
Nucleotide-Binding Leucine-Rich Repeat Genes CsRSF1 and CsRSF2 Are Positive Modulators in the Cucumis sativus Defense Response to Sphaerotheca fuliginea

Abstract
:1. Introduction
2. Results
2.1. Gene Cloning and Sequence Analysis of CsRSF1 and CsRSF2
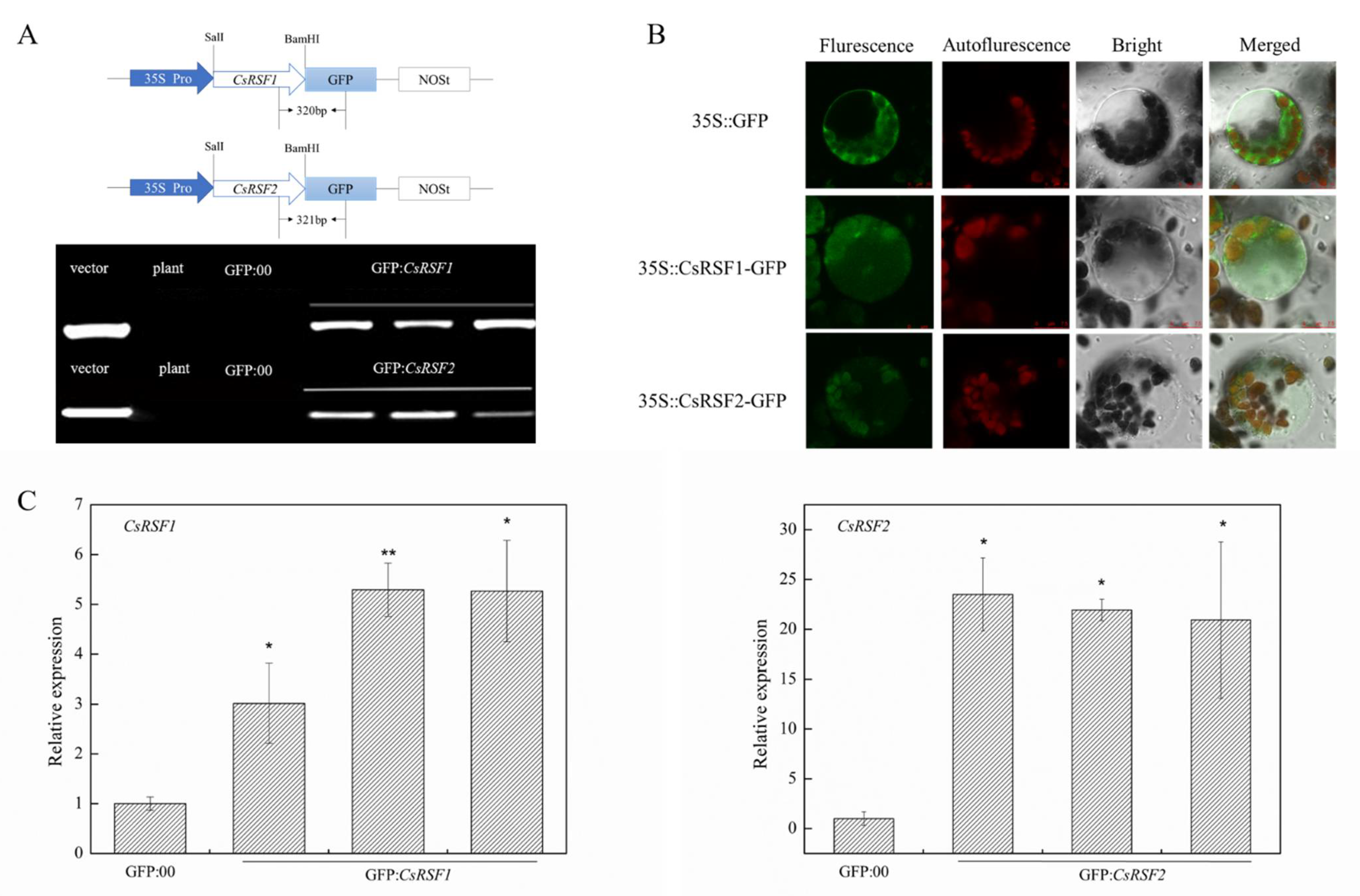
2.2. Subcellular Localization and Expression Patterns of CsRSF1 and CsRSF2
2.3. Response of CsRSF1 and CsRSF2 to Exogenous Substances
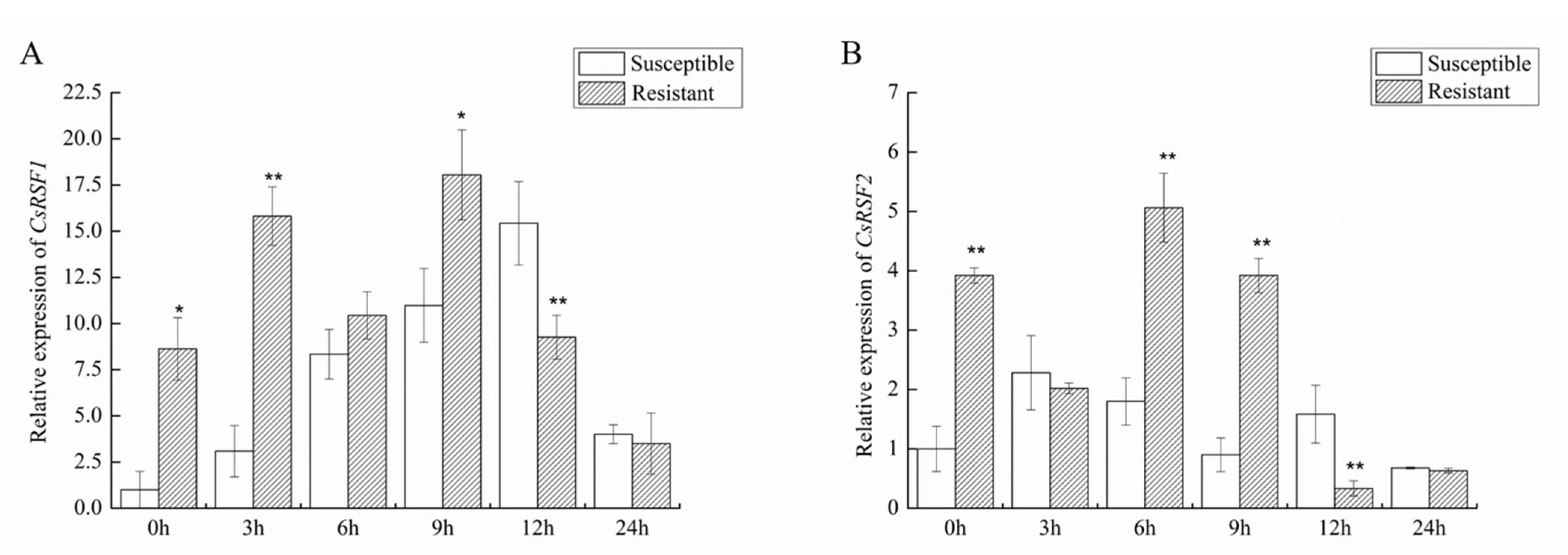
2.4. Expression Patterns of CsRSF1 and CsRSF2 in the Resistant/Susceptible Cucumber Cultivars after S. fuliginea Inoculation
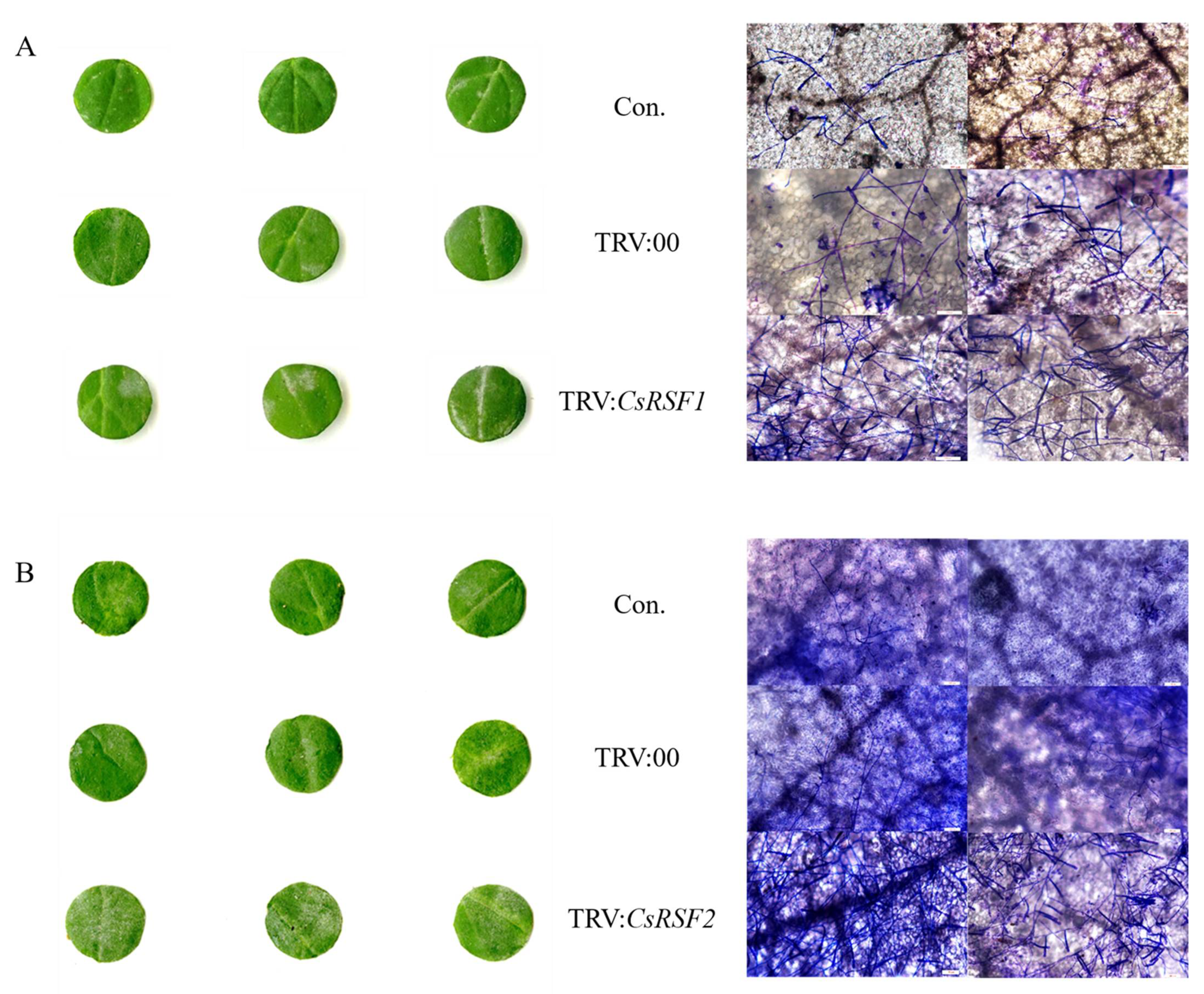
2.5. Silencing of CsRSF1 and CsRSF2 Impairs Host Resistance to S. fuliginea
2.6. CsRSF1 and CsRSF2 Overexpression Improved Resistance to S. fuliginea in Cucumber
2.7. CsRSF1 and CsRSF2 Modulated Expression Levels of Defense-Related Genes
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Treatments
4.2. Pathogen Growth and Inoculation
4.3. Full-Length CDS Cloning and Sequence Analysis
4.4. Subcellular Localization
4.5. Abiotic and Biotic Stresses
4.6. Tobacco Rattle Virus (TRV)-Mediated CsRSF1/CsRSF2 Gene Silencing
4.7. CsRSF1 and CsRSF2 Overexpression Transformation
4.8. Quantitative Reverse-Transcription PCR (qRT–PCR)
4.9. Extraction of Cucumber Protoplasts
4.10. Coomassie Brilliant Blue R250 Staining and Microscopy
4.11. Disease Index
4.12. Data Processing and Statistical Analysis
4.13. Bioinformatics Analysis Content and Tools Website
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shen, H.; You, C.; Zhang, Y.; Cheng, F.; Guo, S.; Sun, J. Research Progress on Comprehensive Control of Cucumber Pow-Dery Mildew. Heilongjiang Agric. Sci. 2017, 8, 133–137. [Google Scholar] [CrossRef]
- Yue, H.; Wu, X.; Hao, J.; Cui, J.; Zhang, S.; Zhang, X.; Bai, Z. Status and Prospects in Molecular Breeding of Powdery Mildew Resistance in Cucumber. J. Plant Genet. Resour. 2014, 15, 120–128. [Google Scholar] [CrossRef]
- Lyon, G.; Newton, A.; Walters, D. Induced Resistance in Crop Protection: The Future, Drivers and Barriers. Induc. Resist. Plant Def. 2007, 243–249. [Google Scholar] [CrossRef]
- Dangl, J.L.; Jones, J.D.G. Plant Pathogens and Integrated Defence Responses to Infection. Nat. Cell Biol. 2001, 411, 826–833. [Google Scholar] [CrossRef] [PubMed]
- Ausubel, F.M. Are Innate Immune Signaling Pathways in Plants and Animals Conserved? Nat. Immunol. 2005, 6, 973–979. [Google Scholar] [CrossRef]
- Hayashi, K.; Fujita, Y.; Ashizawa, T.; Suzuki, F.; Nagamura, Y.; Hayano-Saito, Y. Serotonin Attenuates Biotic Stress and Leads to Lesion Browning Caused by a Hypersensitive Response to Magnaporthe Oryzaepenetration in Rice. Plant J. 2016, 85, 46–56. [Google Scholar] [CrossRef] [Green Version]
- Iwai, T.; Miyasaka, A.; Seo, S.; Ohashi, Y. Contribution of Ethylene Biosynthesis for Resistance to Blast Fungus Infection in Young Rice Plants. Plant Physiol. 2006, 142, 1202–1215. [Google Scholar] [CrossRef] [Green Version]
- Eichmann, R.; Schäfer, P. The Endoplasmic Reticulum in Plant Immunity and Cell Death. Front. Plant Sci. 2012, 3, 200. [Google Scholar] [CrossRef] [Green Version]
- Saha, D.; Rana, R.S.; Sureja, A.K.; Verma, M.; Arya, L.; Das Munshi, A. Cloning and Characterization of NBS-LRR Encoding Resistance Gene Candidates from Tomato Leaf Curl New Delhi Virus Resistant Genotype of Luffa Cylindrica Roem. Physiol. Mol. Plant Pathol. 2013, 81, 107–117. [Google Scholar] [CrossRef]
- Van Ooijen, G.; Burg, H.A.V.D.; Cornelissen, B.J.C.; Takken, F.L.W. Structure and Function of Resistance Proteins in Solanaceous Plants. Annu. Rev. Phytopathol. 2007, 45, 43–72. [Google Scholar] [CrossRef] [Green Version]
- Meyers, B.C.; Kozik, A.; Griego, A.; Kuang, H.; Michelmore, R.W. Genome-Wide Analysis of NBS-LRR–Encoding Genes in Arabidopsis. Plant Cell 2003, 15, 809–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Liu, F.; Zhu, S.; Li, X. The Maize NBS-LRR Gene ZmNBS25 Enhances Disease Resistance in Rice and Arabidopsis. Front. Plant Sci. 2018, 9, 1033. [Google Scholar] [CrossRef] [PubMed]
- Belkin, M.E.; Sigov, A.S. Some Trend in Super-High Frequency Optoelectronics. J. Commun. Technol. Electron. 2009, 54, 855–868. [Google Scholar] [CrossRef]
- Sharma, R.; Rawat, V.; Suresh, C. Genome-Wide Identification and Tissue-Specific Expression Analysis of Nucleotide Binding Site-Leucine Rich Repeat Gene Family in CICER Arietinum (Kabuli Chickpea). Genom. Data 2017, 14, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Pang, Z.; Zhao, S.; Zhang, L.; Lv, Q.; Yin, D.; Li, D.; Liu, X.; Zhao, X.; Li, X.; et al. Importance of OsRac1 and RAI1 in Signalling of Nucleotide-Binding Site Leucine-Rich Repeat Protein-Mediated Resistance to Rice Blast Disease. New Phytol. 2019, 223, 828–838. [Google Scholar] [CrossRef]
- Tameling, W.I.L.; Elzinga, S.D.J.; Darmin, P.S.; Vossen, J.H.; Takken, F.L.W.; Haring, M.A.; Cornelissen, B.J.C. The Tomato R Gene Products I-2 and Mi-1 Are Functional ATP Binding Proteins with ATPase Activity. Plant Cell 2002, 14, 2929–2939. [Google Scholar] [CrossRef]
- Chen, M.; Wu, J.; Wang, L.; Mantri, N.; Zhang, X.; Zhu, Z.; Wang, S. Mapping and Genetic Structure Analysis of the Anthracnose Resistance Locus Co-1HY in the Common Bean (Phaseolus vulgaris L.). PLoS ONE 2017, 12, e0169954. [Google Scholar] [CrossRef]
- Wu, J.; Zhu, J.; Wang, L.; Wang, S. Genome-Wide Association Study Identifies NBS-LRR-Encoding Genes Related with Anthracnose and Common Bacterial Blight in the Common Bean. Front. Plant Sci. 2017, 8, 1398. [Google Scholar] [CrossRef] [Green Version]
- Zou, S.; Wang, H.; Li, Y.; Kong, Z.; Tang, D. The NB-LRR Gene Pm60 Confers Powdery Mildew Resistance in Wheat. New Phytol. 2017, 218, 298–309. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Liu, J.; Bao, S.; Yang, Y.; Zhuang, Y. Molecular Cloning and Characterization of a Wild Eggplant Solanum Aculeatissimum NBS-LRR Gene, Involved in Plant Resistance to Meloidogyne incognita. Int. J. Mol. Sci. 2018, 19, 583. [Google Scholar] [CrossRef] [Green Version]
- Jiang, N.; Cui, J.; Meng, J.; Luan, Y. A Tomato Nucleotide Binding Sites−Leucine-Rich Repeat Gene Is Positively Involved in Plant Resistance to Phytophthora infestans. Phytopathology 2018, 108, 980–987. [Google Scholar] [CrossRef] [Green Version]
- Li, N.-Y.; Zhou, L.; Zhang, D.-D.; Klosterman, S.J.; Li, T.-G.; Gui, Y.-J.; Kong, Z.-Q.; Ma, X.-F.; Short, D.P.G.; Zhang, W.-Q.; et al. Heterologous Expression of the Cotton NBS-LRR Gene GbaNA1 Enhances Verticillium Wilt Resistance in Arabidopsis. Front. Plant Sci. 2018, 9, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calil, I.P.; Fontes, E.P.B. Plant Immunity against Viruses: Antiviral Immune Receptors in Focus. Ann. Bot. 2016, 119, 711–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Q.-H.; Saijo, Y.; Mauch, S.; Biskup, C.; Bieri, S.; Keller, B.; Seki, H.; Ülker, B.; Somssich, I.E.; Schulze-Lefert, P. Nuclear Activity of MLA Immune Receptors Links Isolate-Specific and Basal Disease-Resistance Responses. Science 2007, 315, 1098–1103. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.; Yu, D.; Jiao, J.; Jing, S.; Schulze-Lefert, P.; Shen, Q.-H. Barley MLA Immune Receptors Directly Interfere with Antagonistically Acting Transcription Factors to Initiate Disease Resistance Signaling. Plant Cell 2013, 25, 1158–1173. [Google Scholar] [CrossRef] [Green Version]
- Inoue, H.; Hayashi, N.; Matsushita, A.; Xinqiong, L.; Nakayama, A.; Sugano, S.; Jiang, C.-J.; Takatsuji, H. Blast Resistance of CC-NB-LRR Protein Pb1 is Mediated by WRKY45 through Protein–Protein Interaction. Proc. Natl. Acad. Sci. USA 2013, 110, 9577–9582. [Google Scholar] [CrossRef] [Green Version]
- Wirthmueller, L.; Zhang, Y.; Jones, J.D.; Parker, J.E. Nuclear Accumulation of the Arabidopsis Immune Receptor RPS4 Is Necessary for Triggering EDS1-Dependent Defense. Curr. Biol. 2007, 17, 2023–2029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García, A.V.; Blanvillain-Baufumé, S.; Huibers, R.P.; Wiermer, M.; Li, G.; Gobbato, E.; Rietz, S.; Parker, J.E. Balanced Nuclear and Cytoplasmic Activities of EDS1 Are Required for a Complete Plant Innate Immune Response. PLoS Pathog. 2010, 6, e1000970. [Google Scholar] [CrossRef] [Green Version]
- Heidrich, K.; Wirthmueller, L.; Tasset, C.; Pouzet, C.; Deslandes, L.; Parker, J.E. Arabidopsis EDS1 Connects Pathogen Effector Recognition to Cell Compartment-Specific Immune Responses. Science 2011, 334, 1401–1404. [Google Scholar] [CrossRef] [PubMed]
- Tameling, W.I.; Nooijen, C.; Ludwig, N.; Boter, M.; Slootweg, E.; Goverse, A.; Shirasu, K.; Joosten, M.H. RanGAP2 Mediates Nucleocytoplasmic Partitioning of the NB-LRR Immune Receptor Rx in the Solanaceae, Thereby Dictating Rx Function. Plant Cell 2010, 22, 4176–4194. [Google Scholar] [CrossRef] [Green Version]
- Wan, H.; Yuan, W.; Bo, K.; Shen, J.; Pang, X.; Chen, J. Genome-Wide Analysis of NBS-Encoding Disease Resistance Genes in Cucumis Sativus and Phylogenetic Study of NBS-Encoding Genes in Cucurbitaceae Crops. BMC Genom. 2013, 14, 109. [Google Scholar] [CrossRef] [Green Version]
- Xiao, S.; Ellwood, S.; Calis, O.; Patrick, E.; Li, T.; Coleman, M.; Turner, J.G. Broad-Spectrum Mildew Resistance in Arabidopsis thaliana Mediated by RPW8. Science 2001, 291, 118–120. [Google Scholar] [CrossRef]
- Liu, Z.C. Inheritance Analysis and QTL Mapping of Powdery Mildew Resistance in Cucumber. Ph.D. Thesis, Shanghai Jiao-Tong University, Shanghai, China, 2008. [Google Scholar]
- Luo, J.; Zhang, R.; Qi, X.; Xu, Q.; Chen, X. A Preliminary Study on Cloning of Chitinase Gene and Relationships to Resistance of Powdery Mildew in Cucumber (Cucumis sativus L.). Mol. Plant Breed. 2015, 13, 1584–1591. [Google Scholar]
- Yu, G.C. Differential Proteome Analysis of Cucumber Resistant and Sensitive Sister Leaves in Response to S. fuliginea. Master’s thesis, Shenyang Agricultural University, Shengyang, China, 2008. [Google Scholar]
- Meng, X.; Song, T.; Fan, H.; Yu, Y.; Cui, N.; Zhao, J.; Meng, K. A Comparative Cell Wall Proteomic Analysis of Cucumber Leaves under Sphaerotheca Fuliginea Stress. Acta Physiol. Plant. 2016, 38, 260. [Google Scholar] [CrossRef]
- Panstruga, R.; Kuhn, H. Mutual Interplay between Phytopathogenic Powdery Mildew Fungi and Other Microorganisms. Mol. Plant Pathol. 2018, 20, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Li, D.; Li, Y.; Gu, X.; Huang, S.; Garcia-Mas, J.; Weng, Y. A 1,681-Locus Consensus Genetic Map of Cultivated Cucumber Including 67 NB-LRR Resistance Gene Homolog and Ten Gene Loci. BMC Plant Biol. 2013, 13, 53. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.P.; Liu, M.M.; Miao, H.; Zhang, S.Q.; Yang, Y.H.; Xie, B.Y. QTL Mapping of Resistance Genes to Powdery Mildew in Cucumber (Cucumis sativus L.). Sci. Agric. Sin. 2011, 44, 3584–3593. [Google Scholar]
- Ameline-Torregrosa, C.; Wang, B.-B.; O’Bleness, M.S.; Deshpande, S.; Zhu, H.; Roe, B.; Young, N.D.; Cannon, S.B. Identification and Characterization of Nucleotide-Binding Site-Leucine-Rich Repeat Genes in the Model Plant Medicago truncatula. Plant Physiol. 2008, 146, 5–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Xu, Z.; Jiao, F.; Yu, H.; Xiao, B.; Li, Y.; Lu, X. Cloning, Structural Features, and Expression Analysis of Resistance Gene Analogs in Tobacco. Mol. Biol. Rep. 2009, 37, 345–354. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Zhu, S.; Zhao, R.; Jiang, Z.; Ji, Y.; Ji, J.; Qiu, D.; Li, H.; Bie, T. Pm21, Encoding a Typical CC-NBS-LRR Protein, Confers Broad-Spectrum Resistance to Wheat Powdery Mildew Disease. Mol. Plant 2018, 11, 879–882. [Google Scholar] [CrossRef] [Green Version]
- Dowen, R.H.; Engel, J.L.; Shao, F.; Ecker, J.R.; Dixon, J.E. A Family of Bacterial Cysteine Protease Type III Effectors Utilizes Acylation-Dependent and Independent Strategies to Localize to Plasma Membranes. J. Biol. Chem. 2009, 284, 15867–15879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Elmore, J.M.; Lin, Z.-J.D.; Coaker, G. A Receptor-Like Cytoplasmic Kinase Phosphorylates the Host Target RIN4, Leading to the Activation of a Plant Innate Immune Receptor. Cell Host Microbe 2011, 9, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Jiang, J.; Yang, H.; Zhao, T.; Xu, X.; Li, J. Virus-Induced Gene Silencing (VIGS) of the NBS-LRR Gene SLNLC1 Compromises Sm-Mediated Disease Resistance to Stemphylium Lycopersici in Tomato. Biochem. Biophys. Res. Commun. 2018, 503, 1524–1529. [Google Scholar] [CrossRef]
- Chakraborty, J.; Priya, P.; Dastidar, S.G.; Das, S. Physical Interaction between Nuclear Accumulated CC-NB-ARC-LRR Protein and WRKY64 Promotes EDS1 Dependent Fusarium Wilt Resistance in Chickpea. Plant Sci. 2018, 276, 111–133. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.; Ma, Y.; Zhou, Y.; Zhang, H.; Duan, L.; Chen, H.; Zeng, J.; Zhou, Q.; Wang, S.; Gu, W.; et al. Biosynthesis, Regulation, and Domestication of Bitterness in Cucumber. Science 2014, 346, 1084–1088. [Google Scholar] [CrossRef] [PubMed]
- Bari, R.; Jones, J.D.G. Role of Plant Hormones in Plant Defence Responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef]
- Divi, U.K.; Rahman, T.; Krishna, P. Brassinosteroid-Mediated Stress Tolerance in Arabidopsis Shows Interactions with Abscisic Acid, Ethylene and Salicylic Acid Pathways. BMC Plant Biol. 2010, 10, 151. [Google Scholar] [CrossRef] [Green Version]
- Ton, J.; Flors, V.; Mauch-Mani, B. The Multifaceted Role of ABA in Disease Resistance. Trends Plant Sci. 2009, 14, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.; Tang, S.; Stallmann, A.; Dangl, J.L.; Bonardi, V. Genetic Requirements for Signaling from an Autoactive Plant NB-LRR Intracellular Innate Immune Receptor. PLoS Genet. 2013, 9, e1003465. [Google Scholar] [CrossRef] [Green Version]
- Mohr, P.G.; Cahill, D.M. Relative Roles of Glyceollin, Lignin and the Hypersensitive Response and the Influence of ABA in Compatible and Incompatible Interactions of Soybeans with Phytophthora Sojae. Physiol. Mol. Plant Pathol. 2001, 58, 31–41. [Google Scholar] [CrossRef]
- Adie, B.A.; Pérez-Pérez, J.; Pérez-Pérez, M.M.; Godoy, M.; Sánchez-Serrano, J.-J.; Schmelz, E.A.; Solano, R. ABA Is an Essential Signal for Plant Resistance to Pathogens Affecting JA Biosynthesis and the Activation of Defenses in Arabidopsis. Plant Cell 2007, 19, 1665–1681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vleesschauwer, D.; Van Buyten, E.; Satoh, K.; Balidion, J.; Mauleon, R.; Choi, I.-R.; Vera-Cruz, C.; Kikuchi, S.; Höfte, M. Brassinosteroids Antagonize Gibberellin- and Salicylate-Mediated Root Immunity in Rice. Plant Physiol. 2012, 158, 1833–1846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Torres-Zabala, M.; Truman, W.; Bennett, M.H.; Lafforgue, G.; Mansfield, J.W.; Egea, P.R.; Bögre, L.; Grant, M. Pseudomonas Syringae pv. Tomato Hijacks the Arabidopsis Abscisic Acid Signalling Pathway to Cause Disease. EMBO J. 2007, 26, 1434–1443. [Google Scholar] [CrossRef] [PubMed]
- Browse, J. Jasmonate Passes Muster: A Receptor and Targets for the Defense Hormone. Annu. Rev. Plant Biol. 2009, 60, 183–205. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Liu, J.; Yin, J.; Ni, Y. Induction of PR-1, PR-2, PR-5, Ta-JA2 and Wheat Powdery Mildew Resistance in Response to MeJA Treatment. Acta Phytopathol. Sin. 2001, 41, 270–277. [Google Scholar] [CrossRef]
- Epple, P.; Apel, K.; Bohlmann, H. An Arabidopsis Thaliana Thionin Gene Is Inducible via a Signal Transduction Pathway Different from That for Pathogenesis-Related Proteins. Plant Physiol. 1995, 109, 813–820. [Google Scholar] [CrossRef] [Green Version]
- Penninckx, I.A.; Eggermont, K.; Terras, F.R.; Thomma, B.P.; De Samblanx, G.W.; Buchala, A.; Métraux, J.P.; Manners, J.M.; Broekaert, W.F. Pathogen-Induced Systemic Activation of a Plant Defensin Gene in Arabidopsis Follows a Salicylic Acid-Independent Pathway. Plant Cell 1996, 8, 2309–2323. [Google Scholar] [CrossRef] [Green Version]
- Durner, J.; Shah, J.; Klessig, D.F. Salicylic Acid and Disease Resistance in Plants. Trends Plant Sci. 1997, 2, 266–274. [Google Scholar] [CrossRef]
- Yang, Y.; Shah, J.; Klessig, D.F. Signal Perception and Transduction in Plant Defense Responses. Genes Dev. 1997, 11, 1621–1639. [Google Scholar] [CrossRef] [Green Version]
- Broekaert, W.F.; Delauré, S.L.; De Bolle, M.F.C.; Cammue, B.P.A. The Role of Ethylene in Host-Pathogen Interactions. Annu. Rev. Phytopathol. 2006, 44, 393–416. [Google Scholar] [CrossRef]
- Van Loon, L.C.; Rep, M.; Pieterse, C.M.J. Significance of Inducible Defense-Related Proteins in Infected Plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [Green Version]
- Beckers, G.J.M.; Spoel, S.H. Fine-Tuning Plant Defence Signalling: Salicylate versus Jasmonate. Plant Biol. 2006, 8, 1–10. [Google Scholar] [CrossRef]
- Mur, L.A.; Kenton, P.; Atzorn, R.; Miersch, O.; Wasternack, C. The Outcomes of Concentration-Specific Interactions between Salicylate and Jasmonate Signaling Include Synergy, Antagonism, and Oxidative Stress Leading to Cell Death. Plant Physiol. 2006, 140, 249–262. [Google Scholar] [CrossRef] [Green Version]
- Nahar, K.; Kyndt, T.; Nzogela, Y.B.; Gheysen, G. Abscisic acid Interacts Antagonistically with Classical Defense Pathways in Rice-Migratory Nematode Interaction. New Phytol. 2012, 196, 901–913. [Google Scholar] [CrossRef]
- Vos, I.A.; Emoritz, L.; Pieterse, C.M.J.; Van Wees, S.C.M. Impact of Hormonal Crosstalk on Plant Resistance and Fitness under Multi-Attacker Conditions. Front. Plant Sci. 2015, 6, 639. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Miller, J.; Nozaki, Y.; Takeda, M.; Shah, J.; Hase, S.; Ikegami, M.; Ehara, V.; Dinesh-KumaR, S.P.; Sukamto, R. CY1, an Arabidopsis Thaliana RPP8/HRT Family Resistance Gene, Conferring Resistance to Cucumber Mosaic Virus Requires Salicylic Acid, Ethylene and a Novel Signal Transduction Mechanism. Plant J. 2002, 32, 655–667. [Google Scholar] [CrossRef]
- Navarro, L.; Bari, R.; Achard, P.; Lisón, P.; Nemri, A.; Harberd, N.P.; Jones, J.D. DELLAs Control Plant Immune Responses by Modulating the Balance of Jasmonic Acid and Salicylic Acid Signaling. Curr. Biol. 2008, 18, 650–655. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Lu, C.; Du, L.; Ye, X.; Liu, X.; Coules, A.; Zhang, Z. The Wheat NB-LRR Gene TaRCR1 is Required for Host Defence Response to the Necrotrophic Fungal Pathogen Rhizoctonia Cerealis. Plant Biotechnol. J. 2017, 15, 674–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, V.; Parkhi, V.; Kenerley, C.M.; Rathore, K.S. Defense-Related Gene Expression and Enzyme Activities in Transgenic Cotton Plants Expressing an Endochitinase Gene from Trichoderma Virens in Response to Interaction with Rhizoctonia Solani. Planta 2009, 230, 277–291. [Google Scholar] [CrossRef]
- Wang, Q.; Li, Y.; Ishikawa, K.; Kosami, K.-I.; Uno, K.; Nagawa, S.; Tan, L.; Du, J.; Shimamoto, K.; Kawano, Y. Resistance Protein Pit Interacts with the GEF OsSPK1 to Activate OsRac1 and Trigger Rice Immunity. Proc. Natl. Acad. Sci. USA 2018, 115, E11551–E11560. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Wang, B.; Yin, C.; Zhang, D.; Wang, D.; Song, J.; Zhou, L.; Kong, Z.; Klosterman, S.J.; Li, J.; et al. The Gossypium Hirsutum TIR-NBS-LRR Gene GhDSC1 Mediates Resistance against Verticillium Wilt. Mol. Plant Pathol. 2019, 20, 857–876. [Google Scholar] [CrossRef] [Green Version]
- Ren, H.M. Study on the Inhibitory Mechanism of Chrysophanol on S. fuliginea. Ph.D. Thesis, Hebei Agricultural University, Baoding, China, 2011. [Google Scholar]
- Wang, D.D. Studies on Mechanism, More of Action of Dpintol against Sphaerotheca Fulijinea and Its Mixture with Other Pestcides. Master’s Thesis, Shanghai Jiaotong University, Shanghai, China, 2013. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Material Name | Days Post-Inoculation (dpi) | Disease Index |
|---|---|---|
| Con. | 7 | 22.22 |
| TRV: 00 | 7 | 24.44 |
| TRV:CsRSF1 | 7 | 32.41 |
| Material Name | Days Post-Inoculation (dpi) | Disease Index |
|---|---|---|
| Con. | 7 | 16.68 |
| TRV: 00 | 7 | 17.77 |
| TRV:CsRSF2 | 7 | 36.11 |
| Material Name | Days Post-Inoculation (dpi) | Disease Index |
|---|---|---|
| Con. | 7 | 34.59 |
| GFP: 00 | 7 | 32.00 |
| GFP:CsRSF1 | 7 | 24.88 |
| GFP:CsRSF2 | 7 | 17.33 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Chen, Q.; Huang, J.; Meng, X.; Cui, N.; Yu, Y.; Fan, H. Nucleotide-Binding Leucine-Rich Repeat Genes CsRSF1 and CsRSF2 Are Positive Modulators in the Cucumis sativus Defense Response to Sphaerotheca fuliginea. Int. J. Mol. Sci. 2021, 22, 3986. https://doi.org/10.3390/ijms22083986
Wang X, Chen Q, Huang J, Meng X, Cui N, Yu Y, Fan H. Nucleotide-Binding Leucine-Rich Repeat Genes CsRSF1 and CsRSF2 Are Positive Modulators in the Cucumis sativus Defense Response to Sphaerotheca fuliginea. International Journal of Molecular Sciences. 2021; 22(8):3986. https://doi.org/10.3390/ijms22083986
Chicago/Turabian StyleWang, Xue, Qiumin Chen, Jingnan Huang, Xiangnan Meng, Na Cui, Yang Yu, and Haiyan Fan. 2021. "Nucleotide-Binding Leucine-Rich Repeat Genes CsRSF1 and CsRSF2 Are Positive Modulators in the Cucumis sativus Defense Response to Sphaerotheca fuliginea" International Journal of Molecular Sciences 22, no. 8: 3986. https://doi.org/10.3390/ijms22083986
APA StyleWang, X., Chen, Q., Huang, J., Meng, X., Cui, N., Yu, Y., & Fan, H. (2021). Nucleotide-Binding Leucine-Rich Repeat Genes CsRSF1 and CsRSF2 Are Positive Modulators in the Cucumis sativus Defense Response to Sphaerotheca fuliginea. International Journal of Molecular Sciences, 22(8), 3986. https://doi.org/10.3390/ijms22083986