, L.; Wu, W.-Z.; Qin, L.-X.; Tang, Z.-Y. Human Hepatocellular Carcinoma Tumor–derived Endothelial Cells Manifest Increased Angiogenesis Capability and Drug Resistance Compared with Normal Endothelial Cells. Clin. Cancer Res. 2009, 15, 4838–4846. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Elsner, T.; Botella, L.M.; Velasco, B.; Langa, C.; Bernabéu, C. Endoglin Expression Is Regulated by Transcriptional Cooperation between the Hypoxia and Transforming Growth Factor-β Pathways. J. Biol. Chem. 2002, 277, 43799–43808. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Guo, B.; Ding, S.; Rius, C.; Langa, C.; Kumar, P.; Bernabeu, C.; Kumar, S. TNF Alpha Down-Regulates CD105 Expression in Vascular Endothelial Cells: A Comparative Study with TGF beta 1. Anticancer Res. 2003, 23, 1189–1196. [Google Scholar]
- Tian, F.; Zhou, A.-X.; Smits, A.M.; Larsson, E.; Goumans, M.-J.; Heldin, C.-H.; Borén, J.; Akyürek, L.M. Endothelial Cells Are Activated during Hypoxia via Endoglin/ALK-1/SMAD1/5 Signaling in Vivo and in Vitro. Biochem. Biophys. Res. Commun. 2010, 392, 283–288. [Google Scholar] [CrossRef]
- Wikström, P.; Lissbrant, I.F.; Stattin, P.; Egevad, L.; Bergh, A. Endoglin (CD105) is Expressed on Immature Blood Vessels and Is a Marker for Survival in Prostate Cancer. Prostate 2002, 51, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Darcy, K.M.; Birrer, M.J. Translational Research in the Gynecologic Oncology Group: Evaluation of Ovarian Cancer Markers, Profiles, and Novel Therapies. Gynecol. Oncol. 2010, 117, 429–439. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, F.; Otake, Y.; Yanagihara, K.; Kawano, Y.; Miyahara, R.; Li, M.; Yamada, T.; Hanaoka, N.; Inui, K.; Wada, H. Evaluation of Angiogenesis in Non-small Cell Lung Cancer: Comparison between Anti-CD34 Antibody and Anti-CD105 Antibody. Clin. Cancer Res. 2001, 7, 3410–3415. [Google Scholar]
- Margaritescu, C.; Simionescu, C.; Mogoanta, L.; Badea, P.; Pirici, D.; Stepan, A.; Ciurea, R. Endoglin (CD105) and Microvessel Density in Oral Squamous Cell Carcinoma. Rom. J. Morphol. Embryol. 2008, 49, 321–326. [Google Scholar]
- Henry-Berger, J.; Mouzat, K.; Baron, S.; Bernabeu, C.; Marceau, G.; Saru, J.-P.; Sapin, V.; Lobaccaro, J.-M.A.; Caira, F. Endoglin (CD105) Expression Is Regulated by the Liver X Receptor Alpha (NR1H3) in Human Trophoblast Cell Line JAR1. Biol. Reprod. 2008, 78, 968–975. [Google Scholar] [CrossRef] [Green Version]
- Yagmur, E.; Rizk, M.; Stanzel, S.; Hellerbrand, C.; Lammert, F.; Trautwein, C.; Wasmuth, H.E.; Gressner, A.M. Elevation of Endoglin (CD105) Concentrations in Serum of Patients with Liver Cirrhosis and Carcinoma. Eur. J. Gastroenterol. Hepatol. 2007, 19, 755–761. [Google Scholar] [CrossRef]
- Hawinkels, L.J.; Kuiper, P.; Wiercinska, E.; Verspaget, H.W.; Liu, Z.; Pardali, E.; Sier, C.F.; Dijke, P.T. Matrix Metalloproteinase-14 (MT1-MMP)–Mediated Endoglin Shedding Inhibits Tumor Angiogenesis. Cancer Res. 2010, 70, 4141–4150. [Google Scholar] [CrossRef] [Green Version]
- Gallardo-Vara, E.; Blanco, F.J.; Roqué, M.; Friedman, S.L.; Suzuki, T.; Botella, L.M.; Bernabeu, C. Transcription Factor KLF6 Upregulates Expression of Metalloprotease MMP14 and Subsequent Release of Soluble Endoglin during Vascular Injury. Angiogenesis 2016, 19, 155–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Gómez, E.; Del Castillo, G.; Santibáñez, J.F.; Lêpez-Novoa, J.M.; Bernabéu, C.; Quintanilla, M. The Role of the TGF-β Coreceptor Endoglin in Cancer. Sci. World J. 2010, 10, 2367–2384. [Google Scholar] [CrossRef] [Green Version]
- Duff, S.E.; Li, C.; Garland, J.M.; Kumar, S. CD105 is Important for Angiogenesis: Evidence and Potential Applications. FASEB J. 2003, 17, 984–992. [Google Scholar] [CrossRef]
- Elnemr, D.M.; Abdel-Azeez, H.A.; Labib, H.A.; Abo-Taleb, F.M. Clinical Relevance of Serum Endoglin Level in Egyptian Hepatocellular Carcinoma Patients. Clin. Lab. 2012, 58, 1023–1028. [Google Scholar] [PubMed]
- Teama, S.; Fawzy, A.; Teama, S.; Helal, A.; Drwish, A.D.; Elbaz, T.; Desouky, E. Increased Serum Endoglin and Transforming Growth Factor β1 mRNA Expression and Risk of Hepatocellular Carcinoma in Cirrhotic Egyptian Patients. Asian Pac. J. Cancer Prev. 2016, 17, 2429–2434. [Google Scholar]
- Li, Y.; Zhai, Z.; Liu, D.; Zhong, X.; Meng, X.; Yang, Q.; Liu, J.; Li, H.-Y. CD105 Promotes Hepatocarcinoma Cell Invasion and Metastasis through VEGF. Tumor Biol. 2014, 36, 737–745. [Google Scholar] [CrossRef]
- Kwon, Y.-C.; Bose, S.K.; Steele, R.; Meyer, K.; Di Bisceglie, A.M.; Ray, R.B.; Ray, R. Promotion of Cancer Stem-Like Cell Properties in Hepatitis C Virus-Infected Hepatocytes. J. Virol. 2015, 89, 11549–11556. [Google Scholar] [CrossRef] [Green Version]
- Mardomi, A.; Sabzichi, M.; Somi, M.H.; Shanehbandi, D.; Rahbarghazi, R.; Sanjarani, O.T.; Samadi, N. Trafficking Mechanism of Bone Marrow-Derived Mesenchymal Stem Cells toward Hepatocellular Carcinoma HepG2 Cells by Modulating Endoglin, CXCR4 and TGF-β. Cell. Mol. Boil. 2016, 62, 81–86. [Google Scholar]
- Yu, D.; Zhuang, L.; Sun, X.; Chen, J.; Yao, Y.; Meng, K.; Ding, Y. Particular Distribution and Expression Pattern of Endoglin (CD105) in the Liver of Patients with Hepatocellular Carcinoma. BMC Cancer 2007, 7, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- About, F.; Bibert, S.; Jouanguy, E.; Nalpas, B.; Lorenzo, L.; Rattina, V.; Zarhrate, M.; Hanein, S.; Munteanu, M.; Müllhaupt, B.; et al. Identification of an Endoglin Variant Associated With HCV-Related Liver Fibrosis Progression by Next-Generation Sequencing. Front. Genet. 2019, 10, 1024. [Google Scholar] [CrossRef] [Green Version]
- Meurer, S.; Wimmer, A.E.; Van De Leur, E.; Weiskirchen, R. Leur Endoglin Trafficking/Exosomal Targeting in Liver Cells Depends on N-Glycosylation. Cells 2019, 8, 997. [Google Scholar] [CrossRef] [Green Version]
- Gerrits, T.; Zandbergen, M.; Wolterbeek, R.; Bruijn, J.A.; Baelde, H.J.; Scharpfenecker, M. Endoglin Promotes Myofibroblast Differentiation and Extracellular Matrix Production in Diabetic Nephropathy. Int. J. Mol. Sci. 2020, 21, 7713. [Google Scholar] [CrossRef]
- Zhang, J.; Gu, C.; Song, Q.; Zhu, M.; Xu, Y.; Xiao, M.; Zheng, W. Identifying Cancer-Associated Fibroblasts as Emerging Targets for Hepatocellular Carcinoma. Cell Biosci. 2020, 10, 1–15. [Google Scholar] [CrossRef]
- Liu, Z.; Chen, M.; Zhao, R.; Huang, Y.; Liu, F.; Li, B.; Qin, Y. CAF-Induced Placental Growth Factor Facilitates Neoangiogenesis in Hepatocellular Carcinoma. Acta Biochim. Biophys. Sin. 2019, 52, 18–25. [Google Scholar] [CrossRef]
- Yang, Y.; Guan, Q.; Guo, L.; Han, C. The Prognostic Correlation between CD105 Expression Level in Tumor Tissue and Peripheral Blood and Sunitinib Administration in Advanced Hepatocellular Carcinoma. Cancer Biol. Ther. 2018, 19, 1006–1014. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, O.D.; Canedo, N.H.S.; Pannain, V.L. Immunohistochemical Angiogenic Biomarkers in Hepatocellular Carcinoma and Cirrhosis: Correlation with Pathological Features. Clinical 2016, 71, 639–643. [Google Scholar] [CrossRef]
- Zhong, L.; Zou, H.; Huang, Y.; Gong, W.; He, J.; Tan, J.; Lai, Z.; Li, Y.; Zhou, C.; Zhang, G.; et al. Magnetic Endoglin Aptamer Nanoprobe for Targeted Diagnosis of Solid Tumor. J. Biomed. Nanotechnol. 2019, 15, 352–362. [Google Scholar] [CrossRef] [PubMed]
- Duffy, A.; Ulahannan, S.V.; Cao, L.; Rahma, O.E.; Makarova-Rusher, O.V.; Kleiner, D.E.; Fioravanti, S.; Walker, M.D.; Carey, S.; Yu, Y.; et al. A Phase II Study of TRC105 in Patients with Hepatocellular Carcinoma Who Have Progressed on Sorafenib. United Eur. Gastroenterol. J. 2015, 3, 453–461. [Google Scholar] [CrossRef]
- Karmani, L.; Bouchat, V.; Bouzin, C.; Leveque, P.; LaBar, D.; Bol, A.; Deumer, G.; Marega, R.; Bonifazi, D.; Haufroid, V.; et al. 89Zr-Labeled Anti-endoglin Antibody-Targeted Gold Nanoparticles for Imaging Cancer: Implications for Future Cancer Therapy. Nanomedicine 2014, 9, 1923–1937. [Google Scholar] [CrossRef] [Green Version]
- Duan, C.-L.; Hou, G.-H.; Liu, Y.-P.; Liang, T.; Song, J.; Han, J.-K.; Zhang, C. Tumor Vascular Homing Endgolin-Targeted Radioimmunotherapy in Hepatocellular Carcinoma. Tumor Biol. 2014, 35, 12205–12215. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Paauwe, M.; Nixon, A.B.; Hawinkels, L.J. Endoglin Targeting: Lessons Learned and Questions That Remain. Int. J. Mol. Sci. 2020, 22, 147. [Google Scholar] [CrossRef]
- Schoonderwoerd, M.J.; Koops, M.F.; Angela, R.A.; Koolmoes, B.; Toitou, M.; Paauwe, M.; Barnhoorn, M.C.; Liu, Y.; Sier, C.F.; Hardwick, J.C.; et al. Targeting Endoglin-Expressing Regulatory T Cells in the Tumor Microenvironment Enhances the Effect of PD1 Checkpoint Inhibitor Immunotherapy. Clin. Cancer Res. 2020, 26, 3831–3842. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Trial Description | Condition or Disease | FDA Approval Status | NCT# |
|---|---|---|---|
| Sorafenib and | Hepatoma | Phase 1/2 | NCT01306058 |
| TRC105 in | Liver neoplasms | ||
| hepatocellular | Adenoma, liver | ||
| cancer | hepatocellular carcinoma | ||
| Liver neoplasms, experimental | |||
| TRC105 for | Hepatocellular carcinoma | Phase 2 | NCT01375569 |
| liver cancer | Hepatocellular cancer | ||
| that has not responded to sorafenib | Carcinoma, hepatocellular | ||
| Trial of TRC105 and sorafenib in patients with HCC | Hepatocellular carcinoma | Phase 1/2 | NCT02560779 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeng, K.-S.; Sheen, I.-S.; Lin, S.-S.; Leu, C.-M.; Chang, C.-F. The Role of Endoglin in Hepatocellular Carcinoma. Int. J. Mol. Sci. 2021, 22, 3208. https://doi.org/10.3390/ijms22063208
Jeng K-S, Sheen I-S, Lin S-S, Leu C-M, Chang C-F. The Role of Endoglin in Hepatocellular Carcinoma. International Journal of Molecular Sciences. 2021; 22(6):3208. https://doi.org/10.3390/ijms22063208
Chicago/Turabian StyleJeng, Kuo-Shyang, I-Shyan Sheen, Shu-Sheng Lin, Chuen-Miin Leu, and Chiung-Fang Chang. 2021. "The Role of Endoglin in Hepatocellular Carcinoma" International Journal of Molecular Sciences 22, no. 6: 3208. https://doi.org/10.3390/ijms22063208
APA StyleJeng, K. -S., Sheen, I. -S., Lin, S. -S., Leu, C. -M., & Chang, C. -F. (2021). The Role of Endoglin in Hepatocellular Carcinoma. International Journal of Molecular Sciences, 22(6), 3208. https://doi.org/10.3390/ijms22063208