
Microglia and Inhibitory Circuitry in the Medullary Dorsal Horn: Laminar and Time-Dependent Changes in a Trigeminal Model of Neuropathic Pain

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Behavior
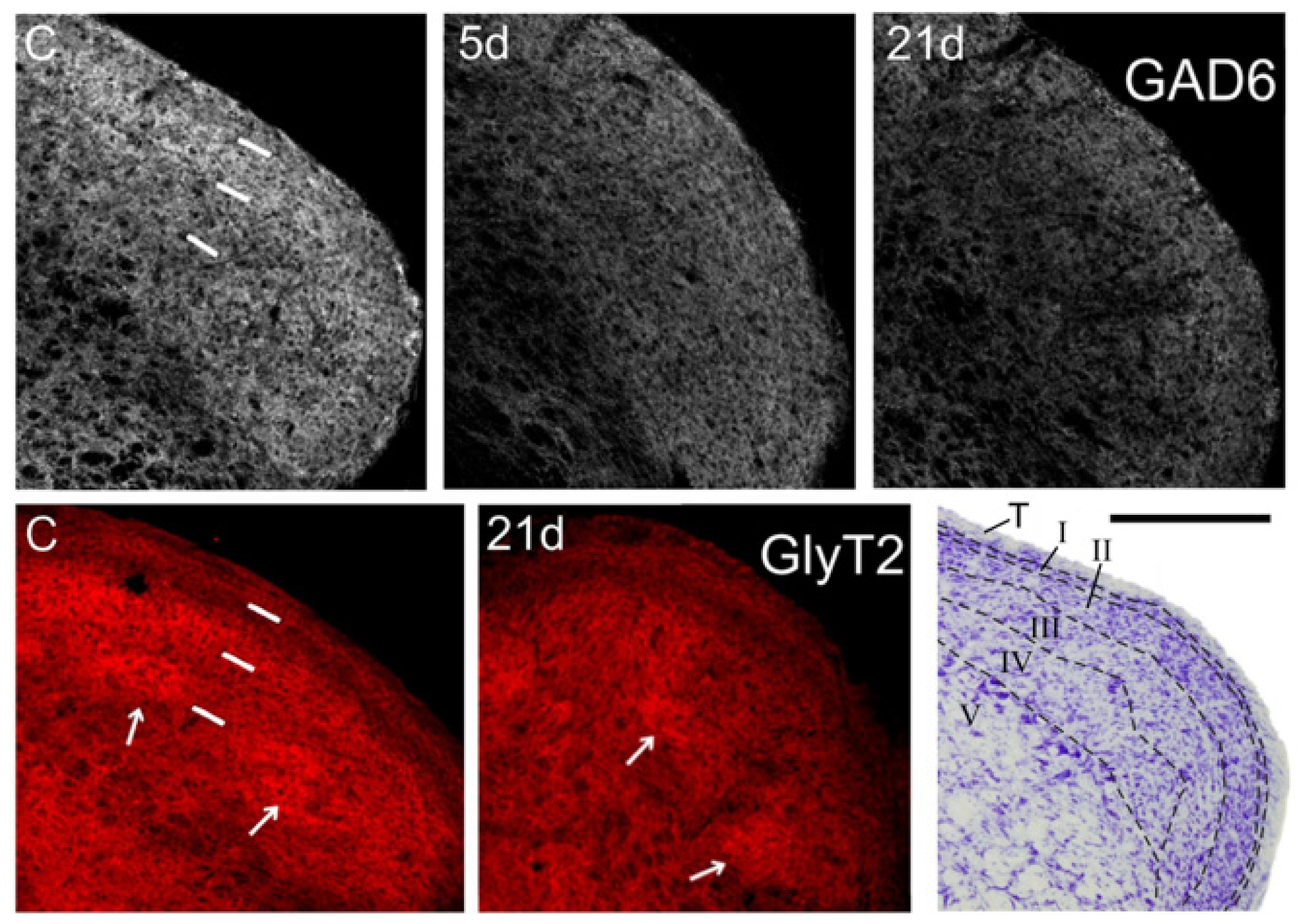
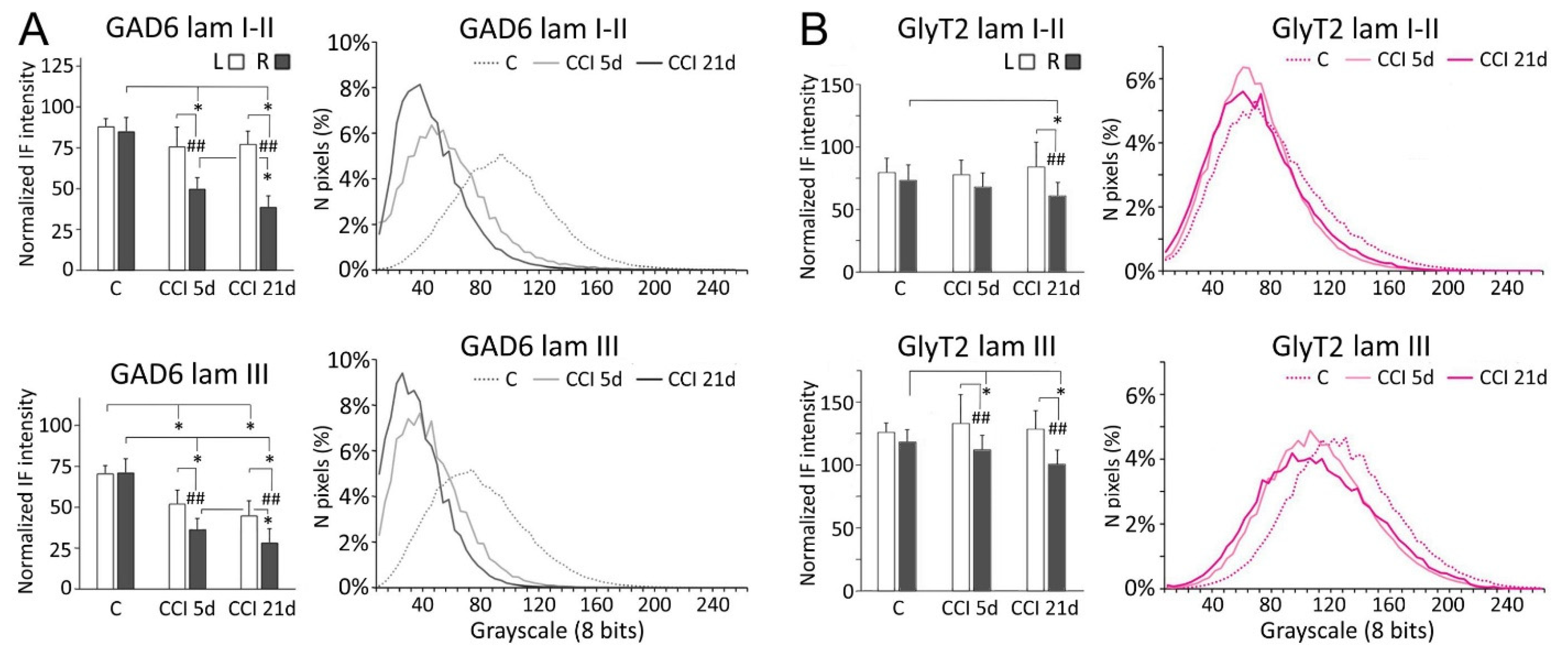
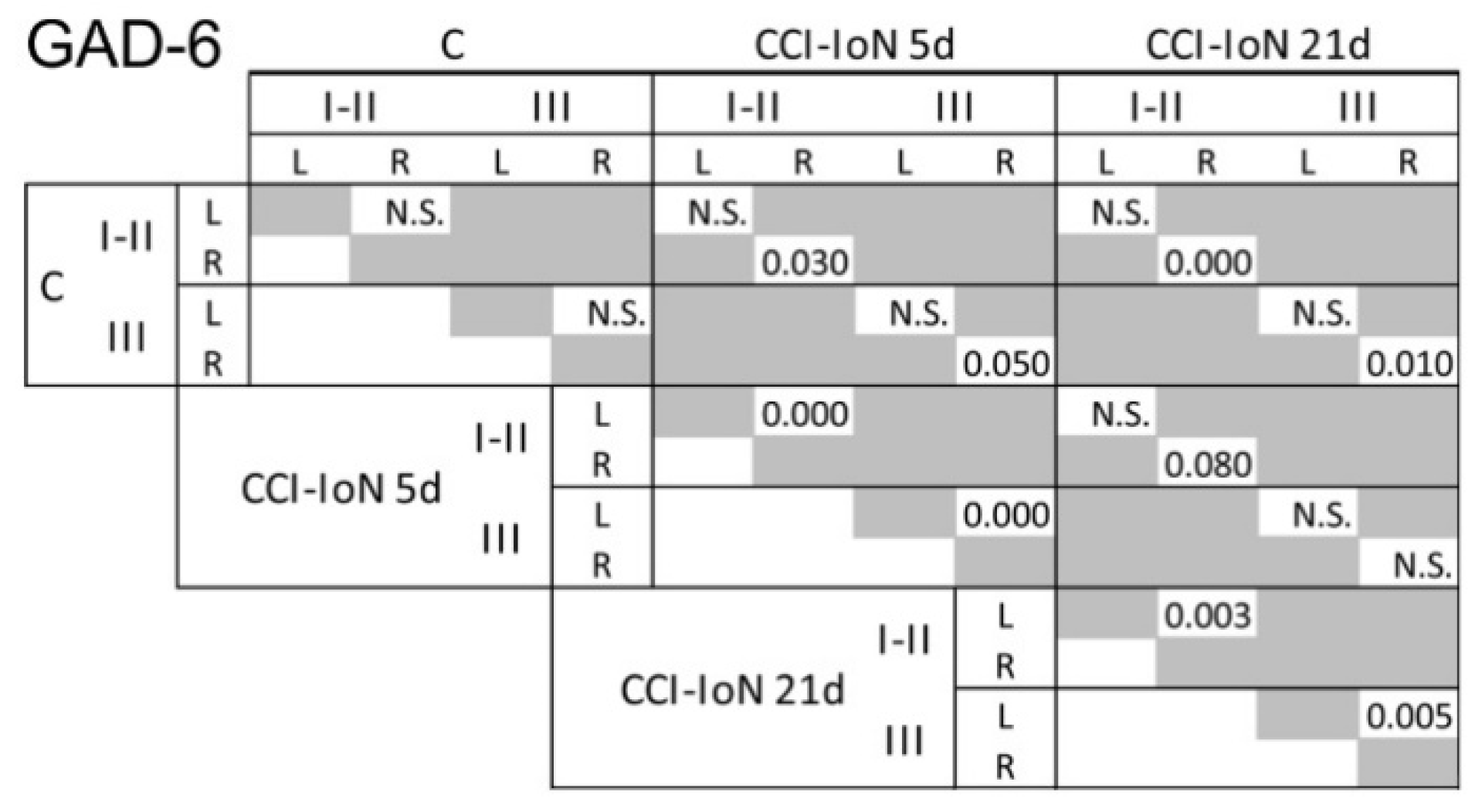
2.2. GAD-6 and GlyT2 Immunostaining
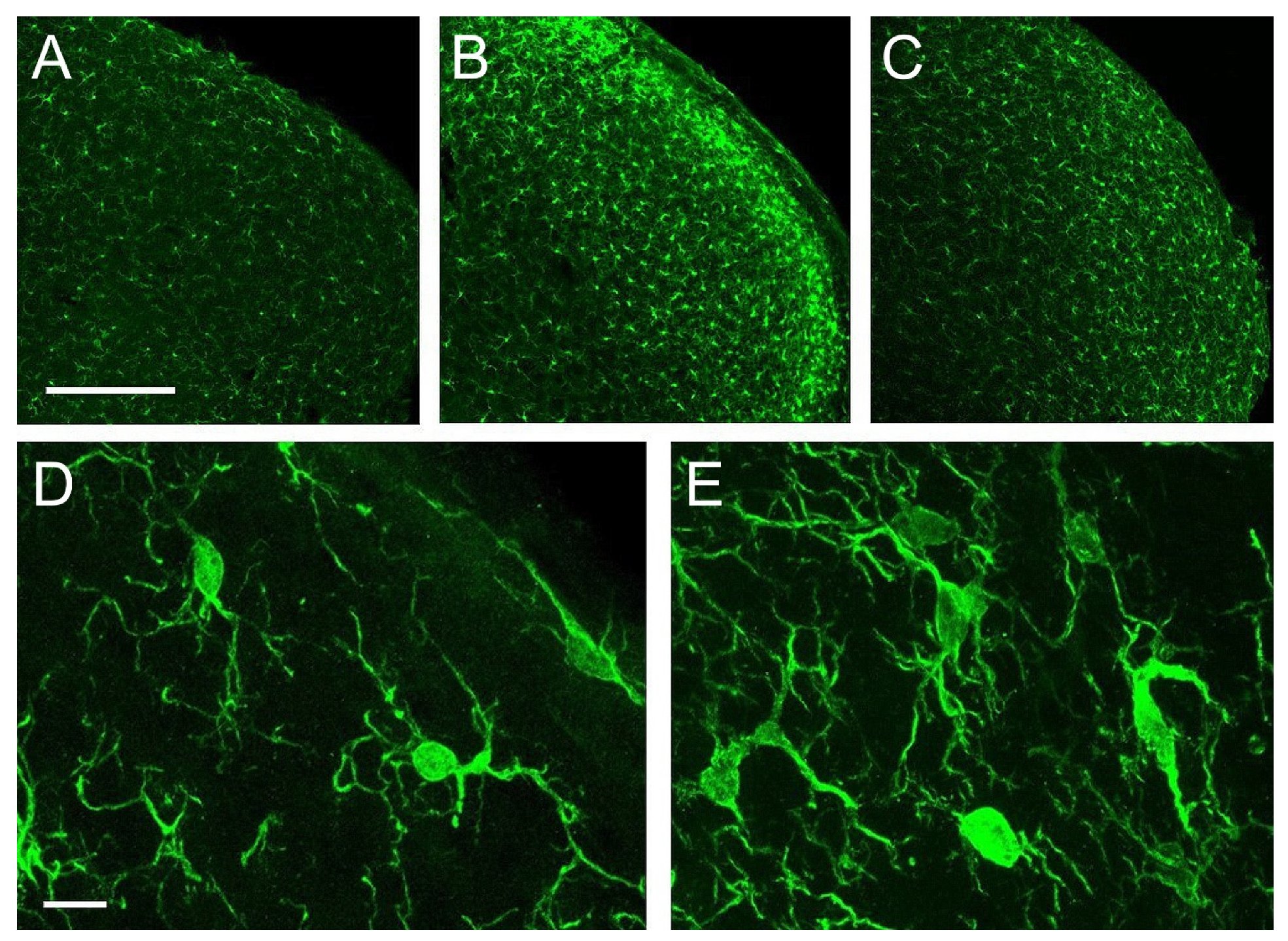
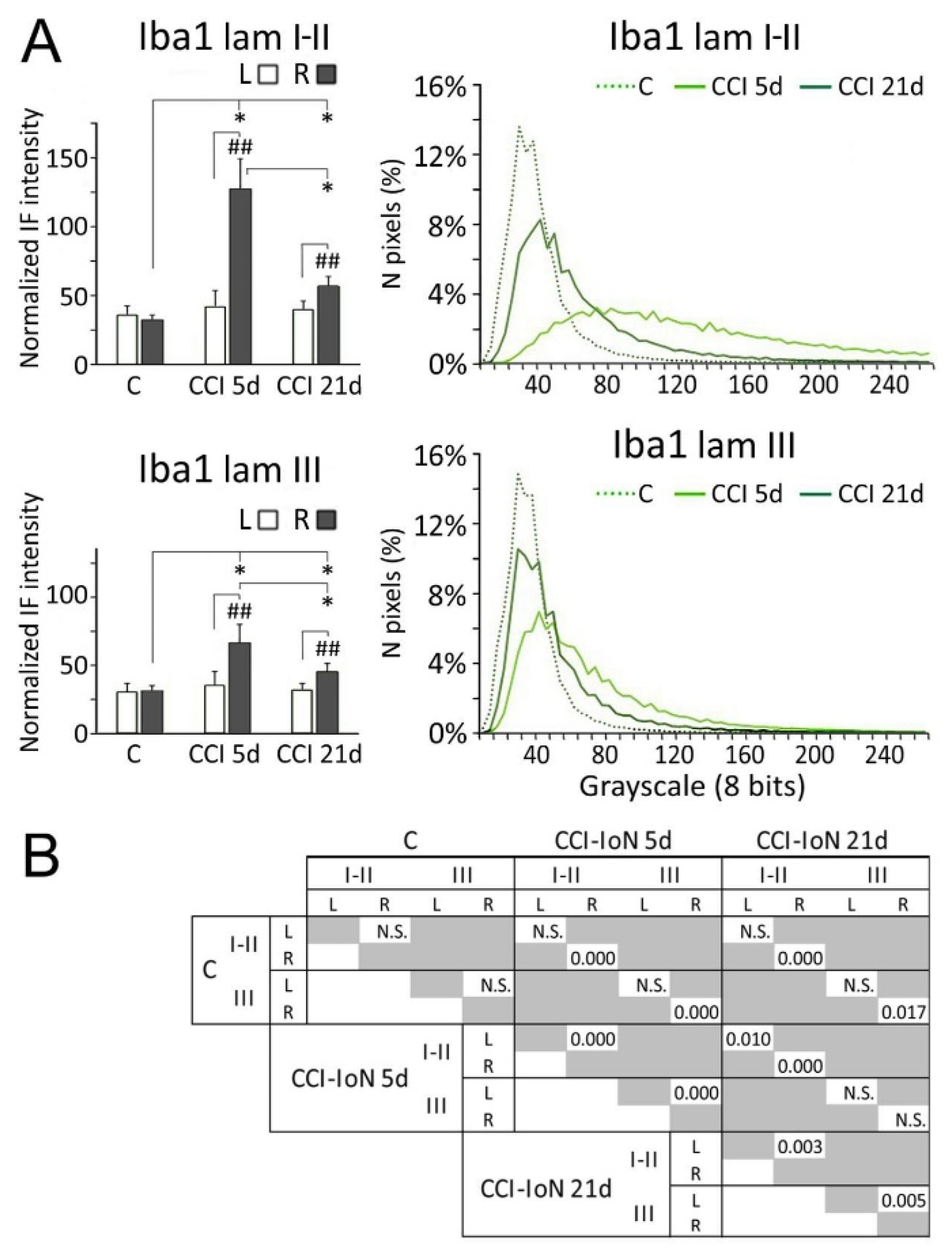
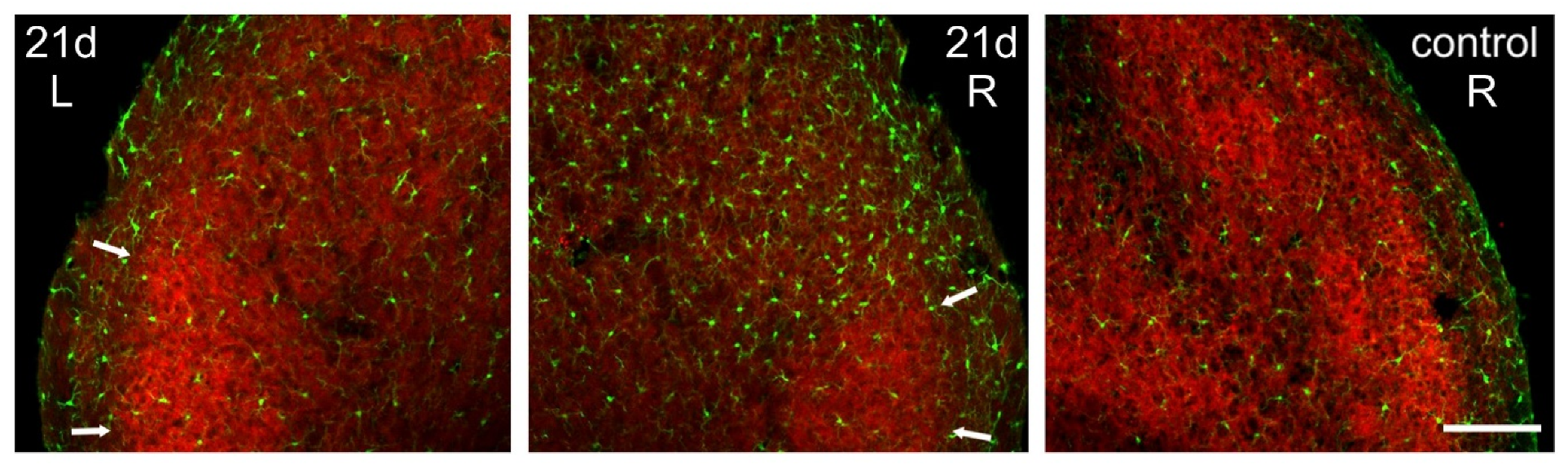
2.3. Microglia
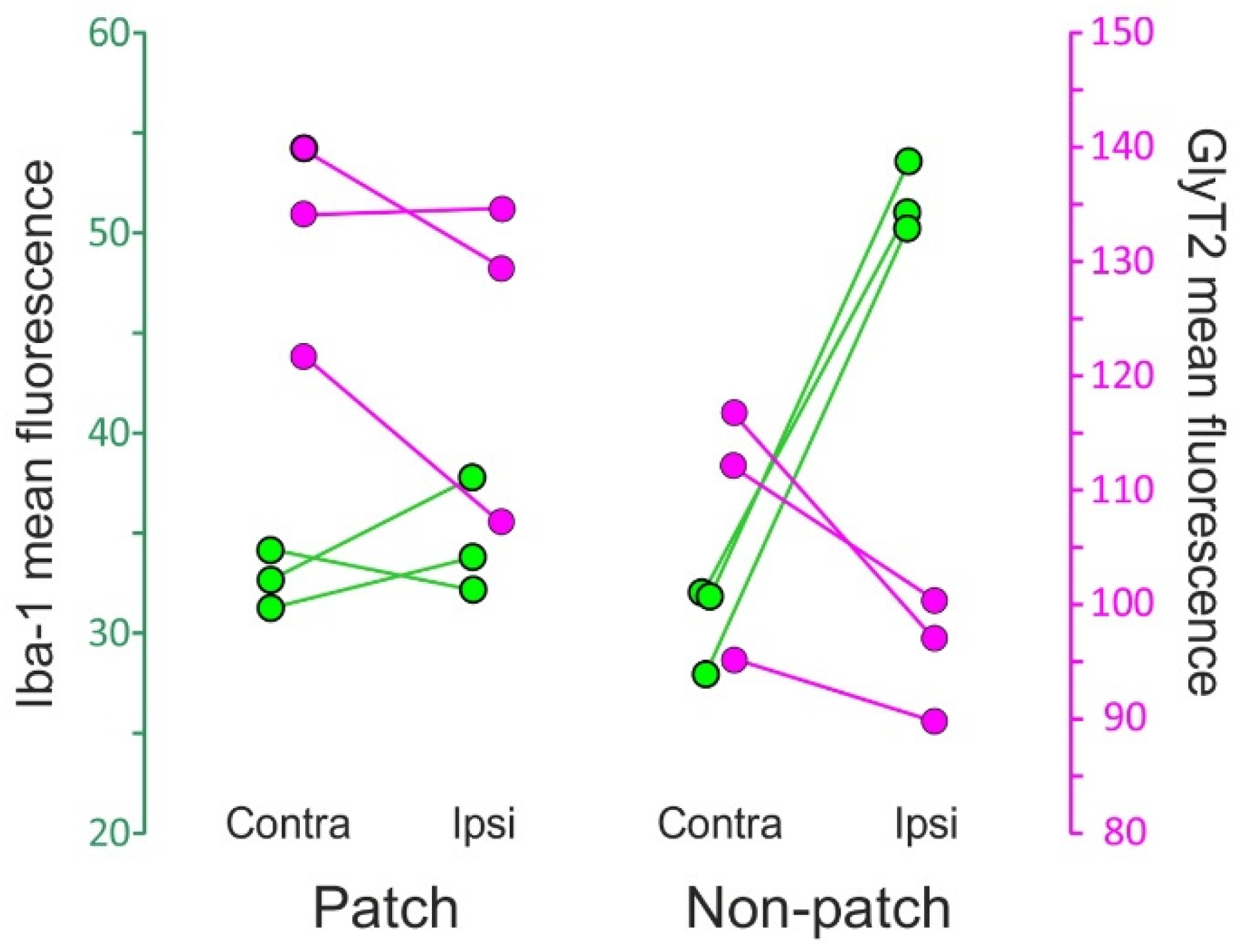
2.4. Microgliosis and GlyT2 Expression
3. Discussion
3.1. Inhibitory Transmitters in Sp5C Are Affected by the CCI-IoN
3.2. Microglia Activation and Its Relationship with the Expression of Inhibitory Molecules
4. Materials and Methods
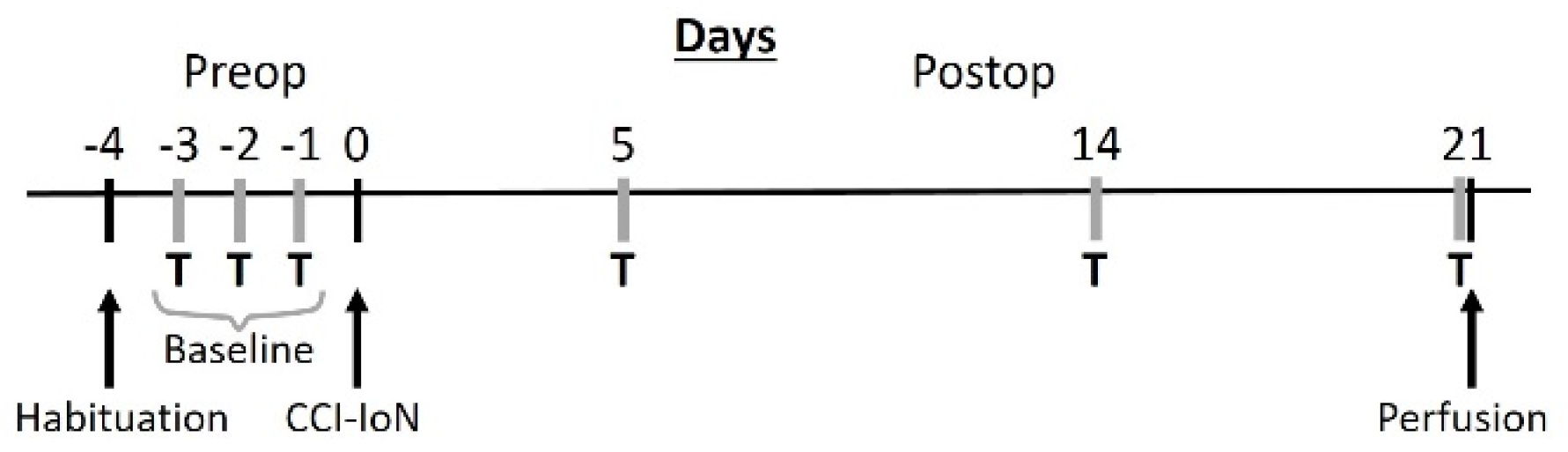
4.1. Experimental Subjects
4.2. Surgery and IoN Constriction
4.3. Behavioral Testing
4.4. Tissue Processing and Immunostaining
4.5. Densitometry of Immunofluorescence
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moore, K.A.; Kohno, T.; Karchewski, L.A.; Scholz, J.; Baba, H.; Woolf, C.J. Partial Peripheral Nerve Injury Promotes a Selective Loss of GABAergic Inhibition in the Superficial Dorsal Horn of the Spinal Cord. J. Neurosci. 2002, 22, 6724–6731. [Google Scholar] [CrossRef]
- Martin, Y.B.; Malmierca, E.; Avendaño, C.; Nuñez, A. Neuronal disinhibition in the trigeminal nucleus caudalis in a model of chronic neuropathic pain. Eur. J. Neurosci. 2010, 32, 399–408. [Google Scholar] [CrossRef]
- Lee, K.Y.; Ratté, S.; Prescott, S.A. Excitatory neurons are more disinhibited than inhibitory neurons by chloride dysregulation in the spinal dorsal horn. eLife 2019, 8, 8. [Google Scholar] [CrossRef]
- Yaksh, T.L. Behavioral and autonomic correlates of the tactile evoked allodynia produced by spinal glycine inhibition: Effects of modulatory receptor systems and excitatory amino acid antagonists. Pain 1989, 37, 111–123. [Google Scholar] [CrossRef]
- Baba, H.; Ji, R.R.; Kohno, T.; Moore, A.K.; Ataka, T.; Wakai, A.; Okamoto, M.; Woolf, C.J. Removal of GABAergic inhibition facilitates polysynaptic A fiber-mediated excitatory transmission to the superficial spinal dorsal horn. Mol. Cell. Neurosci. 2003, 24, 818–830. [Google Scholar] [CrossRef]
- Hwang, J.H.; Yaksh, T.L. The effect of spinal GABA receptor agonists on tactile allodynia in a surgically-induced neuropathic pain model in the rat. Pain 1997, 70, 15–22. [Google Scholar] [CrossRef]
- Kim, J.; Kim, S.J.; Lee, H.; Chang, J.W. Effective neuropathic pain relief through sciatic nerve administration of GAD65-expressing rAAV2. Biochem. Biophys. Res. Commun. 2009, 388, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, K.; Hara, K.; Haranishi, Y.; Terada, T.; Sata, T. The Antinociceptive Effect of SNAP5114, a Gamma-Aminobutyric Acid Transporter-3 Inhibitor, in Rat Experimental Pain Models. Anesth. Analg. 2013, 116, 1162–1169. [Google Scholar] [CrossRef]
- Barthel, F.; Urban, A.; Schlösser, L.; Eulenburg, V.; Werdehausen, R.; Brandenburger, T.; Aragon, C.; Bauer, I.; Hermanns, H. Long-term Application of Glycine Transporter Inhibitors Acts Antineuropathic and Modulates Spinal N-methyl-d-aspartate Receptor Subunit NR-1 Expression in Rats. Anesthesiology 2014, 121, 160–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandenberg, R.J.; Ryan, R.M.; Carland, J.E.; Imlach, W.L.; Christie, M.J. Glycine transport inhibitors for the treatment of pain. Trends Pharmacol. Sci. 2014, 35, 423–430. [Google Scholar] [CrossRef]
- Tsuda, M.; Inoue, K.; Salter, M.W. Neuropathic pain and spinal microglia: A big problem from molecules in ‘small’ glia. Trends Neurosci. 2005, 28, 101–107. [Google Scholar] [CrossRef]
- Tsuda, M.; Shigemoto-Mogami, Y.; Koizumi, S.; Mizokoshi, A.; Kohsaka, S.; Salter, M.W.; Inoue, K. P2X4 receptors induced in spinal microglia gate tactile allodynia after nerve injury. Nat. Cell Biol. 2003, 424, 778–783. [Google Scholar] [CrossRef] [PubMed]
- Coull, J.A.M.; Beggs, S.; Boudreau, D.; Boivin, D.; Tsuda, M.; Inoue, K.; Gravel, C.; Salter, M.W.; De Koninck, Y. BDNF from microglia causes the shift in neuronal anion gradient underlying neuropathic pain. Nat. Cell Biol. 2005, 438, 1017–1021. [Google Scholar] [CrossRef] [PubMed]
- Keller, A.F.; Beggs, S.; Salter, M.W.; De Koninck, Y. Transformation of the Output of Spinal Lamina I Neurons After Nerve Injury and Microglia Stimulation Underlying Neuropathic Pain. Mol. Pain 2007, 3, 27. [Google Scholar] [CrossRef] [Green Version]
- Trang, T.; Beggs, S.; Salter, M.W. Brain-derived neurotrophic factor from microglia: A molecular substrate for neuropathic pain. Neuron Glia Biol. 2011, 7, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Ferrini, F.; De Koninck, Y. Microglia Control Neuronal Network Excitability via BDNF Signalling. Neural Plast. 2013, 2013, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Tsuda, M. Microglia in neuropathic pain: Cellular and molecular mechanisms and therapeutic potential. Nat. Rev. Neurosci. 2018, 19, 138–152. [Google Scholar] [CrossRef] [PubMed]
- Vos, B.P.; Strassman, A.M.; Maciewicz, R.J. Behavioral evidence of trigeminal neuropathic pain following chronic constriction injury to the rat’s infraorbital nerve. J. Neurosci. 1994, 14, 2708–2723. [Google Scholar] [CrossRef] [Green Version]
- Krzyzanowska, A.; Avendaño, C. Behavioral testing in rodent models of orofacial neuropathic and inflammatory pain. Brain Behav. 2012, 2, 678–697. [Google Scholar] [CrossRef]
- Bereiter, D.A.; Hirata, H.; Hu, J.W. Trigeminal subnucleus caudalis: Beyond homologies with the spinal dorsal horn. Pain 2000, 88, 221–224. [Google Scholar] [CrossRef]
- Watson, C.P.N. Management issues of neuropathic trigeminal pain from a medical perspective. J. Orofac. Pain 2004, 18, 366–373. [Google Scholar] [PubMed]
- Aicher, S.A.; Hermes, S.M.; Whittier, K.L.; Hegarty, D.M. Descending projections from the rostral ventromedial medulla (RVM) to trigeminal and spinal dorsal horns are morphologically and neurochemically distinct. J. Chem. Neuroanat. 2012, 43, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Kogelman, L.J.A.; Christensen, R.E.; Pedersen, S.H.; Bertalan, M.; Hansen, T.F.; Jansen-Olesen, I.; Olesen, J. Whole transcriptome expression of trigeminal ganglia compared to dorsal root ganglia in Rattus Norvegicus. Neuroscience 2017, 350, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Megat, S.; Ray, P.R.; Tavares-Ferreira, D.; Moy, J.K.; Sankaranarayanan, I.; Wanghzou, A.; Lou, T.F.; Barragan-Iglesias, P.; Campbell, Z.T.; Dussor, G.; et al. Differences between Dorsal Root and Trigeminal Ganglion Nociceptors in Mice Revealed by Translational Profiling. J. Neurosci. 2019, 39, 6829–6847. [Google Scholar] [CrossRef] [Green Version]
- Latrémolière, A.; Mauborgne, A.; Masson, J.; Bourgoin, S.; Kayser, V.; Hamon, M.; Pohl, M. Differential Implication of Proinflammatory Cytokine Interleukin-6 in the Development of Cephalic versus Extracephalic Neuropathic Pain in Rats. J. Neurosci. 2008, 28, 8489–8501. [Google Scholar] [CrossRef] [Green Version]
- Shibuta, K.; Suzuki, I.; Shinoda, M.; Tsuboi, Y.; Honda, K.; Shimizu, N.; Sessle, B.J.; Iwata, K. Organization of hyperactive microglial cells in trigeminal spinal subnucleus caudalis and upper cervical spinal cord associated with orofacial neuropathic pain. Brain Res. 2012, 1451, 74–86. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, D.L.; Houser, C.R.; Tobin, A.J. Two Forms of the gamma aminobutyric Acid Synthetic Enzyme Glutamate Decarboxylase Have Distinct Intraneuronal Distributions and Cofactor Interactions. J. Neurochem. 1991, 56, 720–723. [Google Scholar] [CrossRef]
- Zafra, F.; Aragon, C.; Olivares, L.; Danbolt, N.; Gimenez, C.; Storm-Mathisen, J. Glycine transporters are differentially expressed among CNS cells. J. Neurosci. 1995, 15, 3952–3969. [Google Scholar] [CrossRef]
- García-Magro, N.; Martin, Y.B.; Palomino-Antolin, A.; Egea, J.; Negredo, P.; Avendaño, C. Multiple Morphometric Assessment of Microglial Cells in Deafferented Spinal Trigeminal Nucleus. Front. Neuroanat. 2020, 13. [Google Scholar] [CrossRef]
- Melzack, R.; Wall, P.D. Pain Mechanisms: A New Theory. Science 1965, 150, 971–978. [Google Scholar] [CrossRef]
- Sivilotti, L.; Woolf, C.J. The contribution of GABAA and glycine receptors to central sensitization: Disinhibition and touch-evoked allodynia in the spinal cord. J. Neurophysiol. 1994, 72, 169–179. [Google Scholar] [CrossRef]
- Sherman, S.E.; Loomis, C.W. Strychnine-sensitive modulation is selective for non-noxious somatosensory input in the spinal cord of the rat. Pain 1996, 66, 321–330. [Google Scholar] [CrossRef]
- Takeda, M.; Tanimoto, T.; Matsumoto, S. Change in mechanical receptive field properties induced by GABA(A) receptor activation in the trigeminal spinal nucleus caudalis neurons in rats. Exp. Brain Res. 2000, 134, 409–416. [Google Scholar] [CrossRef]
- Lu, Y.; Perl, E.R. A Specific Inhibitory Pathway between Substantia Gelatinosa Neurons Receiving Direct C-Fiber Input. J. Neurosci. 2003, 23, 8752–8758. [Google Scholar] [CrossRef] [PubMed]
- Zeilhofer, H.U. The glycinergic control of spinal pain processing. Cell. Mol. Life Sci. 2005, 62, 2027–2035. [Google Scholar] [CrossRef] [PubMed]
- Bardoni, R.; Takazawa, T.; Tong, C.-K.; Choudhury, P.; Scherrer, G.; MacDermott, A.B. Pre- and postsynaptic inhibitory control in the spinal cord dorsal horn. Ann. N. Y. Acad. Sci. 2013, 1279, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Avendaño, C.; Machín, R.; Bermejo, P.E.; Lagares, A. Neuron numbers in the sensory trigeminal nuclei of the rat: A GABA- and glycine-immunocytochemical and stereological analysis. J. Comp. Neurol. 2005, 493, 538–553. [Google Scholar] [CrossRef] [PubMed]
- Castro-Lopes, J.; Tavares, I.; Coimbra, A. GABA decreases in the spinal cord dorsal horn after peripheral neurectomy. Brain Res. 1993, 620, 287–291. [Google Scholar] [CrossRef]
- Huang, W.; Simpson, R. Long-term intrathecal administration of glycine prevents mechanical hyperalgesia in a rat model of neuropathic pain. Neurol. Res. 2000, 22, 160–164. [Google Scholar] [CrossRef] [PubMed]
- Foster, E.; Wildner, H.; Tudeau, L.; Haueter, S.; Ralvenius, W.T.; Jegen, M.; Johannssen, H.; Hösli, L.; Haenraets, K.; Ghanem, A.; et al. Targeted Ablation, Silencing, and Activation Establish Glycinergic Dorsal Horn Neurons as Key Components of a Spinal Gate for Pain and Itch. Neuron 2015, 85, 1289–1304. [Google Scholar] [CrossRef] [Green Version]
- Imlach, W.L.; Bhola, R.F.; Mohammadi, S.A.; Christie, M.J. Glycinergic dysfunction in a subpopulation of dorsal horn interneurons in a rat model of neuropathic pain. Sci. Rep. 2016, 6, 37104. [Google Scholar] [CrossRef] [Green Version]
- Dieb, W.; Hafidi, A. Mechanism of GABA involvement in post-traumatic trigeminal neuropathic pain: Activation of neuronal circuitry composed of PKCγ interneurons and pERK1/2 expressing neurons. Eur. J. Pain 2014, 19, 85–96. [Google Scholar] [CrossRef]
- García-Magro, N.; Negredo, P.; Martin, Y.B.; Nuñez, Á.; Avendaño, C. Modulation of mechanosensory vibrissal responses in the trigeminocervical complex by stimulation of the greater occipital nerve in a rat model of trigeminal neuropathic pain. J. Headache Pain 2020, 21, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Polgár, E.; Hughes, I.D.; Riddell, S.J.; Maxwell, J.D.; Puskár, Z.; Todd, J.A. Selective loss of spinal GABAergic or glycinergic neurons is not necessary for development of thermal hyperalgesia in the chronic constriction injury model of neuropathic pain. Pain 2003, 104, 229–239. [Google Scholar] [CrossRef]
- Price, T.J.; Cervero, F.; Gold, M.S.; Hammond, D.L.; Prescott, S.A. Chloride regulation in the pain pathway. Brain Res. Rev. 2009, 60, 149–170. [Google Scholar] [CrossRef] [Green Version]
- Todd, A.J. Plasticity of Inhibition in the Spinal Cord. Organotypic Models Drug Dev. 2015, 227, 171–190. [Google Scholar] [CrossRef]
- Ahmadi, S.; Muth-Selbach, U.; Lauterbach, A.; Lipfert, P.; Neuhuber, W.L.; Zeilhofer, H.U. Facilitation of Spinal NMDA Receptor Currents by Spillover of Synaptically Released Glycine. Science 2003, 300, 2094–2097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eaton, M.J.; Plunkett, J.A.; Karmally, S.; Martinez, M.; Montanez, K. Changes in GAD- and GABA- immunoreactivity in the spinal dorsal horn after peripheral nerve injury and promotion of recovery by lumbar transplant of immortalized serotonergic precursors. J. Chem. Neuroanat. 1998, 16, 57–72. [Google Scholar] [CrossRef]
- Mackie, M.; Hughes, D.; Maxwell, D.; Tillakaratne, N.; Todd, A. Distribution and colocalisation of glutamate decarboxylase isoforms in the rat spinal cord. Neuroscience 2003, 119, 461–472. [Google Scholar] [CrossRef]
- Lorenzo, L.E.; Magnussen, C.; Bailey, A.L.; Louis, M.S.; De Koninck, Y.; Ribeiro-Da-Silva, A. Spatial and Temporal Pattern of Changes in the Number of GAD65-Immunoreactive Inhibitory Terminals in the Rat Superficial Dorsal Horn following Peripheral Nerve Injury. Mol. Pain 2014, 10, 57. [Google Scholar] [CrossRef] [Green Version]
- Panneton, W.M.; Pan, B.; Gan, Q. Somatotopy in the Medullary Dorsal Horn as a Basis for Orofacial Reflex Behavior. Front. Neurol. 2017, 8, 522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacquin, M.F.; Chiaia, N.L.; Rhoadest, R.W. Trigeminal Projections to Contralateral Dorsal Horn: Central Extent, Peripheral Origins, and Plasticity. Somatosens. Mot. Res. 1990, 7, 153–183. [Google Scholar] [CrossRef]
- Zeilhofer, H.U.; Wildner, H.; Yévenes, G.E. Fast Synaptic Inhibition in Spinal Sensory Processing and Pain Control. Physiol. Rev. 2012, 92, 193–235. [Google Scholar] [CrossRef] [PubMed]
- Petitjean, H.; Pawlowski, S.A.; Fraine, S.L.; Sharif, B.; Hamad, D.; Fatima, T.; Berg, J.; Brown, C.M.; Jan, L.-Y.; Ribeiro-Da-Silva, A.; et al. Dorsal Horn Parvalbumin Neurons Are Gate-Keepers of Touch-Evoked Pain after Nerve Injury. Cell Rep. 2015, 13, 1246–1257. [Google Scholar] [CrossRef] [Green Version]
- Takazawa, T.; Choudhury, P.; Tong, C.-K.; Conway, C.M.; Scherrer, G.; Flood, P.D.; Mukai, J.; MacDermott, A.B. Inhibition Mediated by Glycinergic and GABAergic Receptors on Excitatory Neurons in Mouse Superficial Dorsal Horn Is Location-Specific but Modified by Inflammation. J. Neurosci. 2017, 37, 2336–2348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.C.; Cheng, K.I.; Chen, P.R.; Tseng, K.Y.; Kwan, A.L.; Chang, L.L. Glycine receptors expression in rat spinal cord and dorsal root ganglion in prostaglandin E2 intrathecal injection models. BMC Neurosci. 2018, 19, 72. [Google Scholar] [CrossRef] [PubMed]
- Gilmore, S.A.; Skinner, R.D. Intraspinal non-neuronal cellular responses to peripheral nerve injury. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 1979, 194, 369–387. [Google Scholar] [CrossRef] [PubMed]
- Cova, J.L.; Aldskogius, H.; Arvidsson, J.; Molander, C. Changes in microglial cell numbers in the spinal cord dorsal horn following brachial plexus transection in the adult rat. Exp. Brain Res. 1988, 73, 61–68. [Google Scholar] [CrossRef]
- Gehrmann, J.; Monaco, S.; Kreutzberg, G.W. Spinal cord microglial cells and DRG satellite cells rapidly respond to transection of the rat sciatic nerve. Restor. Neurol. Neurosci. 1991, 2, 181–198. [Google Scholar] [CrossRef]
- Eriksson, N.P.; Persson, J.K.E.; Svensson, M.; Arvidsson, J.; Molander, C.; Aldskogius, H. A quantitative analysis of the microglial cell reaction in central primary sensory projection territories following peripheral nerve injury in the adult rat. Exp. Brain Res. 1993, 96, 19–27. [Google Scholar] [CrossRef]
- Melzer, P.; Zhang, M.-Z.; McKanna, J. Infraorbital nerve transection and whisker follicle removal in adult rats affect microglia and astrocytes in the trigeminal brainstem. A study with lipocortin1- and S100β-immunohistochemistry. Neuroscience 1997, 80, 459–472. [Google Scholar] [CrossRef]
- Inoue, K.; Tsuda, M. Microglia and neuropathic pain. Glia 2009, 57, 1469–1479. [Google Scholar] [CrossRef]
- Calvo, M.; Bennett, D.L. The mechanisms of microgliosis and pain following peripheral nerve injury. Exp. Neurol. 2012, 234, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Zeilhofer, H.U.; Ganley, R. The Oxford Handbook of the Neurobiology of Pain; Wood, J.N., Ed.; Oxford University Press: Oxford, UK, 2019. [Google Scholar]
- Trang, T.; Beggs, S.; Wan, X.; Salter, M.W. P2X4-Receptor-Mediated Synthesis and Release of Brain-Derived Neurotrophic Factor in Microglia Is Dependent on Calcium and p38-Mitogen-Activated Protein Kinase Activation. J. Neurosci. 2009, 29, 3518–3528. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ratnam, J.; Zou, B.; England, P.M.; Basbaum, A.I. TrkB Signaling Is Required for Both the Induction and Maintenance of Tissue and Nerve Injury-Induced Persistent Pain. J. Neurosci. 2009, 29, 5508–5515. [Google Scholar] [CrossRef]
- Geng, S.-J.; Liao, F.-F.; Dang, W.-H.; Ding, X.; Liu, X.-D.; Cai, J.; Han, J.-S.; Wan, Y.; Xing, G.-G. Contribution of the spinal cord BDNF to the development of neuropathic pain by activation of the NR2B-containing NMDA receptors in rats with spinal nerve ligation. Exp. Neurol. 2010, 222, 256–266. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Jiang, E.; Gao, M.; Weng, H.-R. Endogenous activation of presynaptic NMDA receptors enhances glutamate release from the primary afferents in the spinal dorsal horn in a rat model of neuropathic pain. J. Physiol. 2013, 591, 2001–2019. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Walwyn, W.; Ennes, H.S.; Kim, H.; McRoberts, J.A.; Marvizón, J.C.G. BDNF released during neuropathic pain potentiates NMDA receptors in primary afferent terminals. Eur. J. Neurosci. 2014, 39, 1439–1454. [Google Scholar] [CrossRef]
- Hildebrand, M.E.; Xu, J.; Dedek, A.; Li, Y.; Sengar, A.S.; Beggs, S.; Lombroso, P.J.; Salter, M.W. Potentiation of Synaptic GluN2B NMDAR Currents by Fyn Kinase Is Gated through BDNF-Mediated Disinhibition in Spinal Pain Processing. Cell Rep. 2016, 17, 2753–2765. [Google Scholar] [CrossRef] [Green Version]
- Rasmusson, D.D.; Turnbull, B.G. Immediate effects of digit amputation on SI cortex in the raccoon: Unmasking of inhibitory fields. Brain Res. 1983, 288, 368–370. [Google Scholar] [CrossRef]
- Hicks, T.; Dykes, R. Receptive field size for certain neurons in primary somatosensory cortex is determined by GABA-mediated intracortical inhibition. Brain Res. 1983, 274, 160–164. [Google Scholar] [CrossRef]
- Calford, M.B.; Tweedale, R. Acute changes in cutaneous receptive fields in primary somatosensory cortex after digit denervation in adult flying fox. J. Neurophysiol. 1991, 65, 178–187. [Google Scholar] [CrossRef]
- Pettit, M.J.; Schwark, H.D. Receptive field reorganization in dorsal column nuclei during temporary denervation. Science 1993, 262, 2054–2056. [Google Scholar] [CrossRef]
- Panetsos, F.; Nuñez, A.; Avendaño, C. Local anaesthesia induces immediate receptive field changes in nucleus gracilis and cortex. Neuroreport 1995, 7, 150–152. [Google Scholar] [CrossRef]
- Calford, M. Dynamic representational plasticity in sensory cortex. Neuroscience 2002, 111, 709–738. [Google Scholar] [CrossRef]
- Torsney, C. Disinhibition Opens the Gate to Pathological Pain Signaling in Superficial Neurokinin 1 Receptor-Expressing Neurons in Rat Spinal Cord. J. Neurosci. 2006, 26, 1833–1843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winters, B.L.; Rawling, T.; Vandenberg, R.J.; Christie, M.J.; Bhola, R.F.; Imlach, W.L. Activity of novel lipid glycine transporter inhibitors on synaptic signalling in the dorsal horn of the spinal cord. Br. J. Pharmacol. 2018, 175, 2337–2347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohno, K.; Kitano, J.; Kohro, Y.; Tozaki-Saitoh, H.; Inoue, K.; Tsuda, M. Temporal Kinetics of Microgliosis in the Spinal Dorsal Horn after Peripheral Nerve Injury in Rodents. Biol. Pharm. Bull. 2018, 41, 1096–1102. [Google Scholar] [CrossRef] [Green Version]
- Kunori, S.; Matsumura, S.; Okuda-Ashitaka, E.; Katano, T.; Audoly, L.P.; Urade, Y.; Ito, S. A novel role of prostaglandin E2 in neuropathic pain. Glia 2010, 59, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Romero-Sandoval, A.; Chai, N.; Nutile-McMenemy, N.; DeLeo, J.A. A comparison of spinal Iba1 and GFAP expression in rodent models of acute and chronic pain. Brain Res. 2008, 1219, 116–126. [Google Scholar] [CrossRef] [Green Version]
- Linl, S.C.; Yehl, J.H.; Chenl, C.L.; Choul, S.H.; Tsail, Y.J. Effects of local lidocaine treatment before and after median nerve injury on mechanical hypersensitivity and microglia activation in rat cuneate nucleus. Eur. J. Pain 2011, 15, 359–367. [Google Scholar] [CrossRef]
- Xu, F.; Huang, J.; He, Z.; Chen, J.; Tang, X.; Song, Z.; Guo, Q.; Huang, C. Microglial polarization dynamics in dorsal spinal cord in the early stages following chronic sciatic nerve damage. Neurosci. Lett. 2016, 617, 6–13. [Google Scholar] [CrossRef]
- Al-Khrasani, M.; Mohammadzadeh, A.; Balogh, M.; Király, K.; Barsi, S.; Hajnal, B.; Köles, L.; Zádori, Z.S.; Harsing, L.G. Glycine transporter inhibitors: A new avenue for managing neuropathic pain. Brain Res. Bull. 2019, 152, 143–158. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.; Na, Y.-J.; Lee, K.; Kim, Y.H.; Lee, Y.; Kang, M.; Jiang, B.-C.; Yeom, Y.I.; Wu, L.-J.; Gao, Y.-J.; et al. High-resolution transcriptome analysis reveals neuropathic pain gene-expression signatures in spinal microglia after nerve injury. Pain 2016, 157, 964–976. [Google Scholar] [CrossRef] [Green Version]
- Denk, F.; Crow, M.; Didangelos, A.; Lopes, D.M.; McMahon, S.B. Persistent Alterations in Microglial Enhancers in a Model of Chronic Pain. Cell Rep. 2016, 15, 1771–1781. [Google Scholar] [CrossRef] [Green Version]
- Ji, R.-R.; Berta, T.; Nedergaard, M. Glia and pain: Is chronic pain a gliopathy? Pain 2013, 154, S10–S28. [Google Scholar] [CrossRef] [PubMed]
- Coyle, D.E. Partial peripheral nerve injury leads to activation of astroglia and microglia which parallels the development of allodynic behavior. Glia 1998, 23, 75–83. [Google Scholar] [CrossRef]
- Leinders, M.; Knaepen, L.; De Kock, M.; Sommer, C.; Hermans, E.; Deumens, R. Up-regulation of spinal microglial Iba-1 expression persists after resolution of neuropathic pain hypersensitivity. Neurosci. Lett. 2013, 554, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Echeverry, S.; Shi, X.Q.; Yang, M.; Huang, H.; Wu, Y.; Lorenzo, L.E.; Perez-Sanchez, J.; Bonin, R.P.; De Koninck, Y.; Zhang, J. Spinal microglia are required for long-term maintenance of neuropathic pain. Pain 2017, 158, 1792–1801. [Google Scholar] [CrossRef]
- Kosaka, Y.; Yafuso, T.; Shimizu-Okabe, C.; Kim, J.; Kobayashi, S.; Okura, N.; Ando, H.; Okabe, A.; Takayama, C. Development and persistence of neuropathic pain through microglial activation and KCC2 decreasing after mouse tibial nerve injury. Brain Res. 2020, 1733, 146718. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.-X.; Zhuang, Z.-Y.; Woolf, C.J.; Ji, R.-R. p38 Mitogen-Activated Protein Kinase Is Activated after a Spinal Nerve Ligation in Spinal Cord Microglia and Dorsal Root Ganglion Neurons and Contributes to the Generation of Neuropathic Pain. J. Neurosci. 2003, 23, 4017–4022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssen, S.; Truin, M.; Van Kleef, M.; Joosten, E. Differential GABAergic disinhibition during the development of painful peripheral neuropathy. Neuroscience 2011, 184, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Kumada, T.; Furukawa, T.; Inoue, K.; Watanabe, M.; Sato, K.; Fukuda, A. Pre- and post-synaptic switches of GABA actions associated with Cl− homeostatic changes are induced in the spinal nucleus of the trigeminal nerve in a rat model of trigeminal neuropathic pain. Neuroscience 2013, 228, 334–348. [Google Scholar] [CrossRef]
- Masuda, T.; Ozono, Y.; Mikuriya, S.; Kohro, Y.; Tozaki-Saitoh, H.; Iwatsuki, K.; Uneyama, H.; Ichikawa, R.; Salter, M.W.; Tsuda, M.; et al. Dorsal horn neurons release extracellular ATP in a VNUT-dependent manner that underlies neuropathic pain. Nat. Commun. 2016, 7, 12529. [Google Scholar] [CrossRef] [Green Version]
- Krzyzanowska, A.; Pittolo, S.; Cabrerizo, M.; Sánchez-López, J.; Krishnasamy, S.; Venero, C.; Avendaño, C. Assessing nociceptive sensitivity in mouse models of inflammatory and neuropathic trigeminal pain. J. Neurosci. Methods 2011, 201, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Bennett, G.J.; Xie, Y.K. A peripheral mononeuropathy in rat that produces disorders of pain sensation like those seen in man. Pain 1988, 33, 87–107. [Google Scholar] [CrossRef]
- García-Magro, N.; Martin, Y.B.; Negredo, P.; Avendaño, C. The greater occipital nerve and its spinal and brainstem afferent projections: A stereological and tract-tracing study in the rat. J. Comp. Neurol. 2018, 526, 3000–3019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Núñez, E.; Pérez-Siles, G.; Rodenstein, L.; Alonso-Torres, P.; Zafra, F.; Jimenez, E.; Aragón, C.; López-Corcuera, B. Subcellular Localization of the Neuronal Glycine Transporter GLYT2 in Brainstem. Traffic 2009, 10, 829–843. [Google Scholar] [CrossRef]
- Kirkman, T.W. Statistics to Use. Available online: http://www.physics.csbsju.edu/stats/ (accessed on 28 December 2020).









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Magro, N.; Martin, Y.B.; Negredo, P.; Zafra, F.; Avendaño, C. Microglia and Inhibitory Circuitry in the Medullary Dorsal Horn: Laminar and Time-Dependent Changes in a Trigeminal Model of Neuropathic Pain. Int. J. Mol. Sci. 2021, 22, 4564. https://doi.org/10.3390/ijms22094564
García-Magro N, Martin YB, Negredo P, Zafra F, Avendaño C. Microglia and Inhibitory Circuitry in the Medullary Dorsal Horn: Laminar and Time-Dependent Changes in a Trigeminal Model of Neuropathic Pain. International Journal of Molecular Sciences. 2021; 22(9):4564. https://doi.org/10.3390/ijms22094564
Chicago/Turabian StyleGarcía-Magro, Nuria, Yasmina B. Martin, Pilar Negredo, Francisco Zafra, and Carlos Avendaño. 2021. "Microglia and Inhibitory Circuitry in the Medullary Dorsal Horn: Laminar and Time-Dependent Changes in a Trigeminal Model of Neuropathic Pain" International Journal of Molecular Sciences 22, no. 9: 4564. https://doi.org/10.3390/ijms22094564
APA StyleGarcía-Magro, N., Martin, Y. B., Negredo, P., Zafra, F., & Avendaño, C. (2021). Microglia and Inhibitory Circuitry in the Medullary Dorsal Horn: Laminar and Time-Dependent Changes in a Trigeminal Model of Neuropathic Pain. International Journal of Molecular Sciences, 22(9), 4564. https://doi.org/10.3390/ijms22094564