
Application of Upstream Open Reading Frames (uORFs) Editing for the Development of Stress-Tolerant Crops


 ,
,
Abstract
:1. Introduction
2. Upstream Open Reading Frames (uORF)
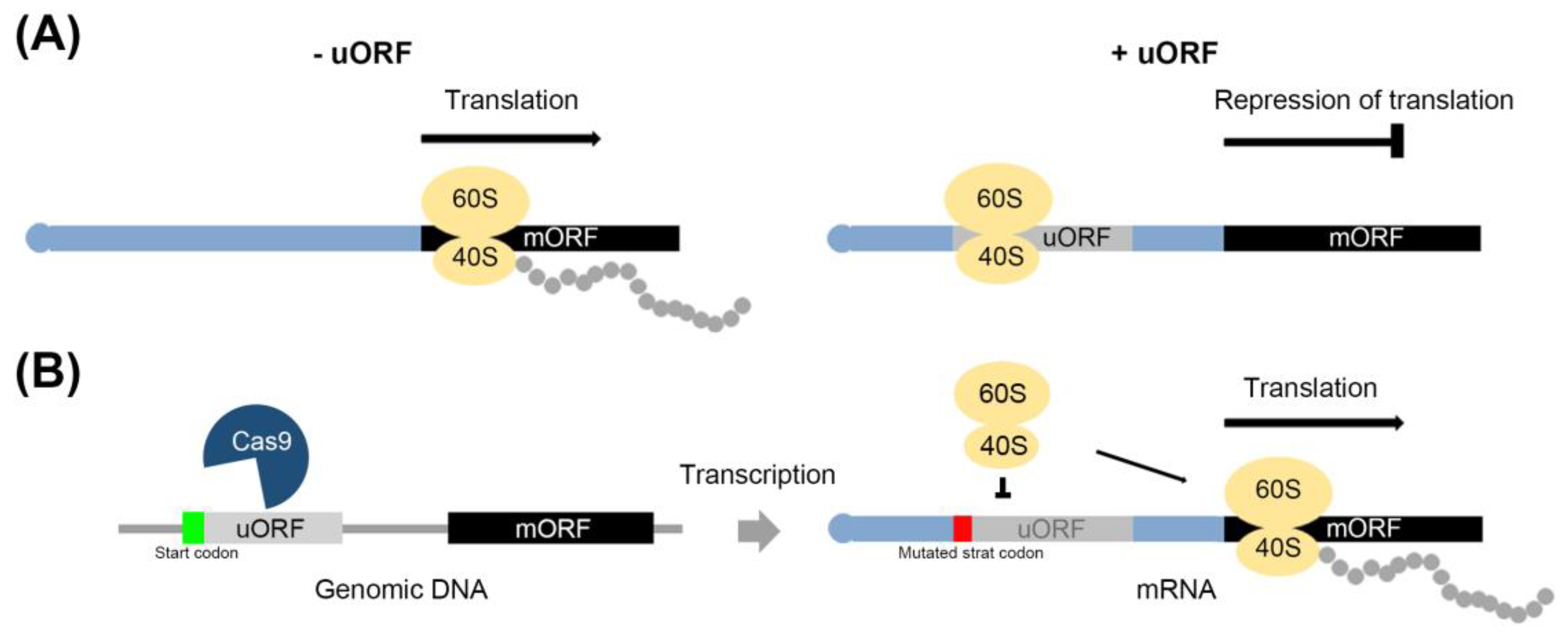
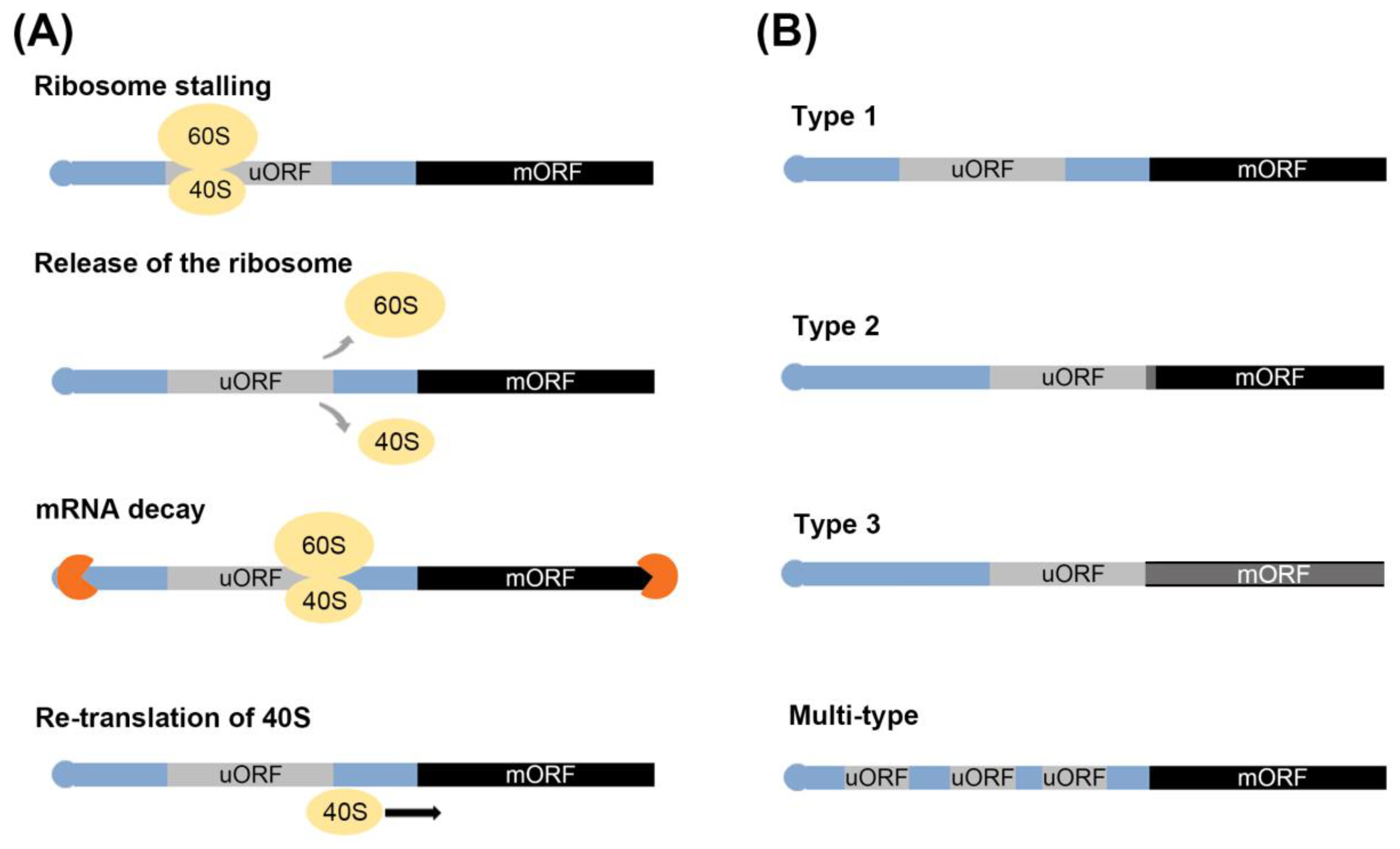
2.1. The Process of Translation of mRNA Mediated by uORF
2.2. The Classification of uORF
2.3. Stress-Responsive uORF-Mediated Transcripts in Rice
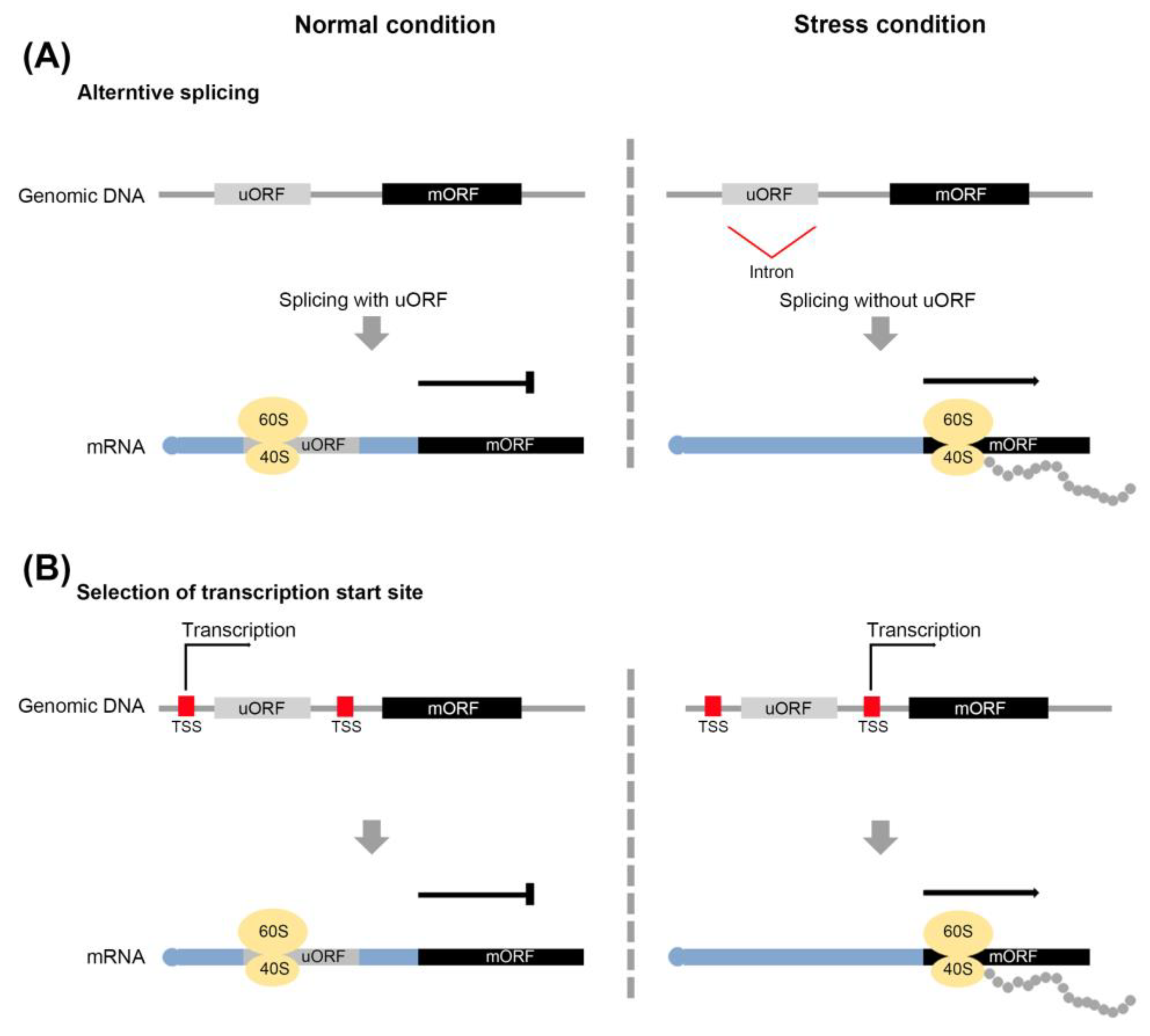
2.4. Avoidance of uORF-Mediated Repression of Gene Expression during Stress Responses

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene ID (Rap-DB) | Transcript ID (MSU) | uORF Type | Expression Response | Reference | ||
|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | |||||
| HAK1 | Os04g0401700 | LOC_Os04g32920.2 | 1 | 0 | 0 | down | [48] |
| LOC_Os04g32920.4 | 4 | 0 | 0 | ||||
| HAK10 | Os06g0625900 | LOC_Os06g42030.1 | 1 | 0 | 0 | up | [49] |
| HKT4 | Os04g0607500 | LOC_Os04g51820.1 | 2 | 0 | 0 | up | [49] |
| LOC_Os04g51820.2 | 2 | 0 | 0 | ||||
| LOC_Os04g51820.3 | 3 | 12 | 0 | ||||
| HKT6 | Os02g0175000 | LOC_Os02g07830.1 | 2 | 1 | 0 | down | [49] |
| HKT8 | Os01g0307500 | LOC_Os01g20160.1 | 1 | 0 | 0 | down | [49] |
| NHX1 | Os07g0666900 | LOC_Os07g47100.1 | 1 | 0 | 0 | up | [50] |
| LOC_Os07g47100.2 | 1 | 0 | 0 | ||||
| LOC_Os07g47100.3 | 1 | 0 | 0 | ||||
| SOS1 | Os12g0641100 | LOC_Os12g44360.1 | 9 | 3 | 0 | up | [50] |
| LOC_Os12g44360.2 | 0 | 1 | 0 | ||||
| LOC_Os12g44360.3 | 0 | 1 | 0 | ||||
| LOC_Os12g44360.4 | 0 | 1 | 0 | ||||
| CAX3 | Os04g0653200 | LOC_Os04g55940.1 | 2 | 0 | 0 | up | [51] |
| LOC_Os04g55940.2 | 1 | 0 | 0 | ||||
| IDS1 | Os03g0818800 | LOC_Os03g60430.1 | 2 | 0 | 0 | down | [52] |
| LOC_Os03g60430.2 | 2 | 0 | 0 | ||||
| NAC6 | Os01g0884300 | LOC_Os01g66120.1 | 1 | 0 | 0 | up | [53] |
| LOC_Os01g66120.2 | 3 | 0 | 0 | ||||
| EIL2 | Os07g0685700 | LOC_Os07g48630.1 | 1 | 0 | 0 | down | [54] |
| LOC_Os07g48630.2 | 12 | 0 | 0 | ||||
| bHLH035 | Os01g0159800 | LOC_Os01g06640.1 | 3 | 0 | 0 | up | [55] |
| LOC_Os01g06640.2 | 2 | 0 | 0 | ||||
| LOC_Os01g06640.3 | 14 | 0 | 0 | ||||
| LEA5 | Os05g0584200 | LOC_Os05g50710.1 | 1 | 0 | 0 | up | [56] |
| JAZ9 | Os03g0180800 | LOC_Os03g08310.1 | 1 | 0 | 0 | down | [57] |
| RSS3 | Os11g0446000 | LOC_Os03g08310.1 | 1 | 0 | 0 | up | [58] |
| ACA6 | Os01g0939100 | LOC_Os01g71240.1 | 3 | 0 | 0 | up | [59] |
| RLCK3 | Os01g0113300 | LOC_Os01g02300.1 | 1 | 0 | 0 | up | [60] |
| RLCK5 | Os01g0114100 | LOC_Os01g02390.1 | 0 | 1 | 0 | up | [60] |
| LOC_Os01g02390.2 | 0 | 1 | 0 | ||||
| RLCK6 | Os01g0114300 | LOC_Os01g02400.1 | 0 | 0 | 0 | up | [60] |
| LOC_Os01g02400.2 | 17 | 1 | 0 | ||||
| LOC_Os01g02400.3 | 39 | 0 | 0 | ||||
| RLCK27 | Os01g0247500 | LOC_Os01g14510.1 | 8 | 0 | 0 | up | [60] |
| RLCK42 | Os01g0602800 | LOC_Os01g41870.1 | 1 | 0 | 0 | down | [60] |
| LOC_Os01g41870.2 | 1 | 0 | 0 | ||||
| RLCK48 | Os01g0852100 | LOC_Os01g63280.1 | 20 | 3 | 0 | down | [60] |
| LOC_Os01g63280.2 | 1 | 1 | 0 | ||||
| LOC_Os01g63280.3 | 14 | 3 | 0 | ||||
| RLCK57 | Os01g0973500 | LOC_Os01g74200.1 | 1 | 0 | 0 | up | [60] |
| RLCK72 | Os02g0513000 | LOC_Os02g30900.1 | 2 | 0 | 0 | up | [60] |
| RLCK80 | Os02g0650500 | LOC_Os02g43430.1 | 1 | 0 | 0 | up | [60] |
| RLCK85 (ER2) | Os02g0777400 | LOC_Os02g53720.1 | 1 | 0 | 0 | down | [60] |
| LOC_Os02g53720.2 | 1 | 0 | 0 | ||||
| RLCK95 | Os03g0113000 | LOC_Os03g02190.1 | 0 | 1 | 0 | down | [60] |
| RLCK101 (PTK3) | Os03g0159100 | LOC_Os03g06330.1 | 9 | 0 | 0 | up | [60] |
| RLCK106 | Os03g0264300 | LOC_Os03g15770.1 | 4 | 0 | 0 | up | [60] |
| LOC_Os03g15770.2 | 1 | 1 | 0 | ||||
| LOC_Os03g15770.3 | 5 | 0 | 0 | ||||
| RLCK110 (PTK5) | Os03g0407900 | LOC_Os03g29410.1 | 2 | 0 | 0 | up | [60] |
| RLCK167 | Os04g0654600 | LOC_Os04g56060.1 | 3 | 0 | 0 | down | [60] |
| RLCK168 | Os04g0655300 | LOC_Os04g56110.1 | 6 | 0 | 0 | up | [60] |
| LOC_Os04g56110.2 | 23 | 0 | 0 | ||||
| LOC_Os04g56110.3 | 5 | 0 | 0 | ||||
| RLCK191 | Os05g0589700 | LOC_Os05g51190.2 | 1 | 0 | 0 | down | [60] |
| LOC_Os05g51190.3 | 1 | 0 | 0 | ||||
| RLCK194 (PUB5) | Os06g0140800 | LOC_Os06g04880.1 | 2 | 0 | 0 | up | [60] |
| RLCK204 | Os06g0203800 | LOC_Os06g10230.1 | 1 | 1 | 0 | down | [60] |
| RLCK216 (RRK1) | Os06g0693200 | LOC_Os06g47820.1 | 1 | 0 | 0 | up | [60] |
| RLCK218 | Os06g0714900 | LOC_Os06g50100.1 | 6 | 0 | 0 | up | [60] |
| RLCK223 | Os07g0159700 | LOC_Os07g06570.1 | 1 | 0 | 0 | down | [60] |
| RLCK242 | Os07g0693000 | LOC_Os07g49240.1 | 1 | 0 | 0 | down | [60] |
| RLCK249 (ER1) | Os06g0203800 | LOC_Os06g10230.1 | 1 | 1 | 0 | down | [60] |
| RLCK301 | Os10g0442800 | LOC_Os10g30600.1 | 1 | 0 | 0 | up | [60] |
| LOC_Os10g30600.2 | 5 | 0 | 0 | ||||
| RLCK334 | Os11g0556400 | LOC_Os11g35274.1 | 1 | 0 | 0 | down | [60] |
| RLCK373 | Os12g0611100 | LOC_Os12g41710.1 | 4 | 0 | 0 | down | [60] |
| LOC_Os12g41710.2 | 30 | 0 | 0 | ||||
| LOC_Os12g41710.3 | 16 | 0 | 0 | ||||
| LOC_Os12g41710.4 | 29 | 0 | 0 | ||||
| RLCK375 | Os12g0615300 | LOC_Os12g42070.1 | 7 | 0 | 0 | down | [60] |
| Aldh2a | Os02g0730000 | LOC_Os02g49720.6 | 7 | 1 | 0 | up | [61] |
| ABA2 | Os03g0810800 | LOC_Os03g59610.1 | 1 | 0 | 0 | up | [55] |
3. Genetically Modified (GM) Crops
3.1. Development of the First GM Crops
3.2. Hormone Regulation in Plant Salt Stress Response
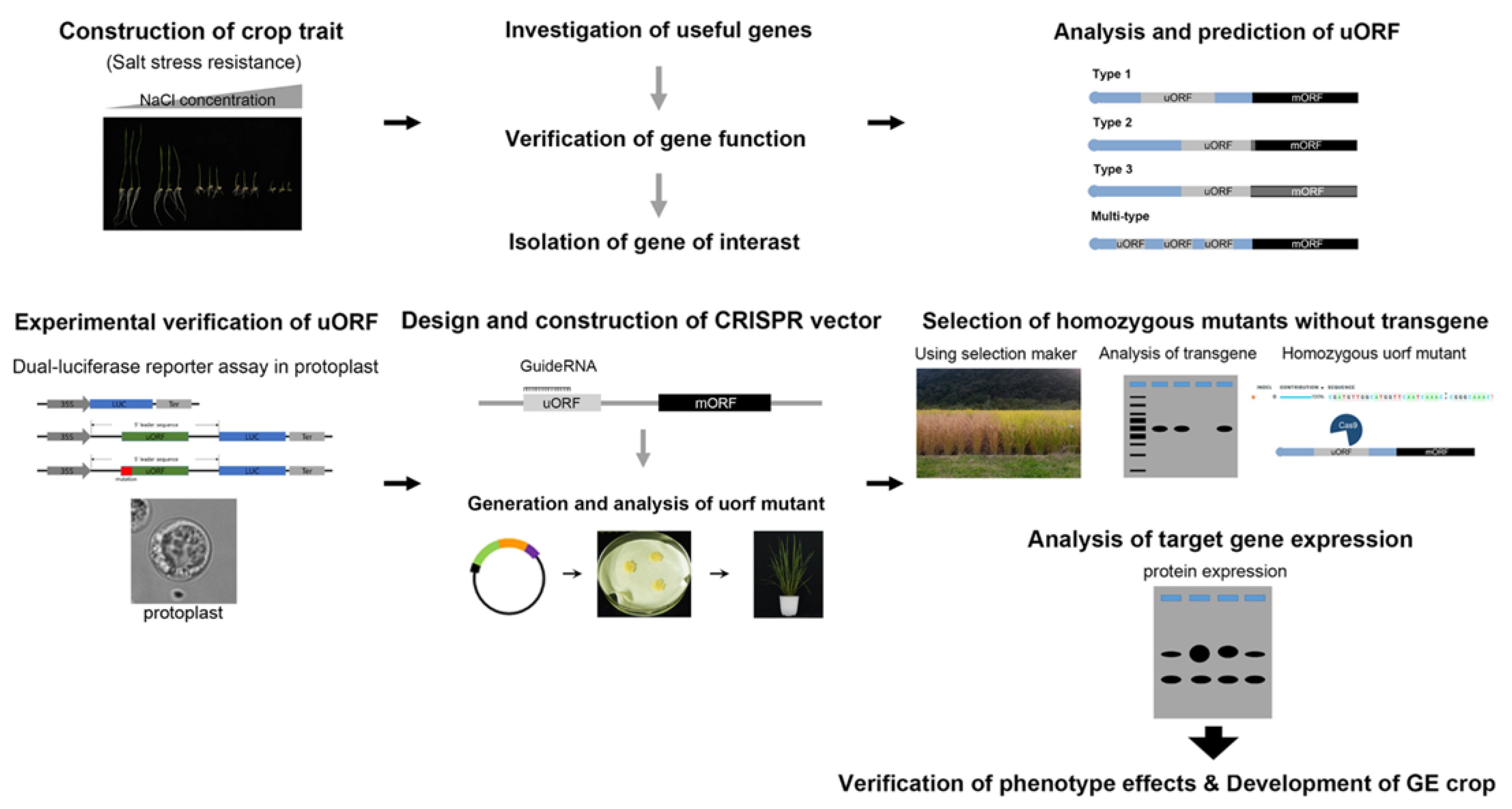
4. Enhancing Gene Expression in Crops by Editing uORFs Using the CRISPR System as an Alternative to GM Approaches
5. Future Prospects
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhao, C.; Liu, B.; Piao, S.; Wang, X.; Lobell, D.B.; Huang, Y.; Huang, M.; Yao, Y.; Bassu, S.; Ciais, P.; et al. Temperature increase reduces global yields of major crops in four independent estimates. Proc. Natl. Acad. Sci. USA 2017, 114, 9326–9331. [Google Scholar] [CrossRef] [Green Version]
- d’Amour, C.B.; Reitsma, F.; Baiocchi, G.; Barthel, S.; Güneralp, B.; Erb, K.-H.; Haberl, H.; Creutzig, F.; Seto, K.C. Future urban land expansion and implications for global croplands. Proc. Natl. Acad. Sci. USA 2017, 114, 8939–8944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Röös, E.; Bajželj, B.; Smith, P.; Patel, M.; Little, D.; Garnett, T. Greedy or needy? Land use and climate impacts of food in 2050 under different livestock futures. Glob. Environ. Chang. 2017, 47, 1–12. [Google Scholar] [CrossRef]
- Kamthan, A.; Chaudhuri, A.; Kamthan, M.; Datta, A. Genetically modified (GM) crops: Milestones and new advances in crop improvement. Theor. Appl. Genet. 2016, 129, 1639–1655. [Google Scholar] [CrossRef]
- Sakamoto, T. Phytohormones and rice crop yield: Strategies and opportunities for genetic improvement. Transgenic Res. 2006, 15, 399–404. [Google Scholar] [CrossRef]
- McSteen, P.; Zhao, Y. Plant hormones and signaling: Common themes and new developments. Dev. Cell 2008, 14, 467–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spartz, A.K.; Gray, W.M. Plant hormone receptors: New perceptions. Genes Dev. 2008, 22, 2139–2148. [Google Scholar] [CrossRef] [Green Version]
- Gaj, T.; Gersbach, C.A.; Barbas, C.F., III. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends Biotechnol. 2013, 31, 397–405. [Google Scholar] [CrossRef] [Green Version]
- Calyxt Inc. First Commercial Sale of Calyxt High Oleic Soybean Oil on the U.S. Market; Calyxt Inc.: St. Paul, MN, USA, 2019; Available online: https://calyxt.com/first-commercial-sale-of-calyxt-high-oleic-soybean-oil-on-the-u-s-market (accessed on 26 February 2019).
- Sikora, P.; Chawade, A.; Larsson, M.; Olsson, J.; Olsson, O. Mutagenesis as a tool in plant genetics, functional genomics, and breeding. Int. J. Plant Genom. 2011, 2011, 314829. [Google Scholar] [CrossRef] [Green Version]
- Grohmann, L.; Keilwagen, J.; Duensing, N.; Dagand, E.; Hartung, F.; Wilhelm, R.; Bendiek, J.; Sprink, T. Detection and identification of genome editing in plants: Challenges and opportunities. Front. Plant Sci. 2019, 10, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menz, J.; Modrzejewski, D.; Hartung, F.; Wilhelm, R.; Sprink, T. Genome edited crops touch the market: A view on the global development and regulatory environment. Front. Plant Sci. 2020, 11, 586027. [Google Scholar] [CrossRef] [PubMed]
- Kurihara, Y. uORF Shuffling Fine-Tunes Gene Expression at a Deep Level of the Process. Plants 2020, 9, 608. [Google Scholar] [CrossRef]
- Morris, D.R.; Geballe, A.P. Upstream open reading frames as regulators of mRNA translation. Mol. Cell. Biol. 2000, 20, 8635–8642. [Google Scholar] [CrossRef] [Green Version]
- Bazin, J.; Baerenfaller, K.; Gosai, S.J.; Gregory, B.D.; Crespi, M.; Bailey-Serres, J. Global analysis of ribosome-associated noncoding RNAs unveils new modes of translational regulation. Proc. Natl. Acad. Sci. USA 2017, 114, E10018–E10027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvo, S.E.; Pagliarini, D.J.; Mootha, V.K. Upstream open reading frames cause widespread reduction of protein expression and are polymorphic among humans. Proc. Natl. Acad. Sci. USA 2009, 106, 7507–7512. [Google Scholar] [CrossRef] [Green Version]
- Von Arnim, A.G.; Jia, Q.; Vaughn, J.N. Regulation of plant translation by upstream open reading frames. Plant Sci. 2014, 214, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ebina, I.; Takemoto-Tsutsumi, M.; Watanabe, S.; Koyama, H.; Endo, Y.; Kimata, K.; Igarashi, T.; Murakami, K.; Kudo, R.; Ohsumi, A. Identification of novel Arabidopsis thaliana upstream open reading frames that control expression of the main coding sequences in a peptide sequence-dependent manner. Nucleic Acids Res. 2015, 43, 1562–1576. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, N.; Sasaki, S.; Takahashi, H.; Yamashita, Y.; Naito, S.; Onouchi, H. Identification of Arabidopsis thaliana upstream open reading frames encoding peptide sequences that cause ribosomal arrest. Nucleic Acids Res. 2017, 45, 8844–8858. [Google Scholar] [CrossRef]
- Barbosa, C.; Peixeiro, I.; Romão, L. Gene expression regulation by upstream open reading frames and human disease. PLoS Genet. 2013, 9, e1003529. [Google Scholar] [CrossRef] [Green Version]
- Wittmann, J.; Hol, E.M.; Jäck, H.-M. hUPF2 silencing identifies physiologic substrates of mammalian nonsense-mediated mRNA decay. Mol. Cell. Biol. 2006, 26, 1272–1287. [Google Scholar] [CrossRef] [Green Version]
- Yepiskoposyan, H.; Aeschimann, F.; Nilsson, D.; Okoniewski, M.; Mühlemann, O. Autoregulation of the nonsense-mediated mRNA decay pathway in human cells. RNA 2011, 17, 2108–2118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spriggs, K.A.; Bushell, M.; Willis, A.E. Translational regulation of gene expression during conditions of cell stress. Mol. Cell 2010, 40, 228–237. [Google Scholar] [CrossRef]
- Niu, R.; Zhou, Y.; Zhang, Y.; Mou, R.; Tang, Z.; Wang, Z.; Zhou, G.; Guo, S.; Yuan, M.; Xu, G. uORFlight: A vehicle toward uORF-mediated translational regulation mechanisms in eukaryotes. Database (Oxford) 2020, 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittler, R. Abiotic stress, the field environment and stress combination. Trends Plant Sci. 2006, 11, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Peet, M.; Kramer, P. Effects of decreasing source/sink ratio in soybeans on photosynthesis, photorespiration, transpiration and yield. Plant Cell Environ. 1980, 3, 201–206. [Google Scholar]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Bartels, D.; Sunkar, R. Drought and salt tolerance in plants. Crit. Rev. Plant Sci. 2005, 24, 23–58. [Google Scholar] [CrossRef]
- Verslues, P.E.; Agarwal, M.; Katiyar-Agarwal, S.; Zhu, J.; Zhu, J.K. Methods and concepts in quantifying resistance to drought, salt and freezing, abiotic stresses that affect plant water status. Plant J. 2006, 45, 523–539. [Google Scholar] [CrossRef] [PubMed]
- Wellman, C.H.; Gray, J. The microfossil record of early land plants. Philos. Trans. R. Soc. B Biol. Sci. 2000, 355, 717–732. [Google Scholar] [CrossRef]
- Wellman, C.H.; Osterloff, P.L.; Mohiuddin, U. Fragments of the earliest land plants. Nature 2003, 425, 282. [Google Scholar] [CrossRef] [PubMed]
- Eckardt, N.A. Sequencing the rice genome. Am. Soc. Plant Biol. 2000. [Google Scholar] [CrossRef] [Green Version]
- Fairhurst, T.; Dobermann, A. Rice in the global food supply. World 2002, 5, 454, 349–511, 675. [Google Scholar]
- Mindell, J.; Maduke, M. ClC chloride channels. Genome Biol. 2001, 2, reviews3003.1. [Google Scholar] [CrossRef] [PubMed]
- Jentsch, T.J. CLC chloride channels and transporters: From genes to protein structure, pathology and physiology. Crit. Rev. Biochem. Mol. Biol. 2008, 43, 3–36. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, A.; Fukuda, A.; Sakai, S.; Tanaka, Y. Molecular cloning, functional expression and subcellular localization of two putative vacuolar voltage-gated chloride channels in rice (Oryza sativa L.). Plant Cell Physiol. 2006, 47, 32–42. [Google Scholar] [CrossRef]
- Um, T.Y.; Lee, S.; Kim, J.-K.; Jang, G.; Do Choi, Y. CHLORIDE CHANNEL 1 promotes drought tolerance in rice, leading to increased grain yield. Plant Biotechnol. Rep. 2018, 12, 283–293. [Google Scholar] [CrossRef]
- Huot, B.; Yao, J.; Montgomery, B.L.; He, S.Y. Growth–defense tradeoffs in plants: A balancing act to optimize fitness. Mol. Plant 2014, 7, 1267–1287. [Google Scholar] [CrossRef] [Green Version]
- Züst, T.; Agrawal, A.A. Trade-offs between plant growth and defense against insect herbivory: An emerging mechanistic synthesis. Annu. Rev. Plant Biol. 2017, 68, 513–534. [Google Scholar] [CrossRef] [Green Version]
- Hou, X.; Ding, L.; Yu, H. Crosstalk between GA and JA signaling mediates plant growth and defense. Plant Cell Rep. 2013, 32, 1067–1074. [Google Scholar] [CrossRef]
- Yang, D.-L.; Yao, J.; Mei, C.-S.; Tong, X.-H.; Zeng, L.-J.; Li, Q.; Xiao, L.-T.; Sun, T.-P.; Li, J.; Deng, X.-W. Plant hormone jasmonate prioritizes defense over growth by interfering with gibberellin signaling cascade. Proc. Natl. Acad. Sci. USA 2012, 109, E1192–E1200. [Google Scholar] [CrossRef] [Green Version]
- Um, T.Y.; Lee, H.Y.; Lee, S.; Chang, S.H.; Chung, P.J.; Oh, K.-B.; Kim, J.-K.; Jang, G.; Choi, Y.D. JASMONATE ZIM-DOMAIN PROTEIN 9 interacts with SLENDER RICE 1 to mediate the antagonistic interaction between jasmonic and gibberellic acid signals in rice. Front. Plant Sci. 2018, 9, 1866. [Google Scholar] [CrossRef]
- van der Horst, S.; Filipovska, T.; Hanson, J.; Smeekens, S. Metabolite control of translation by conserved peptide uORFs: The ribosome as a metabolite multisensor. Plant Physiol. 2020, 182, 110–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozak, M. Effects of intercistronic length on the efficiency of reinitiation by eucaryotic ribosomes. Mol. Cell. Biol. 1987, 7, 3438–3445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, A.K.; Lu, Y.; Zinta, G.; Lang, Z.; Zhu, J.-K. UTR-dependent control of gene expression in plants. Trends Plant Sci. 2018, 23, 248–259. [Google Scholar] [CrossRef]
- Tokizawa, M.; Kusunoki, K.; Koyama, H.; Kurotani, A.; Sakurai, T.; Suzuki, Y.; Sakamoto, T.; Kurata, T.; Yamamoto, Y.Y. Identification of Arabidopsis genic and non-genic promoters by paired-end sequencing of TSS tags. Plant J. 2017, 90, 587–605. [Google Scholar] [CrossRef]
- Chen, G.; Hu, Q.; Luo, L.; Yang, T.; Zhang, S.; Hu, Y.; Yu, L.; Xu, G. Rice potassium transporter O s HAK 1 is essential for maintaining potassium-mediated growth and functions in salt tolerance over low and high potassium concentration ranges. Plant Cell Environ. 2015, 38, 2747–2765. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, M.; Guo, R.; Shi, D.; Liu, B.; Lin, X.; Yang, C. Effects of salt stress on ion balance and nitrogen metabolism of old and young leaves in rice (Oryza sativa L.). BMC Plant Biol. 2012, 12, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossain, M.M.; Imran, S.; Islam, M.A.; Abdul, M. Expressional analysis of OsNHX1, OsNHX2, OsSOS1 and Os DREB transporters in salt tolerant (FR13A) and salt sensitive rice (brri dhan29) induced by salinity stress. IOSR J. Agric. Vet. Sci. 2017, 10, 64–70. [Google Scholar]
- Yamada, N.; Theerawitaya, C.; Cha-um, S.; Kirdmanee, C.; Takabe, T. Expression and functional analysis of putative vacuolar Ca 2+-transporters (CAXs and ACAs) in roots of salt tolerant and sensitive rice cultivars. Protoplasma 2014, 251, 1067–1075. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Zhang, S.; Tao, W.; Zhang, X.; Liu, J.; Sun, J.; Zhang, H.; Pu, L.; Huang, R.; Chen, T. INDETERMINATE SPIKELET1 recruits histone deacetylase and a transcriptional repression complex to regulate rice salt tolerance. Plant Physiol. 2018, 178, 824–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakashima, K.; Tran, L.S.P.; Van Nguyen, D.; Fujita, M.; Maruyama, K.; Todaka, D.; Ito, Y.; Hayashi, N.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional analysis of a NAC-type transcription factor OsNAC6 involved in abiotic and biotic stress-responsive gene expression in rice. Plant J. 2007, 51, 617–630. [Google Scholar] [CrossRef]
- Yang, C.; Ma, B.; He, S.-J.; Xiong, Q.; Duan, K.-X.; Yin, C.-C.; Chen, H.; Lu, X.; Chen, S.-Y.; Zhang, J.-S. MAOHUZI6/ETHYLENE INSENSITIVE3-LIKE1 and ETHYLENE INSENSITIVE3-LIKE2 regulate ethylene response of roots and coleoptiles and negatively affect salt tolerance in rice. Plant Physiol. 2015, 169, 148–165. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.-C.; Cheng, W.-H.; Hong, C.-Y.; Chang, Y.-S.; Chang, M.-C. The transcription factor OsbHLH035 mediates seed germination and enables seedling recovery from salt stress through ABA-dependent and ABA-independent pathways, respectively. Rice 2018, 11, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-S.; Zhu, H.-B.; Jin, G.-L.; Liu, H.-L.; Wu, W.-R.; Zhu, J. Genome-scale identification and analysis of LEA genes in rice (Oryza sativa L.). Plant Sci. 2007, 172, 414–420. [Google Scholar] [CrossRef]
- Wu, H.; Ye, H.; Yao, R.; Zhang, T.; Xiong, L. OsJAZ9 acts as a transcriptional regulator in jasmonate signaling and modulates salt stress tolerance in rice. Plant Sci. 2015, 232, 1–12. [Google Scholar] [CrossRef]
- Toda, Y.; Tanaka, M.; Ogawa, D.; Kurata, K.; Kurotani, K.-I.; Habu, Y.; Ando, T.; Sugimoto, K.; Mitsuda, N.; Katoh, E. RICE SALT SENSITIVE3 forms a ternary complex with JAZ and class-C bHLH factors and regulates jasmonate-induced gene expression and root cell elongation. Plant Cell 2013, 25, 1709–1725. [Google Scholar] [CrossRef] [Green Version]
- Huda, K.M.; Banu, M.S.A.; Garg, B.; Tula, S.; Tuteja, R.; Tuteja, N. Os ACA 6, a P-type IIB Ca2+ ATP ase promotes salinity and drought stress tolerance in tobacco by ROS scavenging and enhancing the expression of stress-responsive genes. Plant J. 2013, 76, 997–1015. [Google Scholar] [CrossRef]
- Vij, S.; Giri, J.; Dansana, P.K.; Kapoor, S.; Tyagi, A.K. The receptor-like cytoplasmic kinase (OsRLCK) gene family in rice: Organization, phylogenetic relationship, and expression during development and stress. Mol. Plant 2008, 1, 732–750. [Google Scholar] [CrossRef]
- Ranawake, A.L.; Mori, N.; Nakamura, C. Expression profiles of stress responsive genes in rice (Oryza sativa L.) under abiotic stresses. Biotechnol. Biotechnol. Equip. 2012, 26, 2838–2843. [Google Scholar] [CrossRef]
- Ahmad, P.; Ashraf, M.; Younis, M.; Hu, X.; Kumar, A.; Akram, N.A.; Al-Qurainy, F. Role of transgenic plants in agriculture and biopharming. Biotechnol. Adv. 2012, 30, 524–540. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Xia, X.; Peng, S.; Lumpkin, T.A. Meeting demands for increased cereal production in China. J. Cereal Sci. 2014, 59, 235–244. [Google Scholar] [CrossRef]
- Barton, K.A.; Binns, A.N.; Matzke, A.J.; Chilton, M.-D. Regeneration of intact tobacco plants containing full length copies of genetically engineered T-DNA, and transmission of T-DNA to R1 progeny. Cell 1983, 32, 1033–1043. [Google Scholar] [CrossRef]
- Gatehouse, J.A. Biotechnological prospects for engineering insect-resistant plants. Plant Physiol. 2008, 146, 881–887. [Google Scholar] [CrossRef] [Green Version]
- Herrera-Estrella, L.; Depicker, A.; Van Montagu, M.; Schell, J. Expression of chimaeric genes transferred into plant cells using a Ti-plasmid-derived vector. Nature 1983, 303, 209. [Google Scholar] [CrossRef]
- Meinke, D.W.; Cherry, J.M.; Dean, C.; Rounsley, S.D.; Koornneef, M. Arabidopsis thaliana: A model plant for genome analysis. Science 1998, 282, 662–682. [Google Scholar] [CrossRef] [Green Version]
- Gordon-Kamm, W.J.; Spencer, T.M.; Mangano, M.L.; Adams, T.R.; Daines, R.J.; Start, W.G.; O’Brien, J.V.; Chambers, S.A.; Adams, W.R.; Willetts, N.G. Transformation of maize cells and regeneration of fertile transgenic plants. Plant Cell 1990, 2, 603–618. [Google Scholar] [CrossRef]
- McCabe, D.E.; Swain, W.F.; Martinell, B.J.; Christou, P. Stable transformation of soybean (Glycine max) by particle acceleration. Bio/Technology 1988, 6, 923. [Google Scholar] [CrossRef]
- Umbeck, P.; Johnson, G.; Barton, K.; Swain, W. Genetically transformed cotton (Gossypium hirsutum L.) plants. Bio/Technology 1987, 5, 263. [Google Scholar] [CrossRef]
- Cerdeira, A.L.; Duke, S.O. The current status and environmental impacts of glyphosate-resistant crops. J. Environ. Qual. 2006, 35, 1633–1658. [Google Scholar] [CrossRef]
- Raven, P.H. Does the use of transgenic plants diminish or promote biodiversity? New Biotechnol. 2010, 27, 528–533. [Google Scholar] [CrossRef] [PubMed]
- ISAAA. Global Status of Commercialized Biotech/GM Crops in 2019: Biotech Crops Drive Socio-Economic Development and Sustainable Environment in the New Frontier; ISAAA Brief No. 55; ISAAA Ithaca: New York, NY, USA, 2019. [Google Scholar]
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef]
- Boter, M.; Ruíz-Rivero, O.; Abdeen, A.; Prat, S. Conserved MYC transcription factors play a key role in jasmonate signaling both in tomato and Arabidopsis. Genes Dev. 2004, 18, 1577–1591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, J.-S.; Joo, J.; Kim, M.-J.; Kim, Y.-K.; Nahm, B.H.; Song, S.I.; Cheong, J.-J.; Lee, J.S.; Kim, J.-K.; Choi, Y.D. OsbHLH148, a basic helix-loop-helix protein, interacts with OsJAZ proteins in a jasmonate signaling pathway leading to drought tolerance in rice. Plant J. 2011, 65, 907–921. [Google Scholar] [CrossRef] [PubMed]
- Nambara, E.; Marion-Poll, A. Abscisic Acid Biosynthesis and Catabolism. Annu. Rev. Plant Biol. 2005, 56, 165–185. [Google Scholar] [CrossRef] [Green Version]
- Finkelstein, R. Abscisic Acid synthesis and response. Arab. Book 2013, 11, e0166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deinlein, U.; Stephan, A.B.; Horie, T.; Luo, W.; Xu, G.; Schroeder, J.I. Plant salt-tolerance mechanisms. Trends Plant Sci. 2014, 19, 371–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamamoto, S.; Horie, T.; Hauser, F.; Deinlein, U.; Schroeder, J.I.; Uozumi, N. HKT transporters mediate salt stress resistance in plants: From structure and function to the field. Curr. Opin. Biotechnol. 2015, 32, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Le Page, M. The second great food war. New Sci. 2018, 239, 22–23. [Google Scholar] [CrossRef]
- Gaj, T.; Sirk, S.J.; Shui, S.-L.; Liu, J. Genome-editing technologies: Principles and applications. Cold Spring Harb. Perspect. Biol. 2016, 8, a023754. [Google Scholar] [CrossRef] [Green Version]
- Ahmar, S.; Saeed, S.; Khan, M.H.U.; Ullah Khan, S.; Mora-Poblete, F.; Kamran, M.; Faheem, A.; Maqsood, A.; Rauf, M.; Saleem, S. A Revolution toward Gene-Editing Technology and Its Application to Crop Improvement. Int. J. Mol. Sci. 2020, 21, 5665. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Si, X.; Ji, X.; Fan, R.; Liu, J.; Chen, K.; Wang, D.; Gao, C. Genome editing of upstream open reading frames enables translational control in plants. Nat. Biotechnol. 2018, 36, 894–898. [Google Scholar] [CrossRef] [PubMed]
- Xing, S.; Chen, K.; Zhu, H.; Zhang, R.; Zhang, H.; Li, B.; Gao, C. Fine-tuning sugar content in strawberry. BMC Genome Biol. 2020, 21, 230. [Google Scholar]
- Kim, E.H.; Kim, Y.S.; Park, S.-H.; Koo, Y.J.; Do Choi, Y.; Chung, Y.-Y.; Lee, I.-J.; Kim, J.-K. Methyl jasmonate reduces grain yield by mediating stress signals to alter spikelet development in rice. Plant Physiol. 2009, 149, 1751–1760. [Google Scholar] [CrossRef] [Green Version]
- Si, X.; Zhang, H.; Wang, Y.; Chen, K.; Gao, C. Manipulating gene translation in plants by CRISPR–Cas9-mediated genome editing of upstream open reading frames. Nat. Protoc. 2020, 15, 338–363. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Um, T.; Park, T.; Shim, J.S.; Kim, Y.S.; Lee, G.-S.; Choi, I.-Y.; Kim, J.-K.; Seo, J.S.; Park, S.C. Application of Upstream Open Reading Frames (uORFs) Editing for the Development of Stress-Tolerant Crops. Int. J. Mol. Sci. 2021, 22, 3743. https://doi.org/10.3390/ijms22073743
Um T, Park T, Shim JS, Kim YS, Lee G-S, Choi I-Y, Kim J-K, Seo JS, Park SC. Application of Upstream Open Reading Frames (uORFs) Editing for the Development of Stress-Tolerant Crops. International Journal of Molecular Sciences. 2021; 22(7):3743. https://doi.org/10.3390/ijms22073743
Chicago/Turabian StyleUm, Taeyoung, Taehyeon Park, Jae Sung Shim, Youn Shic Kim, Gang-Seob Lee, Ik-Young Choi, Ju-Kon Kim, Jun Sung Seo, and Soo Chul Park. 2021. "Application of Upstream Open Reading Frames (uORFs) Editing for the Development of Stress-Tolerant Crops" International Journal of Molecular Sciences 22, no. 7: 3743. https://doi.org/10.3390/ijms22073743
APA StyleUm, T., Park, T., Shim, J. S., Kim, Y. S., Lee, G. -S., Choi, I. -Y., Kim, J. -K., Seo, J. S., & Park, S. C. (2021). Application of Upstream Open Reading Frames (uORFs) Editing for the Development of Stress-Tolerant Crops. International Journal of Molecular Sciences, 22(7), 3743. https://doi.org/10.3390/ijms22073743