
Emerging Molecular Connections between NM23 Proteins, Telomeres and Telomere-Associated Factors: Implications in Cancer Metastasis and Ageing

Abstract
:1. Introduction
2. Direct Interaction of NM23 Proteins with Telomeres and Telomere-Binding Proteins
2.1. NM23 Proteins Bind with the Telomeric DNA Sequence In Vitro
2.2. NM23-H2 Interaction with Telomere-Like G-Quadruplex Motif at the c-MYC Promoter
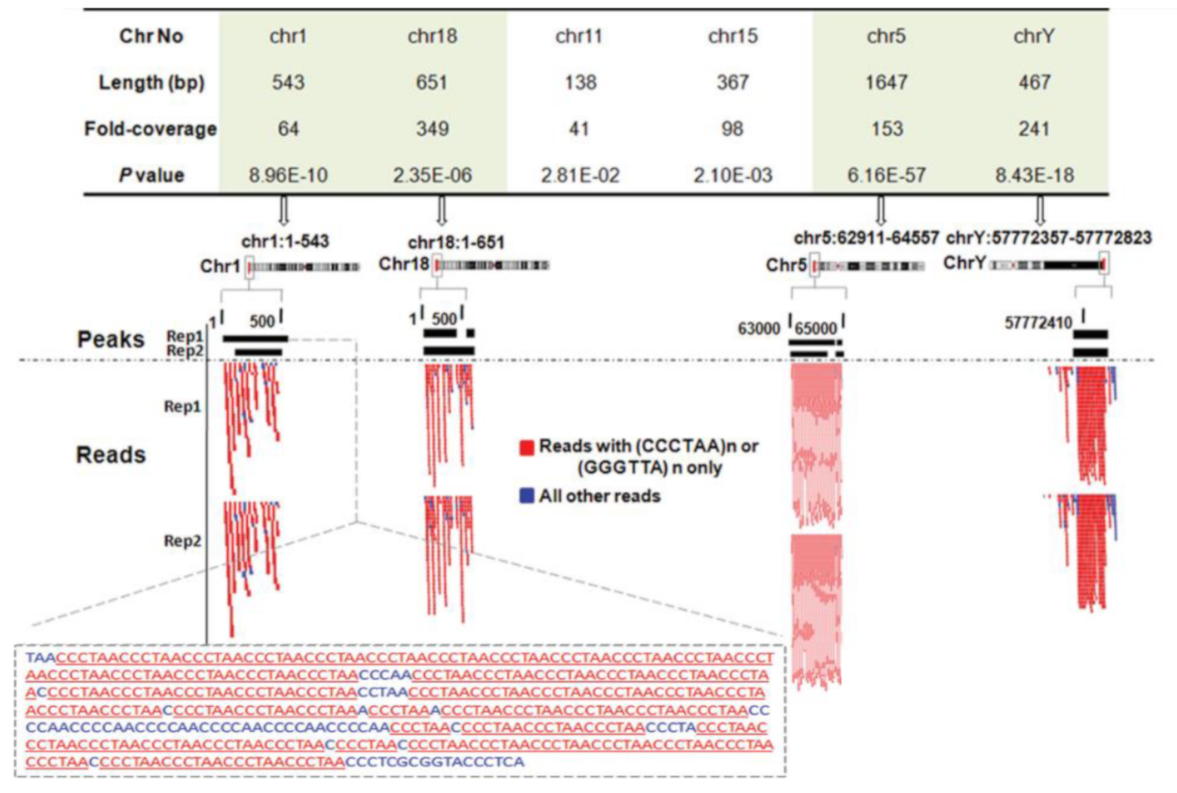
2.3. NM23-H2 Interacts with Telomeres Inside Cells
2.4. Interaction of the Telomere Binding Factor TRF1 and TRF2 with NM23-H2
2.5. Human Telomerase Physically Associates with NM23-H2
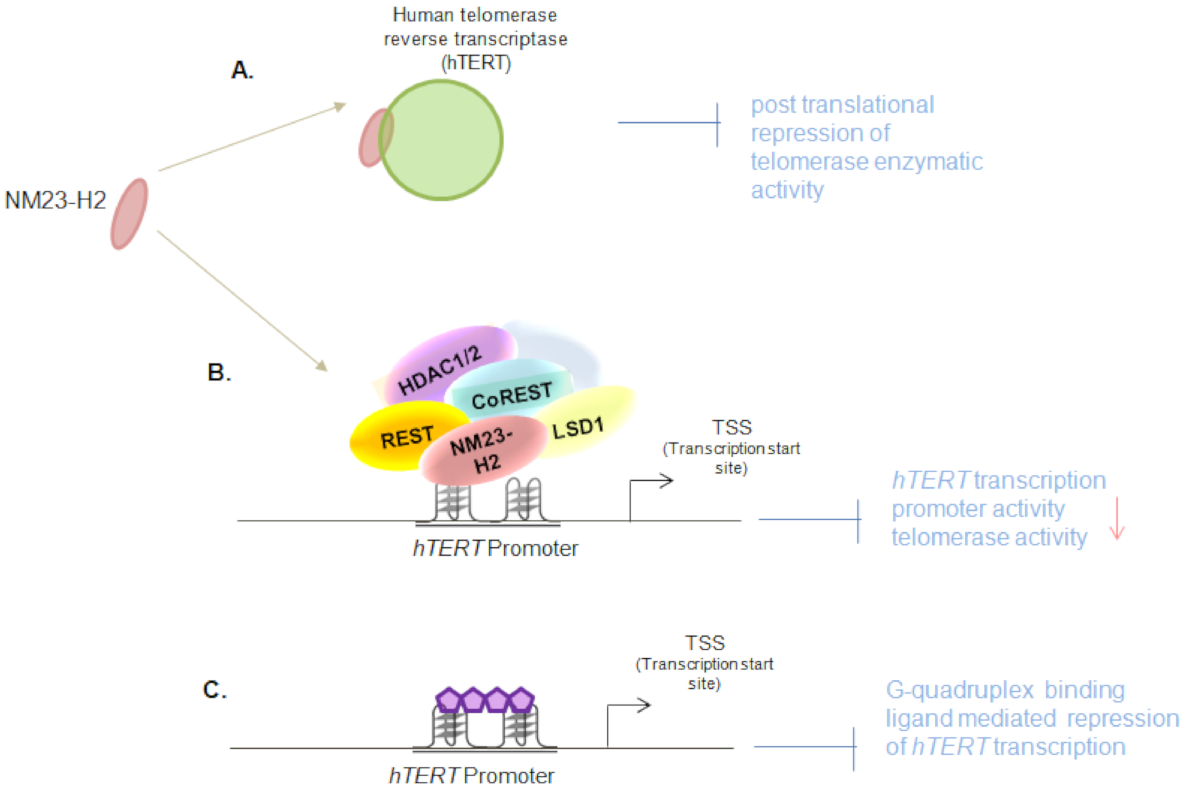
2.6. NM23-H2-Mediated Epigenetic Regulation of Human Telomerase Transcription
3. Putative Telomere Dependent and Independent Role of the NM23NDPK Variants in Ageing and Cancer
3.1. Potential Role of NM23-H4 in Regulation of Ageing
3.2. NM23-H1 Levels Alter during Ageing-Related Disorders
3.3. Cytoskeleton Modulation by NM23/NDPK Proteins in Cancer and Ageing
4. Role of NM23/NDPKs in T-Cell Activation—Potential Telomerase Connection
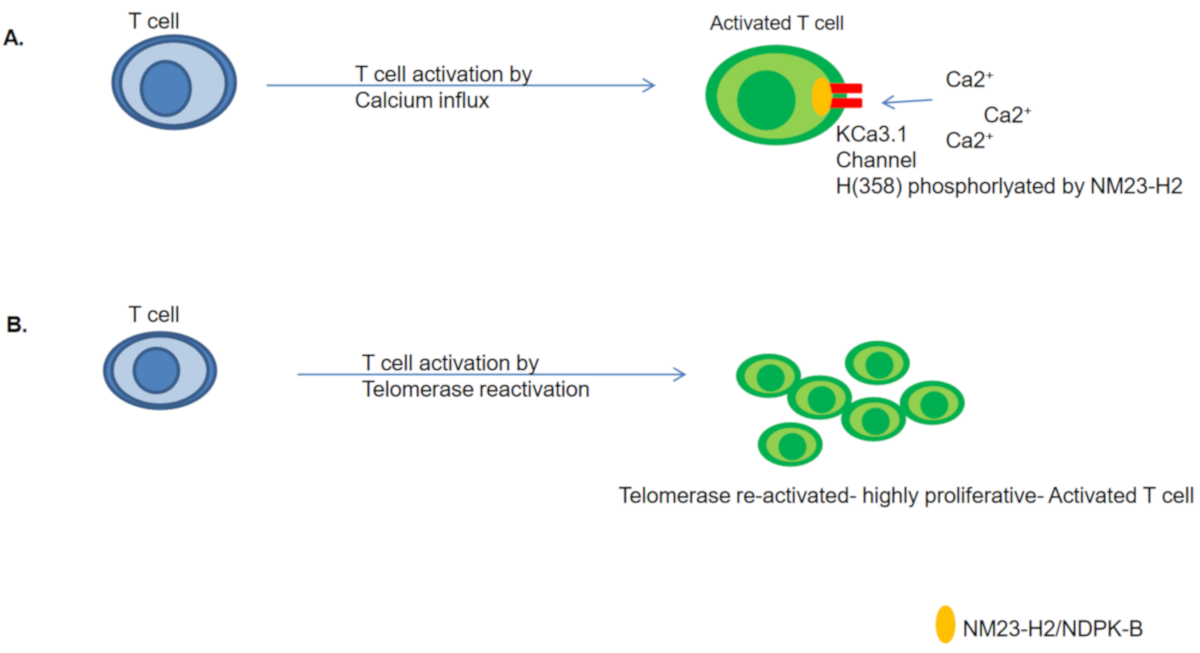
4.1. NM23-H2/NDPK-B Mediated T Cell Activation
4.2. Potential Role of NM23-H2 in Telomerase Re-Activation during T Cell Activation
5. Emerging Aspects and Future Questions
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Boissan, M.; Dabernat, S.; Peuchant, E.; Schlattner, U.; Lascu, I.; Lacombe, M.-L. The Mammalian Nm23/NDPK Family: From Metastasis Control to Cilia Movement. Mol. Cell. Biochem. 2009, 329, 51–62. [Google Scholar] [CrossRef]
- Sharma, S.; Sengupta, A.; Chowdhury, S. NM23/NDPK Proteins in Transcription Regulatory Functions and Chromatin Modulation: Emerging Trends. Lab. Investig. 2018, 98, 175–181. [Google Scholar] [CrossRef] [Green Version]
- Boissan, M.; Schlattner, U.; Lacombe, M.-L. The NDPK/NME Superfamily: State of the Art. Lab. Investig. 2018, 98, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Hartsough, M.T.; Steeg, P.S. Nm23/Nucleoside Diphosphate Kinase in Human Cancers. J. Bioenerg. Biomembr. 2000, 32, 301–308. [Google Scholar] [CrossRef]
- Kar, A.; Saha, D.; Purohit, G.; Singh, A.; Kumar, P.; Yadav, V.K.; Kumar, P.; Thakur, R.K.; Chowdhury, S. Metastases Suppressor NME2 Associates with Telomere Ends and Telomerase and Reduces Telomerase Activity within Cells. Nucleic Acids Res. 2012, 40, 2554–2565. [Google Scholar] [CrossRef] [Green Version]
- Nosaka, K.; Kawahara, M.; Masuda, M.; Satomi, Y.; Nishino, H. Association of Nucleoside Diphosphate Kinase Nm23-H2 with Human Telomeres. Biochem. Biophys. Res. Commun. 1998, 243, 342–348. [Google Scholar] [CrossRef]
- Lee, J.-S.; Lee, S.-H. Cloning and Characterization of CDNA Encoding Zebrafish Danio Rerio NM23-B Gene. Gene 2000, 245, 75–79. [Google Scholar] [CrossRef]
- Perina, D.; Korolija, M.; Mikoč, A.; Halasz, M.; Herak Bosnar, M.; Ćetković, H. Characterization of Nme5-Like Gene/Protein from the Red Alga Chondrus Crispus. Mar. Drugs 2019, 18, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Postel, E.H.; Berberich, S.J.; Flint, S.J.; Ferrone, C.A. Human C-Myc Transcription Factor PuF Identified as Nm23-H2 Nucleoside Diphosphate Kinase, a Candidate Suppressor of Tumor Metastasis. Science 1993, 261, 478–480. [Google Scholar] [CrossRef]
- Thakur, R.K.; Kumar, P.; Halder, K.; Verma, A.; Kar, A.; Parent, J.-L.; Basundra, R.; Kumar, A.; Chowdhury, S. Metastases Suppressor NM23-H2 Interaction with G-Quadruplex DNA within c-MYC Promoter Nuclease Hypersensitive Element Induces c-MYC Expression. Nucleic Acids Res. 2009, 37, 172–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sen, D.; Gilbert, W. Formation of Parallel Four-Stranded Complexes by Guanine-Rich Motifs in DNA and Its Implications for Meiosis. Nature 1988, 334, 364–366. [Google Scholar] [CrossRef] [PubMed]
- Saha, D.; Singh, A.; Hussain, T.; Srivastava, V.; Sengupta, S.; Kar, A.; Dhapola, P.; Dhople, V.; Ummanni, R.; Chowdhury, S. Epigenetic Suppression of Human Telomerase (HTERT) Is Mediated by the Metastasis Suppressor NME2 in a G-Quadruplex-Dependent Fashion. J. Biol. Chem. 2017, 292, 15205–15215. [Google Scholar] [CrossRef] [Green Version]
- Hildebrandt, M.; Lacombe, M.L.; Mesnildrey, S.; Véron, M. A Human NDP-Kinase B Specifically Binds Single-Stranded Poly-Pyrimidine Sequences. Nucleic Acids Res. 1995, 23, 3858–3864. [Google Scholar] [CrossRef] [Green Version]
- Sfeir, A.; de Lange, T. Removal of Shelterin Reveals the Telomere End-Protection Problem. Science 2012, 336, 593–597. [Google Scholar] [CrossRef] [Green Version]
- Roake, C.M.; Artandi, S.E. Regulation of Human Telomerase in Homeostasis and Disease. Nat. Rev. Mol. Cell Biol. 2020, 21, 384–397. [Google Scholar] [CrossRef]
- De Lange, T. Shelterin: The Protein Complex That Shapes and Safeguards Human Telomeres. Genes Dev. 2005, 19, 2100–2110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jafri, M.A.; Ansari, S.A.; Alqahtani, M.H.; Shay, J.W. Roles of Telomeres and Telomerase in Cancer, and Advances in Telomerase-Targeted Therapies. Genome Med. 2016, 8, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shay, J.W.; Wright, W.E. Role of Telomeres and Telomerase in Cancer. Semin. Cancer Biol. 2011, 21, 349–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagheri, S.; Nosrati, M.; Li, S.; Fong, S.; Torabian, S.; Rangel, J.; Moore, D.H.; Federman, S.; Laposa, R.R.; Baehner, F.L.; et al. Genes and Pathways Downstream of Telomerase in Melanoma Metastasis. Proc. Natl. Acad. Sci. USA 2006, 103, 11306–11311. [Google Scholar] [CrossRef] [Green Version]
- Saito, Y.; Kosugi, S.; Suda, T.; Wakabayashi, Y.; Mishima, Y.; Hatakeyama, K.; Kominami, R. Telomerase Activity and Metastasis: Expansion of Cells Having Higher Telomerase Activity within Culture Lines and Tumor Tissues. Jpn. J. Cancer Res. 1997, 88, 732–737. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.; Khoo, C.M.; Naylor, M.L.; Maser, R.S.; DePinho, R.A. Telomere-Based Crisis: Functional Differences between Telomerase Activation and ALT in Tumor Progression. Genes Dev. 2003, 17, 88–100. [Google Scholar] [CrossRef] [Green Version]
- Robinson, N.J.; Schiemann, W.P. Means to the Ends: The Role of Telomeres and Telomere Processing Machinery in Metastasis. Biochim. Biophys. Acta 2016, 1866, 320–329. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Marcelo, T.; Gómez, A.; Pascua, I.; de Juan, C.; Head, J.; Hernando, F.; Jarabo, J.-R.; Calatayud, J.; Torres-García, A.-J.; Iniesta, P. Telomere Length and Telomerase Activity in Non-Small Cell Lung Cancer Prognosis: Clinical Usefulness of a Specific Telomere Status. J. Exp. Clin. Cancer Res. 2015, 34, 78. [Google Scholar] [CrossRef] [Green Version]
- Viceconte, N.; Dheur, M.-S.; Majerova, E.; Pierreux, C.E.; Baurain, J.-F.; van Baren, N.; Decottignies, A. Highly Aggressive Metastatic Melanoma Cells Unable to Maintain Telomere Length. Cell Rep. 2017, 19, 2529–2543. [Google Scholar] [CrossRef] [Green Version]
- Bojovic, B.; Crowe, D.L. Dysfunctional Telomeres Promote Genomic Instability and Metastasis in the Absence of Telomerase Activity in Oncogene Induced Mammary Cancer. Mol. Carcinog. 2013, 52, 103–117. [Google Scholar] [CrossRef]
- Griffith, J.K.; Bryant, J.E.; Fordyce, C.A.; Gilliland, F.D.; Joste, N.E.; Moyzis, R.K. Reduced Telomere DNA Content Is Correlated with Genomic Instability and Metastasis in Invasive Human Breast Carcinoma. Breast Cancer Res. Treat. 1999, 54, 59–64. [Google Scholar] [CrossRef]
- Kar, A.; Chowdhury, S. Inhibition of Telomerase Activity by NME2: Impact on Metastasis Suppression? Naunyn Schmiedebergs Arch. Pharm. 2015, 388, 235–241. [Google Scholar] [CrossRef] [Green Version]
- Iizuka, N.; Mori, N.; Tamesa, T.; Tangoku, A.; Oka, M. Telomerase Activity and Nm23-H2 Protein Expression in Hepatocellular Carcinoma. Anticancer Res. 2003, 23, 43–47. [Google Scholar]
- Lee, M.-J.; Xu, D.-Y.; Li, H.; Yu, G.-R.; Leem, S.-H.; Chu, I.-S.; Kim, I.-H.; Kim, D.-G. Pro-Oncogenic Potential of NM23-H2 in Hepatocellular Carcinoma. Exp. Mol. Med. 2012, 44, 214–224. [Google Scholar] [CrossRef]
- Saharia, A.; Guittat, L.; Crocker, S.; Lim, A.; Steffen, M.; Kulkarni, S.; Stewart, S.A. Flap Endonuclease 1 Contributes to Telomere Stability. Curr. Biol. 2008, 18, 496–500. [Google Scholar] [CrossRef] [Green Version]
- Dechat, T.; Gajewski, A.; Korbei, B.; Gerlich, D.; Daigle, N.; Haraguchi, T.; Furukawa, K.; Ellenberg, J.; Foisner, R. LAP2alpha and BAF Transiently Localize to Telomeres and Specific Regions on Chromatin during Nuclear Assembly. J. Cell. Sci. 2004, 117, 6117–6128. [Google Scholar] [CrossRef] [Green Version]
- Gonzalo, S.; Eissenberg, J.C. Tying up Loose Ends: Telomeres, Genomic Instability and Lamins. Curr. Opin. Genet. Dev. 2016, 37, 109–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puts, G.S.; Leonard, M.K.; Pamidimukkala, N.V.; Snyder, D.E.; Kaetzel, D.M. Nuclear Functions of NME Proteins. Lab. Investig. 2018, 98, 211–218. [Google Scholar] [CrossRef]
- Kaetzel, D.M.; Zhang, Q.; Yang, M.; McCorkle, J.R.; Ma, D.; Craven, R.J. Potential Roles of 3’-5’ Exonuclease Activity of NM23-H1 in DNA Repair and Malignant Progression. J. Bioenerg. Biomembr. 2006, 38, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.K.; Sharma, S.; Chowdhury, S. Non-Duplex G-Quadruplex Structures Emerge as Mediators of Epigenetic Modifications. Trends Genet. 2019, 35, 129–144. [Google Scholar] [CrossRef] [Green Version]
- Sengupta, A.; Ganguly, A.; Chowdhury, S. Promise of G-Quadruplex Structure Binding Ligands as Epigenetic Modifiers with Anti-Cancer Effects. Molecules 2019, 24, 582. [Google Scholar] [CrossRef] [Green Version]
- Horn, S.; Figl, A.; Rachakonda, P.S.; Fischer, C.; Sucker, A.; Gast, A.; Kadel, S.; Moll, I.; Nagore, E.; Hemminki, K.; et al. TERT Promoter Mutations in Familial and Sporadic Melanoma. Science 2013, 339, 959–961. [Google Scholar] [CrossRef] [Green Version]
- Huang, F.W.; Hodis, E.; Xu, M.J.; Kryukov, G.V.; Chin, L.; Garraway, L.A. Highly Recurrent TERT Promoter Mutations in Human Melanoma. Science 2013, 339, 957–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.-J.; Cui, Y.; Yin, H.; Scheid, A.; Hendricks, W.P.D.; Schmidt, J.; Sekulic, A.; Kong, D.; Trent, J.M.; Gokhale, V.; et al. A Pharmacological Chaperone Molecule Induces Cancer Cell Death by Restoring Tertiary DNA Structures in Mutant HTERT Promoters. J. Am. Chem. Soc. 2016. [Google Scholar] [CrossRef] [Green Version]
- Schlattner, U.; Tokarska-Schlattner, M.; Ramirez, S.; Tyurina, Y.Y.; Amoscato, A.A.; Mohammadyani, D.; Huang, Z.; Jiang, J.; Yanamala, N.; Seffouh, A.; et al. Dual Function of Mitochondrial Nm23-H4 Protein in Phosphotransfer and Intermembrane Lipid Transfer: A Cardiolipin-Dependent Switch. J. Biol. Chem. 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kagan, V.E.; Jiang, J.; Huang, Z.; Tyurina, Y.Y.; Desbourdes, C.; Cottet-Rousselle, C.; Dar, H.H.; Verma, M.; Tyurin, V.A.; Kapralov, A.A.; et al. NDPK-D (NM23-H4)-Mediated Externalization of Cardiolipin Enables Elimination of Depolarized Mitochondria by Mitophagy. Cell Death Differ. 2016, 23, 1140–1151. [Google Scholar] [CrossRef] [Green Version]
- Schlattner, U.; Tokarska-Schlattner, M.; Epand, R.M.; Boissan, M.; Lacombe, M.L.; Kagan, V.E. NME4/Nucleoside Diphosphate Kinase D in Cardiolipin Signaling and Mitophagy. Lab. Investig. 2018, 98, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.T.; Ji, J.; Dagda, R.K.; Jiang, J.F.; Tyurina, Y.Y.; Kapralov, A.A.; Tyurin, V.A.; Yanamala, N.; Shrivastava, I.H.; Mohammadyani, D.; et al. Cardiolipin Externalization to the Outer Mitochondrial Membrane Acts as an Elimination Signal for Mitophagy in Neuronal Cells. Nat. Cell Biol. 2013, 15, 1197–1205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Q.; Jeong, Y.Y. Mitophagy in Alzheimer’s Disease and Other Age-Related Neurodegenerative Diseases. Cells 2020, 9, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujita, Y.; Fujiwara, K.; Zenitani, S.; Yamashita, T. Acetylation of NDPK-D Regulates Its Subcellular Localization and Cell Survival. PLoS ONE 2015, 10, e0139616. [Google Scholar] [CrossRef] [Green Version]
- Miller, C.; Wang, L.; Ostergaard, E.; Dan, P.; Saada, A. The Interplay between SUCLA2, SUCLG2, and Mitochondrial DNA Depletion. Biochim. Biophys. Acta Mol. Basis Dis. 2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Tian, J.; Sui, S.; Yuan, X.; Chen, H.; Qu, C.; Du, Y.; Guo, L.; Du, H. Loss of Succinyl-CoA Synthase ADP-Forming β Subunit Disrupts MtDNA Stability and Mitochondrial Dynamics in Neurons. Sci. Rep. 2017. [Google Scholar] [CrossRef] [Green Version]
- Ishijima, Y.; Shimada, N.; Fukuda, M.; Miyazaki, H.; Orlov, N.Y.; Orlova, T.G.; Yamada, T.; Kimura, N. Overexpression of Nucleoside Diphosphate Kinases Induces Neurite Outgrowth and Their Substitution to Inactive Forms Leads to Suppression of Nerve Growth Factor- and Dibutyryl Cyclic AMP-Induced Effects in PC12D Cells. FEBS Lett. 1999, 445, 155–159. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Fountoulakis, M.; Cairns, N.J.; Lubec, G. Human Brain Nucleoside Diphosphate Kinase Activity Is Decreased in Alzheimer’s Disease and Down Syndrome. Biochem. Biophys. Res. Commun. 2002, 296, 970–975. [Google Scholar] [CrossRef]
- Lionello, M.; Blandamura, S.; Lovato, A.; Franchella, S.; Giacomelli, L.; Ottaviano, G.; Stellini, E.; Staffieri, A.; Marioni, G. A High Nuclear Nm23-H1 Expression Is Associated with a Better Prognosis in Elderly Patients with Laryngeal Carcinoma. Acta Otolaryngol. 2013, 133, 874–880. [Google Scholar] [CrossRef] [PubMed]
- Kounakis, K.; Tavernarakis, N. The Cytoskeleton as a Modulator of Aging and Neurodegeneration. Adv. Exp. Med. Biol. 2019, 1178, 227–245. [Google Scholar] [PubMed]
- Fife, C.M.; McCarroll, J.A.; Kavallaris, M. Movers and Shakers: Cell Cytoskeleton in Cancer Metastasis: Cytoskeleton and Cancer Metastasis. Br. J. Pharm. 2014, 171, 5507–5523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snider, N.T.; Altshuler, P.J.; Omary, M.B. Modulation of Cytoskeletal Dynamics by Mammalian Nucleoside Diphosphate Kinase (NDPK) Proteins. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2015. [Google Scholar] [CrossRef] [Green Version]
- Marino, N.; Marshall, J.-C.; Collins, J.W.; Zhou, M.; Qian, Y.; Veenstra, T.; Steeg, P.S. Nm23-H1 Binds to Gelsolin and Inactivates Its Actin-Severing Capacity to Promote Tumor Cell Motility and Metastasis. Cancer Res. 2013, 73, 5949–5962. [Google Scholar] [CrossRef] [Green Version]
- Boissan, M.; Montagnac, G.; Shen, Q.; Griparic, L.; Guitton, J.; Romao, M.; Sauvonnet, N.; Lagache, T.; Lascu, I.; Raposo, G.; et al. Membrane Trafficking. Nucleoside Diphosphate Kinases Fuel Dynamin Superfamily Proteins with GTP for Membrane Remodeling. Science 2014, 344, 1510–1515. [Google Scholar] [CrossRef] [Green Version]
- Conery, A.R.; Sever, S.; Harlow, E. Nucleoside Diphosphate Kinase Nm23-H1 Regulates Chromosomal Stability by Activating the GTPase Dynamin during Cytokinesis. Proc. Natl. Acad. Sci. USA 2010, 107, 15461–15466. [Google Scholar] [CrossRef] [Green Version]
- Baker, N.M.; Zeitlin, S.G.; Shi, L.Z.; Shah, J.; Berns, M.W. Chromosome Tips Damaged in Anaphase Inhibit Cytokinesis. PLoS ONE 2010, 5, e12398. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, J.; Benavente, R.; Hodzic, D.; Höög, C.; Stewart, C.L.; Alsheimer, M. Transmembrane Protein Sun2 Is Involved in Tethering Mammalian Meiotic Telomeres to the Nuclear Envelope. Proc. Natl. Acad. Sci. USA 2007, 104, 7426–7431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burla, R.; La Torre, M.; Saggio, I. Mammalian Telomeres and Their Partnership with Lamins. Nucleus 2016, 7, 187–202. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Wang, J.; Chan, K.M.; Tjia, W.M.; Deng, W.; Guan, X.; Huang, J.; Li, K.M.; Chau, P.Y.; Chen, D.J.; et al. Genomic Instability in Laminopathy-Based Premature Aging. Nat. Med. 2005, 11, 780–785. [Google Scholar] [CrossRef]
- Andersen, S.S. Spindle Assembly and the Art of Regulating Microtubule Dynamics by MAPs and Stathmin/Op18. Trends Cell Biol. 2000, 10, 261–267. [Google Scholar] [CrossRef]
- Cheon, M.S.; Fountoulakis, M.; Cairns, N.J.; Dierssen, M.; Herkner, K.; Lubec, G. Decreased Protein Levels of Stathmin in Adult Brains with Down Syndrome and Alzheimer’s Disease. J. Neural Transm. Suppl. 2001, 281–288. [Google Scholar] [CrossRef]
- Dumont, F.J.; Staruch, M.J.; Koprak, S.L.; Melino, M.R.; Sigal, N.H. Distinct Mechanisms of Suppression of Murine T Cell Activation by the Related Macrolides FK-506 and Rapamycin. J. Immunol. 1990, 144, 251–258. [Google Scholar]
- Chen, L.; Flies, D.B. Molecular Mechanisms of T Cell Co-Stimulation and Co-Inhibition. Nat. Rev. Immunol. 2013, 13, 227–242. [Google Scholar] [CrossRef] [PubMed]
- Siller-Farfán, J.A.; Dushek, O. Molecular Mechanisms of T Cell Sensitivity to Antigen. Immunol. Rev. 2018, 285, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Chatila, T.; Silverman, L.; Miller, R.; Geha, R. Mechanisms of T Cell Activation by the Calcium Ionophore Ionomycin. J. Immunol. 1989, 143, 1283–1289. [Google Scholar]
- Srivastava, S.; Li, Z.; Ko, K.; Choudhury, P.; Albaqumi, M.; Johnson, A.K.; Yan, Y.; Backer, J.M.; Unutmaz, D.; Coetzee, W.A.; et al. Histidine Phosphorylation of the Potassium Channel KCa3.1 by Nucleoside Diphosphate Kinase B Is Required for Activation of KCa3.1 and CD4 T Cells. Mol. Cell 2006, 24, 665–675. [Google Scholar] [CrossRef]
- Di, L.; Srivastava, S.; Zhdanova, O.; Sun, Y.; Li, Z.; Skolnik, E.Y. Nucleoside Diphosphate Kinase B Knock-out Mice Have Impaired Activation of the K+ Channel KCa3.1, Resulting in Defective T Cell Activation. J. Biol. Chem. 2010, 285, 38765–38771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, S.; Zhdanova, O.; Di, L.; Li, Z.; Albaqumi, M.; Wulff, H.; Skolnik, E.Y. Protein Histidine Phosphatase 1 Negatively Regulates CD4 T Cells by Inhibiting the K+ Channel KCa3.1. Proc. Natl. Acad. Sci. USA 2008, 105, 14442–14446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Son, N.H.; Murray, S.; Yanovski, J.; Hodes, R.J.; Weng, N. Lineage-Specific Telomere Shortening and Unaltered Capacity for Telomerase Expression in Human T and B Lymphocytes with Age. J. Immunol. 2000, 165, 1191–1196. [Google Scholar] [CrossRef] [Green Version]
- Weng, N.P.; Levine, B.L.; June, C.H.; Hodes, R.J. Regulated Expression of Telomerase Activity in Human T Lymphocyte Development and Activation. J. Exp. Med. 1996, 183, 2471–2479. [Google Scholar] [CrossRef]
- Huang, E.E.; Tedone, E.; O’Hara, R.; Cornelius, C.; Lai, T.-P.; Ludlow, A.; Wright, W.E.; Shay, J.W. The Maintenance of Telomere Length in CD28+ T Cells During T Lymphocyte Stimulation. Sci. Rep. 2017, 7, 6785. [Google Scholar] [CrossRef] [Green Version]
- Bodnar, A.G.; Kim, N.W.; Effros, R.B.; Chiu, C.P. Mechanism of Telomerase Induction during T Cell Activation. Exp. Cell Res. 1996, 228, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Kaban, A.P.; Gunina, L.M. Use of the indices of the kallikrein-kinin system of the blood in evaluating the efficacy of the prevention and treatment of postoperative purulent complications in oncologic patients. Klin. Khir. 1989, 10–12. [Google Scholar]
- Sawai, Y.; Sawasaki, Y.; Tsukada, K. Ribonuclease H Activity in Developing Rat Brain. Life Sci. 1977, 21, 1351–1356. [Google Scholar] [CrossRef]
- Patrick, M.S.; Cheng, N.-L.; Kim, J.; An, J.; Dong, F.; Yang, Q.; Zou, I.; Weng, N.-P. Human T Cell Differentiation Negatively Regulates Telomerase Expression Resulting in Reduced Activation-Induced Proliferation and Survival. Front. Immunol. 2019, 10, 1993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tedone, E.; Huang, E.; O’Hara, R.; Batten, K.; Ludlow, A.T.; Lai, T.-P.; Arosio, B.; Mari, D.; Wright, W.E.; Shay, J.W. Telomere Length and Telomerase Activity in T Cells Are Biomarkers of High-Performing Centenarians. Aging Cell 2019, 18, e12859. [Google Scholar] [CrossRef]
- Najarro, K.; Nguyen, H.; Chen, G.; Xu, M.; Alcorta, S.; Yao, X.; Zukley, L.; Metter, E.J.; Truong, T.; Lin, Y.; et al. Telomere Length as an Indicator of the Robustness of B- and T-Cell Response to Influenza in Older Adults. J. Infect. Dis. 2015, 212, 1261–1269. [Google Scholar] [CrossRef]
- O’Callaghan, N.J.; Bull, C.; Fenech, M. Elevated Plasma Magnesium and Calcium May Be Associated with Shorter Telomeres in Older South Australian Women. J. Nutr. Health Aging 2014, 18, 131–136. [Google Scholar] [CrossRef]
- Akbar, A.N.; Vukmanovic-Stejic, M. Telomerase in T Lymphocytes: Use It and Lose It? J. Immunol. 2007, 178, 6689–6694. [Google Scholar] [CrossRef] [Green Version]
- Weng, N.P.; Hathcock, K.S.; Hodes, R.J. Regulation of Telomere Length and Telomerase in T and B Cells: A Mechanism for Maintaining Replicative Potential. Immunity 1998, 9, 151–157. [Google Scholar] [CrossRef] [Green Version]
- Patrick, M.; Weng, N.-P. Expression and Regulation of Telomerase in Human T Cell Differentiation, Activation, Aging and Diseases. Cell Immunol. 2019, 345, 103989. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, B.; Garinis, G.A.; Hoeijmakers, J.H.J. Age to Survive: DNA Damage and Aging. Trends Genet. 2008, 24, 77–85. [Google Scholar] [CrossRef]
- Li, J.S.Z.; Miralles Fusté, J.; Simavorian, T.; Bartocci, C.; Tsai, J.; Karlseder, J.; Lazzerini Denchi, E. TZAP: A Telomere-Associated Protein Involved in Telomere Length Control. Science 2017, 355, 638–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarek, G.; Kotsantis, P.; Ruis, P.; Van Ly, D.; Margalef, P.; Borel, V.; Zheng, X.-F.; Flynn, H.R.; Snijders, A.P.; Chowdhury, D.; et al. CDK Phosphorylation of TRF2 Controls T-Loop Dynamics during the Cell Cycle. Nature 2019, 575, 523–527. [Google Scholar] [CrossRef] [PubMed]
- Wojtyla, A.; Gladych, M.; Rubis, B. Human Telomerase Activity Regulation. Mol. Biol. Rep. 2011, 38, 3339–3349. [Google Scholar] [CrossRef] [Green Version]
- Walker, J.R.; Zhu, X.-D. Post-Translational Modifications of TRF1 and TRF2 and Their Roles in Telomere Maintenance. Mech Ageing Dev. 2012, 133, 421–434. [Google Scholar] [CrossRef] [PubMed]
- Shin, W.H.; Chung, K.C. Human Telomerase Reverse Transcriptase Positively Regulates Mitophagy by Inhibiting the Processing and Cytoplasmic Release of Mitochondrial PINK1. Cell Death Dis. 2020, 11, 425. [Google Scholar] [CrossRef]
- Milon, L.; Meyer, P.; Chiadmi, M.; Munier, A.; Johansson, M.; Karlsson, A.; Lascu, I.; Capeau, J.; Janin, J.; Lacombe, M.L. The Human Nm23-H4 Gene Product Is a Mitochondrial Nucleoside Diphosphate Kinase. J. Biol. Chem. 2000, 275, 14264–14272. [Google Scholar] [CrossRef] [Green Version]
- Saretzki, G. Extra-Telomeric Functions of Human Telomerase: Cancer, Mitochondria and Oxidative Stress. Curr. Pharm. Des. 2014, 20, 6386–6403. [Google Scholar] [CrossRef]
- Chiodi, I.; Mondello, C. Telomere-Independent Functions of Telomerase in Nuclei, Cytoplasm, and Mitochondria. Front. Oncol. 2012, 2, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, A.K.; Sharma, S.; Sengupta, S.; Saha, D.; Kumar, P.; Hussain, T.; Srivastava, V.; Roy, S.D.; Shay, J.W.; Chowdhury, S. Telomere Length-Dependent Transcription and Epigenetic Modifications in Promoters Remote from Telomere Ends. PLoS Genet. 2018, 14, e1007782. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.K.; Sharma, S.; Bagri, S.; Kutum, R.; Kumar, P.; Hussain, A.; Singh, P.; Saha, D.; Kar, A.; Dash, D.; et al. Telomere Repeat–Binding Factor 2 Binds Extensively to Extra-Telomeric G-Quadruplexes and Regulates the Epigenetic Status of Several Gene Promoters. J. Biol. Chem. 2019, 294, 17709–17722. [Google Scholar] [CrossRef] [Green Version]
- Pedroso, I.M.; Hayward, W.; Fletcher, T.M. The Effect of the TRF2 N-Terminal and TRFH Regions on Telomeric G-Quadruplex Structures. Nucleic Acids Res. 2009, 37, 1541–1554. [Google Scholar] [CrossRef] [Green Version]
- Biffi, G.; Tannahill, D.; Balasubramanian, S. An Intramolecular G-Quadruplex Structure Is Required for Binding of Telomeric Repeat-Containing RNA to the Telomeric Protein TRF2. J. Am. Chem. Soc. 2012, 134, 11974–11976. [Google Scholar] [CrossRef]
- Sharma, S.; Mukherjee, A.K.; Roy, S.S.; Bagri, S.; Lier, S.; Verma, M.; Sengupta, A.; Kumar, M.; Nesse, G.; Pandey, D.P.; et al. Human Telomerase Expression Is under Direct Transcriptional Control of the Telomere-Binding-Factor TRF2. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Wood, A.M.; Rendtlew Danielsen, J.M.; Lucas, C.A.; Rice, E.L.; Scalzo, D.; Shimi, T.; Goldman, R.D.; Smith, E.D.; Le Beau, M.M.; Kosak, S.T. TRF2 and Lamin A/C Interact to Facilitate the Functional Organization of Chromosome Ends. Nat. Commun. 2014, 5, 5467. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.D.; Garza-Gongora, A.G.; MacQuarrie, K.L.; Kosak, S.T. Interstitial Telomeric Loops and Implications of the Interaction between TRF2 and Lamin A/C. Differentiation 2018, 102, 19–26. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| NM23 Protein | Telomere Binding | Interaction with Telomere Associated Protein | Implication in Ageing and Cancer |
|---|---|---|---|
| NM23-H1 | ✕ [6] | ✕ [6] | √ [48,49,50,61,62] |
| NM23-H2 | √ [5,6] | √ [5,12,30,31,32] | √ [12,61] |
| NM23-H4 | - | - | [46,47] |
| NM23-H2 homolog, NM23B | √ [7] | - | - |
| NM23-H5 homolog, NME5-like Cc | √ [8] | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, S.; Sengupta, A.; Chowdhury, S. Emerging Molecular Connections between NM23 Proteins, Telomeres and Telomere-Associated Factors: Implications in Cancer Metastasis and Ageing. Int. J. Mol. Sci. 2021, 22, 3457. https://doi.org/10.3390/ijms22073457
Sharma S, Sengupta A, Chowdhury S. Emerging Molecular Connections between NM23 Proteins, Telomeres and Telomere-Associated Factors: Implications in Cancer Metastasis and Ageing. International Journal of Molecular Sciences. 2021; 22(7):3457. https://doi.org/10.3390/ijms22073457
Chicago/Turabian StyleSharma, Shalu, Antara Sengupta, and Shantanu Chowdhury. 2021. "Emerging Molecular Connections between NM23 Proteins, Telomeres and Telomere-Associated Factors: Implications in Cancer Metastasis and Ageing" International Journal of Molecular Sciences 22, no. 7: 3457. https://doi.org/10.3390/ijms22073457
APA StyleSharma, S., Sengupta, A., & Chowdhury, S. (2021). Emerging Molecular Connections between NM23 Proteins, Telomeres and Telomere-Associated Factors: Implications in Cancer Metastasis and Ageing. International Journal of Molecular Sciences, 22(7), 3457. https://doi.org/10.3390/ijms22073457