
Clinical Application of Novel Therapies for Coronary Angiogenesis: Overview, Challenges, and Prospects

 ,
,
Abstract
:1. Introduction
2. Structure of the Vasculature
3. Mechanisms of Vascularization
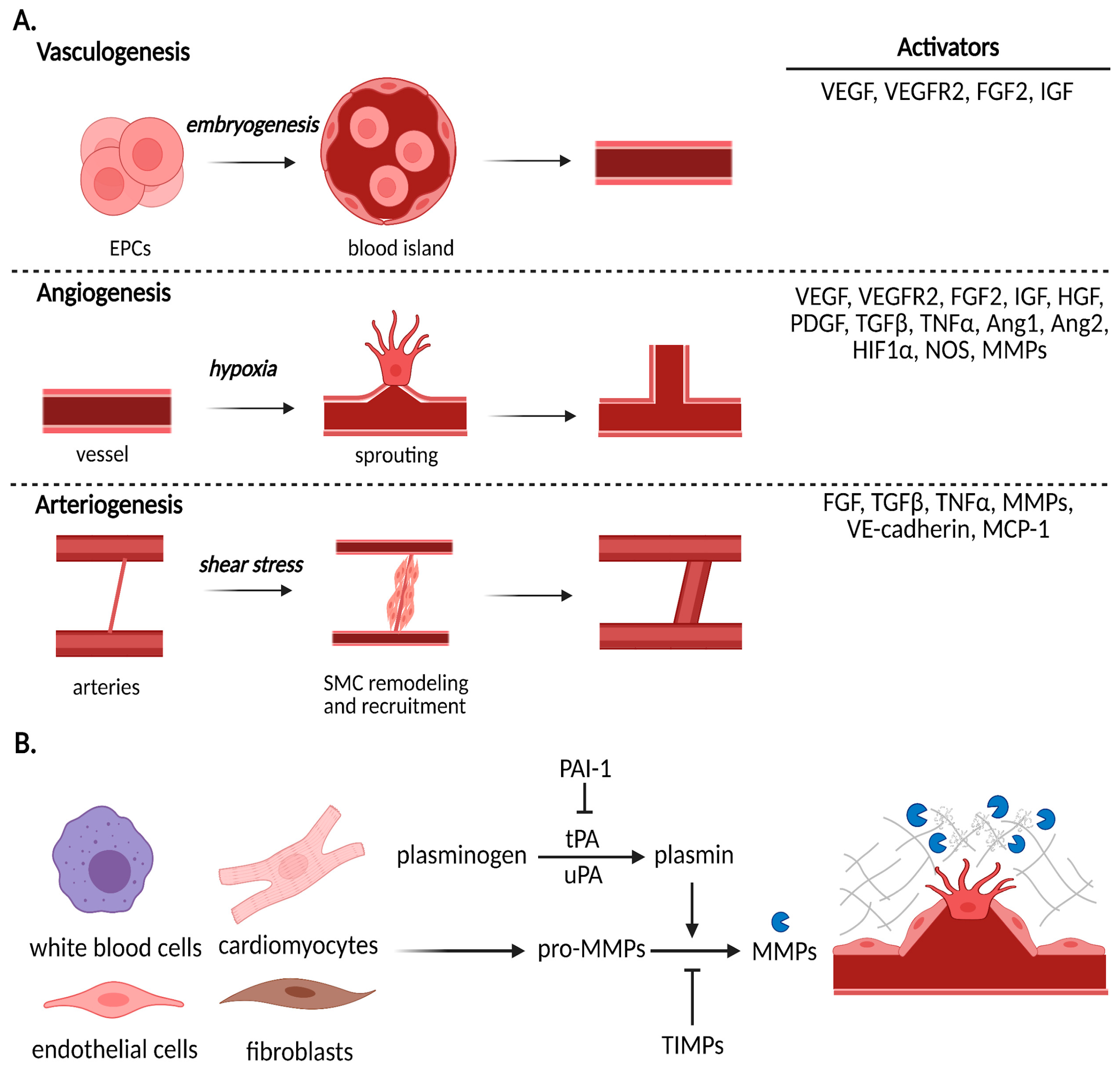
3.1. Vasculogenesis
3.2. Angiogenesis
3.3. Arteriogenesis
4. Preclinical Studies and Clinical Trials
4.1. Protein Therapy
4.2. Gene Therapy
4.3. Stem Cell Therapy
4.4. Extracellular Vesicle Therapy
5. Future of Therapeutic Angiogenesis
5.1. Patient Selection
5.2. Comorbidities
5.3. Combination Therapies
5.4. Delivery
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Roth, G.A.; Johnson, C.; Abajobir, A.; Abd-Allah, F.; Abera, S.F.; Abyu, G.; Ahmed, M.; Aksut, B.; Alam, T.; Alam, K.; et al. Global, Regional, and National Burden of Cardiovascular Diseases for 10 Causes, 1990 to 2015. J. Am. Coll. Cardiol. 2017, 70, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Folkman, J. Tumor angiogenesis: Therapeutic implications. N. Engl. J. Med. 1971, 285, 1182–1186. [Google Scholar] [CrossRef] [PubMed]
- Harada, K.; Friedman, M.; Lopez, J.J.; Wang, S.Y.; Li, J.; Prasad, P.V.; Pearlman, J.D.; Edelman, E.R.; Sellke, F.W.; Simons, M. Vascular endothelial growth factor administration in chronic myocardial ischemia. Am. J. Physiol. Circ. Physiol. 1996, 270, H1791–H1802. [Google Scholar] [CrossRef]
- Sellke, F.W.; Laham, R.J.; Edelman, E.R.; Pearlman, J.D.; Simons, M. Therapeutic angiogenesis with basic fibroblast growth factor: Technique and early results. Ann. Thorac. Surg. 1998, 65, 1540–1544. [Google Scholar] [CrossRef]
- Taimeh, Z.; Loughran, J.; Birks, E.J.; Bolli, R. Vascular endothelial growth factor in heart failure. Nat. Rev. Cardiol. 2013, 10, 519–530. [Google Scholar] [CrossRef]
- Buschmann, I.; Schaper, W. Arteriogenesis Versus Angiogenesis: Two Mechanisms of Vessel Growth. Physiology 1999, 14, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Conway, E.M.; Collen, D.; Carmeliet, P. Molecular mechanisms of blood vessel growth. Cardiovasc. Res. 2001, 49, 507–521. [Google Scholar] [CrossRef] [Green Version]
- Patan, S. Vasculogenesis and Angiogenesis. In BT—Angiogenesis in Brain Tumors; Kirsch, M., Black, P.M., Eds.; Springer: Boston, MA, USA, 2004; pp. 3–32. ISBN 978-1-4419-8871-3. [Google Scholar]
- Ribatti, D.; Vacca, A.; Nico, B.; Roncali, L.; Dammacco, F. Postnatal vasculogenesis. Mech. Dev. 2001, 100, 157–163. [Google Scholar] [CrossRef]
- Kimura, H.; Weisz, A.; Kurashima, Y.; Hashimoto, K.; Ogura, T.; D’Acquisto, F.; Addeo, R.; Makuuchi, M.; Esumi, H. Hypoxia response element of the human vascular endothelial growth factor gene mediates transcriptional regulation by nitric oxide: Control of hypoxia-inducible factor-1 activity by nitric oxide. Blood 2000, 95, 189–197. [Google Scholar] [CrossRef]
- Nelson, A.R.; Fingleton, B.; Rothenberg, M.L.; Matrisian, L.M. Matrix metalloproteinases: Biologic activity and clinical implications. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2000, 18, 1135–1149. [Google Scholar] [CrossRef] [PubMed]
- Brew, K.; Dinakarpandian, D.; Nagase, H. Tissue inhibitors of metalloproteinases: Evolution, structure and function. Biochim. Biophys. Acta 2000, 1477, 267–283. [Google Scholar] [CrossRef]
- Iruela-Arispe, M.L.; Davis, G.E. Cellular and Molecular Mechanisms of Vascular Lumen Formation. Dev. Cell 2009, 16, 222–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmeliet, P. Developmental biology. Controlling the cellular brakes. Nature 1999, 401, 657–658. [Google Scholar] [CrossRef]
- Cook, D.R.; Doumit, M.E.; Merkel, R.A. Transforming growth factor-beta, basic fibroblast growth factor, and platelet-derived growth factor-BB interact to affect proliferation of clonally derived porcine satellite cells. J. Cell. Physiol. 1993, 157, 307–312. [Google Scholar] [CrossRef]
- Cai, W.; Schaper, W. Mechanisms of arteriogenesis. Acta Biochim. Biophys. Sin. 2008, 40, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Meier, P.; Seiler, C. The coronary collateral circulation--past, present and future. Curr. Cardiol. Rev. 2014, 10, 1. [Google Scholar] [CrossRef] [Green Version]
- Lin, Q.; Lu, J.; Yanagisawa, H.; Webb, R.; Lyons, G.E.; Richardson, J.A.; Olson, E.N. Requirement of the MADS-box transcription factor MEF2C for vascular development. Development 1998, 125, 4565–4574. [Google Scholar]
- Olsson, A.-K.; Dimberg, A.; Kreuger, J.; Claesson-Welsh, L. VEGF receptor signalling—In control of vascular function. Nat. Rev. Mol. Cell Biol. 2006, 7, 359–371. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ibrahimi, O.A.; Olsen, S.K.; Umemori, H.; Mohammadi, M.; Ornitz, D.M. Receptor specificity of the fibroblast growth factor family. The complete mammalian FGF family. J. Biol. Chem. 2006, 281, 15694–15700. [Google Scholar] [CrossRef] [Green Version]
- Zymek, P.; Bujak, M.; Chatila, K.; Cieslak, A.; Thakker, G.; Entman, M.L.; Frangogiannis, N.G. The role of platelet-derived growth factor signaling in healing myocardial infarcts. J. Am. Coll. Cardiol. 2006, 48, 2315–2323. [Google Scholar] [CrossRef] [Green Version]
- Shyu, K.G.; Manor, O.; Magner, M.; Yancopoulos, G.D.; Isner, J.M. Direct intramuscular injection of plasmid DNA encoding angiopoietin-1 but not angiopoietin-2 augments revascularization in the rabbit ischemic hindlimb. Circulation 1998, 98, 2081–2087. [Google Scholar] [CrossRef]
- Yang, Z.; Chen, B.; Sheng, Z.; Zhang, D.; Jia, E.; Wang, W.; Ma, D.; Zhu, T.; Wang, L.; Li, C.; et al. Improvement of heart function in postinfarct heart failure swine models after hepatocyte growth factor gene transfer: Comparison of low-, medium- and high-dose groups. Mol. Biol. Rep. 2010, 37, 2075–2081. [Google Scholar] [CrossRef]
- Rong, S.; Wang, X.; Wang, Y.; Wu, H.; Zhou, X.; Wang, Z.; Wang, Y.; Xue, C.; Li, B.; Gao, D. Anti-inflammatory activities of hepatocyte growth factor in post-ischemic heart failure. Acta Pharmacol. Sin. 2018, 39, 1613–1621. [Google Scholar] [CrossRef] [Green Version]
- Henry, T.D.; Annex, B.H.; McKendall, G.R.; Azrin, M.A.; Lopez, J.J.; Giordano, F.J.; Shah, P.K.; Willerson, J.T.; Benza, R.L.; Berman, D.S.; et al. The VIVA trial: Vascular endothelial growth factor in Ischemia for Vascular Angiogenesis. Circulation 2003, 107, 1359–1365. [Google Scholar] [CrossRef]
- Simons, M.; Annex, B.H.; Laham, R.J.; Kleiman, N.; Henry, T.; Dauerman, H.; Udelson, J.E.; Gervino, E.V.; Pike, M.; Whitehouse, M.J.; et al. Pharmacological treatment of coronary artery disease with recombinant fibroblast growth factor-2: Double-blind, randomized, controlled clinical trial. Circulation 2002, 105, 788–793. [Google Scholar] [CrossRef]
- Ruel, M.; Beanlands, R.S.; Lortie, M.; Chan, V.; Camack, N.; deKemp, R.A.; Suuronen, E.J.; Rubens, F.D.; DaSilva, J.N.; Sellke, F.W.; et al. Concomitant treatment with oral L-arginine improves the efficacy of surgical angiogenesis in patients with severe diffuse coronary artery disease: The Endothelial Modulation in Angiogenic Therapy randomized controlled trial. J. Thorac. Cardiovasc. Surg. 2008, 135, 762–770.e1. [Google Scholar] [CrossRef] [Green Version]
- Lathi, K.G.; Cespedes, R.M.; Losordo, D.W.; Vale, P.R.; Symes, J.F.; Isner, J.M. Direct intramyocardial gene therapy with vegf for inoperable coronary artery disease: Preliminary clinical results. Anesth. Analg. 1999, 88, 73SCA. [Google Scholar] [CrossRef]
- Hartikainen, J.; Hassinen, I.; Hedman, A.; Kivelä, A.; Saraste, A.; Knuuti, J.; Husso, M.; Mussalo, H.; Hedman, M.; Rissanen, T.T.; et al. Adenoviral intramyocardial VEGF-DΔNΔC gene transfer increases myocardial perfusion reserve in refractory angina patients: A phase I/IIa study with 1-year follow-up. Eur. Heart J. 2017, 38, 2547–2555. [Google Scholar] [CrossRef] [Green Version]
- Stewart, D.J.; Hilton, J.D.; Arnold, J.M.O.; Gregoire, J.; Rivard, A.; Archer, S.L.; Charbonneau, F.; Cohen, E.; Curtis, M.; Buller, C.E.; et al. Angiogenic gene therapy in patients with nonrevascularizable ischemic heart disease: A phase 2 randomized, controlled trial of AdVEGF121 (AdVEGF121) versus maximum medical treatment. Gene Ther. 2006, 13, 1503–1511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedman, M.; Hartikainen, J.; Syvänne, M.; Stjernvall, J.; Hedman, A.; Kivelä, A.; Vanninen, E.; Mussalo, H.; Kauppila, E.; Simula, S.; et al. Safety and feasibility of catheter-based local intracoronary vascular endothelial growth factor gene transfer in the prevention of postangioplasty and in-stent restenosis and in the treatment of chronic myocardial ischemia: Phase II results of the Kuopio. Circulation 2003, 107, 2677–2683. [Google Scholar] [CrossRef]
- Grines, C.L.; Watkins, M.W.; Helmer, G.; Penny, W.; Brinker, J.; Marmur, J.D.; West, A.; Rade, J.J.; Marrott, P.; Hammond, H.K.; et al. Angiogenic Gene Therapy (AGENT) trial in patients with stable angina pectoris. Circulation 2002, 105, 1291–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikol, S.; Baumgartner, I.; Van Belle, E.; Diehm, C.; Visoná, A.; Capogrossi, M.C.; Ferreira-Maldent, N.; Gallino, A.; Graham Wyatt, M.; Dinesh Wijesinghe, L.; et al. Therapeutic Angiogenesis With Intramuscular NV1FGF Improves Amputation-free Survival in Patients With Critical Limb Ischemia. Mol. Ther. 2008, 16, 972–978. [Google Scholar] [CrossRef]
- Lunde, K.; Solheim, S.; Aakhus, S.; Arnesen, H.; Abdelnoor, M.; Egeland, T.; Endresen, K.; Ilebekk, A.; Mangschau, A.; Fjeld, J.G.; et al. Intracoronary injection of mononuclear bone marrow cells in acute myocardial infarction. N. Engl. J. Med. 2006, 355, 1199–1209. [Google Scholar] [CrossRef] [Green Version]
- Perin, E.C.; Willerson, J.T.; Pepine, C.J.; Henry, T.D.; Ellis, S.G.; Zhao, D.X.M.; Silva, G.V.; Lai, D.; Thomas, J.D.; Kronenberg, M.W.; et al. Effect of Transendocardial Delivery of Autologous Bone Marrow Mononuclear Cells on Functional Capacity, Left Ventricular Function, and Perfusion in Chronic Heart Failure: The FOCUS-CCTRN Trial. JAMA 2012, 307, 1717–1726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, G.P.; Wollert, K.C.; Lotz, J.; Steffens, J.; Lippolt, P.; Fichtner, S.; Hecker, H.; Schaefer, A.; Arseniev, L.; Hertenstein, B.; et al. Intracoronary bone marrow cell transfer after myocardial infarction: Eighteen months’ follow-up data from the randomized, controlled BOOST (BOne marrOw transfer to enhance ST-elevation infarct regeneration) trial. Circulation 2006, 113, 1287–1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartunek, J.; Terzic, A.; Davison, B.A.; Filippatos, G.S.; Radovanovic, S.; Beleslin, B.; Merkely, B.; Musialek, P.; Wojakowski, W.; Andreka, P.; et al. Cardiopoietic cell therapy for advanced ischaemic heart failure: Results at 39 weeks of the prospective, randomized, double blind, sham-controlled CHART-1 clinical trial. Eur. Heart J. 2017, 38, 648–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartunek, J.; Behfar, A.; Dolatabadi, D.; Vanderheyden, M.; Ostojic, M.; Dens, J.; El Nakadi, B.; Banovic, M.; Beleslin, B.; Vrolix, M.; et al. Cardiopoietic stem cell therapy in heart failure: The C-CURE (Cardiopoietic stem Cell therapy in heart failURE) multicenter randomized trial with lineage-specified biologics. J. Am. Coll. Cardiol. 2013, 61, 2329–2338. [Google Scholar] [CrossRef] [Green Version]
- Assmus, B.; Honold, J.; Schächinger, V.; Britten, M.B.; Fischer-Rasokat, U.; Lehmann, R.; Teupe, C.; Pistorius, K.; Martin, H.; Abolmaali, N.D.; et al. Transcoronary transplantation of progenitor cells after myocardial infarction. N. Engl. J. Med. 2006, 355, 1222–1232. [Google Scholar] [CrossRef] [Green Version]
- Henry, T.D.; Grines, C.L.; Watkins, M.W.; Dib, N.; Barbeau, G.; Moreadith, R.; Andrasfay, T.; Engler, R.L. Effects of Ad5FGF-4 in patients with angina: An analysis of pooled data from the AGENT-3 and AGENT-4 trials. J. Am. Coll. Cardiol. 2007, 50, 1038–1046. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Zhou, G.; Wu, W.; Duan, Y.; Ma, G.; Song, J.; Xiao, R.; Vandenberghe, L.; Zhang, F.; D’Amore, P.A.; et al. Genome editing abrogates angiogenesis in vivo. Nat. Commun. 2017, 8, 112. [Google Scholar] [CrossRef]
- Suzuki, G.; Iyer, V.; Lee, T.C.; Canty, J.M., Jr. Autologous Mesenchymal Stem Cells Mobilize cKit+ and CD133+ Bone Marrow Progenitor Cells and Improve Regional Function in Hibernating Myocardium. Circ. Res. 2011, 109, 1044–1054. [Google Scholar] [CrossRef] [Green Version]
- Weil, B.R.; Suzuki, G.; Leiker, M.M.; Fallavollita, J.A.; Canty, J.M., Jr. Comparative Efficacy of Intracoronary Allogeneic Mesenchymal Stem Cells and Cardiosphere-Derived Cells in Swine with Hibernating Myocardium. Circ. Res. 2015, 117, 634–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaefer, A.; Zwadlo, C.; Fuchs, M.; Meyer, G.P.; Lippolt, P.; Wollert, K.C.; Drexler, H. Long-term effects of intracoronary bone marrow cell transfer on diastolic function in patients after acute myocardial infarction: 5-year results from the randomized-controlled BOOST trial--an echocardiographic study. Eur. J. Echocardiogr. 2010, 11, 165–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Noronha, N.C.; Mizukami, A.; Caliári-Oliveira, C.; Cominal, J.G.; Rocha, J.L.M.; Covas, D.T.; Swiech, K.; Malmegrim, K.C.R. Priming approaches to improve the efficacy of mesenchymal stromal cell-based therapies. Stem Cell Res. Ther. 2019, 10, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhury, S.M.; Abou-Elkacem, L.; Lee, T.; Dahl, J.; Lutz, A.M. Ultrasound and microbubble mediated therapeutic delivery: Underlying mechanisms and future outlook. J. Control. Release 2020, 326, 75–90. [Google Scholar] [CrossRef]
- Gnecchi, M.; He, H.; Liang, O.D.; Melo, L.G.; Morello, F.; Mu, H.; Noiseux, N.; Zhang, L.; Pratt, R.E.; Ingwall, J.S.; et al. Paracrine action accounts for marked protection of ischemic heart by Akt-modified mesenchymal stem cells. Nat. Med. 2005, 11, 367–368. [Google Scholar] [CrossRef]
- Gnecchi, M.; He, H.; Noiseux, N.; Liang, O.D.; Zhang, L.; Morello, F.; Mu, H.; Melo, L.G.; Pratt, R.E.; Ingwall, J.S.; et al. Evidence supporting paracrine hypothesis for Akt-modified mesenchymal stem cell-mediated cardiac protection and functional improvement. FASEB J. 2006, 20, 661–669. [Google Scholar] [CrossRef]
- Alibhai, F.J.; Tobin, S.W.; Yeganeh, A.; Weisel, R.D.; Li, R.K. Emerging roles of extracellular vesicles in cardiac repair and rejuvenation. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H733–H744. [Google Scholar] [CrossRef]
- Potz, B.A.; Scrimgeour, L.A.; Pavlov, V.I.; Sodha, N.R.; Abid, M.R.; Sellke, F.W. Extracellular Vesicle Injection Improves Myocardial Function and Increases Angiogenesis in a Swine Model of Chronic Ischemia. J. Am. Heart Assoc. 2018, 7, e008344. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.D.; Johansson, H.J.; Graham, C.S.; Vesterlund, M.; Pham, M.T.; Bramlett, C.S.; Montgomery, E.N.; Mellema, M.S.; Bardini, R.L.; Contreras, Z.; et al. Comprehensive Proteomic Analysis of Mesenchymal Stem Cell Exosomes Reveals Modulation of Angiogenesis via Nuclear Factor-KappaB Signaling. Stem Cells 2016, 34, 601–613. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-S.; Choi, D.-Y.; Yun, S.J.; Choi, S.-M.; Kang, J.W.; Jung, J.W.; Hwang, D.; Kim, K.P.; Kim, D.-W. Proteomic Analysis of Microvesicles Derived from Human Mesenchymal Stem Cells. J. Proteome Res. 2012, 11, 839–849. [Google Scholar] [CrossRef] [PubMed]
- Ruel, M.; Wu, G.F.; Khan, T.A.; Voisine, P.; Bianchi, C.; Li, J.; Li, J.; Laham, R.J.; Sellke, F.W. Inhibition of the cardiac angiogenic response to surgical FGF-2 therapy in a Swine endothelial dysfunction model. Circulation 2003, 108 (Suppl. 1), II335–II340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voisine, P.; Bianchi, C.; Ruel, M.; Malik, T.; Rosinberg, A.; Feng, J.; Khan, T.A.; Xu, S.-H.; Sandmeyer, J.; Laham, R.J.; et al. Inhibition of the cardiac angiogenic response to exogenous vascular endothelial growth factor. Surgery 2004, 136, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Boodhwani, M.; Nakai, Y.; Voisine, P.; Feng, J.; Li, J.; Mieno, S.; Ramlawi, B.; Bianchi, C.; Laham, R.; Sellke, F.W. High-dose atorvastatin improves hypercholesterolemic coronary endothelial dysfunction without improving the angiogenic response. Circulation 2006, 114, I402–I408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boodhwani, M.; Mieno, S.; Voisine, P.; Feng, J.; Sodha, N.; Li, J.; Sellke, F.W. High-dose atorvastatin is associated with impaired myocardial angiogenesis in response to vascular endothelial growth factor in hypercholesterolemic swine. J. Thorac. Cardiovasc. Surg. 2006, 132, 1299–1306. [Google Scholar] [CrossRef] [Green Version]
- Klauber, N.; Browne, F.; Anand-Apte, B.; D’Amato, R.J. New Activity of Spironolactone. Circulation 1996, 94, 2566–2571. [Google Scholar] [CrossRef]
- Volpert, O.V.; Ward, W.F.; Lingen, M.W.; Chesler, L.; Solt, D.B.; Johnson, M.D.; Molteni, A.; Polverini, P.J.; Bouck, N.P. Captopril inhibits angiogenesis and slows the growth of experimental tumors in rats. J. Clin. Investig. 1996, 98, 671–679. [Google Scholar] [CrossRef]
- Dai, X.; Yan, J.; Fu, X.; Pan, Q.; Sun, D.; Xu, Y.; Wang, J.; Nie, L.; Tong, L.; Shen, A.; et al. Aspirin Inhibits Cancer Metastasis and Angiogenesis via Targeting Heparanase. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2017, 23, 6267–6278. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Lou, X.; Wang, R.; Sun, C.; Liu, X.; Liu, S.; Wang, Z.; Ni, C. Aspirin, a Potential GLUT1 Inhibitor in a Vascular Endothelial Cell Line. Open Med. 2019, 14, 552–560. [Google Scholar] [CrossRef]
- Sun, H.; Davison, B.A.; Cotter, G.; Pencina, M.J.; Koch, G.G. Evaluating Treatment Efficacy by Multiple End Points in Phase II Acute Heart Failure Clinical Trials. Circ. Heart Fail. 2012, 5, 742–749. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention. National Diabetes Statistics Report, 2020; Centers for Disease Control and Prevention, US Department of Health and Human Services: Atlanta, GA, USA, 2020.
- Hales, C.M.; Carroll, M.D.; Fryar, C.D.; Ogden, C.L. Prevalence of obesity and severe obesity among adults: United States, 2017–2018; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2020; p. 360.
- Sodha, N.R.; Boodhwani, M.; Clements, R.T.; Xu, S.-H.; Khabbaz, K.R.; Sellke, F.W. Increased antiangiogenic protein expression in the skeletal muscle of diabetic swine and patients. Arch. Surg. 2008, 143, 463–470. [Google Scholar] [CrossRef] [Green Version]
- Boodhwani, M.; Nakai, Y.; Mieno, S.; Voisine, P.; Bianchi, C.; Araujo, E.G.; Feng, J.; Michael, K.; Li, J.; Sellke, F.W. Hypercholesterolemia impairs the myocardial angiogenic response in a swine model of chronic ischemia: Role of endostatin and oxidative stress. Ann. Thorac. Surg. 2006, 81, 634–641. [Google Scholar] [CrossRef] [PubMed]
- Sodha, N.R.; Clements, R.T.; Boodhwani, M.; Xu, S.-H.; Laham, R.J.; Bianchi, C.; Sellke, F.W. Endostatin and angiostatin are increased in diabetic patients with coronary artery disease and associated with impaired coronary collateral formation. Am. J. Physiol. Heart Circ. Physiol. 2009, 296, H428–H434. [Google Scholar] [CrossRef] [Green Version]
- Aboulgheit, A.; Potz, B.A.; Scrimgeour, L.A.; Karbasiafshar, C.; Shi, G.; Zhang, Z.; Machan, J.T.; Schorl, C.; Brodsky, A.S.; Braga, K.; et al. Effects of High Fat Versus Normal Diet on Extracellular Vesicle-Induced Angiogenesis in a Swine Model of Chronic Myocardial Ischemia. J. Am. Heart Assoc. 2021, 10, e017437. [Google Scholar] [CrossRef] [PubMed]
- Boodhwani, M.; Sodha, N.R.; Mieno, S.; Xu, S.-H.; Feng, J.; Ramlawi, B.; Clements, R.T.; Sellke, F.W. Functional, cellular, and molecular characterization of the angiogenic response to chronic myocardial ischemia in diabetes. Circulation 2007, 116, 31–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boodhwani, M.; Sodha, N.R.; Mieno, S.; Ramlawi, B.; Xu, S.-H.; Feng, J.; Clements, R.T.; Ruel, M.; Sellke, F.W. Insulin treatment enhances the myocardial angiogenic response in diabetes. J. Thorac. Cardiovasc. Surg. 2007, 134, 1453–1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, T.; Zhao, L.; Manuel, G.; Taylor, H.; Liu, D. Approaches to therapeutic angiogenesis for ischemic heart disease. J. Mol. Med. 2019, 97, 141–151. [Google Scholar] [CrossRef]
- Bai, Y.; Leng, Y.; Yin, G.; Pu, X.; Huang, Z.; Liao, X.; Chen, X.; Yao, Y. Effects of combinations of BMP-2 with FGF-2 and/or VEGF on HUVECs angiogenesis in vitro and CAM angiogenesis in vivo. Cell Tissue Res. 2014, 356, 109–121. [Google Scholar] [CrossRef]
- Park, B.-W.; Jung, S.-H.; Das, S.; Lee, S.M.; Park, J.-H.; Kim, H.; Hwang, J.-W.; Lee, S.; Kim, H.-J.; Kim, H.-Y.; et al. In vivo priming of human mesenchymal stem cells with hepatocyte growth factor–engineered mesenchymal stem cells promotes therapeutic potential for cardiac repair. Sci. Adv. 2020, 6, eaay6994. [Google Scholar] [CrossRef] [Green Version]
- Bandara, N.; Gurusinghe, S.; Chen, H.; Chen, S.; Wang, L.; Lim, S.Y.; Strappe, P. Minicircle DNA-mediated endothelial nitric oxide synthase gene transfer enhances angiogenic responses of bone marrow-derived mesenchymal stem cells. Stem Cell Res. Ther. 2016, 7, 48. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Shen, H.; Shao, L.; Teng, X.; Chen, Y.; Liu, X.; Yang, Z.; Shen, Z. HIF-1α overexpression in mesenchymal stem cell-derived exosomes mediates cardioprotection in myocardial infarction by enhanced angiogenesis. Stem Cell Res. Ther. 2020, 11, 373. [Google Scholar] [CrossRef] [PubMed]
- Vale, P.R.; Losordo, D.W.; Milliken, C.E.; McDonald, M.C.; Gravelin, L.M.; Curry, C.M.; Esakof, D.D.; Maysky, M.; Symes, J.F.; Isner, J.M. Randomized, single-blind, placebo-controlled pilot study of catheter-based myocardial gene transfer for therapeutic angiogenesis using left ventricular electromechanical mapping in patients with chronic myocardial ischemia. Circulation 2001, 103, 2138–2143. [Google Scholar] [CrossRef] [Green Version]
- Chu, H.; Wang, Y. Therapeutic angiogenesis: Controlled delivery of angiogenic factors. Ther. Deliv. 2012, 3, 693–714. [Google Scholar] [CrossRef] [Green Version]
- Silva, E.A.; Mooney, D.J. Spatiotemporal control of vascular endothelial growth factor delivery from injectable hydrogels enhances angiogenesis. J. Thromb. Haemost. 2007, 5, 590–598. [Google Scholar] [CrossRef]
- Zahid, M.; Phillips, B.E.; Albers, S.M.; Giannoukakis, N.; Watkins, S.C.; Robbins, P.D. Identification of a cardiac specific protein transduction domain by in vivo biopanning using a M13 phage peptide display library in mice. PLoS ONE 2010, 5, e12252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanki, S.; Jaalouk, D.E.; Lee, S.; Yu, A.Y.C.; Gannon, J.; Lee, R.T. Identification of targeting peptides for ischemic myocardium by in vivo phage display. J. Mol. Cell. Cardiol. 2011, 50, 841–848. [Google Scholar] [CrossRef] [Green Version]
- Vandergriff, A.; Huang, K.; Shen, D.; Hu, S.; Hensley, M.T.; Caranasos, T.G.; Qian, L.; Cheng, K. Targeting regenerative exosomes to myocardial infarction using cardiac homing peptide. Theranostics 2018, 8, 1869–1878. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, Y.; Zhao, Z.; Meng, Q.; Yu, Y.; Sun, J.; Yang, Z.; Chen, Y.; Li, J.; Ma, T.; et al. Engineered Exosomes With Ischemic Myocardium-Targeting Peptide for Targeted Therapy in Myocardial Infarction. J. Am. Heart Assoc. 2018, 7, e008737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, H.; Gao, J.; Chen, C.-W.; Huard, J.; Wang, Y. Injectable fibroblast growth factor-2 coacervate for persistent angiogenesis. Proc. Natl. Acad. Sci. USA 2011, 108, 13444–13449. [Google Scholar] [CrossRef] [Green Version]
- Chu, H.; Johnson, N.R.; Mason, N.S.; Wang, Y. A [polycation:heparin] complex releases growth factors with enhanced bioactivity. J. Control. Release 2011, 150, 157–163. [Google Scholar] [CrossRef]
- Wu, J.; Ye, J.; Zhu, J.; Xiao, Z.; He, C.; Shi, H.; Wang, Y.; Lin, C.; Zhang, H.; Zhao, Y.; et al. Heparin-Based Coacervate of FGF2 Improves Dermal Regeneration by Asserting a Synergistic Role with Cell Proliferation and Endogenous Facilitated VEGF for Cutaneous Wound Healing. Biomacromolecules 2016, 17, 2168–2177. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Agent | Study Design (Disease; Delivery; Dose; Number of Patients) | Outcome | Ref. |
|---|---|---|---|
| Protein Therapy | |||
| VEGF | CAD; IC day 0 and IV day 3,6,9; 17 ng/kg/min or 50 ng/kg/min; n = 178) | No improvement in exercise time 60 days post treatment | [25] |
| FGF | CAD; IC; single injection of 0, 0.3, 3, or 30 µg/kg; n = 337 | Exercise tolerance and angina symptoms improved at 90 days; no difference at 180 days | [26] |
| CAD; IC via heparin-alginate slow-release device; 1 or 10 µg; n = 8 | Exercise tolerance and myocardial perfusion showed a trend toward improvement at 90 days, but not at 180 days | [4] | |
| Gene Therapy | |||
| VEGF | CAD; IM 10×; 200 µg supplemented with 6 g l-arginine per day for 3 months; n = 19 | Improved anterior wall perfusion and anterior wall contractility at 3 months | [27] |
| CAD, IM; 125 or 250 µg; n = 15 | Angina was significantly reduced and myocardial perfusion was improved | [28] | |
| Angina, IM, 200 µL at 10 sites; n = 30 | Myocardial perfusion reserve significantly increased at 3 months and 12 months compared to baseline, although no significance between 3 and 12 months. | [29] | |
| IHD, IM, 4 × 1010 pfu, n = 67 | Total exercise duration and time were improved at 12 and 26 weeks | [30] | |
| IHD, IC, 2 × 1010 pfu, n = 103 | Myocardial perfusion was significantly improved at 6 months; no changes in minimal lumen diameter nor % of diameter stenosis were also reported | [31] | |
| FGF | Angina, IC, 5 different dose groups, n = 79 | Increased exercise time at 4 weeks | [32] |
| CLI; intramuscular; 4 mg at day 1, 15, 30, and 45; n = 125 | Complete healing of at least one ulcer in the treated limb at week 25; treatment also significantly reduced the risk of all amputations by two-fold | [33] | |
| Stem-Cell Therapy | |||
| BM-MSC | MI; IC; day 6 post-MI on average; 7 × 105 cells; n = 101 | LVEF was increased at 6 months; no change in LV EDV nor infarct size was observed. | [34] |
| CAD; transendocardial injection; 1 × 108; n = 92 | LV ESV nor maximal oxygen consumption were improved at 6 months | [35] | |
| (MI; IC; 24.6 ± 9.4 × 108 nucleated cells, 9.5 ± 6.3 × 106 CD34+ cells, and 3.6 ± 3.4 × 106 hematopoietic cells ~4.8 days post-MI; n = 60) | LVEF was improved at 6 months, but was not significant at 18 months | [36] | |
| CPC | IHD; IM; injections at 17 sites; n = 315 | No significant improvements in primary endpoints of MLHFQ score, 6 min walk distance; LV ESV and LV EF at 39 weeks | [37] |
| IHD; IM; 600 × 106 to 1200 × 106 cells; n = 319 | LVEF was improved with reduction in LV ESV, and improved 6-min walk distance | [38] | |
| BMC or CPC | MI, IC, mean of 22 × 106 CPC or 205 × 106 BMC, n = 75 | BMC treatment significantly increased LVEF compared to CPC and control groups at 3 months. | [39] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sabra, M.; Karbasiafshar, C.; Aboulgheit, A.; Raj, S.; Abid, M.R.; Sellke, F.W. Clinical Application of Novel Therapies for Coronary Angiogenesis: Overview, Challenges, and Prospects. Int. J. Mol. Sci. 2021, 22, 3722. https://doi.org/10.3390/ijms22073722
Sabra M, Karbasiafshar C, Aboulgheit A, Raj S, Abid MR, Sellke FW. Clinical Application of Novel Therapies for Coronary Angiogenesis: Overview, Challenges, and Prospects. International Journal of Molecular Sciences. 2021; 22(7):3722. https://doi.org/10.3390/ijms22073722
Chicago/Turabian StyleSabra, Mohamed, Catherine Karbasiafshar, Ahmed Aboulgheit, Sidharth Raj, M. Ruhul Abid, and Frank W. Sellke. 2021. "Clinical Application of Novel Therapies for Coronary Angiogenesis: Overview, Challenges, and Prospects" International Journal of Molecular Sciences 22, no. 7: 3722. https://doi.org/10.3390/ijms22073722
APA StyleSabra, M., Karbasiafshar, C., Aboulgheit, A., Raj, S., Abid, M. R., & Sellke, F. W. (2021). Clinical Application of Novel Therapies for Coronary Angiogenesis: Overview, Challenges, and Prospects. International Journal of Molecular Sciences, 22(7), 3722. https://doi.org/10.3390/ijms22073722