
The OSMR Gene Is Involved in Hirschsprung Associated Enterocolitis Susceptibility through an Altered Downstream Signaling

 , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Results
2.1. Variants Detection by Whole-Exome Sequencing
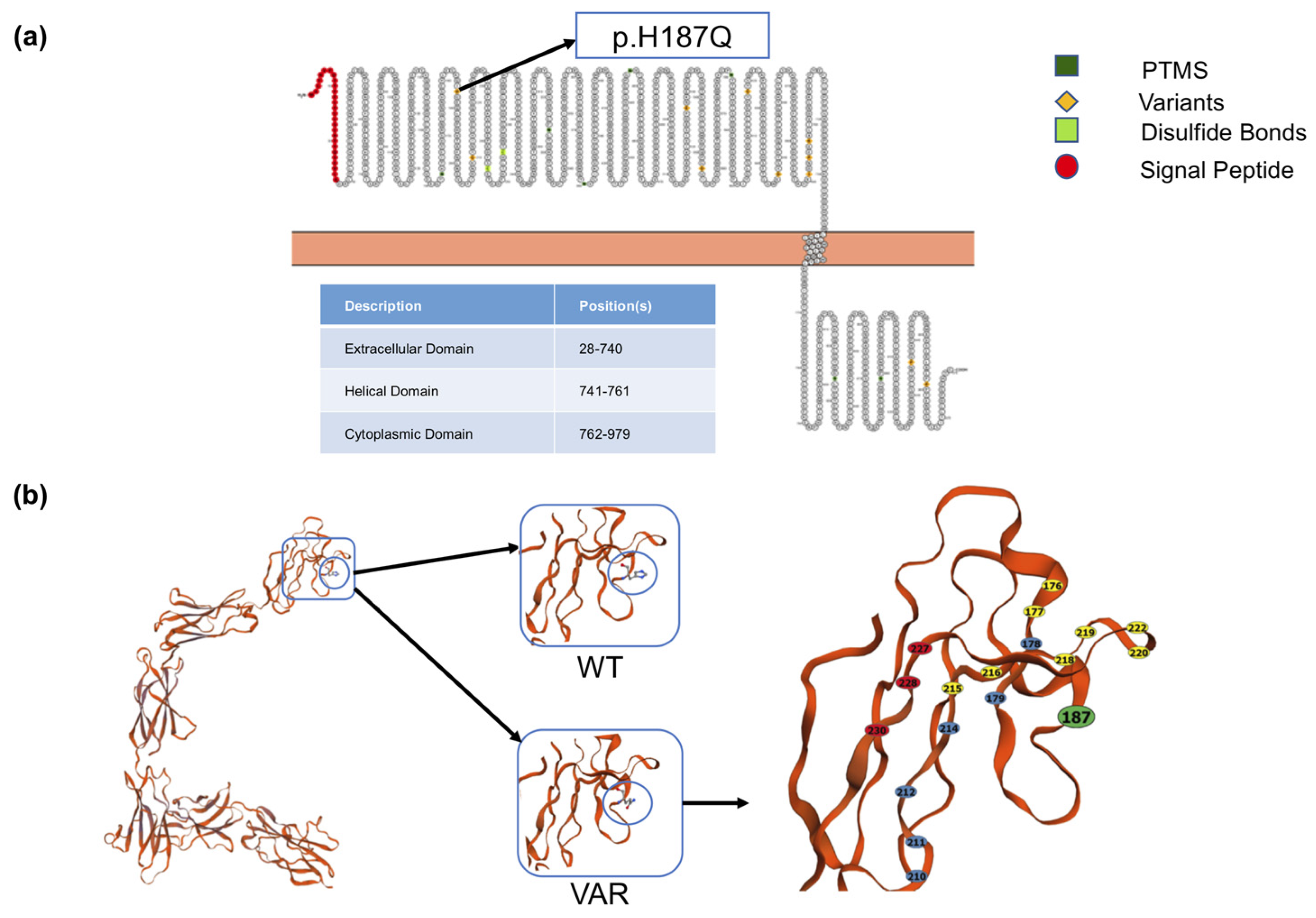
2.2. In Silico Analysis of the Effects of rs34675408 on the OSMR Structure
2.3. OSMR Expression in HAEC Lymphobastoid Cell Lines
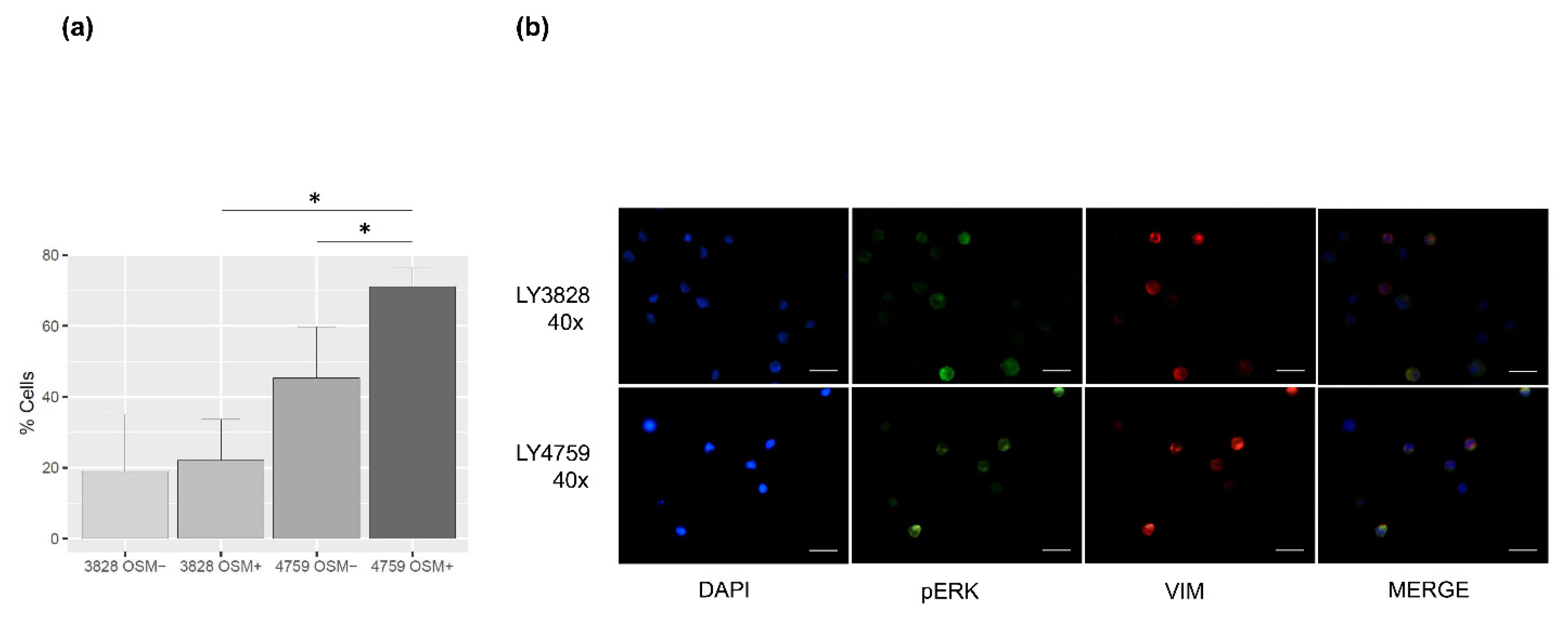
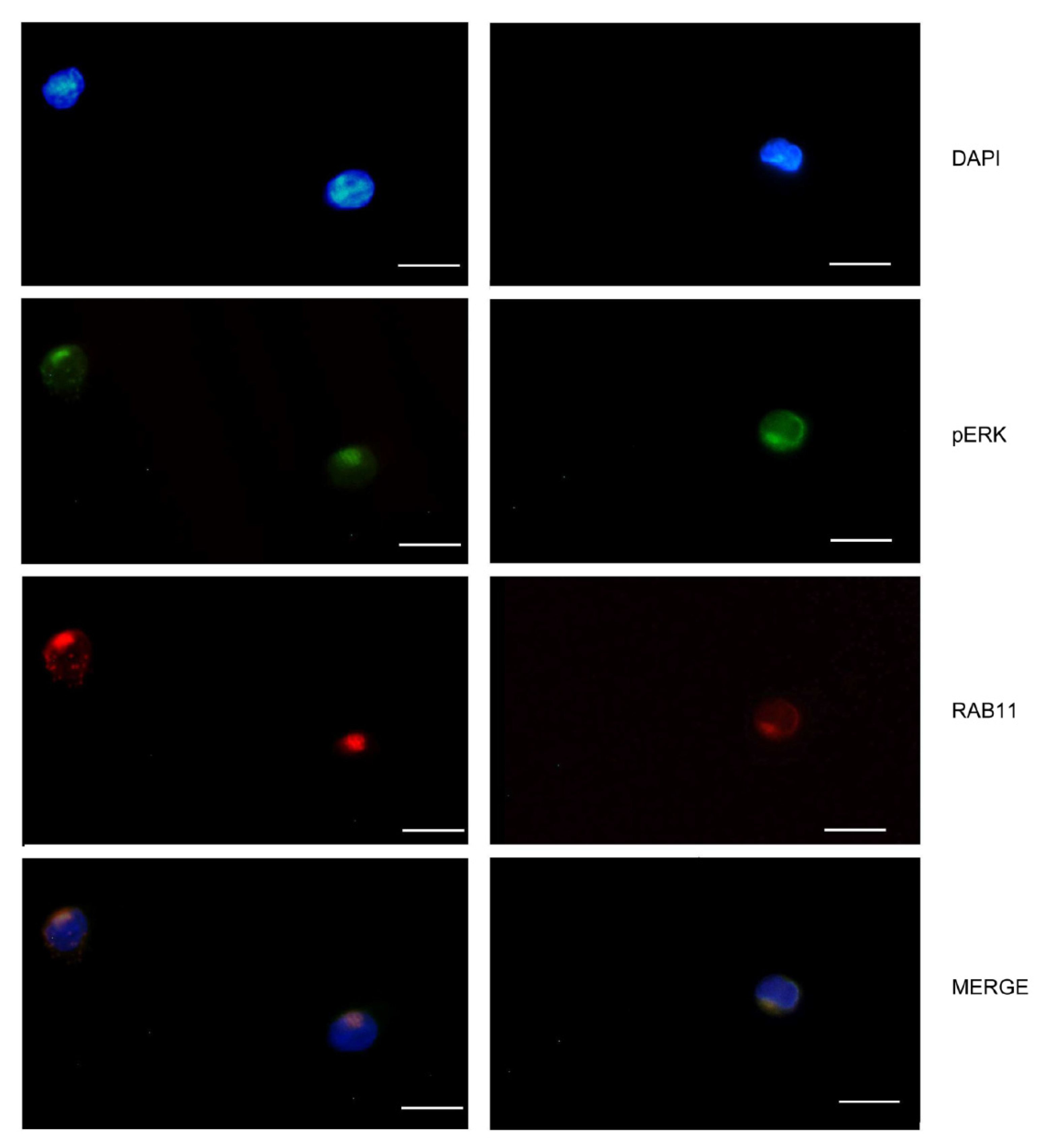
2.4. Phospho-ERK Cellular Distribution Is Induced by OSMR Stimulation in HSCR and HAEC Lymphoblasts
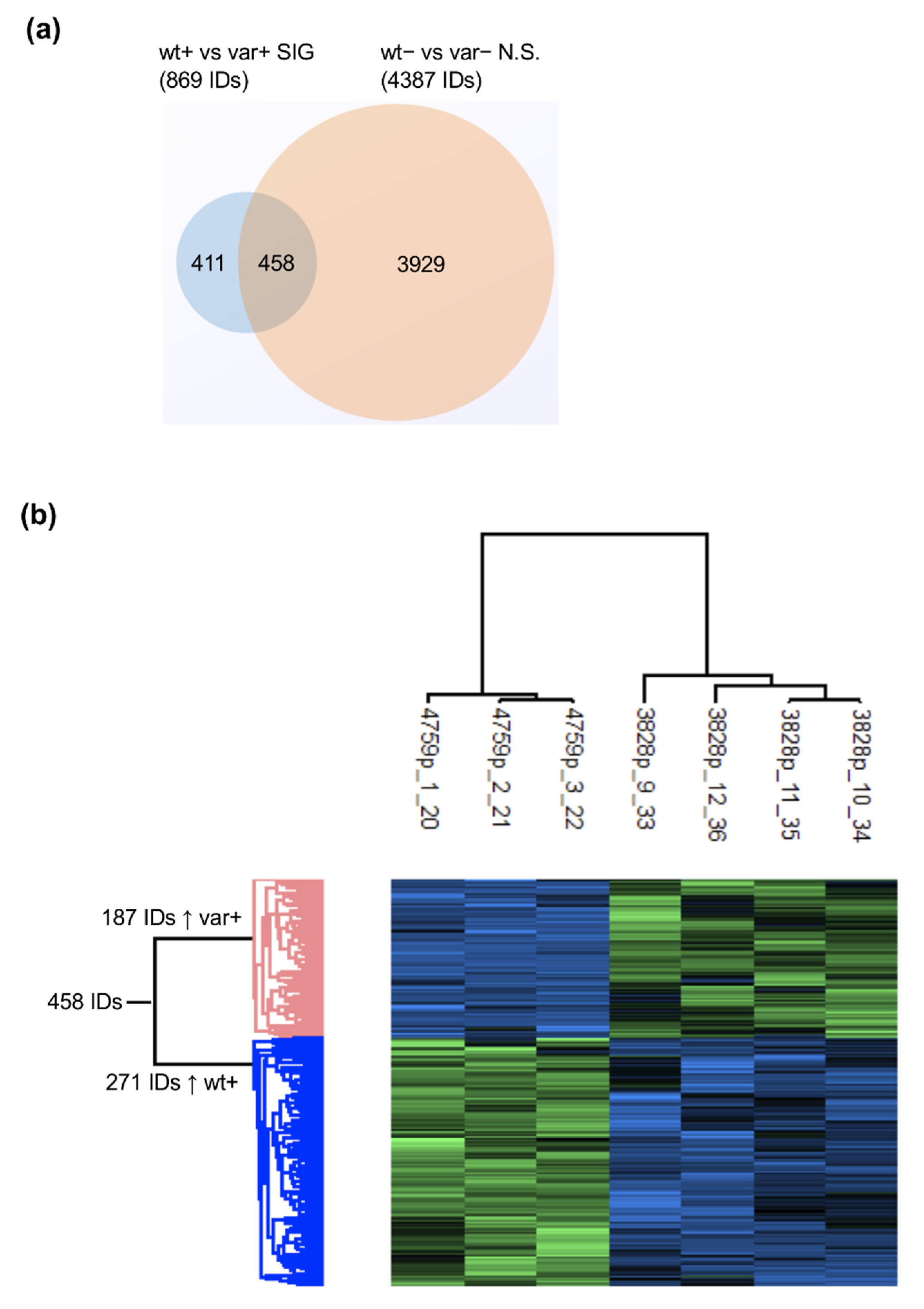
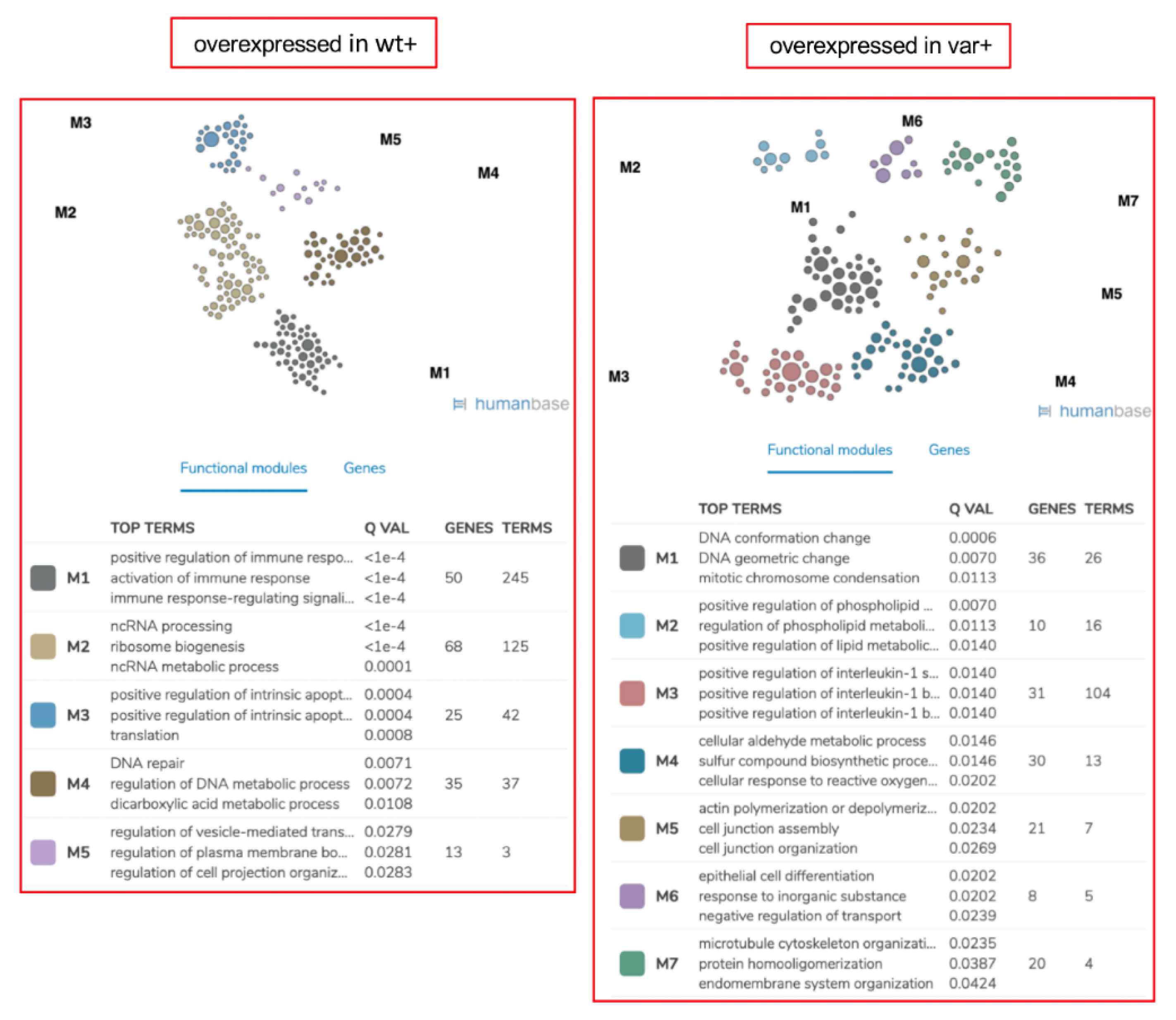
2.5. Proteomics and Pathway Analyses Reveal Pathway Perturbations Driven by the OSMR Variant
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Whole-Exome Sequencing and Variants Filtering, Prioritization, and Validation
4.3. In Silico Analyses of OSMR
4.4. Cell Cultures Preparation
4.5. Western Blotting Assay
4.6. Immunofluorescence
4.7. Sample Preparation, NanoLC, and Mass Spectrometer Setup
4.8. Statistical Analysis
4.8.1. Genetic Analysis
4.8.2. ERK Activation
4.8.3. Mass Spectrometry
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Pini Prato, A.; Rossi, V.; Mosconi, M.; Holm, C.; Lantieri, F.; Griseri, P.; Ceccherini, I.; Mavilio, D.; Jasonni, V.; Tuo, G.; et al. A prospective observational study of associated anomalies in Hirschsprung’s disease. Orphanet J. Rare Dis. 2013, 8. [Google Scholar] [CrossRef] [Green Version]
- Emison, E.; Garcia-Barcelo, M.; Grice, E.; Lantieri, F.; Amiel, J.; Burzynski, G.; Fernandez, R.; Hao, L.; Kashuk, C.; West, K.; et al. Differential contributions of rare and common, coding and noncoding Ret mutations to multifactorial Hirschsprung disease liability. Am. J. Hum. Genet. 2010, 87, 60–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lantieri, F.; Griseri, P.; Puppo, F.; Campus, R.; Martucciello, G.; Ravazzolo, R.; Devoto, M.; Ceccherini, I. Haplotypes of the human RET proto-oncogene associated with Hirschsprung disease in the Italian population derive from a single ancestral combination of alleles. Ann. Hum. Genet. 2006, 70, 12–26. [Google Scholar] [CrossRef] [PubMed]
- Amiel, J.; Sproat-Emison, E.; Garcia-Barcelo, M.; Lantieri, F.; Burzynski, G.; Borrego, S.; Pelet, A.; Arnold, S.; Miao, X.; Griseri, P.; et al. Hirschsprung disease, associated syndromes and genetics: A review. J. Med Genet. 2008, 45, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Q.; Arnold, S.; Heanue, T.; Kilambi, K.; Doan, B.; Kapoor, A.; Ling, A.; Sosa, M.; Guy, M.; Jiang, Q.; et al. Functional loss of semaphorin 3C and/or semaphorin 3D and their epistatic interaction with ret are critical to Hirschsprung disease liability. Am. J. Hum. Genet. 2015, 96, 581–596. [Google Scholar] [CrossRef] [Green Version]
- Tang, C.; Gui, H.; Kapoor, A.; Kim, J.; Luzón-Toro, B.; Pelet, A.; Burzynski, G.; Lantieri, F.; So, M.; Berrios, C.; et al. Trans-ethnic meta-analysis of genome-wide association studies for Hirschsprung disease. Hum. Mol. Genet. 2016, 25, 5265–5275. [Google Scholar] [CrossRef] [Green Version]
- Gosain, A.; Barlow-Anacker, A.; Erickson, C.; Pierre, J.; Heneghan, A.; Epstein, M.; Kudsk, K. Impaired Cellular Immunity in the Murine Neural Crest Conditional Deletion of Endothelin Receptor-B Model of Hirschsprung’s Disease. PLoS ONE 2015, 10, e0128822. [Google Scholar] [CrossRef]
- Allaire, J.; Crowley, S.; Law, H.; Chang, S.; Ko, H.; Vallance, B. The Intestinal Epithelium: Central Coordinator of Mucosal Immunity. Trends Immunol. 2018, 39, 677–696. [Google Scholar] [CrossRef]
- Nakamura, H.; Lim, T.; Puri, P. Inflammatory bowel disease in patients with Hirschsprung’s disease: A systematic review and meta-analysis. Pediatric Surg. Int. 2018, 34, 149–154. [Google Scholar] [CrossRef]
- Thiagarajah, J.; Yildiz, H.; Carlson, T.; Thomas, A.; Steiger, C.; Pieretti, A.; Zukerberg, L.; Carrier, R.; Goldstein, A. Altered goblet cell differentiation and surface mucus properties in Hirschsprung disease. PLoS ONE 2014, 9, e99944. [Google Scholar] [CrossRef]
- Zambetti, L.P.; Mortellaro, A. NLRPs, microbiota, and gut homeostasis: Unravelling the connection. J. Pathol. 2020, 233, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Pini Prato, A.; Bartow-McKenney, C.; Hudspeth, K.; Mosconi, M.; Rossi, V.; Avanzini, S.; Faticato, M.G.; Ceccherini, I.; Lantieri, F.; Mattioli, G.; et al. A Metagenomics Study on Hirschsprung’s Disease Associated Enterocolitis: Biodiversity and Gut Microbial Homeostasis Depend on Resection Length and Patient’s Clinical History. Front. Pediatrics 2019, 7. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Poroyko, V.; Yan, Z.; Pan, L.; Feng, Y.; Zhao, P.; Xie, Z.; Hong, L. Characterization of Intestinal Microbiomes of Hirschsprung’s Disease Patients with or without Enterocolitis Using Illumina-MiSeq High-Throughput Sequencing. PLoS ONE 2016, 11, e0162079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Z.; Poroyko, V.; Gu, S.; Zhang, Z.; Pan, L.; Wang, J.; Bao, N.; Hong, L. Characterization of the intestinal microbiome of Hirschsprung’s disease with and without enterocolitis. Biochem. Biophys. Res. Commun. 2014, 445, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Porokuokka, L.; Virtanen, H.; Lindén, J.; Sidorova, Y.; Danilova, T.; Lindahl, M.; Saarma, M.; Andressoo, J. Gfra1 Underexpression Causes Hirschsprung’s Disease and Associated Enterocolitis in Mice. Cell. Mol. Gastroenterol. Hepatol. 2019, 7, 655–678. [Google Scholar] [CrossRef] [Green Version]
- Perea, D.; Guiu, J.; Hudry, B.; Konstantinidou, C.; Milona, A.; Hadjieconomou, D.; Carroll, T.; Hoyer, N.; Natarajan, D.; Kallijärvi, J.; et al. Ret receptor tyrosine kinase sustains proliferation and tissue maturation in intestinal epithelia. EMBO J. 2017, 36, 3029–3045. [Google Scholar] [CrossRef] [Green Version]
- Rusmini, M.; Griseri, P.; Lantieri, F.; Matera, I.; Hudspeth, K.; Roberto, A.; Mikulak, J.; Avanzini, S.; Rossi, V.; Mattioli, G.; et al. Induction of RET dependent and independent pro-inflammatory programs in human peripheral blood mononuclear cells from Hirschsprung patients. PLoS ONE 2013, 8, e59066. [Google Scholar] [CrossRef]
- Veiga-Fernandes, H.; Coles, M.; Foster, K.; Patel, A.; Williams, A.; Natarajan, D.; Barlow, A.; Pachnis, V.; Kioussis, D. Tyrosine kinase receptor RET is a key regulator of Peyer’s patch organogenesis. Nature 2007, 446, 547–551. [Google Scholar] [CrossRef]
- Du, Q.; Qian, Y.; Xue, W. Molecular Simulation of Oncostatin M and Receptor (OSM-OSMR) Interaction as a Potential Therapeutic Target for Inflammatory Bowel Disease. Front. Mol. Biosci. 2020, 7. [Google Scholar] [CrossRef] [Green Version]
- Biasini, M.; Bienert, S.; Waterhouse, A.; Arnold, K.; Studer, G.; Schmidt, T.; Kiefer, F.; Gallo Cassarino, T.; Bertoni, M.; Bordoli, L.; et al. SWISS-MODEL: Modelling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Res. 2014, 42, W252–W258. [Google Scholar] [CrossRef]
- Sakai, R.; Fukuda, R.; Unida, S.; Aki, M.; Ono, Y.; Endo, A.; Kusumi, S.; Koga, D.; Fukushima, T.; Komada, M.; et al. The integral function of the endocytic recycling compartment is regulated by RFFL-mediated ubiquitylation of Rab11 effectors. J. Cell Sci. 2019, 132, jcs228007. [Google Scholar] [CrossRef] [Green Version]
- Kapp-Barnea, Y.; Ninio-Many, L.; Hirschberg, K.; Fukuda, M.; Jeromin, A.; Sagi-Eisenberg, R. Neuronal calcium sensor-1 and phosphatidylinositol 4-kinase beta stimulate extracellular signal-regulated kinase 1/2 signaling by accelerating recycling through the endocytic recycling compartment. Mol. Biol. Cell 2006, 17, 4130–4141. [Google Scholar] [CrossRef] [Green Version]
- Dumas, A.; Lagarde, S.; Laflamme, C.; Pouliot, M. Oncostatin M decreases interleukin-1 β secretion by human synovial fibroblasts and attenuates an acute inflammatory reaction in vivo. J. Cell Mol. Med. 2012, 16, 1274–1285. [Google Scholar] [CrossRef]
- Hermanns, H. Oncostatin M and interleukin-31: Cytokines, receptors, signal transduction and physiology. Cytokine Growth Factor Rev. 2015, 26, 545–558. [Google Scholar] [CrossRef]
- Li, H.; Feng, C.; Fan, C.; Yang, Y.; Yang, X.; Lu, H.; Lu, Q.; Zhu, F.; Xiang, C.; Zhang, Z.; et al. Intervention of oncostatin M-driven mucosal inflammation by berberine exerts therapeutic property in chronic ulcerative colitis. Cell Death Dis. 2020, 11, 271. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, F.; Xie, Y.; Yang, Q.; Luo, H.; Jia, P.; Shi, Z.; Wang, S.; Zheng, X. Feiyangchangweiyan capsule protects against ulcerative colitis in mice by modulating the OSM/OSMR pathway and improving gut microbiota. Phytomed. Int. J. Phytother. Phytopharm. 2021, 80. [Google Scholar] [CrossRef]
- Beigel, F.; Friedrich, M.; Probst, C.; Sotlar, K.; Göke, B.; Diegelmann, J.; Brand, S. Oncostatin M mediates STAT3-dependent intestinal epithelial restitution via increased cell proliferation, decreased apoptosis and upregulation of SERPIN family members. PLoS ONE 2014, 9, e93498. [Google Scholar] [CrossRef]
- West, N.; Hegazy, A.; Owens, B.; Bullers, S.; Linggi, B.; Buonocore, S.; Coccia, M.; Görtz, D.; This, S.; Stockenhuber, K.; et al. Oncostatin M drives intestinal inflammation and predicts response to tumor necrosis factor-neutralizing therapy in patients with inflammatory bowel disease. Nat. Med. 2017, 23, 579–589. [Google Scholar] [CrossRef]
- Pothoven, K.; Schleimer, R. The barrier hypothesis and Oncostatin M: Restoration of epithelial barrier function as a novel therapeutic strategy for the treatment of type 2 inflammatory disease. Tissue Barriers 2017, 5, e1341367. [Google Scholar] [CrossRef]
- Spehlmann, M.; Eckmann, L. Nuclear factor-kappa B in intestinal protection and destruction. Curr. Opin. Gastroenterol. 2009, 25, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, T.; Fujita, N.; Jang, M.; Uematsu, S.; Yang, B.; Satoh, T.; Omori, H.; Noda, T.; Yamamoto, N.; Komatsu, M.; et al. Loss of the autophagy protein Atg16L1 enhances endotoxin-induced IL-1beta production. Nature 2008, 456, 264–268. [Google Scholar] [CrossRef]
- Bachetti, T.; Ceccherini, I. Tumor necrosis factor receptor-associated periodic syndrome as a model linking autophagy and inflammation in protein aggregation diseases. J. Mol. Med. (Berl. Ger.) 2014, 92, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Carbonero, F.; Benefiel, A.; Gaskins, H. Contributions of the microbial hydrogen economy to colonic homeostasis. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 504–518. [Google Scholar] [CrossRef]
- Demehri, F.; Frykman, P.; Cheng, Z.; Ruan, C.; Wester, T.; Nordenskjöld, A.; Kawaguchi, A.; Hui, T.; Granström, A.; Funari, V.; et al. Altered fecal short chain fatty acid composition in children with a history of Hirschsprung-associated enterocolitis. J. Pediatric Surg. 2016, 51, 81–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickert, G.; Neufert, C.; Leppkes, M.; Zheng, Y.; Wittkopf, N.; Warntjen, M.; Lehr, H.; Hirth, S.; Weigmann, B.; Wirtz, S.; et al. STAT3 links IL-22 signaling in intestinal epithelial cells to mucosal wound healing. J. Exp. Med. 2009, 206, 1465–1472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Cheng, J.; Zou, S.; D’Souza, A.; Koff, J.; Lu, J.; Lee, P.; Krause, D.; Egan, M.; Bruscia, E. Pharmacological modulation of the AKT/microRNA-199a-5p/CAV1 pathway ameliorates cystic fibrosis lung hyper-inflammation. Nat. Commun. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Rathor, N.; Chung, H.K.; Wang, S.R.; Wang, J.; Turner, D.J.; Rao, J.N. Caveolin-1 enhances rapid mucosal restitution by activating TRPC1-mediated Ca2+ signaling. Physiol. Rep. 2014, e12193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, H.; O’Donnell, A.; Tomuschat, C.; Coyle, D.; Puri, P. Altered expression of caveolin-1 in the colon of patients with Hirschsprung’s disease. Pediatric Surg. Int. 2019, 35, 929–934. [Google Scholar] [CrossRef]
- Lu, N.; Malemud, C. Extracellular Signal-Regulated Kinase: A Regulator of Cell Growth, Inflammation, Chondrocyte and Bone Cell Receptor-Mediated Gene Expression. Int. J. Mol. Sci. 2019, 20, 3792. [Google Scholar] [CrossRef] [Green Version]
- Pastor, A.; Osman, F.; Teitelbaum, D.; Caty, M.; Langer, J. Development of a standardized definition for Hirschsprung’s-associated enterocolitis: A Delphi analysis. J. Pediatric Surg. 2009, 44, 251–256. [Google Scholar] [CrossRef]
- Elhalaby, E.; Coran, A.; Blane, C.; Hirschl, R.; Teitelbaum, D. Enterocolitis associated with Hirschsprung’s disease: A clinical-radiological characterization based on 168 patients. J. Pediatric Surg. 1995, 30, 76–83. [Google Scholar] [CrossRef]
- Fuentes Fajardo, K.; Adams, D.; Mason, C.; Sincan, M.; Tifft, C.; Toro, C.; Boerkoel, C.; Gahl, W.; Markello, T. Detecting false-positive signals in exome sequencing. Hum. Mutat. 2012, 33, 609–613. [Google Scholar] [CrossRef] [Green Version]
- Omasits, U.; Ahrens, C.; Müller, S.; Wollscheid, B. Protter: Interactive protein feature visualization and integration with experimental proteomic data. Bioinform. (Oxf. Engl. ) 2014, 30, 884–886. [Google Scholar] [CrossRef] [Green Version]
- Migita, K.; Komori, A.; Torigoshi, T.; Maeda, Y.; Izumi, Y.; Jiuchi, Y.; Miyashita, T.; Nakamura, M.; Motokawa, S.; Ishibashi, H. CP690,550 inhibits oncostatin M-induced JAK/STAT signaling pathway in rheumatoid synoviocytes. Arthritis Res. Ther. 2011, 13, R72. [Google Scholar] [CrossRef] [Green Version]
- Tsang, M.; Gantchev, J.; Ghazawi, F.; Litvinov, I. Protocol for adhesion and immunostaining of lymphocytes and other non-adherent cells in culture. BioTechniques 2017, 63, 230–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulak, N.; Pichler, G.; Paron, I.; Nagaraj, N.; Mann, M. Minimal, encapsulated proteomic-sample processing applied to copy-number estimation in eukaryotic cells. Nat. Methods 2014, 11, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef]
- Greene, C.; Krishnan, A.; Wong, A.; Ricciotti, E.; Zelaya, R.; Himmelstein, D.; Zhang, R.; Hartmann, B.; Zaslavsky, E.; Sealfon, S.; et al. Understanding multicellular function and disease with human tissue-specific networks. Nat. Genet. 2015, 47, 569–576. [Google Scholar] [CrossRef] [Green Version]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NGS | Replicate | Total | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Rank | Chromosomal Position | Gene | AA | rs# | REF/ALT | gnomAD ° | MAF | MAF | Genotypes (omo/HT/wt) | MAF | p Value * | |||
| Cases ^ | Cases | Controls | Cases | Controls | Cases | Controls | ||||||||
| 1 | chr19:17940842 | JAK3 | p.Ala1094Ala | rs3212780 | G/A | 0.281 | 0.375 | 0.250 | 0.321 | 3/14/17 | 3/21/30 | 0.294 | 0.250 | 0.6002 |
| 2 | chr21:48069682 | PRMT2 | p.Arg229Trp | rs76937225 § | C/T | 0.039 | 0.292 | - | - | - | - | - | - | - |
| 3–4 | chr5:38884071 | OSMR | p.His187Gln | rs34675408 | T/G | 0.070 | 0.292 | 0.117 | 0.057 | 2/17/53 | 0/11/97 | 0.146 | 0.051 | 0.0024 |
| 3–4 | chr2:209190632 | PIKFYVE | p.Ser1033Ala | rs999890 | T/G | 0.140 | 0.292 | 0.150 | 0.143 | 2/9/21 | 0/6/27 | 0.203 | 0.091 | 0.0850 |
| 5–6 | chr11:7059981 | NLRP14 | p.Arg55Gln | rs61063081 | G/A | 0.207 | 0.333 | 0.206 | 0.268 | 1/13/15 | 5/12/36 | 0.259 | 0.208 | 0.5581 |
| 5–6 | chr11:7091569 | NLRP14 | p.Leu1010Phe | rs17280682 § | C/T | 0.207 | 0.333 | - | - | - | - | - | - | - |
| CASES (HAEC) | |||||||
| Gene | SNP | REF | ALT | T:U * | OR (95%CI) | Chi-Square | p-Value |
| OSMR | rs34675408 | T | G | 11:2 | 5.5 (1.2–24.8) | 6.231 | 0.0126 |
| CTRL (HSCR only) | |||||||
| OSMR | rs34675408 | T | G | 2:5 | 0.4 (0.1–2.1) | 1.286 | 0.257 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bachetti, T.; Rosamilia, F.; Bartolucci, M.; Santamaria, G.; Mosconi, M.; Sartori, S.; De Filippo, M.R.; Di Duca, M.; Obino, V.; Avanzini, S.; et al. The OSMR Gene Is Involved in Hirschsprung Associated Enterocolitis Susceptibility through an Altered Downstream Signaling. Int. J. Mol. Sci. 2021, 22, 3831. https://doi.org/10.3390/ijms22083831
Bachetti T, Rosamilia F, Bartolucci M, Santamaria G, Mosconi M, Sartori S, De Filippo MR, Di Duca M, Obino V, Avanzini S, et al. The OSMR Gene Is Involved in Hirschsprung Associated Enterocolitis Susceptibility through an Altered Downstream Signaling. International Journal of Molecular Sciences. 2021; 22(8):3831. https://doi.org/10.3390/ijms22083831
Chicago/Turabian StyleBachetti, Tiziana, Francesca Rosamilia, Martina Bartolucci, Giuseppe Santamaria, Manuela Mosconi, Serenella Sartori, Maria Rosaria De Filippo, Marco Di Duca, Valentina Obino, Stefano Avanzini, and et al. 2021. "The OSMR Gene Is Involved in Hirschsprung Associated Enterocolitis Susceptibility through an Altered Downstream Signaling" International Journal of Molecular Sciences 22, no. 8: 3831. https://doi.org/10.3390/ijms22083831
APA StyleBachetti, T., Rosamilia, F., Bartolucci, M., Santamaria, G., Mosconi, M., Sartori, S., De Filippo, M. R., Di Duca, M., Obino, V., Avanzini, S., Mavilio, D., Candiani, S., Petretto, A., Pini Prato, A., Ceccherini, I., & Lantieri, F. (2021). The OSMR Gene Is Involved in Hirschsprung Associated Enterocolitis Susceptibility through an Altered Downstream Signaling. International Journal of Molecular Sciences, 22(8), 3831. https://doi.org/10.3390/ijms22083831