
A Highlight on the Inhibition of Fungal Carbonic Anhydrases as Drug Targets for the Antifungal Armamentarium



Abstract
:1. Introduction
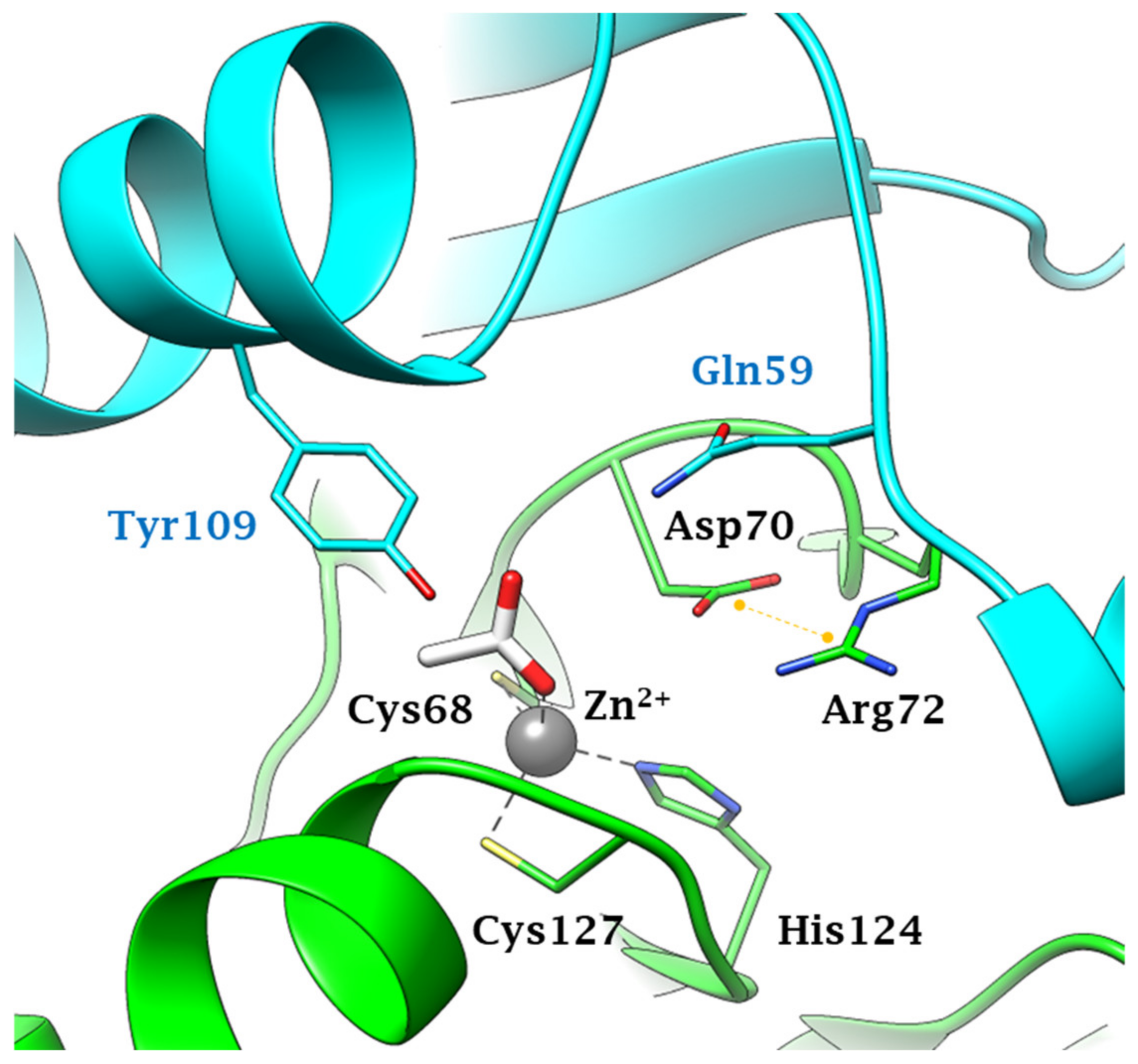
Fungal Enzymes Involved in the Bicarbonate Production (Physiological Role and Structural Features)
2. Main Class of CA Inhibitors (Sulfonamides and Anions)
2.1. Substituted Benzene-Sulfonamides
2.2. Inorganic Metal-Complexing Anions or More Complicated Species
3. Other Classes of Less Investigated CAIs
3.1. Dithiocarbamates
3.2. Phenols
4. Kinetic Parameters and Inhibition Profiles of Various Fungal CAs
4.1. Saccharomyces Cerevisiae CA
4.1.1. Sulfonamide Inhibition
4.1.2. Anion Inhibition
4.1.3. Dithiocarbamate Inhibition
4.1.4. Phenols as Inhibitors
4.2. Candida Glabrata CA
4.2.1. Sulfonamide Inhibition
4.2.2. Anion Inhibition
4.3. Sordaria Macrospora CAs
4.3.1. Sulfonamide Inhibition
4.3.2. Anion Inhibition
4.4. Malassezia Globosa and M. Restricta
4.4.1. Malassezia globosa CA
Sulfonamide Inhibition
Anion Inhibition
Dithiocarbamate Inhibition
Monothiocarbamate Inhibition
Phenols as Inhibitors
Phosphonamidates
4.4.2. Malassezia restricta CA
Sulfonamide Inhibition
Anion Inhibition
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Canadell, J.G.; Le Quere, C.; Raupach, M.R.; Field, C.B.; Buitenhuis, E.T.; Ciais, P.; Conway, T.J.; Gillett, N.P.; Houghton, R.A.; Marland, G. Contributions to accelerating atmospheric CO2 growth from economic activity, carbon intensity, and efficiency of natural sinks. Proc. Natl. Acad. Sci. USA 2007, 104, 18866–18870. [Google Scholar] [CrossRef] [Green Version]
- Cramer, W.; Bondeau, A.; Schaphoff, S.; Lucht, W.; Smith, B.; Sitch, S. Tropical forests and the global carbon cycle: Impacts of atmospheric carbon dioxide, climate change and rate of deforestation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2004, 359, 331–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, L.; Booker, F.L.; Tu, C.; Burkey, K.O.; Zhou, L.; Shew, H.D.; Rufty, T.W.; Hu, S. Arbuscular mycorrhizal fungi increase organic carbon decomposition under elevated CO2. Science 2012, 337, 1084–1087. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, A.P. Fungal CO2 sensing: A breath of fresh air. Curr. Biol. 2005, 15, R934–R936. [Google Scholar] [CrossRef] [Green Version]
- Siscar-Lewin, S.; Hube, B.; Brunke, S. Antivirulence and avirulence genes in human pathogenic fungi. Virulence 2019, 10, 935–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Lara, M.F.; Ostrosky-Zeichner, L. Invasive Candidiasis. Semin. Respir. Crit. Care Med. 2020, 41, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Ledoux, M.P.; Guffroy, B.; Nivoix, Y.; Simand, C.; Herbrecht, R. Invasive Pulmonary Aspergillosis. Semin. Respir. Crit. Care Med. 2020, 41, 80–98. [Google Scholar] [CrossRef] [PubMed]
- Reid, G.; Lynch, J.P., 3rd; Fishbein, M.C.; Clark, N.M. Mucormycosis. Semin. Respir. Crit. Care Med. 2020, 41, 99–114. [Google Scholar] [CrossRef]
- Zavala, S.; Baddley, J.W. Cryptococcosis. Semin. Respir. Crit. Care Med. 2020, 41, 69–79. [Google Scholar] [CrossRef]
- Ohkuni, K.; Hayashi, M.; Yamashita, I. Bicarbonate-mediated social communication stimulates meiosis and sporulation of Saccharomyces cerevisiae. Yeast 1998, 14, 623–631. [Google Scholar] [CrossRef]
- Uno, I.; Matsumoto, K.; Hirata, A.; Ishikawa, T. Outer plaque assembly and spore encapsulation are defective during sporulation of adenylate cyclase-deficient mutants of Saccharomyces cerevisiae. J. Cell Biol. 1985, 100, 1854–1862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, J.C.; Oude-Elferink, R.P. Role of the bicarbonate-responsive soluble adenylyl cyclase in pH sensing and metabolic regulation. Front. Physiol. 2014, 5, 42. [Google Scholar] [CrossRef] [Green Version]
- Klengel, T.; Liang, W.J.; Chaloupka, J.; Ruoff, C.; Schroppel, K.; Naglik, J.R.; Eckert, S.E.; Mogensen, E.G.; Haynes, K.; Tuite, M.F.; et al. Fungal adenylyl cyclase integrates CO2 sensing with cAMP signaling and virulence. Curr. Biol. 2005, 15, 2021–2026. [Google Scholar] [CrossRef] [Green Version]
- Chadwick, B.J.; Lin, X. On the History and Applications of Congenic Strains in Cryptococcus Research. Pathogens 2020, 9, 750. [Google Scholar] [CrossRef] [PubMed]
- Alspaugh, J.A.; Perfect, J.R.; Heitman, J. Cryptococcus neoformans mating and virulence are regulated by the G-protein alpha subunit GPA1 and cAMP. Genes Dev. 1997, 11, 3206–3217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Cann, M.J.; Litvin, T.N.; Iourgenko, V.; Sinclair, M.L.; Levin, L.R.; Buck, J. Soluble adenylyl cyclase as an evolutionarily conserved bicarbonate sensor. Science 2000, 289, 625–628. [Google Scholar] [CrossRef] [Green Version]
- Granger, D.L.; Perfect, J.R.; Durack, D.T. Virulence of Cryptococcus neoformans. Regulation of capsule synthesis by carbon dioxide. J. Clin. Investig. 1985, 76, 508–516. [Google Scholar] [CrossRef] [Green Version]
- Buck, J.; Levin, L.R. Physiological sensing of carbon dioxide/bicarbonate/pH via cyclic nucleotide signaling. Sensors 2011, 11, 2112–2128. [Google Scholar] [CrossRef] [Green Version]
- D’Souza, C.A.; Heitman, J. Conserved cAMP signaling cascades regulate fungal development and virulence. FEMS Microbiol. Rev. 2001, 25, 349–364. [Google Scholar] [CrossRef]
- Bahn, Y.S.; Cox, G.M.; Perfect, J.R.; Heitman, J. Carbonic anhydrase and CO2 sensing during Cryptococcus neoformans growth, differentiation, and virulence. Curr. Biol. 2005, 15, 2013–2020. [Google Scholar] [CrossRef] [Green Version]
- Staniszewska, M. Virulence Factors in Candida species. Curr. Protein Pept. Sci. 2020, 21, 313–323. [Google Scholar] [CrossRef]
- Rutherford, J.C.; Bahn, Y.S.; van den Berg, B.; Heitman, J.; Xue, C. Nutrient and Stress Sensing in Pathogenic Yeasts. Front. Microbiol. 2019, 10, 442. [Google Scholar] [CrossRef] [PubMed]
- Kohler, J.R.; Hube, B.; Puccia, R.; Casadevall, A.; Perfect, J.R. Fungi that Infect Humans. Microbiol. Spectr. 2017, 5, 811–843. [Google Scholar]
- Guzel-Akdemir, O.; Carradori, S.; Grande, R.; Demir-Yazici, K.; Angeli, A.; Supuran, C.T.; Akdemir, A. Development of Thiazolidinones as Fungal Carbonic Anhydrase Inhibitors. Int. J. Mol. Sci. 2020, 21, 2960. [Google Scholar] [CrossRef] [Green Version]
- Annunziato, G.; Angeli, A.; D’Alba, F.; Bruno, A.; Pieroni, M.; Vullo, D.; De Luca, V.; Capasso, C.; Supuran, C.T.; Costantino, G. Discovery of New Potential Anti-Infective Compounds Based on Carbonic Anhydrase Inhibitors by Rational Target-Focused Repurposing Approaches. ChemMedChem 2016, 11, 1904–1914. [Google Scholar] [CrossRef] [PubMed]
- Ozensoy Guler, O.; Capasso, C.; Supuran, C.T. A magnificent enzyme superfamily: Carbonic anhydrases, their purification and characterization. J. Enzym. Inhib. Med. Chem. 2016, 31, 689–694. [Google Scholar] [CrossRef] [PubMed]
- Del Prete, S.; Vullo, D.; De Luca, V.; Carginale, V.; Ferraroni, M.; Osman, S.M.; AlOthman, Z.; Supuran, C.T.; Capasso, C. Sulfonamide inhibition studies of the beta-carbonic anhydrase from the pathogenic bacterium Vibrio cholerae. Bioorg. Med. Chem. 2016, 24, 1115–1120. [Google Scholar] [CrossRef]
- Del Prete, S.; De Luca, V.; De Simone, G.; Supuran, C.T.; Capasso, C. Cloning, expression and purification of the complete domain of the eta-carbonic anhydrase from Plasmodium falciparum. J. Enzym. Inhib. Med. Chem. 2016, 31 (Suppl. S4), 1–6. [Google Scholar] [CrossRef] [Green Version]
- Capasso, C.; Supuran, C.T. An Overview of the Carbonic Anhydrases from Two Pathogens of the Oral Cavity: Streptococcus mutans and Porphyromonas gingivalis. Curr. Top. Med. Chem. 2016, 16, 2359–2368. [Google Scholar] [CrossRef]
- Capasso, C.; Supuran, C.T. An overview of the alpha-, beta- and gamma-carbonic anhydrases from Bacteria: Can bacterial carbonic anhydrases shed new light on evolution of bacteria? J. Enzym. Inhib. Med. Chem. 2015, 30, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Supuran, C.T.; Capasso, C. An Overview of the Bacterial Carbonic Anhydrases. Metabolites 2017, 7, 56. [Google Scholar] [CrossRef] [Green Version]
- Supuran, C.T. Structure and function of carbonic anhydrases. Biochem. J. 2016, 473, 2023–2032. [Google Scholar] [CrossRef] [PubMed]
- McKenna, R.; Supuran, C.T. Carbonic anhydrase inhibitors drug design. Subcell Biochem. 2014, 75, 291–323. [Google Scholar] [PubMed]
- Neri, D.; Supuran, C.T. Interfering with pH regulation in tumours as a therapeutic strategy. Nat. Rev. Drug Discov. 2011, 10, 767–777. [Google Scholar] [CrossRef] [Green Version]
- Supuran, C.T. Carbonic anhydrases: Novel therapeutic applications for inhibitors and activators. Nat. Rev. Drug. Discov. 2008, 7, 168–181. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T. Carbonic anhydrases—An overview. Curr. Pharm. Des. 2008, 14, 603–614. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T.; Capasso, C. Biomedical applications of prokaryotic carbonic anhydrases. Expert Opin. Ther. Pat. 2018, 28, 745–754. [Google Scholar] [CrossRef]
- Supuran, C.T.; Capasso, C. Carbonic Anhydrase from Porphyromonas Gingivalis as a Drug Target. Pathogens 2017, 6, 30. [Google Scholar] [CrossRef] [Green Version]
- Capasso, C.; Supuran, C.T. An Overview of the Selectivity and Efficiency of the Bacterial Carbonic Anhydrase Inhibitors. Curr. Med. Chem. 2015, 22, 2130–2139. [Google Scholar] [CrossRef]
- Rogato, A.; Del Prete, S.; Nocentini, A.; Carginale, V.; Supuran, C.T.; Capasso, C. Phaeodactylum tricornutum as a model organism for testing the membrane penetrability of sulphonamide carbonic anhydrase inhibitors. J. Enzym. Inhib. Med. Chem. 2019, 34, 510–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angeli, A.; Pinteala, M.; Maier, S.S.; Del Prete, S.; Capasso, C.; Simionescu, B.C.; Supuran, C.T. Inhibition of alpha-, beta-, gamma-, delta-, zeta- and eta-class carbonic anhydrases from bacteria, fungi, algae, diatoms and protozoans with famotidine. J Enzym. Inhib. Med. Chem. 2019, 34, 644–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berrino, E.; Bozdag, M.; Del Prete, S.; Alasmary, F.A.S.; Alqahtani, L.S.; AlOthman, Z.; Capasso, C.; Supuran, C.T. Inhibition of alpha-, beta-, gamma-, and delta-carbonic anhydrases from bacteria and diatoms with N’-aryl-N-hydroxy-ureas. J. Enzym. Inhib. Med. Chem. 2018, 33, 1194–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angeli, A.; Del Prete, S.; Alasmary, F.A.S.; Alqahtani, L.S.; AlOthman, Z.; Donald, W.A.; Capasso, C.; Supuran, C.T. The first activation studies of the eta-carbonic anhydrase from the malaria parasite Plasmodium falciparum with amines and amino acids. Bioorg. Chem. 2018, 80, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Mogensen, E.G.; Janbon, G.; Chaloupka, J.; Steegborn, C.; Fu, M.S.; Moyrand, F.; Klengel, T.; Pearson, D.S.; Geeves, M.A.; Buck, J.; et al. Cryptococcus neoformans senses CO2 through the carbonic anhydrase Can2 and the adenylyl cyclase Cac1. Eukaryot Cell 2006, 5, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Schlicker, C.; Hall, R.A.; Vullo, D.; Middelhaufe, S.; Gertz, M.; Supuran, C.T.; Muhlschlegel, F.A.; Steegborn, C. Structure and inhibition of the CO2-sensing carbonic anhydrase Can2 from the pathogenic fungus Cryptococcus neoformans. J. Mol. Biol. 2009, 385, 1207–1220. [Google Scholar] [CrossRef]
- Isik, S.; Kockar, F.; Aydin, M.; Arslan, O.; Guler, O.O.; Innocenti, A.; Scozzafava, A.; Supuran, C.T. Carbonic anhydrase inhibitors: Inhibition of the beta-class enzyme from the yeast Saccharomyces cerevisiae with sulfonamides and sulfamates. Bioorg. Med. Chem. 2009, 17, 1158–1163. [Google Scholar] [CrossRef]
- Isik, S.; Kockar, F.; Arslan, O.; Guler, O.O.; Innocenti, A.; Supuran, C.T. Carbonic anhydrase inhibitors. Inhibition of the beta-class enzyme from the yeast Saccharomyces cerevisiae with anions. Bioorg. Med. Chem. Lett. 2008, 18, 6327–6331. [Google Scholar] [CrossRef]
- Cuesta-Seijo, J.A.; Borchert, M.S.; Navarro-Poulsen, J.C.; Schnorr, K.M.; Mortensen, S.B.; Lo Leggio, L. Structure of a dimeric fungal alpha-type carbonic anhydrase. FEBS Lett. 2011, 585, 1042–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehneck, R.; Neumann, P.; Vullo, D.; Elleuche, S.; Supuran, C.T.; Ficner, R.; Poggeler, S. Crystal structures of two tetrameric beta-carbonic anhydrases from the filamentous ascomycete Sordaria macrospora. FEBS J. 2014, 281, 1759–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Yeon, J.; Sung, J.; Kim, N.J.; Hong, S.; Jin, M.S. Structural insights into novel mechanisms of inhibition of the major beta-carbonic anhydrase CafB from the pathogenic fungus Aspergillus fumigatus. J. Struct. Biol. 2021, 213, 107700. [Google Scholar] [CrossRef]
- Kim, S.; Yeon, J.; Sung, J.; Jin, M.S. Crystal Structure of beta-Carbonic Anhydrase CafA from the Fungal Pathogen Aspergillus fumigatus. Mol. Cells 2020, 43, 831–840. [Google Scholar] [PubMed]
- Covarrubias, A.S.; Bergfors, T.; Jones, T.A.; Hogbom, M. Structural mechanics of the pH-dependent activity of beta-carbonic anhydrase from Mycobacterium tuberculosis. J. Biol. Chem. 2006, 281, 4993–4999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hewitson, K.S.; Vullo, D.; Scozzafava, A.; Mastrolorenzo, A.; Supuran, C.T. Molecular cloning, characterization, and inhibition studies of a beta-carbonic anhydrase from Malassezia globosa, a potential antidandruff target. J. Med. Chem. 2012, 55, 3513–3520. [Google Scholar] [CrossRef] [PubMed]
- Sparber, F.; LeibundGut-Landmann, S. Infecting Mice with Malassezia spp. to Study the Fungus-Host Interaction. J. Vis. Exp. 2019, 153, e60175. [Google Scholar] [CrossRef]
- Kaur, J.; Cao, X.; Abutaleb, N.S.; Elkashif, A.; Graboski, A.L.; Krabill, A.D.; AbdelKhalek, A.H.; An, W.; Bhardwaj, A.; Seleem, M.N.; et al. Optimization of Acetazolamide-Based Scaffold as Potent Inhibitors of Vancomycin-Resistant Enterococcus. J. Med. Chem. 2020, 63, 9540–9562. [Google Scholar] [CrossRef] [PubMed]
- Otten, H. Domagk and the development of the sulphonamides. J. Antimicrob. Chem. 1986, 17, 689–696. [Google Scholar] [CrossRef]
- Capasso, C.; Supuran, C.T. Sulfa and trimethoprim-like drugs—Antimetabolites acting as carbonic anhydrase, dihydropteroate synthase and dihydrofolate reductase inhibitors. J. Enzym. Inhib. Med. Chem. 2014, 29, 379–387. [Google Scholar] [CrossRef]
- Achari, A.; Somers, D.O.; Champness, J.N.; Bryant, P.K.; Rosemond, J.; Stammers, D.K. Crystal structure of the anti-bacterial sulfonamide drug target dihydropteroate synthase. Nat. Struct. Biol. 1997, 4, 490–497. [Google Scholar] [CrossRef]
- Supuran, C.T. Carbonic anhydrase inhibition and the management of neuropathic pain. Expert Rev. Neurother. 2016, 16, 961–968. [Google Scholar] [CrossRef]
- Supuran, C.T. Drug interaction considerations in the therapeutic use of carbonic anhydrase inhibitors. Expert Opin. Drug Metab. Toxicol. 2016, 12, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T. Advances in structure-based drug discovery of carbonic anhydrase inhibitors. Expert Opin. Drug Discov. 2017, 12, 61–88. [Google Scholar] [CrossRef]
- Supuran, C.T. How many carbonic anhydrase inhibition mechanisms exist? J. Enzym. Inhib. Med. Chem. 2016, 31, 345–360. [Google Scholar] [CrossRef] [PubMed]
- Vullo, D.; Del Prete, S.; Fisher, G.M.; Andrews, K.T.; Poulsen, S.A.; Capasso, C.; Supuran, C.T. Sulfonamide inhibition studies of the eta-class carbonic anhydrase from the malaria pathogen Plasmodium falciparum. Bioorg. Med. Chem. 2015, 23, 526–531. [Google Scholar] [CrossRef] [PubMed]
- Vullo, D.; De Luca, V.; Del Prete, S.; Carginale, V.; Scozzafava, A.; Capasso, C.; Supuran, C.T. Sulfonamide inhibition studies of the gamma-carbonic anhydrase from the Antarctic bacterium Pseudoalteromonas haloplanktis. Bioorg. Med. Chem. Lett. 2015, 25, 3550–3555. [Google Scholar] [CrossRef] [Green Version]
- Vullo, D.; De Luca, V.; Del Prete, S.; Carginale, V.; Scozzafava, A.; Capasso, C.; Supuran, C.T. Sulfonamide inhibition studies of the gamma-carbonic anhydrase from the Antarctic cyanobacterium Nostoc commune. Bioorg. Med. Chem. 2015, 23, 1728–1734. [Google Scholar] [CrossRef] [PubMed]
- Dedeoglu, N.; DeLuca, V.; Isik, S.; Yildirim, H.; Kockar, F.; Capasso, C.; Supuran, C.T. Sulfonamide inhibition study of the beta-class carbonic anhydrase from the caries producing pathogen Streptococcus mutans. Bioorg. Med. Chem. Lett. 2015, 25, 2291–2297. [Google Scholar] [CrossRef] [PubMed]
- Alafeefy, A.M.; Ceruso, M.; Al-Tamimi, A.M.; Del Prete, S.; Supuran, C.T.; Capasso, C. Inhibition studies of quinazoline-sulfonamide derivatives against the gamma-CA (PgiCA) from the pathogenic bacterium, Porphyromonas gingivalis. J. Enzym. Inhib. Med. Chem. 2015, 30, 592–596. [Google Scholar] [CrossRef]
- Alafeefy, A.M.; Abdel-Aziz, H.A.; Vullo, D.; Al-Tamimi, A.M.; Awaad, A.S.; Mohamed, M.A.; Capasso, C.; Supuran, C.T. Inhibition of human carbonic anhydrase isozymes I, II, IX and XII with a new series of sulfonamides incorporating aroylhydrazone-, [1,2,4]triazolo [3,4-b][1,3,4]thiadiazinyl- or 2-(cyanophenylmethylene)-1,3,4-thiadiazol-3(2H)-yl moieties. J. Enzym. Inhib. Med. Chem. 2015, 30, 52–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz, J.R.; Fernandez Baldo, M.; Echeverria, G.; Baldoni, H.; Vullo, D.; Soria, D.B.; Supuran, C.T.; Cami, G.E. A substituted sulfonamide and its Co (II), Cu (II), and Zn (II) complexes as potential antifungal agents. J. Enzym. Inhib. Med. Chem. 2016, 31 (Suppl. S2), 51–62. [Google Scholar] [CrossRef] [Green Version]
- Del Prete, S.; Vullo, D.; De Luca, V.; Carginale, V.; Osman, S.M.; AlOthman, Z.; Supuran, C.T.; Capasso, C. Comparison of the sulfonamide inhibition profiles of the alpha-, beta- and gamma-carbonic anhydrases from the pathogenic bacterium Vibrio cholerae. Bioorg. Med. Chem. Lett. 2016, 26, 1941–1946. [Google Scholar] [CrossRef] [PubMed]
- Del Prete, S.; Vullo, D.; De Luca, V.; Carginale, V.; Osman, S.M.; AlOthman, Z.; Supuran, C.T.; Capasso, C. Cloning, expression, purification and sulfonamide inhibition profile of the complete domain of the eta-carbonic anhydrase from Plasmodium falciparum. Bioorg. Med. Chem. Lett. 2016, 26, 4184–4190. [Google Scholar] [CrossRef]
- Abdel Gawad, N.M.; Amin, N.H.; Elsaadi, M.T.; Mohamed, F.M.; Angeli, A.; De Luca, V.; Capasso, C.; Supuran, C.T. Synthesis of 4-(thiazol-2-ylamino)-benzenesulfonamides with carbonic anhydrase I, II and IX inhibitory activity and cytotoxic effects against breast cancer cell lines. Bioorg. Med. Chem. 2016, 24, 3043–3051. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T. Legionella pneumophila Carbonic Anhydrases: Underexplored Antibacterial Drug Targets. Pathogens 2016, 5, 44. [Google Scholar] [CrossRef] [Green Version]
- Nishimori, I.; Vullo, D.; Minakuchi, T.; Scozzafava, A.; Capasso, C.; Supuran, C.T. Sulfonamide inhibition studies of two beta-carbonic anhydrases from the bacterial pathogen Legionella pneumophila. Bioorg. Med. Chem. 2014, 22, 2939–2946. [Google Scholar] [CrossRef]
- Vullo, D.; Sai Kumar, R.S.; Scozzafava, A.; Capasso, C.; Ferry, J.G.; Supuran, C.T. Anion inhibition studies of a beta-carbonic anhydrase from Clostridium perfringens. Bioorg. Med. Chem. Lett. 2013, 23, 6706–6710. [Google Scholar] [CrossRef]
- Nishimori, I.; Minakuchi, T.; Maresca, A.; Carta, F.; Scozzafava, A.; Supuran, C.T. The beta-carbonic anhydrases from Mycobacterium tuberculosis as drug targets. Curr. Pharm. Des. 2010, 16, 3300–3309. [Google Scholar] [CrossRef]
- Carta, F.; Maresca, A.; Covarrubias, A.S.; Mowbray, S.L.; Jones, T.A.; Supuran, C.T. Carbonic anhydrase inhibitors. Characterization and inhibition studies of the most active beta-carbonic anhydrase from Mycobacterium tuberculosis, Rv3588c. Bioorg. Med. Chem. Lett. 2009, 19, 6649–6654. [Google Scholar] [CrossRef]
- Supuran, C.T. Special Issue: Sulfonamides. Molecules 2017, 22, 1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, K.; Ahlawat, R. Famotidine. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Komiya, T.; Huang, C.H. Updates in the Clinical Development of Epacadostat and Other Indoleamine 2,3-Dioxygenase 1 Inhibitors (IDO1) for Human Cancers. Front. Oncol. 2018, 8, 423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carta, F.; Osman, S.M.; Vullo, D.; AlOthman, Z.; Del Prete, S.; Capasso, C.; Supuran, C.T. Poly(amidoamine) dendrimers show carbonic anhydrase inhibitory activity against alpha-, beta-, gamma- and eta-class enzymes. Bioorg. Med. Chem. 2015, 23, 6794–6798. [Google Scholar] [CrossRef] [PubMed]
- Akdemir, A.; Guzel-Akdemir, O.; Karali, N.; Supuran, C.T. Isatin analogs as novel inhibitors of Candida spp. beta-carbonic anhydrase enzymes. Bioorg. Med. Chem. 2016, 24, 1648–1652. [Google Scholar] [CrossRef] [PubMed]
- Teichert, I.; Poggeler, S.; Nowrousian, M. Sordaria macrospora: 25 years as a model organism for studying the molecular mechanisms of fruiting body development. Appl. Microbiol. Biotechnol. 2020, 104, 3691–3704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elleuche, S.; Poggeler, S. Carbonic anhydrases in fungi. Microbiology 2010, 156, 23–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Clavaud, C.; Bar-Hen, A.; Cui, M.; Gao, J.; Liu, Y.; Liu, C.; Shibagaki, N.; Gueniche, A.; Jourdain, R.; et al. Characterization of the major bacterial-fungal populations colonizing dandruff scalps in Shanghai, China, shows microbial disequilibrium. Exp. Dermatol. 2015, 24, 398–400. [Google Scholar] [CrossRef] [PubMed]
- De Simone, G.; Supuran, C.T. (In)organic anions as carbonic anhydrase inhibitors. J. Inorg. Biochem. 2012, 111, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Nocentini, A.; Cadoni, R.; Del Prete, S.; Capasso, C.; Dumy, P.; Gratteri, P.; Supuran, C.T.; Winum, J.Y. Benzoxaboroles as Efficient Inhibitors of the beta-Carbonic Anhydrases from Pathogenic Fungi: Activity and Modeling Study. ACS Med. Chem. Lett. 2017, 8, 1194–1198. [Google Scholar] [CrossRef] [PubMed]
- Nocentini, A.; Vullo, D.; Del Prete, S.; Osman, S.M.; Alasmary, F.A.S.; AlOthman, Z.; Capasso, C.; Carta, F.; Gratteri, P.; Supuran, C.T. Inhibition of the beta-carbonic anhydrase from the dandruff-producing fungus Malassezia globosa with monothiocarbamates. J. Enzym. Inhib. Med. Chem. 2017, 32, 1064–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carta, F.; Aggarwal, M.; Maresca, A.; Scozzafava, A.; McKenna, R.; Masini, E.; Supuran, C.T. Dithiocarbamates strongly inhibit carbonic anhydrases and show antiglaucoma action in vivo. J. Med. Chem. 2012, 55, 1721–1730. [Google Scholar] [CrossRef] [Green Version]
- Carta, F.; Aggarwal, M.; Maresca, A.; Scozzafava, A.; McKenna, R.; Supuran, C.T. Dithiocarbamates: A new class of carbonic anhydrase inhibitors. Crystallographic and kinetic investigations. Chem. Commun. 2012, 48, 1868–1870. [Google Scholar] [CrossRef] [Green Version]
- Monti, S.M.; Maresca, A.; Viparelli, F.; Carta, F.; De Simone, G.; Muhlschlegel, F.A.; Scozzafava, A.; Supuran, C.T. Dithiocarbamates are strong inhibitors of the beta-class fungal carbonic anhydrases from Cryptococcus neoformans, Candida albicans and Candida glabrata. Bioorg. Med. Chem. Lett. 2012, 22, 859–862. [Google Scholar] [CrossRef] [Green Version]
- Maresca, A.; Carta, F.; Vullo, D.; Supuran, C.T. Dithiocarbamates strongly inhibit the beta-class carbonic anhydrases from Mycobacterium tuberculosis. J. Enzym. Inhib. Med. Chem. 2013, 28, 407–411. [Google Scholar] [CrossRef]
- Scozzafava, A.; Mastrolorenzo, A.; Supuran, C.T. Arylsulfonyl-N,N-dialkyl-dithiocarbamates as tumor cell growth inhibitors: Novel agents targeting beta-tubulin? J. Enzym. Inhib. Med. Chem. 2001, 16, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Scozzafava, A.; Mastrolorenzo, A.; Supuran, C.T. Arylsulfonyl-N,N-diethyl-dithiocarbamates: A novel class of antitumor agents. Bioorg. Med. Chem. Lett. 2000, 10, 1887–1891. [Google Scholar] [CrossRef]
- Innocenti, A.; Scozzafava, A.; Supuran, C.T. Carbonic anhydrase inhibitors. Inhibition of transmembrane isoforms IX, XII, and XIV with less investigated anions including trithiocarbonate and dithiocarbamate. Bioorg. Med. Chem. Lett. 2010, 20, 1548–1550. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T. Structure-based drug discovery of carbonic anhydrase inhibitors. J. Enzym. Inhib. Med. Chem. 2012, 27, 759–772. [Google Scholar] [CrossRef] [PubMed]
- Adak, A.K.; Leonov, A.P.; Ding, N.; Thundimadathil, J.; Kularatne, S.; Low, P.S.; Wei, A. Bishydrazide glycoconjugates for lectin recognition and capture of bacterial pathogens. Bioconjug. Chem. 2010, 21, 2065–2075. [Google Scholar] [CrossRef] [Green Version]
- Lomelino, C.L.; Supuran, C.T.; McKenna, R. Non-Classical Inhibition of Carbonic Anhydrase. Int. J. Mol. Sci. 2016, 17, 1150. [Google Scholar] [CrossRef] [Green Version]
- Resnick, M.A.; Cox, B.S. Yeast as an honorary mammal. Mutat. Res. 2000, 451, 1–11. [Google Scholar] [CrossRef]
- Stefanini, I.; Trabocchi, A.; Marchi, E.; Guarna, A.; Cavalieri, D. A systems biology approach to dissection of the effects of small bicyclic peptidomimetics on a panel of Saccharomyces cerevisiae mutants. J. Biol. Chem. 2010, 285, 23477–23485. [Google Scholar] [CrossRef] [Green Version]
- Alafeefy, A.M.; Isik, S.; Al-Jaber, N.A.; Vullo, D.; Abdel-Aziz, H.A.; Ashour, A.E.; Awaad, A.S.; Capasso, C.; Supuran, C.T. Carbonic anhydrase inhibitors. Benzenesulfonamides incorporating cyanoacrylamide moieties strongly inhibit Saccharomyces cerevisiae beta-carbonic anhydrase. Bioorg. Med. Chem. Lett. 2013, 23, 3570–3575. [Google Scholar] [CrossRef]
- Isik, S.; Guler, O.O.; Kockar, F.; Aydin, M.; Arslan, O.; Supuran, C.T. Saccharomyces cerevisiae beta-carbonic anhydrase: Inhibition and activation studies. Curr. Pharm. Des. 2010, 16, 3327–3336. [Google Scholar] [CrossRef]
- Bozdag, M.; Carta, F.; Vullo, D.; Isik, S.; AlOthman, Z.; Osman, S.M.; Scozzafava, A.; Supuran, C.T. Dithiocarbamates with potent inhibitory activity against the Saccharomyces cerevisiae beta-carbonic anhydrase. J. Enzym. Inhib. Med. Chem. 2016, 31, 132–136. [Google Scholar] [CrossRef] [PubMed]
- Bilginer, S.; Unluer, E.; Gul, H.I.; Mete, E.; Isik, S.; Vullo, D.; Ozensoy-Guler, O.; Beyaztas, S.; Capasso, C.; Supuran, C.T. Carbonic anhydrase inhibitors. Phenols incorporating 2- or 3-pyridyl-ethenylcarbonyl and tertiary amine moieties strongly inhibit Saccharomyces cerevisiae beta-carbonic anhydrase. J. Enzym. Inhib. Med. Chem. 2014, 29, 495–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fidel, P.L., Jr.; Vazquez, J.A.; Sobel, J.D. Candida glabrata: Review of epidemiology, pathogenesis, and clinical disease with comparison to C. albicans. Clin. Microbiol. Rev. 1999, 12, 80–96. [Google Scholar] [CrossRef] [Green Version]
- Innocenti, A.; Leewattanapasuk, W.; Muhlschlegel, F.A.; Mastrolorenzo, A.; Supuran, C.T. Carbonic anhydrase inhibitors. Inhibition of the beta-class enzyme from the pathogenic yeast Candida glabrata with anions. Bioorg. Med. Chem. Lett. 2009, 19, 4802–4805. [Google Scholar] [CrossRef] [PubMed]
- Aksu, K.; Nar, M.; Tanc, M.; Vullo, D.; Gulcin, I.; Goksu, S.; Tumer, F.; Supuran, C.T. Synthesis and carbonic anhydrase inhibitory properties of sulfamides structurally related to dopamine. Bioorg. Med. Chem. 2013, 21, 2925–2931. [Google Scholar] [CrossRef] [PubMed]
- Elleuche, S.; Poggeler, S. Evolution of carbonic anhydrases in fungi. Curr. Genet. 2009, 55, 211–222. [Google Scholar] [CrossRef] [Green Version]
- Strop, P.; Smith, K.S.; Iverson, T.M.; Ferry, J.G.; Rees, D.C. Crystal structure of the “cab”-type beta class carbonic anhydrase from the archaeon Methanobacterium thermoautotrophicum. J. Biol. Chem. 2001, 276, 10299–10305. [Google Scholar] [CrossRef] [Green Version]
- Vullo, D.; Lehneck, R.; Donald, W.A.; Poggeler, S.; Supuran, C.T. Anion Inhibition Studies of the beta-Class Carbonic Anhydrase CAS3 from the Filamentous Ascomycete Sordaria macrospora. Metabolites 2020, 10, 93. [Google Scholar] [CrossRef] [Green Version]
- Vullo, D.; Lehneck, R.; Donald, W.A.; Poggeler, S.; Supuran, C.T. Sulfonamide Inhibition Studies of the beta-Class Carbonic Anhydrase CAS3 from the Filamentous Ascomycete Sordaria macrospora. Molecules 2020, 25, 1036. [Google Scholar] [CrossRef] [Green Version]
- Vullo, D.; Lehneck, R.; Poggeler, S.; Supuran, C.T. Sulfonamide inhibition studies of two beta-carbonic anhydrases from the ascomycete fungus Sordaria macrospora, CAS1 and CAS2. J. Enzym. Inhib. Med. Chem. 2018, 33, 390–396. [Google Scholar] [CrossRef] [Green Version]
- Barac, A.; Pekmezovic, M.; Milobratovic, D.; Otasevic-Tasic, S.; Radunovic, M.; Arsic Arsenijevic, V. Presence, species distribution, and density of Malassezia yeast in patients with seborrhoeic dermatitis—A community-based case-control study and review of literature. Mycoses 2015, 58, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Gaitanis, G.; Magiatis, P.; Hantschke, M.; Bassukas, I.D.; Velegraki, A. The Malassezia genus in skin and systemic diseases. Clin. Microbiol. Rev. 2012, 25, 106–141. [Google Scholar] [CrossRef] [Green Version]
- Stalhberger, T.; Simenel, C.; Clavaud, C.; Eijsink, V.G.; Jourdain, R.; Delepierre, M.; Latge, J.P.; Breton, L.; Fontaine, T. Chemical organization of the cell wall polysaccharide core of Malassezia restricta. J. Biol. Chem. 2014, 289, 12647–12656. [Google Scholar] [CrossRef] [Green Version]
- Grice, E.A.; Dawson, T.L., Jr. Host-microbe interactions: Malassezia and human skin. Curr. Opin. Microbiol. 2017, 40, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Loden, M.; Wessman, C. The antidandruff efficacy of a shampoo containing piroctone olamine and salicylic acid in comparison to that of a zinc pyrithione shampoo. Int. J. Cosmet. Sci. 2000, 22, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Hay, R.J. Malassezia, dandruff and seborrhoeic dermatitis: An overview. Br. J. Dermatol. 2011, 165 (Suppl. S2), 2–8. [Google Scholar] [CrossRef]
- Del Prete, S.; Vullo, D.; Osman, S.M.; AlOthman, Z.; Capasso, C.; Supuran, C.T. Anion inhibition studies of the dandruff-producing fungus Malassezia globosa beta-carbonic anhydrase MgCA. Bioorg. Med. Chem. Lett. 2015, 25, 5194–5198. [Google Scholar] [CrossRef]
- Del Prete, S.; Angeli, A.; Ghobril, C.; Hitce, J.; Clavaud, C.; Marat, X.; Supuran, C.T.; Capasso, C. Sulfonamide Inhibition Profile of the beta-Carbonic Anhydrase from Malassezia restricta, An Opportunistic Pathogen Triggering Scalp Conditions. Metabolites 2020, 10, 39. [Google Scholar] [CrossRef] [Green Version]
- Vullo, D.; Del Prete, S.; Nocentini, A.; Osman, S.M.; AlOthman, Z.; Capasso, C.; Bozdag, M.; Carta, F.; Gratteri, P.; Supuran, C.T. Dithiocarbamates effectively inhibit the beta-carbonic anhydrase from the dandruff-producing fungus Malassezia globosa. Bioorg. Med. Chem. 2017, 25, 1260–1265. [Google Scholar] [CrossRef]
- Entezari Heravi, Y.; Bua, S.; Nocentini, A.; Del Prete, S.; Saboury, A.A.; Sereshti, H.; Capasso, C.; Gratteri, P.; Supuran, C.T. Inhibition of Malassezia globosa carbonic anhydrase with phenols. Bioorg. Med. Chem. 2017, 25, 2577–2582. [Google Scholar] [CrossRef]
- Paloukopoulou, C.; Govari, S.; Soulioti, A.; Stefanis, I.; Angeli, A.; Matheeussen, A.; Capasso, C.; Cos, P.; Supuran, C.T.; Karioti, A. Phenols from Origanum dictamnus L. and Thymus vulgaris L. and their activity against Malassezia globosa carbonic anhydrase. Nat. Prod. Res. 2021, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Alissa, S.A.; Alghulikah, H.A.; Alothman, Z.A.; Osman, S.M.; Del Prete, S.; Capasso, C.; Nocentini, A.; Supuran, C.T. Phosphonamidates are the first phosphorus-based zinc binding motif to show inhibition of beta-class carbonic anhydrases from bacteria, fungi, and protozoa. J. Enzym. Inhib. Med. Chem. 2020, 35, 59–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Prete, S.; Vullo, D.; Ghobril, C.; Hitce, J.; Clavaud, C.; Marat, X.; Capasso, C.; Supuran, C.T. Cloning, Purification, and Characterization of a beta-Carbonic Anhydrase from Malassezia restricta, an Opportunistic Pathogen Involved in Dandruff and Seborrheic Dermatitis. Int. J. Mol. Sci. 2019, 20, 2447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prete, S.D.; Angeli, A.; Ghobril, C.; Hitce, J.; Clavaud, C.; Marat, X.; Supuran, C.T.; Capasso, C. Anion Inhibition Profile of the beta-Carbonic Anhydrase from the Opportunist Pathogenic Fungus Malassezia Restricta Involved in Dandruff and Seborrheic Dermatitis. Metabolites 2019, 9, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Inhibitor | KI (nM) * | ||||||
|---|---|---|---|---|---|---|---|
| hCA I 1 (α-CA) | hCA II 1 (α-CA) | MgCA 2 (β-CA) | MreCA 2 (β-CA) | CAS1 1 (β-CA) | CAS2 1 (β-CA) | CAS3 1 (β-CA) | |
| 1 | 28,000 | 300 | 980 | 412 | 361 | 386 | 90 |
| 2 | 25,000 | 240 | 24.5 | 462 | 144 | 3480 | 84 |
| 3 | 79 | 8 | 15.2 | >10,000 | 225 | 3630 | 83 |
| 4 | 78,500 | 320 | 674 | 404 | 47.1 | 6900 | 560 |
| 5 | 25,000 | 170 | 17.4 | >10,000 | 323 | 8720 | 726 |
| 6 | 21,000 | 160 | 7.9 | >10,000 | 241 | 7650 | 441 |
| 7 | 8300 | 60 | 11.6 | 459 | 43.2 | 7360 | 585 |
| 8 | 9800 | 110 | 12.1 | >10,000 | 79.6 | 9120 | 2078 |
| 9 | 6500 | 40 | 34.9 | >10,000 | 580 | 12,000 | 712 |
| 10 | 7300 | 54 | 54.3 | >10,000 | >50,000 | 23,500 | 350 |
| 11 | 5800 | 63 | 9 | 676 | 890 | 18,700 | 235 |
| 12 | 8400 | 75 | 9.2 | >10,000 | 3350 | >50,000 | 90 |
| 13 | 8600 | 60 | 7900 | >10,000 | 8650 | 48.1 | 88 |
| 14 | 9300 | 19 | 8500 | >10,000 | 7215 | 280 | 94 |
| 15 | 5500 | 80 | 23.6 | >10,000 | 3160 | 143 | 605 |
| 16 | 9500 | 94 | 10.4 | 651 | 4520 | 92.5 | 82 |
| 17 | 21,000 | 125 | 6.3 | >10,000 | >50,000 | 390 | 507 |
| 18 | 164 | 46 | 6.8 | >10,000 | 4443 | 3250 | 226 |
| 19 | 109 | 33 | 3500 | 779 | 475 | 6760 | 91 |
| 20 | 6 | 2 | 23.4 | 91 | 363 | 9880 | 85 |
| 21 | 69 | 11 | 11.8 | 740 | 4550 | 4060 | 95 |
| 22 | 164 | 46 | 9.4 | 374 | 1985 | 25,200 | 85 |
| 23 | 109 | 33 | 4530 | >10,000 | 282 | >50,000 | 89 |
| 24 | 95 | 30 | 256 | >10,000 | 294 | >50,000 | 84 |
| AAZ | 250 | 12 | 7600 | 10 | 445 | 816 | 94 |
| MZA | 50 | 14 | 7455 | 390 | 421 | 8140 | 91 |
| EZA | 25 | 8 | 3800 | 379 | 440 | 3170 | 95 |
| DCP | 1200 | 38 | 34.6 | 306 | 1220 | 5790 | 73 |
| DZA | 50,000 | 9 | 7900 | 81 | 360 | 742 | 274 |
| BRZ | 45,000 | 3 | 8400 | 70 | 451 | 739 | 61 |
| BZA | 15 | 9 | 48.2 | 715 | 2115 | 410 | 54 |
| TPM | 250 | 10 | 146 | 383 | 414 | 673 | 363 |
| ZNS | 56 | 35 | 765 | >10,000 | 1820 | 1885 | 710 |
| SLP | 1200 | 40 | 32 | 485 | 1715 | 670 | 493 |
| IND | 31 | 15 | n.d. | 87 | 4240 | 216 | 94 |
| VLX | 54,000 | 43 | 3150 | 77 | 4425 | 3730 | 831 |
| CLX | 50,000 | 21 | 3480 | 140 | 2513 | 857 | 669 |
| SLT | 374 | 9 | n.d. | 67 | 3210 | 496 | 4838 |
| SAC | 18,540 | 5959 | n.d. | 620 | 5280 | 7075 | 191 |
| HCT | 328 | 290 | n.d. | 850 | 3350 | 6680 | 545 |
| FAM | n.d. | n.d. | n.d. | >10,000 | n.d. | n.d. | n.d. |
| EPA | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Anion | KI (mM) * | ||||||
|---|---|---|---|---|---|---|---|
| hCA I 1 (α-CA) | hCA II 1 (α-CA) | MgCA 2 (β-CA) | MreCA 2 (β-CA) | CAS1 1 (β-CA) | CAS2 1 (β-CA) | CAS3 1 (β-CA) | |
| F− | >300 | >300 | 7.13 | >50 | >100 | >100 | >100 |
| Cl− | 6 | 200 | 7.98 | >50 | 9.2 | >100 | >100 |
| Br− | 4 | 63 | 18.6 | >50 | 9.3 | >100 | >100 |
| I− | 0.3 | 26 | 8.73 | 8.6 | 8.6 | 7.7 | 9.9 |
| CNO− | 0.0007 | 0.03 | 6.81 | >50 | 0.9 | 0.82 | 3.2 |
| SCN− | 0.2 | 1.60 | 8.39 | >50 | 5.4 | 5.6 | 7.3 |
| CN− | 0.0005 | 0.02 | 7.19 | >50 | 0.94 | 0.75 | 8.7 |
| N3− | 0.0012 | 1.51 | 45.2 | >50 | >100 | 6.1 | 7.2 |
| NO2− | 8.4 | 63 | 7.56 | >50 | >100 | >100 | 8.3 |
| NO3− | 7 | 35 | 8.13 | 9 | >100 | >100 | 8.5 |
| HCO3− | 12 | 85 | 0.59 | 0.86 | 3.3 | 7.3 | >100 |
| CO32− | 15 | 73 | >100 | >50 | >100 | 8.8 | 8 |
| HSO3− | 18 | 89 | >100 | >50 | 3.3 | 7.3 | >100 |
| SO42− | 63 | >200 | 19.5 | >50 | >100 | 4.8 | >100 |
| HS− | 0.0006 | 0.04 | 11.9 | >50 | 0.89 | 8.5 | 8.3 |
| SnO32− | 0.57 | 0.83 | 5.07 | 0.56 | 4.3 | 0.92 | 7.9 |
| SeO42− | 118 | 112 | 7.41 | 1.7 | 2.4 | 9.2 | 3.4 |
| TeO42− | 0.66 | 0.92 | 5.75 | 0.56 | 2.5 | 6.3 | 8.1 |
| OsO52− | 0.92 | 0.95 | 6.16 | 8.5 | n.d. | n.d. | n.d. |
| P2O74− | 25.77 | 48.50 | 6.03 | >50 | 3.1 | 0.96 | >100 |
| V2O74− | 0.54 | 0.57 | 6.89 | >50 | >100 | 1.4 | >100 |
| B4O72− | 0.64 | 0.95 | 8.45 | 0.4 | 6.7 | 6.9 | 5.9 |
| ReO4− | 0.11 | 0.75 | 16.7 | >50 | 8.2 | >100 | 8.8 |
| RuO4− | 0.101 | 0.69 | 8.82 | 7.4 | 3.9 | >100 | 9.2 |
| S2O82− | 0.107 | 0.084 | >100 | >50 | 5 | >100 | >100 |
| SeCN− | 0.085 | 0.086 | 1.73 | 0.65 | 2.9 | 9.3 | 7.1 |
| CS32− | 0.0087 | 0.0088 | 1.77 | 0.92 | 0.79 | >100 | 8.6 |
| Et2NCS2− | 0.00079 | 0.0031 | 0.30 | 0.075 | 0.38 | 0.93 | 0.89 |
| CF3SO3− | n.d. | n.d. | 2.28 | 4.5 | n.d. | n.d. | n.d. |
| PF6− | n.d. | n.d. | 6.47 | 3.9 | n.d. | n.d. | n.d. |
| ClO4− | >200 | >200 | >100 | 9.2 | >100 | >100 | >100 |
| BF4− | >200 | >200 | >100 | 383 | >100 | >100 | >100 |
| FSO3− | 0.79 | 0.46 | 4.06 | >50 | 0.93 | 8.4 | >100 |
| NH(SO3)22− | 0.31 | 0.76 | 21.4 | >50 | 0.88 | 9.2 | >100 |
| H2NSO2NH | 0.31 | 1.13 | 0.094 | 0.72 | 0.084 | 0.048 | 0.094 |
| H2NSO3H | 0.021 | 0.39 | 0.083 | 7.7 | 0.069 | 0.072 | 0.095 |
| Ph-B(OH)2 | 58.6 | 23.1 | 0.089 | 8.7 | 0.009 | 0.056 | 0.097 |
| Ph-AsO3H2 | 31.7 | 49.2 | 0.090 | 0.83 | 0.035 | 0.054 | 0.091 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Supuran, C.T.; Capasso, C. A Highlight on the Inhibition of Fungal Carbonic Anhydrases as Drug Targets for the Antifungal Armamentarium. Int. J. Mol. Sci. 2021, 22, 4324. https://doi.org/10.3390/ijms22094324
Supuran CT, Capasso C. A Highlight on the Inhibition of Fungal Carbonic Anhydrases as Drug Targets for the Antifungal Armamentarium. International Journal of Molecular Sciences. 2021; 22(9):4324. https://doi.org/10.3390/ijms22094324
Chicago/Turabian StyleSupuran, Claudiu T., and Clemente Capasso. 2021. "A Highlight on the Inhibition of Fungal Carbonic Anhydrases as Drug Targets for the Antifungal Armamentarium" International Journal of Molecular Sciences 22, no. 9: 4324. https://doi.org/10.3390/ijms22094324
APA StyleSupuran, C. T., & Capasso, C. (2021). A Highlight on the Inhibition of Fungal Carbonic Anhydrases as Drug Targets for the Antifungal Armamentarium. International Journal of Molecular Sciences, 22(9), 4324. https://doi.org/10.3390/ijms22094324