
Acid-Sphingomyelinase Triggered Fluorescently Labeled Sphingomyelin Containing Liposomes in Tumor Diagnosis after Radiation-Induced Stress

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
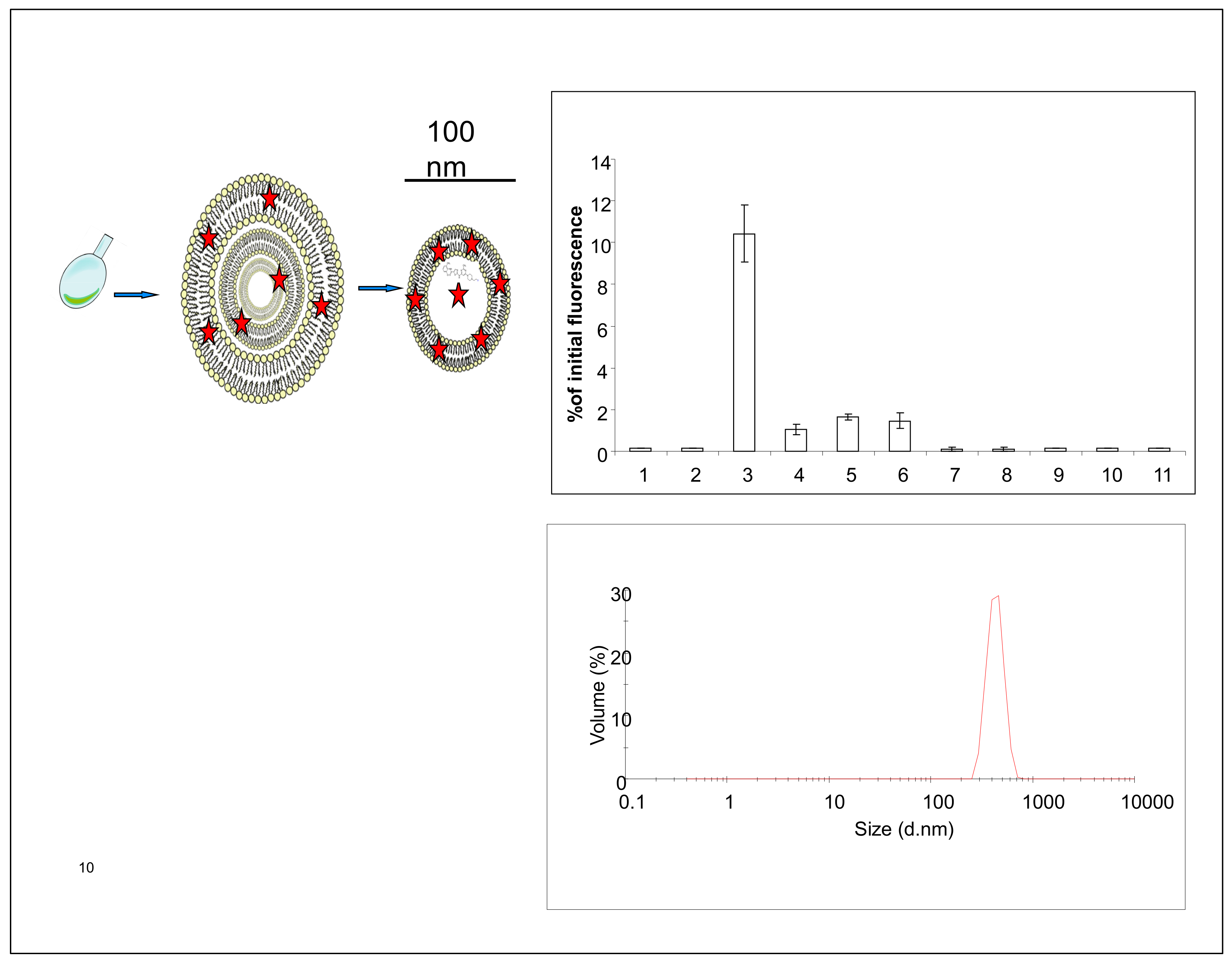
2.1. Liposome Preparation
2.2. In Vitro Release Assay
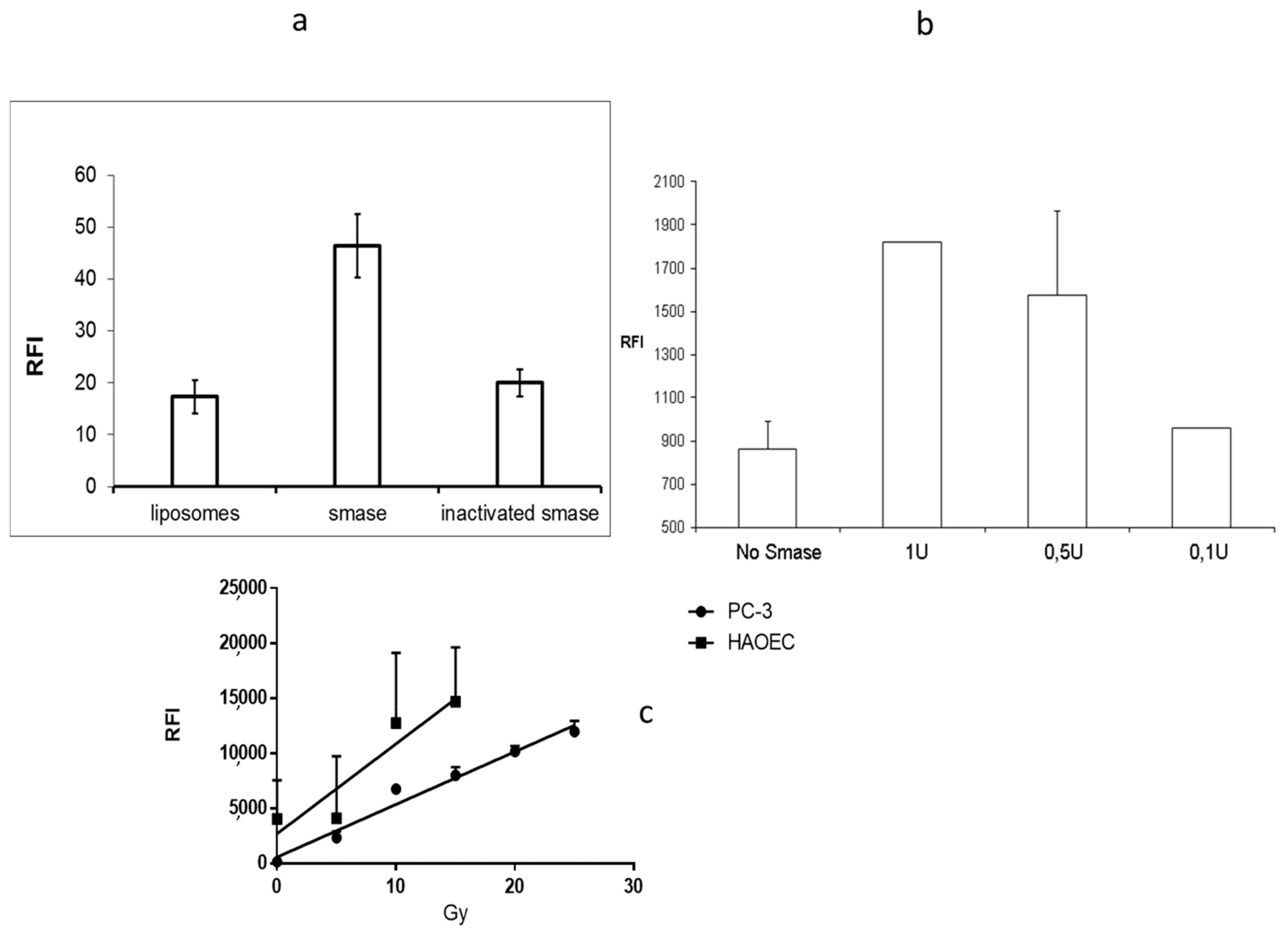
2.3. In Vitro Acid Sphingomyelinase Activity
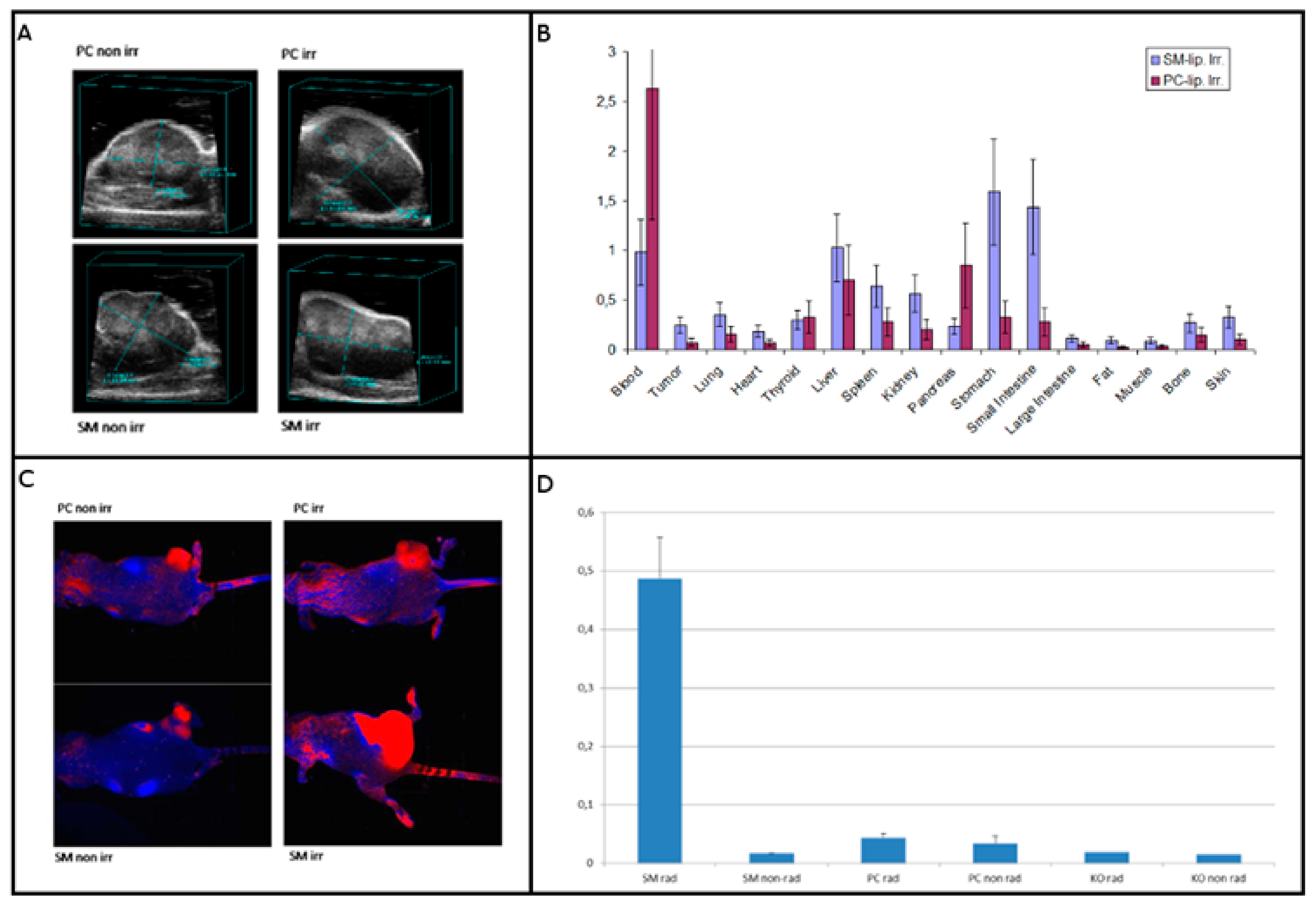
2.4. Testing Liposomes in Tumor Models
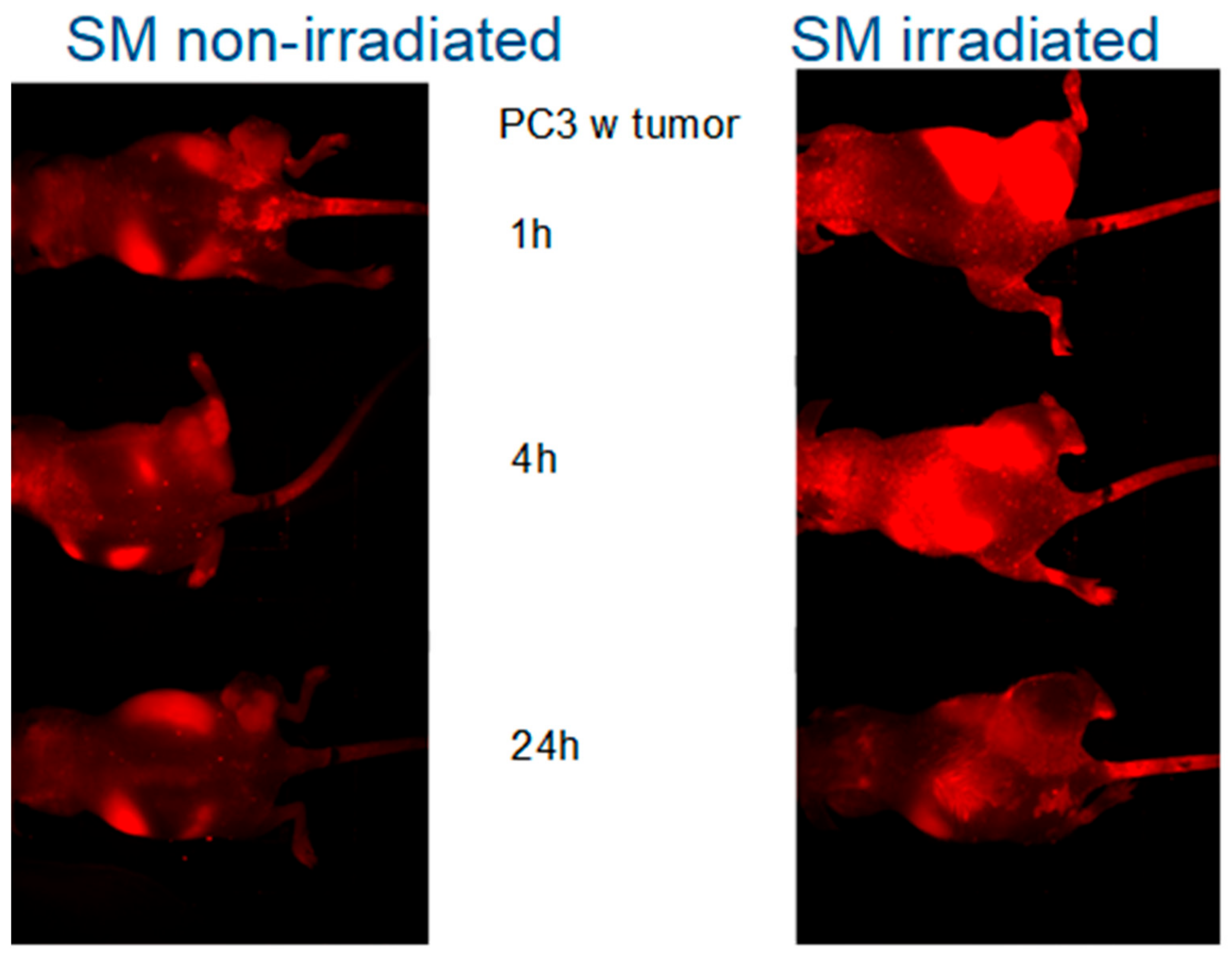
2.5. Fluorescence Imaging of SM-Liposomes
3. Discussion
4. Materials and Methods
4.1. Liposomal Preparation
4.2. Mouse Tumor Model
4.3. Tumor Irradiation and SM-Liposome Biodistribution
4.4. Fluorescence Imaging
5. Patents
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SM | Sphingomyelin |
| PC | phosphatidyl choline |
| GUV | giant unilamellar vesicle |
| EPR | enhanced permeability and retention |
| SMase | Sphingomyelinase |
| PET | positron emission tomography |
| MRI | magnetic resonance imaging |
| DLS | dynamic light scattering |
| HAoEC | human aortic endothelial cells |
| PC-3 | Caucasian prostate adenocarcinoma |
References
- Heneweer, C.; Gendy, S.E.; Peñate-Medina, O. Liposomes and inorganic nanoparticles for drug delivery and cancer imaging. Ther. Deliv. 2012, 3, 645–656. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, P.; Shi, J.; Hao, Y.; Meng, D.; Zhao, Y.; Yanyan, Y.; Li, D.; Chang, J.; Zhang, Z. Radiofrequency-triggered tumor-targeting delivery system for theranostics application. ACS Appl. Mater. Interfaces 2015, 18, 5736–5747. [Google Scholar] [CrossRef] [PubMed]
- Rothman, J.E.; Lenard, J. Membrane asymmetry. Science 1977, 195, 743–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stancevic, B.; Kolesnick, R. Ceramide-rich platforms in transmembrane signaling. FEBS Lett. 2010, 584, 1728–1740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Argüello, M.B.; Basáñez, G.; Goñi, F.M.; Alonso, A. Different effects of enzyme-generated ceramides and diacylglycerols in phospholipid membrane fusion and leakage. J. Biol. Chem. 1996, 271, 26616–26621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holopainen, J.M.; Subramanian, M.; Kinnunen, P.J. Sphingomyelinase induced lipid microdomain formation in a fluid phosphati-dylcholine/sphingomyelin membrane. Biochemistry 1998, 37, 17562–17570. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Argüello, M.B.; Goñi, F.M.; Alonso, A. Vesicle membrane fusion induced by the concerted activities of sphingomyelinase and phospholipase C. J. Biol. Chem. 1998, 273, 22977–22982. [Google Scholar] [CrossRef] [Green Version]
- Nurminen, T.A.; Holopainen, J.M.; Zhao, H.; Kinnunen, P.K. Observation of topical catalysis by sphingomyelinase coupled to microspheres. J. Am. Chem. Soc. 2002, 16, 12129–12134. [Google Scholar] [CrossRef]
- Sot, J.; Bagatolli, L.A.; Goñi, F.M.; Alonso, A. Detergent-Resistant, Ceramide-Enriched Domains in Sphingomyelin/Ceramide Bilayers. Biophys. J. 2006, 90, 903–914. [Google Scholar] [CrossRef] [Green Version]
- Goñi, F.M.; Alonso, A. Sphingomyelinases: Enzymology and membrane activity. FEBS Lett. 2002, 531, 38–46. [Google Scholar] [CrossRef] [Green Version]
- Kornhuber, J.; Rhein, C.; Müller, C.P.; Mühle, C. Secretory sphingomyelinase in health and disease. Biol. Chem. 2015, 396, 707–736. [Google Scholar] [CrossRef]
- Santana, P.; Peña, L.A.; Haimovitz-Friedman, A.; Martin, S.; Green, D.; McLoughlin, M.; Cordon-Cardo, C.; Schuchman, E.H.; Fuks, Z.; Kolesnick, R. Acid sphingomyelinase-deficient human lymphoblasts and mice are defective in radiation-induced apoptosis. Cell 1996, 86, 189–199. [Google Scholar] [CrossRef] [Green Version]
- Norma, W.A. Solving the secretory acid sphingomyelinase puzzle: Insights from lysosome-mediated parasite invasion and plasma membrane repair. Cell Microbiol. 2019, 21, e13065. [Google Scholar]
- Rotolo, J.A.; Zhang, J.; Donepudi, M.; Lee, H.; Fuks, Z.; Kolesnick, R. Caspase-dependent and -independent activation of acid sphingomyelinase signaling. J. Biol. Chem. 2005, 280, 26425–26434. [Google Scholar] [CrossRef] [Green Version]
- Peñate-Medina, T.; Kraas, E.; Luo, K.; Humbert, J.; Zhu, H.; Mertens, F.; Gerle, M.; Rohwedder, A.; Damoah, C.; Will, O.; et al. Utilizing ICG Spectroscopical Properties for Real-Time Nanoparticle Release Quantification In vitro and In vivo in Imaging Setups. Curr. Pharm. Des. 2020, 26, 3828–3833. [Google Scholar] [CrossRef]
- Penate Medina, O.; Haikola, M.; Tahtinen, M.; Simpura, I.; Kaukinen, S.; Valtanen, H.; Zhu, Y.; Kuosmanen, S.; Cao, W.; Reunanen, J.; et al. Liposomal Tumor Targeting in Drug Delivery Utilizing MMP-2- and MMP-9-Binding Ligands. J. Drug Deliv. 2011, 160515. [Google Scholar] [CrossRef]
- Swiecicki, J.-M.; Thiebaut, F.; Di Pisa, M.; Gourdin -Bertin, S.; Tailhades, J.; Mansuy, C.; Burlina, F.; Chwetzoff, S.; Trugnan, G.; Chassaing, G.; et al. How to Unveil Self-Quenched Fluorophores and Subsequently Map the Subcellular Distribution of Exogenous Peptides. Sci. Rep. 2016, 6, 20237. [Google Scholar] [CrossRef] [Green Version]
- Obeid, L.M.; Hannun, Y.A. Ceramide: A stress signal and mediator of growth suppression and apoptosis. J. Cell. Biochem. 1995, 58, 191–198. [Google Scholar] [CrossRef]
- Jan, J.T.; Chatterje, E.; Griffin, D.E. Sindbis virus entry into cells triggers apoptosis by activating sphingomyelinase, leading to the release of ceramide. J. Virol. 2000, 74, 6425–6432. [Google Scholar] [CrossRef] [Green Version]
- Grassme, H.; Gulbins, E.; Brenner, B.; Ferlin, K.; Sandhoff, K.; Harzer, K.; Lang, F.; Meyer, T.F. Acidic sphingomyelinase mediates entry of N. gonorrhoeae into nonphagocytic cells. Cell 1997, 91, 605–615. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Barros, M.; Paris, F.; Cordon-Cardo, C.; Lyden, D.; Rafii, S.; Haimovitz-Friedman, A.; Fuks, Z.; Kolesnick, R. Tumor response to radiotherapy regulated by endothelial cell apoptosis. Science 2003, 300, 1155–1159. [Google Scholar] [CrossRef] [Green Version]
- Cremesti, A.; Paris, F.; Grassmé, H.; Holler, N.; Tschopp, J.; Fuks, Z.; Gulbins, E.; Kolesnick, R. Ceramide enables fas to cap and kill. J. Biol. Chem. 2001, 276, 23954–23961. [Google Scholar] [CrossRef] [Green Version]
- Grassme, H.; Jekle, A.; Riehle, A.; Schwarz, H.; Berger, J.; Sandhoff, K.; Kolesnick, R.; Gulbins, E. CD95 signaling via ceramide-rich membrane rafts. J. Biol. Chem. 2001, 276, 20589–20596. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Rotolo, J.A.; Mesicek, J.; Penate-Medina, T.; Rimner, A.; Liao, W.C.; Yin, X.; Ragupathi, G.; Ehleiter, D.; Gulbins, E.; et al. Mitochondrial ceramide-rich macrodomains functionalize Bax upon irradiation. PLoS ONE 2011, 6, e19783. [Google Scholar] [CrossRef] [Green Version]
- Holme, M.N.; Rana, S.; Barriga, H.M.G.; Kauscher, U.; Brooks, N.J.; Stevens, M.M. A Robust Liposomal Platform for Direct Colorimetric Detection of Sphingomyelinase Enzyme and Inhibitors. ACS Nano 2018, 12, 8197–8207. [Google Scholar] [CrossRef] [Green Version]
- Aldosari, M.H.; de Vries, R.P.; Rodriguez, L.R.; Hesen, N.A.; Beztsinna, N.; van Kuilenburg, A.B.P.; Hollak, C.E.M.; Schellekens, H.; Mastrobattista, E. Liposome-targeted recombinant human acid sphingomyelinase: Production, formulation, and in vitro evaluation. Eur. J. Pharm. Biopharm. 2019, 137, 185–195. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heneweer, C.; Peñate Medina, T.; Tower, R.; Kalthoff, H.; Kolesnick, R.; Larson, S.; Peñate Medina, O. Acid-Sphingomyelinase Triggered Fluorescently Labeled Sphingomyelin Containing Liposomes in Tumor Diagnosis after Radiation-Induced Stress. Int. J. Mol. Sci. 2021, 22, 3864. https://doi.org/10.3390/ijms22083864
Heneweer C, Peñate Medina T, Tower R, Kalthoff H, Kolesnick R, Larson S, Peñate Medina O. Acid-Sphingomyelinase Triggered Fluorescently Labeled Sphingomyelin Containing Liposomes in Tumor Diagnosis after Radiation-Induced Stress. International Journal of Molecular Sciences. 2021; 22(8):3864. https://doi.org/10.3390/ijms22083864
Chicago/Turabian StyleHeneweer, Carola, Tuula Peñate Medina, Robert Tower, Holger Kalthoff, Richard Kolesnick, Steven Larson, and Oula Peñate Medina. 2021. "Acid-Sphingomyelinase Triggered Fluorescently Labeled Sphingomyelin Containing Liposomes in Tumor Diagnosis after Radiation-Induced Stress" International Journal of Molecular Sciences 22, no. 8: 3864. https://doi.org/10.3390/ijms22083864
APA StyleHeneweer, C., Peñate Medina, T., Tower, R., Kalthoff, H., Kolesnick, R., Larson, S., & Peñate Medina, O. (2021). Acid-Sphingomyelinase Triggered Fluorescently Labeled Sphingomyelin Containing Liposomes in Tumor Diagnosis after Radiation-Induced Stress. International Journal of Molecular Sciences, 22(8), 3864. https://doi.org/10.3390/ijms22083864