
Genome-Wide Investigation of the NF-X1 Gene Family in Populus trichocarpa Expression Profiles during Development and Stress

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Identification and Analysis of NF-X1 in A. thaliana, Oryza sativa, and P. trichocarpa
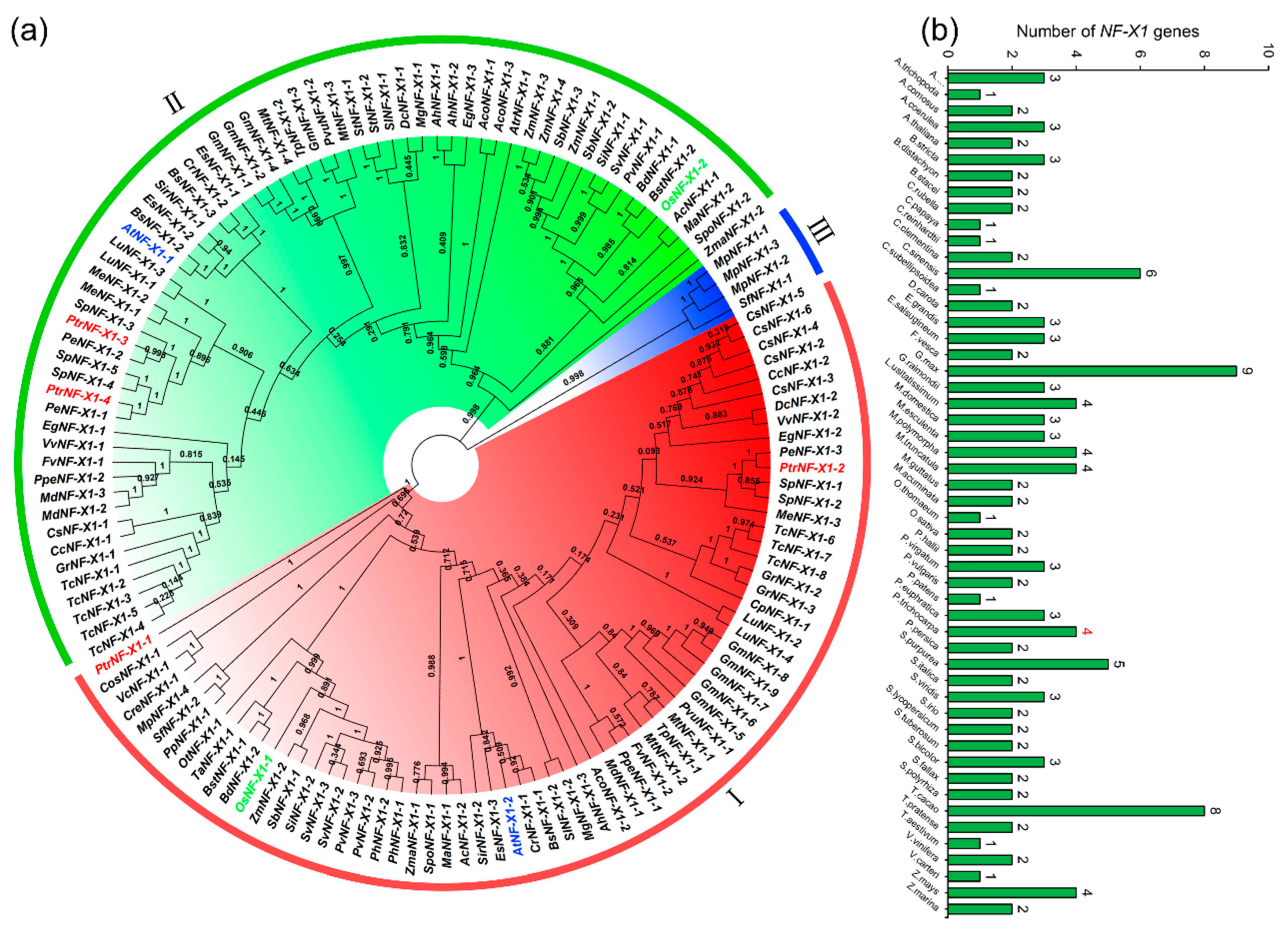
2.2. Evolutionary and Phylogenetic Analysis of the NF-X1 Family
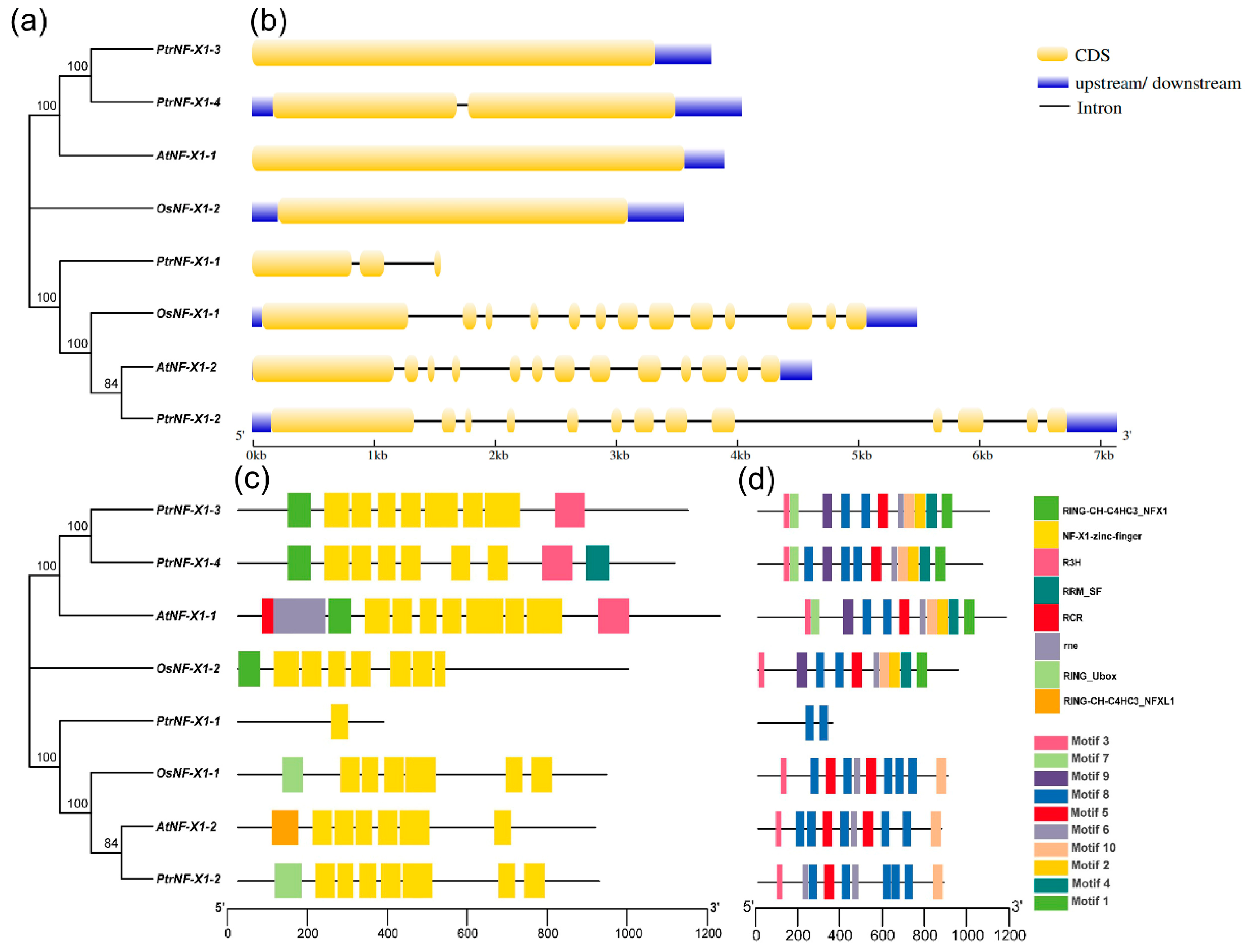
2.3. Gene Structure, Conserved Domain, and Motif
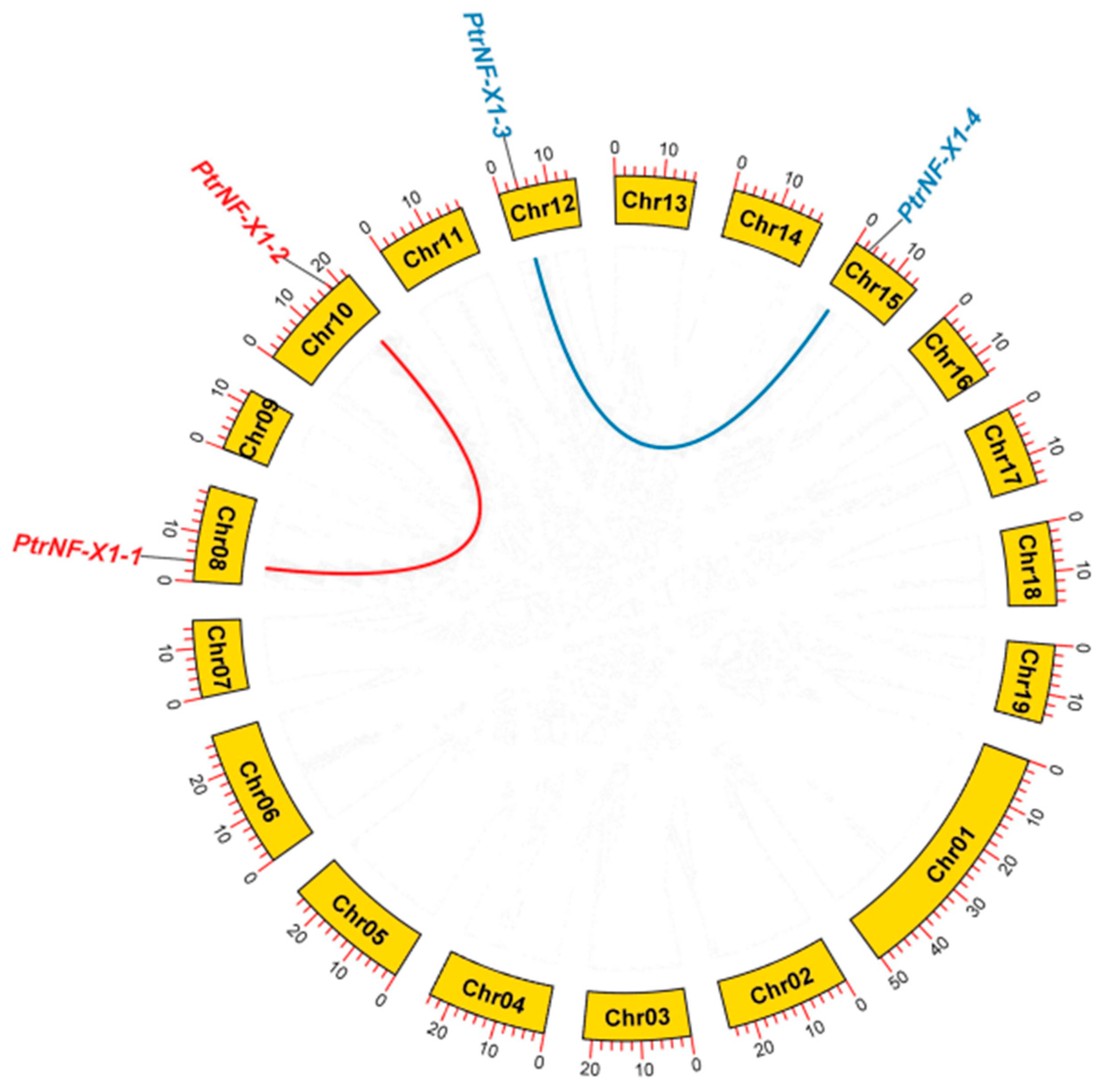
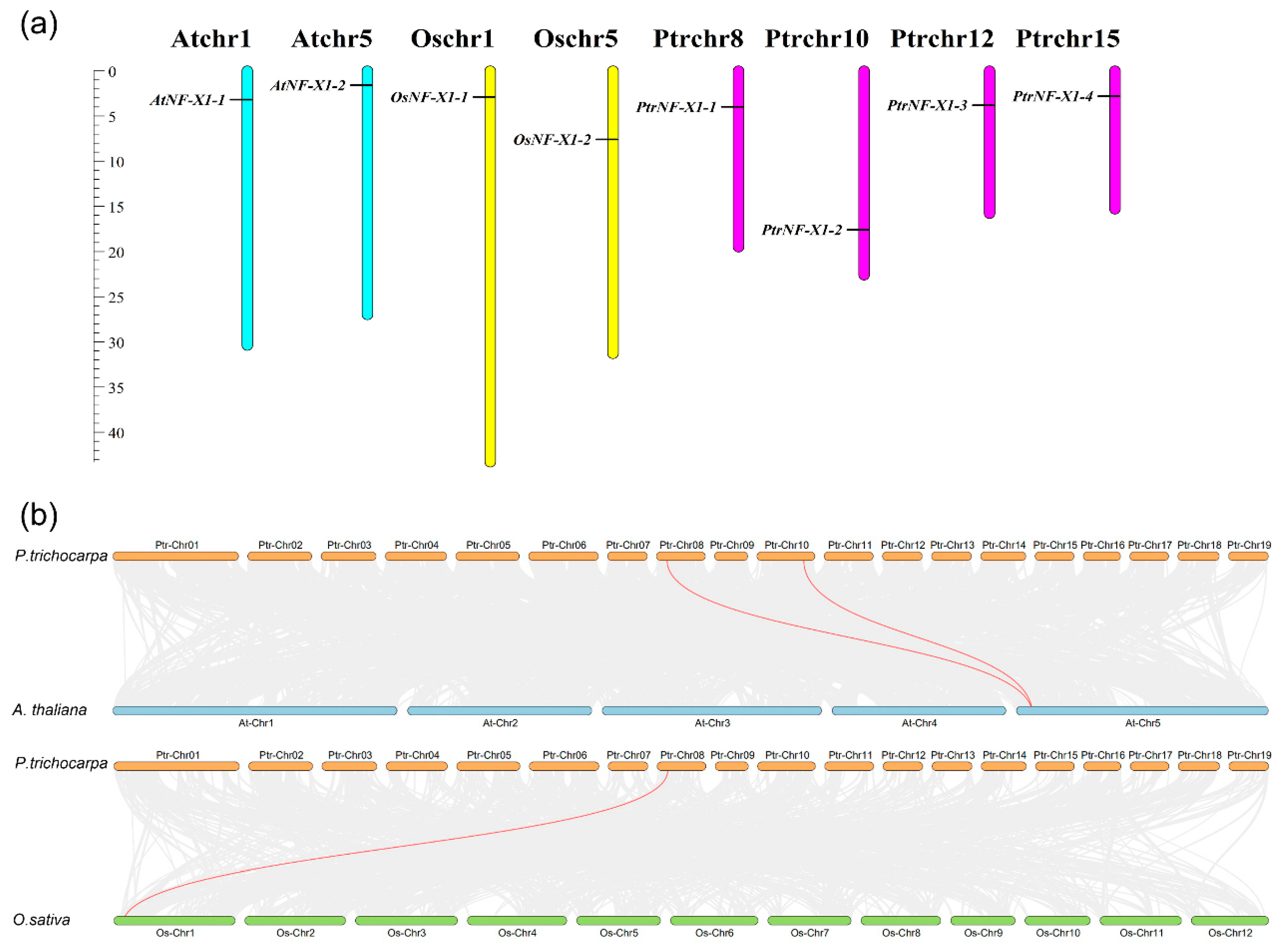
2.4. Analysis of Chromosomal Location and Gene Duplication
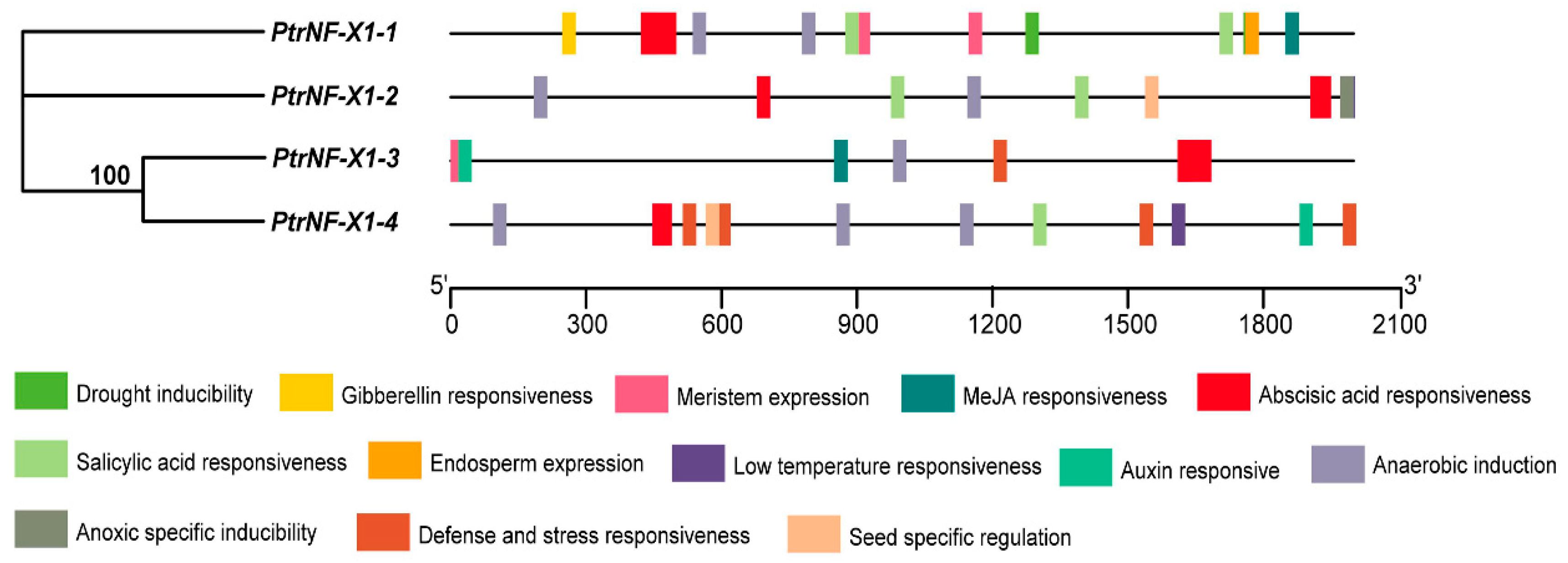
2.5. Cis-Elements Analysis of the PtrNF-X1
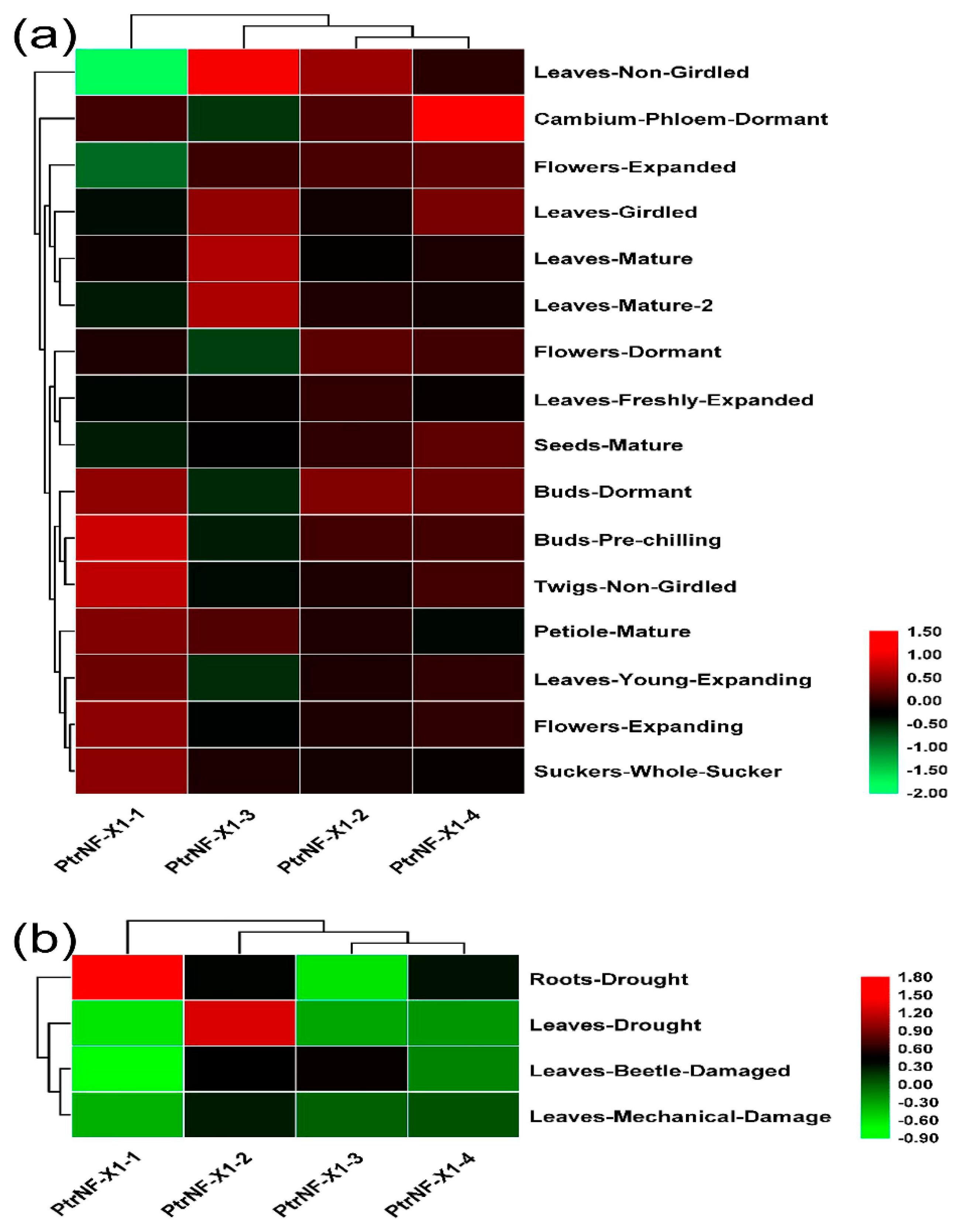
2.6. Expression Level of Poplar NF-X1 Genes with RNA-seq
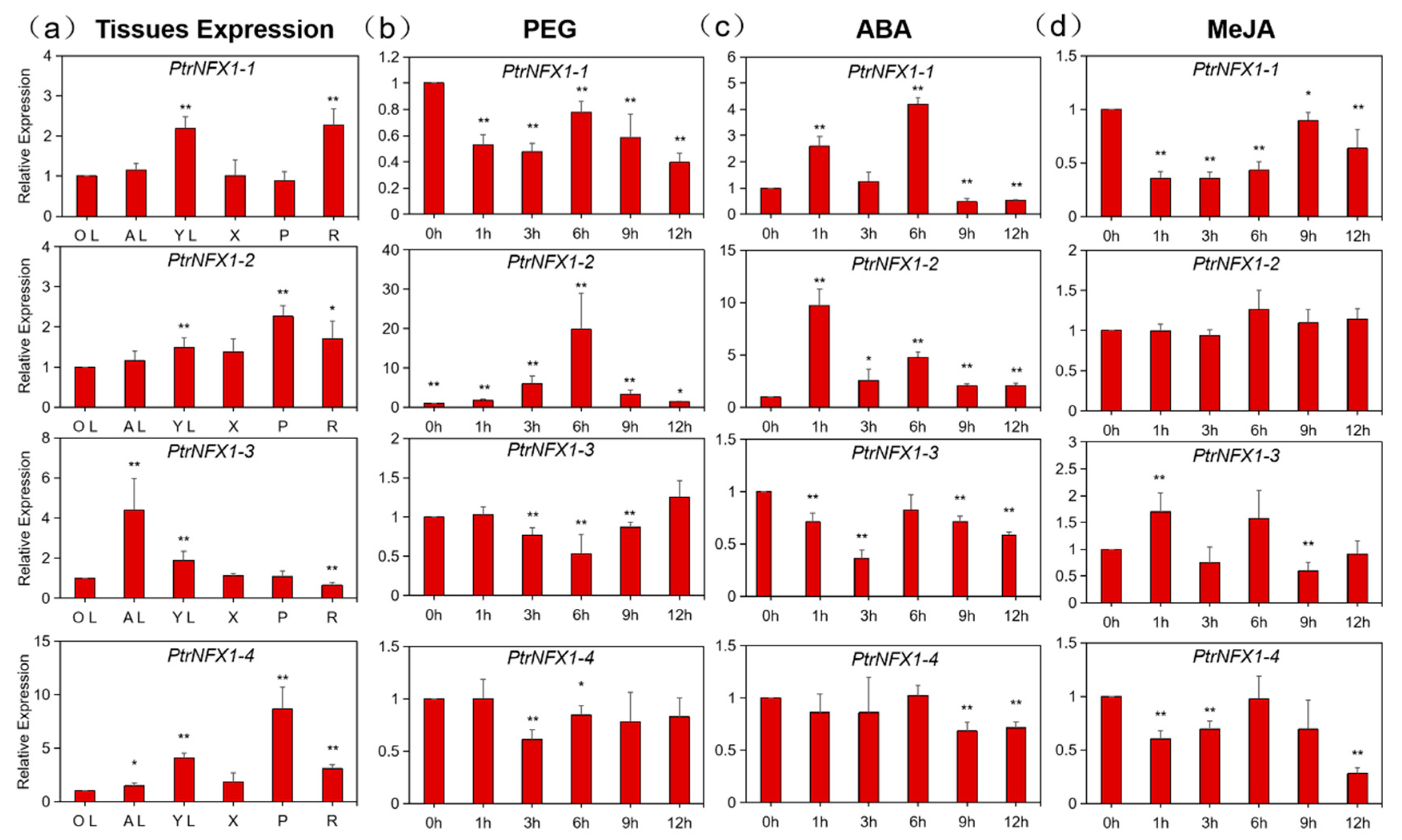
2.7. Expression Profile of PtrNF-X1 Genes in Different Plant Tissues and in Response to Different Treatments
3. Discussion
4. Materials and Methods
4.1. Retrieval of NF-X1 Genes in A. thaliana, O. sativa, and P. trichocarpa
4.2. Evolutionary Relationships of NF-X1 Genes
4.3. Analysis of Gene Structure, Protein Structure, and Motifs
4.4. Chromosomal Locations and Gene Duplication
4.5. Analysis of Cis-Regulatory Elements
4.6. Transcriptomic Data Sets to Analyze the Expression Patterns of PtrNF-X1
4.7. Plant Materials and Treatments
4.8. RNA Extraction and Quantitative Real-Time (qRT-PCR) Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhou, J.M.; Zhang, Y. Plant Immunity: Danger Perception and Signaling. Cell 2020, 181, 978–989. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Li, B.; Iwakawa, H.O.; Pan, Y.; Tang, X.; Ling-Hu, Q.; Liu, Y.; Sheng, S.; Feng, L.; Zhang, H.; et al. Plant 22-nt siRNAs mediate translational repression and stress adaptation. Nature 2020, 581, 89–93. [Google Scholar] [CrossRef]
- Mauch-Mani, B.; Baccelli, I.; Luna, E.; Flors, V. Defense Priming: An Adaptive Part of Induced Resistance. Annu. Rev. Plant Biol. 2017, 68, 485–512. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Zhu, Y.X.; Balint-Kurti, P.J.; Wang, G.F. Fine-Tuning Immunity: Players and Regulators for Plant NLRs. Trends Plant Sci. 2020, 25, 695–713. [Google Scholar] [CrossRef] [PubMed]
- Xing, H.; Fu, X.; Yang, C.; Tang, X.; Guo, L.; Li, C.; Xu, C.; Luo, K. Genome-wide investigation of pentatricopeptide repeat gene family in poplar and their expression analysis in response to biotic and abiotic stresses. Sci. Rep. 2018, 8, 2817. [Google Scholar] [CrossRef] [Green Version]
- Filichkin, S.A.; Hamilton, M.; Dharmawardhana, P.D.; Singh, S.K.; Sullivan, C.; Ben-Hur, A.; Reddy, A.S.N.; Jaiswal, P. Abiotic Stresses Modulate Landscape of Poplar Transcriptome via Alternative Splicing, Differential Intron Retention, and Isoform Ratio Switching. Front. Plant Sci. 2018, 9, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, T.; Shi, C.; Peng, Y.; Tan, H.; Xin, P.; Yang, Y.; Wang, F.; Li, X.; Chu, J.; Huang, J.; et al. Brassinosteroid-Activated BRI1-EMS-SUPPRESSOR 1 Inhibits Flavonoid Biosynthesis and Coordinates Growth and UV-B Stress Responses in Plants. Plant Cell 2020, 32, 3224–3239. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Liu, J.H.; Ma, X.; Luo, D.X.; Gong, Z.H.; Lu, M.H. The Plant Heat Stress Transcription Factors (HSFs): Structure, Regulation, and Function in Response to Abiotic Stresses. Front. Plant Sci 2016, 7, 114. [Google Scholar] [CrossRef] [Green Version]
- Lozano-Juste, J.; Alrefaei, A.F.; Rodriguez, P.L. Plant Osmotic Stress Signaling: MAPKKKs Meet SnRK2s. Trends Plant Sci. 2020. [Google Scholar] [CrossRef]
- Lin, Z.; Li, Y.; Zhang, Z.; Liu, X.; Hsu, C.C.; Du, Y.; Sang, T.; Zhu, C.; Wang, Y.; Satheesh, V.; et al. A RAF-SnRK2 kinase cascade mediates early osmotic stress signaling in higher plants. Nat. Commun. 2020, 11, 613. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, J.P.; Tian, F.; Yang, D.C.; Meng, Y.Q.; Kong, L.; Luo, J.C.; Gao, G. PlantTFDB 4.0: Toward a central hub for transcription factors and regulatory interactions in plants. Nucleic Acids Res. 2017, 45, D1040–D1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Udvardi, M.K.; Kakar, K.; Wandrey, M.; Montanari, O.; Murray, J.; Andriankaja, A.; Zhang, J.-Y.; Benedito, V.; Hofer, J.M.I.; Chueng, F.; et al. Legume Transcription Factors: Global Regulators of Plant Development and Response to the Environment. Plant Physiol. 2007, 144, 538–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akhtar, M.; Jaiswal, A.; Taj, G.; Jaiswal, J.P.; Qureshi, M.I.; Singh, N.K. DREB1/CBF transcription factors: Their structure, function and role in abiotic stress tolerance in plants. J. Genet. 2012, 91, 385–395. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Dong, Q.; Shao, Y.; Jiang, H.; Zhu, S.; Cheng, B.; Xiang, Y. Genome-wide survey and characterization of the WRKY gene family in Populus trichocarpa. Plant Cell Rep. 2012, 31, 1199–1217. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Wang, M.; Gong, X.; Liu, J.H. The transcription factor FcWRKY40 of Fortunella crassifolia functions positively in salt tolerance through modulation of ion homeostasis and proline biosynthesis by directly regulating SOS2 and P5CS1 homologs. New Phytol. 2018, 219, 972–989. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Song, M.; Guo, Y.; Liu, L.; Xue, H.; Dai, H.; Zhang, Z. MdMYB46 could enhance salt and osmotic stress tolerance in apple by directly activating stress-responsive signals. Plant Biotechnol. J. 2019. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Guo, X.; Zhang, M.; Wang, X.; Zhao, Y.; Yin, Z.; Zhang, Z.; Wang, Y.; Xiong, H.; Zhang, H.; et al. OsERF71 confers drought tolerance via modulating ABA signaling and proline biosynthesis. Plant Sci. 2018, 270, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. BMC Plant Biol. 2016, 16. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, D.J.; Conde, J.V.; Berckhan, S.; Prasad, G.; Mendiondo, G.M.; Holdsworth, M.J. Group VII Ethylene Response Factors Coordinate Oxygen and Nitric Oxide Signal Transduction and Stress Responses in Plants. Plant Physiol. 2015, 169, 23–31. [Google Scholar] [CrossRef]
- Mussig, C.; Schroder, F.; Usadel, B.; Lisso, J. Structure and putative function of NFX1-like proteins in plants. Plant Biol. 2010, 12, 381–394. [Google Scholar] [CrossRef]
- Song, Z.; Krishna, S.; Thanos, D.; Strominger, J.L.; Ono, S.J. A Novel Cysteine-rich Sequence-specific DNA-binding Protein Interacts with the Conserved X-box Motif of the Human Major Histocompatibility Complex Class II Genes Via a Repeated Cys-His Domain and Functions as a Transcriptional Repressor. J. Exp. Med. 1994, 180, 1763–1774. [Google Scholar] [CrossRef] [Green Version]
- Stroumbakis, N.D.; Li, Z.; Tolias, P.P. A Homolog of Human Transcription Factor NF-X1 Encoded by the Drosophila shuttle craft Gene Is Required in the Embryonic Central Nervous System. Am. Soc. Microbiol. 1996, 16, 192–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.S.; Lindsay, L.; Hoots, B.; Keys, J.; Franceschi, S.; Winer, R.; Clifford, G.M. Human papillomavirus type distribution in invasive cervical cancer and high-grade cervical lesions: A meta-analysis update. Int. J. Cancer 2007, 121, 621–632. [Google Scholar] [CrossRef] [PubMed]
- Felix, A.; Alemany, L.; Tous, S.; de Sanjose, S.; Bosch, F.X. HPV distribution in cervical cancer in Portugal. A retrospective study from 1928 to 2005. Papillomavirus Res. 2016, 2, 41–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lisso, J.; Altmann, T.; Mussig, C. The AtNFXL1 gene encodes a NF-X1 type zinc finger protein required for growth under salt stress. FEBS Lett. 2006, 580, 4851–4856. [Google Scholar] [CrossRef] [Green Version]
- Satapathy, L.; Mukhopadhyay, K. Transcriptome-wide analysis of NFX1 transcription factors in wheat (Triticum aestivum L.) and their leaf rust responsive expression profiling. Int. J. Pharm. Pharm. Sci. 2016, 8, 60–65. [Google Scholar]
- Righetti, K.; Vu, J.L.; Pelletier, S.; Vu, B.L.; Glaab, E.; Lalanne, D.; Pasha, A.; Patel, R.V.; Provart, N.J.; Verdier, J.; et al. Inference of Longevity-Related Genes from a Robust Coexpression Network of Seed Maturation Identifies Regulators Linking Seed Storability to Biotic Defense-Related Pathways. Plant Cell 2015, 27, 2692–2708. [Google Scholar] [CrossRef] [Green Version]
- He, F.; Li, H.G.; Wang, J.J.; Su, Y.; Wang, H.L.; Feng, C.H.; Yang, Y.; Niu, M.X.; Liu, C.; Yin, W.; et al. PeSTZ1, a C2H2-type zinc finger transcription factor from Populus euphratica, enhances freezing tolerance through modulation of ROS scavenging by directly regulating PeAPX2. Plant Biotechnol. J. 2019. [Google Scholar] [CrossRef] [Green Version]
- Xie, T.; Chen, C.; Li, C.; Liu, J.; Liu, C.; He, Y. Genome-wide investigation of WRKY gene family in pineapple: Evolution and expression profiles during development and stress. BMC Genom. 2018, 19, 490. [Google Scholar] [CrossRef] [Green Version]
- Jansson, S.; Douglas, C.J. Populus: A model system for plant biology. Annu. Rev. Plant Biol. 2007, 58, 435–458. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Xu, X.; Li, C. Identification and expression of CAMTA genes in Populus trichocarpa under biotic and abiotic stress. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, J.; Cai, B.; Peng, R.H.; Zhu, B.; Jin, X.F.; Xue, Y.; Gao, F.; Fu, X.Y.; Tian, Y.S.; Zhao, W.; et al. Genome-wide analysis of the AP2/ERF gene family in Populus trichocarpa. Biochem. Biophys. Res. Commun. 2008, 371, 468–474. [Google Scholar] [CrossRef]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Niu, M.X.; Feng, C.H.; Li, H.G.; Su, Y.; Su, W.L.; Pang, H.; Yang, Y.; Yu, X.; Wang, H.L.; et al. PeSTZ1 confers salt stress tolerance by scavenging the accumulation of ROS through regulating the expression of PeZAT12 and PeAPX2 in Populus. Tree Physiol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Zhang, H.; Song, C.; Zhu, J.-K.; Shabala, S. Mechanisms of Plant Responses and Adaptation to Soil Salinity. Innovation 2020, 1, 100017. [Google Scholar] [CrossRef]
- Yang, Y.; Li, H.G.; Wang, J.; Wang, H.L.; He, F.; Su, Y.; Zhang, Y.; Feng, C.H.; Niu, M.; Li, Z.; et al. PeABF3 Enhances Drought Tolerance via Promoting ABA-induced Stomatal Closure by directly Regulating PeADF5 in Poplar. J. Exp. Bot. 2020. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Meng, S.; Su, W.; Bao, Y.; Lu, Y.; Yin, W.; Liu, C.; Xia, X. Genome-Wide Analysis of Multiple Organellar RNA Editing Factor Family in Poplar Reveals Evolution and Roles in Drought Stress. Int. J. Mol. Sci. 2019, 20, 1425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, P.; Wang, D.; Wang, R.; Kong, N.; Zhang, C.; Yang, C.; Wu, W.; Ma, H.; Chen, Q. Genome-wide analysis of the potato Hsp20 gene family: Identification, genomic organization and expression profiles in response to heat stress. BMC Genom. 2018, 19, 61. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, D.; Hu, H.; Li, W.; Hu, Y.; Xie, J.; Huang, S.; Wang, W. Genome-wide characterization of a SRO gene family involved in response to biotic and abiotic stresses in banana (Musa spp.). BMC Plant Biol. 2019, 19, 211. [Google Scholar] [CrossRef]
- Moturu, T.R.; Thula, S.; Singh, R.K.; Nodzynski, T.; Varekova, R.S.; Friml, J.; Simon, S. Molecular evolution and diversification of the SMXL gene family. J. Exp. Bot. 2018, 69, 2367–2378. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Wang, R.; Liu, Z.; Wu, X.; Wang, J. Genome-wide identification and analysis of the WUSCHEL-related homeobox (WOX) gene family in allotetraploid Brassica napus reveals changes in WOX genes during polyploidization. BMC Genom. 2019, 20, 317. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Wang, Y.; Chai, F.; Li, S.; Xin, H.; Liang, Z. Genome-wide identification and characterization of the 14-3-3 family in Vitis vinifera L. during berry development and cold- and heat-stress response. BMC Genom. 2018, 19, 579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, S.; Dhugga, K.S.; Beech, R.; Singh, J. Genome-wide analysis of the cellulose synthase-like (Csl) gene family in bread wheat (Triticum aestivum L.). BMC Plant Biol. 2017, 17, 193. [Google Scholar] [CrossRef] [Green Version]
- Asano, T.; Masuda, D.; Yasuda, M.; Nakashita, H.; Kudo, T.; Kimura, M.; Yamaguchi, K.; Nishiuchi, T. AtNFXL1, an Arabidopsis homologue of the human transcription factor NF-X1, functions as a negative regulator of the trichothecene phytotoxin-induced defense response. Plant J. 2008, 53, 450–464. [Google Scholar] [CrossRef]
- Hewage, K.A.H.; Yang, J.F.; Wang, D.; Hao, G.F.; Yang, G.F.; Zhu, J.K. Chemical Manipulation of Abscisic Acid Signaling: A New Approach to Abiotic and Biotic Stress Management in Agriculture. Adv. Sci. 2020, 7, 2001265. [Google Scholar] [CrossRef] [PubMed]
- Aziz, E.; Batool, R.; Akhtar, W.; Rehman, S.; Gregersen, P.L.; Mahmood, T. Expression analysis of the polyphenol oxidase gene in response to signaling molecules, herbivory and wounding in antisense transgenic tobacco plants. 3 Biotech 2019, 9, 55. [Google Scholar] [CrossRef]
- Li, C.; Li, D.; Li, J.; Shao, F.; Lu, S. Characterization of the polyphenol oxidase gene family reveals a novel microRNA involved in posttranscriptional regulation of PPOs in Salvia miltiorrhiza. Sci. Rep. 2017, 7, 44622. [Google Scholar] [CrossRef] [Green Version]
- An, Y.; Zhou, Y.; Han, X.; Shen, C.; Wang, S.; Liu, C.; Yin, W.; Xia, X. The GATA transcription factor GNC plays an important role in photosynthesis and growth in poplar. J. Exp. Bot. 2020, 71, 1969–1984. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [Green Version]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Sundell, D.; Mannapperuma, C.; Netotea, S.; Delhomme, N.; Lin, Y.C.; Sjodin, A.; Van de Peer, Y.; Jansson, S.; Hvidsten, T.R.; Street, N.R. The Plant Genome Integrative Explorer Resource: PlantGenIE.org. New Phytol. 2015, 208, 1149–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuskan, G.A.; Difazio, S.; Jansson, S.; Bohlmann, J.; Grigoriev, I.; Hellsten, U.; Putnam, N.; Ralph, S.; Rombauts, S.; Salamov, A.; et al. The genome of black cottonwood, Populus trichocarpa (Torr. & Gray). Science 2006, 313, 1596–1604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, F.; Wang, H.L.; Li, H.G.; Su, Y.; Li, S.; Yang, Y.; Feng, C.H.; Yin, W.; Xia, X. PeCHYR1, a ubiquitin E3 ligase from Populus euphratica, enhances drought tolerance via ABA-induced stomatal closure by ROS production in Populus. Plant Biotechnol. J. 2018. [Google Scholar] [CrossRef] [Green Version]
- Lim, C.W.; Baek, W.; Lee, S.C. The Pepper RING-Type E3 Ligase CaAIRF1 Regulates ABA and Drought Signaling via CaADIP1 Protein Phosphatase Degradation. Plant Physiol. 2017, 173, 2323–2339. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Li, Y.; Yin, Z.; Jiang, J.; Zhang, M.; Guo, X.; Ye, Z.; Zhao, Y.; Xiong, H.; Zhang, Z.; et al. OsASR5 enhances drought tolerance through a stomatal closure pathway associated with ABA and H2 O2 signalling in rice. Plant Biotechnol. J. 2017, 15, 183–196. [Google Scholar] [CrossRef]
- Duan, M.; Zhang, R.; Zhu, F.; Zhang, Z.; Gou, L.; Wen, J.; Dong, J.; Wang, T. A Lipid-Anchored NAC Transcription Factor Is Translocated into the Nucleus and Activates Glyoxalase I Expression during Drought Stress. Plant Cell 2017, 29, 1748–1772. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Gene Model ID | Description | Pfam ID | gDNA | Transcript | CDS | Domains | AA | MW | GRAVY | PI | Homologs in Arabidopsis |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AtNF-X1-1 | AT1G10170.1 | NF-X1 family protein | PF01422 | 3897 | 3897 | 3567 | 58-134/86-215/222-279/312-373/380-428/448-489/503-550/562-622/601-653/658-705/711-766/749-798/888-963 | 1188 | 130.72 | −0.601 | 8.54 | / |
| AtNF-X1-2 | AT5G05660.1 | NF-X1 family protein | PF01422 | 4617 | 2908 | 2643 | 82-149/183-231/237-285/290-330/344-393/397-445/424-472/631-672 | 880 | 98.88 | −0.605 | 8.73 | / |
| OsNF-X1-1 | LOC_Os01g06550.1 | NF-X1 family protein | PF01422 | 5482 | 3225 | 2730 | 109-160/252-300/305-345/359-408/412-460/439-487/659-700/723-774 | 910 | 99.56 | −0.432 | 8.87 | AT5G05660.1(AtNF-X1-2) |
| OsNF-X1-2 | LOC_Os06g14190.1 | NF-X1 family protein | PF01422 | 3559 | 3559 | 2886 | 1-54/87-150/157-205/221-264/279-327/374-426/431-479/484-510 | 962 | 103.89 | −0.346 | 8.34 | AT1G10170.1(AtNF-X1-1) |
| PtrNF-X1-1 | Potri.008G068500.1 | NF-X1 family protein | PF01422 | 1557 | 1077 | 1077 | 228-272 | 358 | 39.77 | −0.099 | 8.67 | AT5G05660.1(AtNF-X1-2) |
| PtrNF-X1-2 | Potri.010G188700.1 | NF-X1 family protein | PF01422 | 7128 | 3237 | 2673 | 90-157/190-238/244-284/299-340/351-400/404-452/431-479/641-682/705-756 | 891 | 99.75 | −0.616 | 8.81 | AT5G05660.1(AtNF-X1-2) |
| PtrNF-X1-3 | Potri.012G043700.1 | NF-X1 family protein | PF01422/PF01424 | 3786 | 3786 | 3327 | 122-179/212-273/280-328/344-387/402-450/460-519/498-541/555-603/608-663/646-695/781-854 | 1109 | 119.81 | −0.429 | 8.24 | AT1G10170.1(AtNF-X1-1) |
| PtrNF-X1-4 | Potri.015G034500.1 | NF-X1 family protein | PF01422 | 4035 | 3920 | 3231 | 122-179/212-273/280-328344-387/402-450/524-572/615-664/750-823/858-915 | 1077 | 116.40 | −0.375 | 8.55 | AT1G10170.1(AtNF-X1-1) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, F.; Shi, Y.-J.; Mi, J.-X.; Zhao, K.-J.; Cui, X.-L.; Chen, L.-H.; Yang, H.-B.; Zhang, F.; Zhao, Q.; Huang, J.-L.; et al. Genome-Wide Investigation of the NF-X1 Gene Family in Populus trichocarpa Expression Profiles during Development and Stress. Int. J. Mol. Sci. 2021, 22, 4664. https://doi.org/10.3390/ijms22094664
He F, Shi Y-J, Mi J-X, Zhao K-J, Cui X-L, Chen L-H, Yang H-B, Zhang F, Zhao Q, Huang J-L, et al. Genome-Wide Investigation of the NF-X1 Gene Family in Populus trichocarpa Expression Profiles during Development and Stress. International Journal of Molecular Sciences. 2021; 22(9):4664. https://doi.org/10.3390/ijms22094664
Chicago/Turabian StyleHe, Fang, Yu-Jie Shi, Jia-Xuan Mi, Kuang-Ji Zhao, Xing-Lei Cui, Liang-Hua Chen, Han-Bo Yang, Fan Zhang, Qian Zhao, Jin-Liang Huang, and et al. 2021. "Genome-Wide Investigation of the NF-X1 Gene Family in Populus trichocarpa Expression Profiles during Development and Stress" International Journal of Molecular Sciences 22, no. 9: 4664. https://doi.org/10.3390/ijms22094664
APA StyleHe, F., Shi, Y. -J., Mi, J. -X., Zhao, K. -J., Cui, X. -L., Chen, L. -H., Yang, H. -B., Zhang, F., Zhao, Q., Huang, J. -L., & Wan, X. -Q. (2021). Genome-Wide Investigation of the NF-X1 Gene Family in Populus trichocarpa Expression Profiles during Development and Stress. International Journal of Molecular Sciences, 22(9), 4664. https://doi.org/10.3390/ijms22094664