
Identification and Expression Analysis of SLAC/SLAH Gene Family in Brassica napus L.


 ,
,
Abstract
:1. Introduction
2. Results
2.1. Identification and Classification of SLAC/SLAH Genes in B. napus
2.2. Phylogenetic Analysis of the SLAC/SLAHs between B. napus and Other Species and Identification of Evolutionary Selection Pressure on BnSLAC/SLAHs
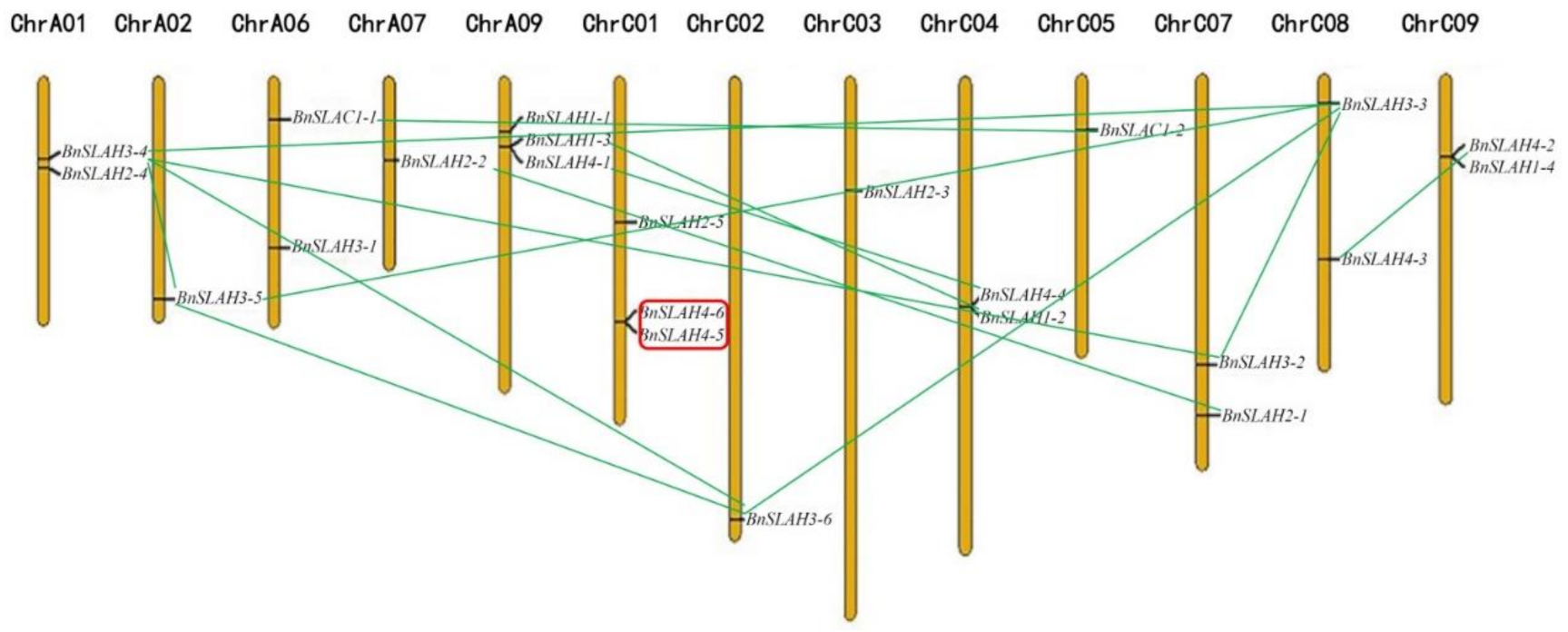
2.3. Chromosomal Location and Duplication Patterns
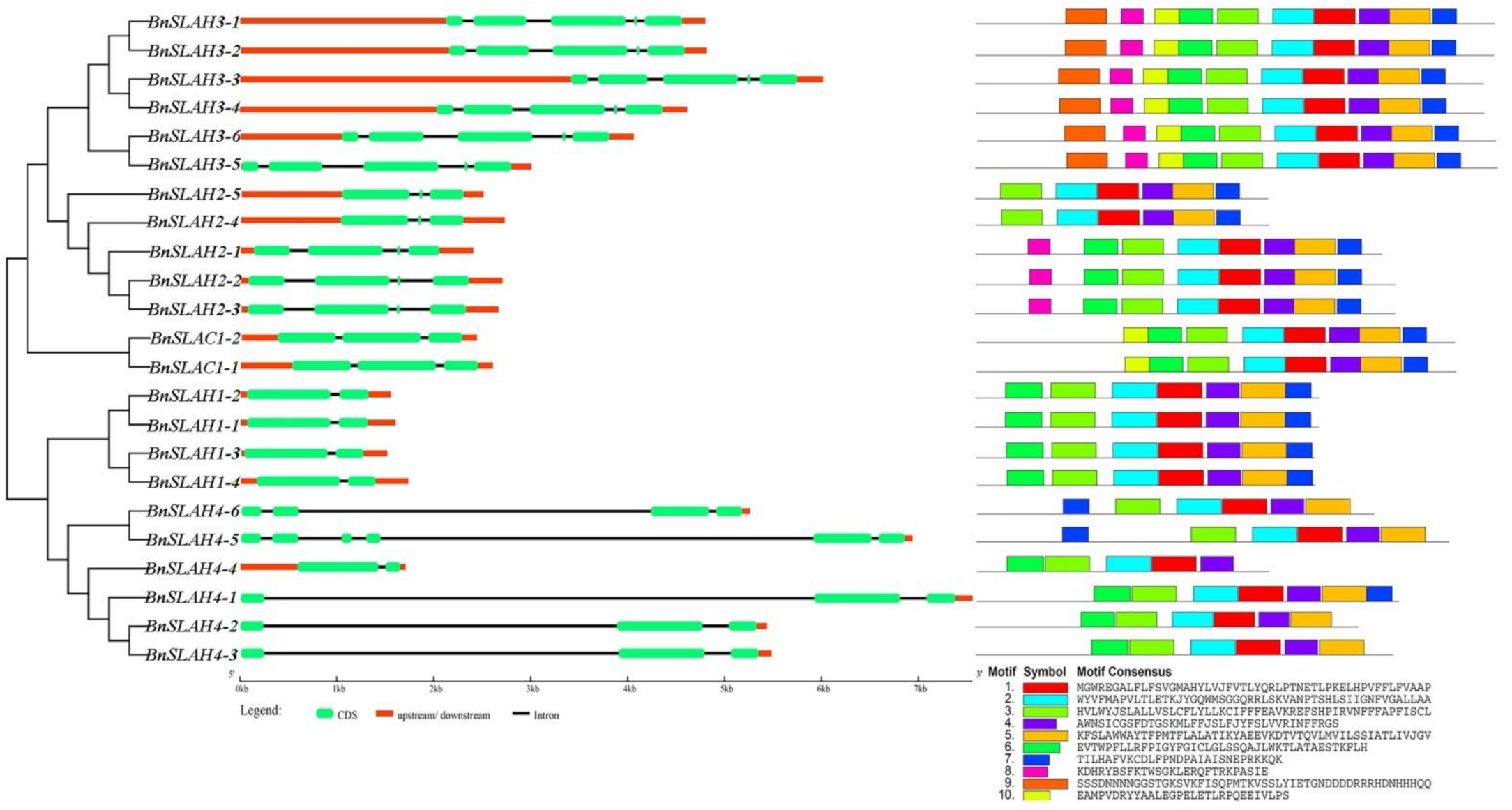
2.4. Gene Structure and Conserved Motif Analysis of the SLAC/SLAH Family Members in B. napus
2.5. Promoter cis-Acting Element Analysis and Interaction Protein Prediction
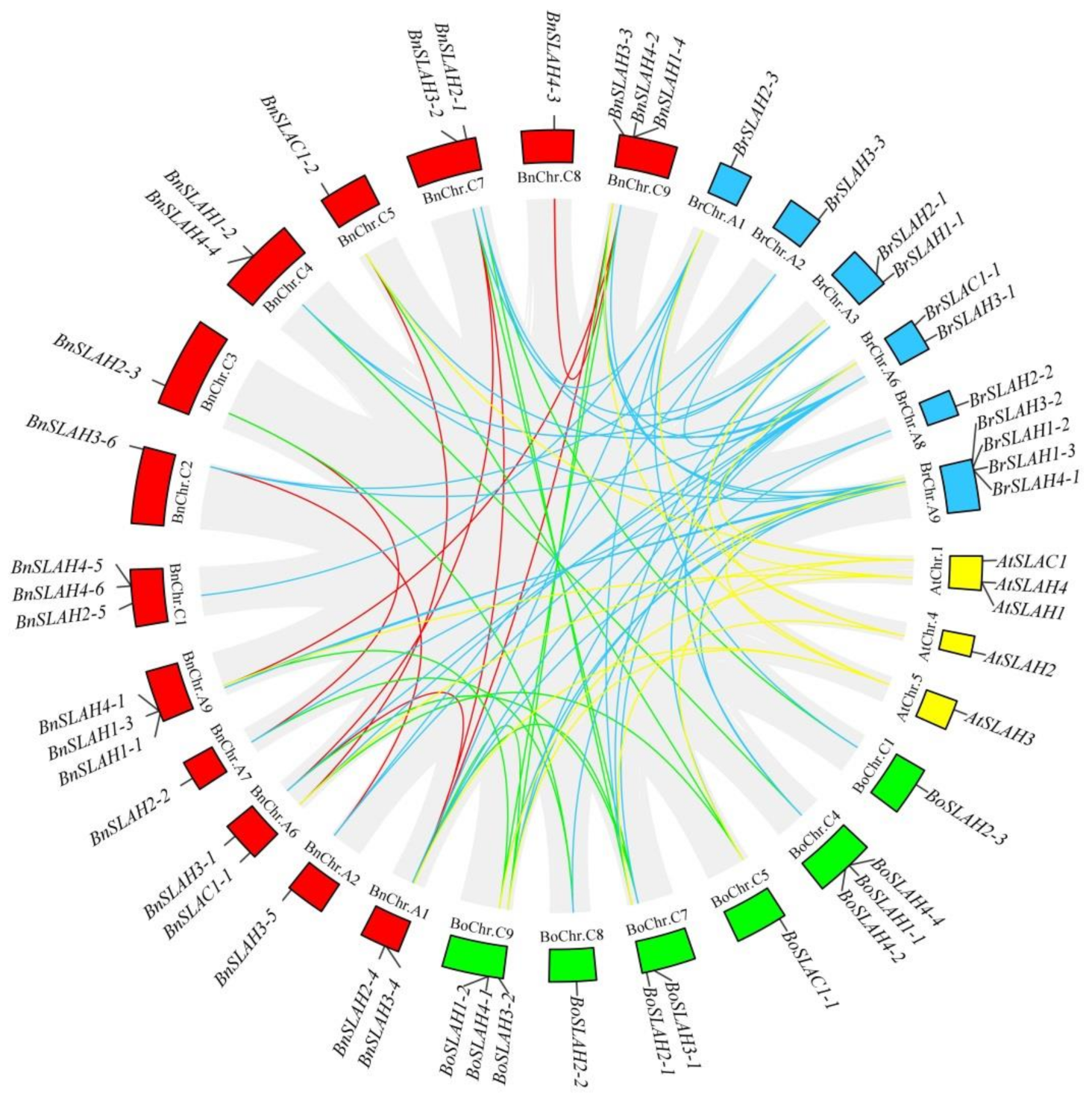
2.6. Synteny Analysis
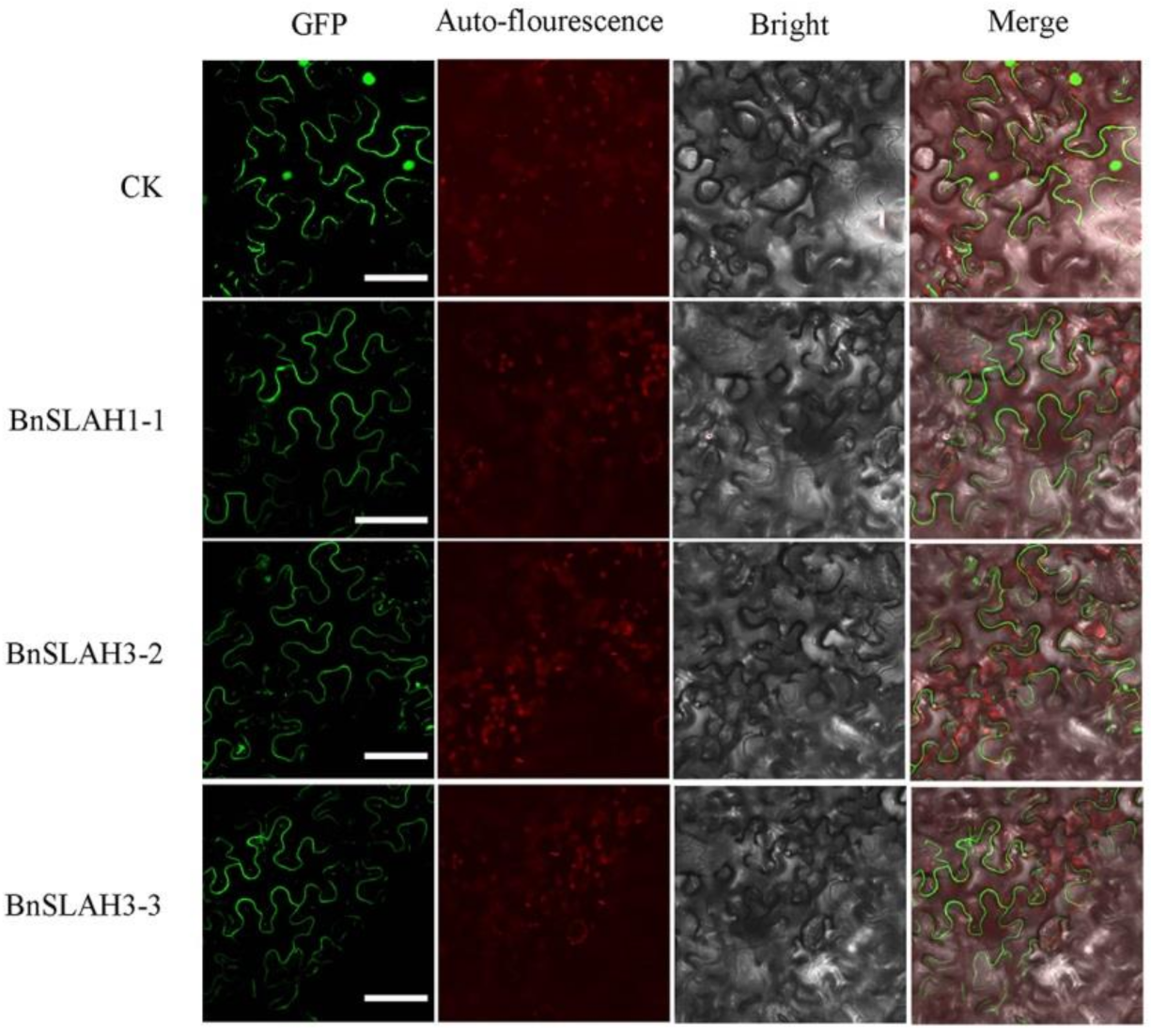
2.7. Subcellular Localization of the SLAC/SLAH Genes of B. napus
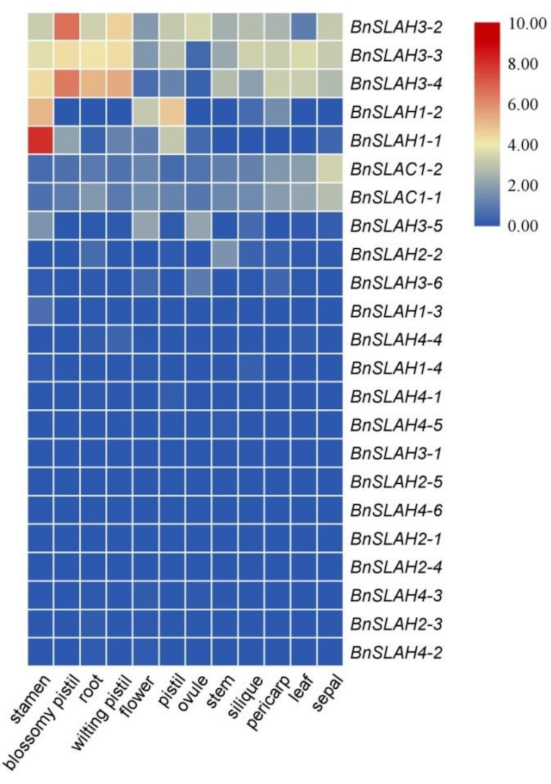
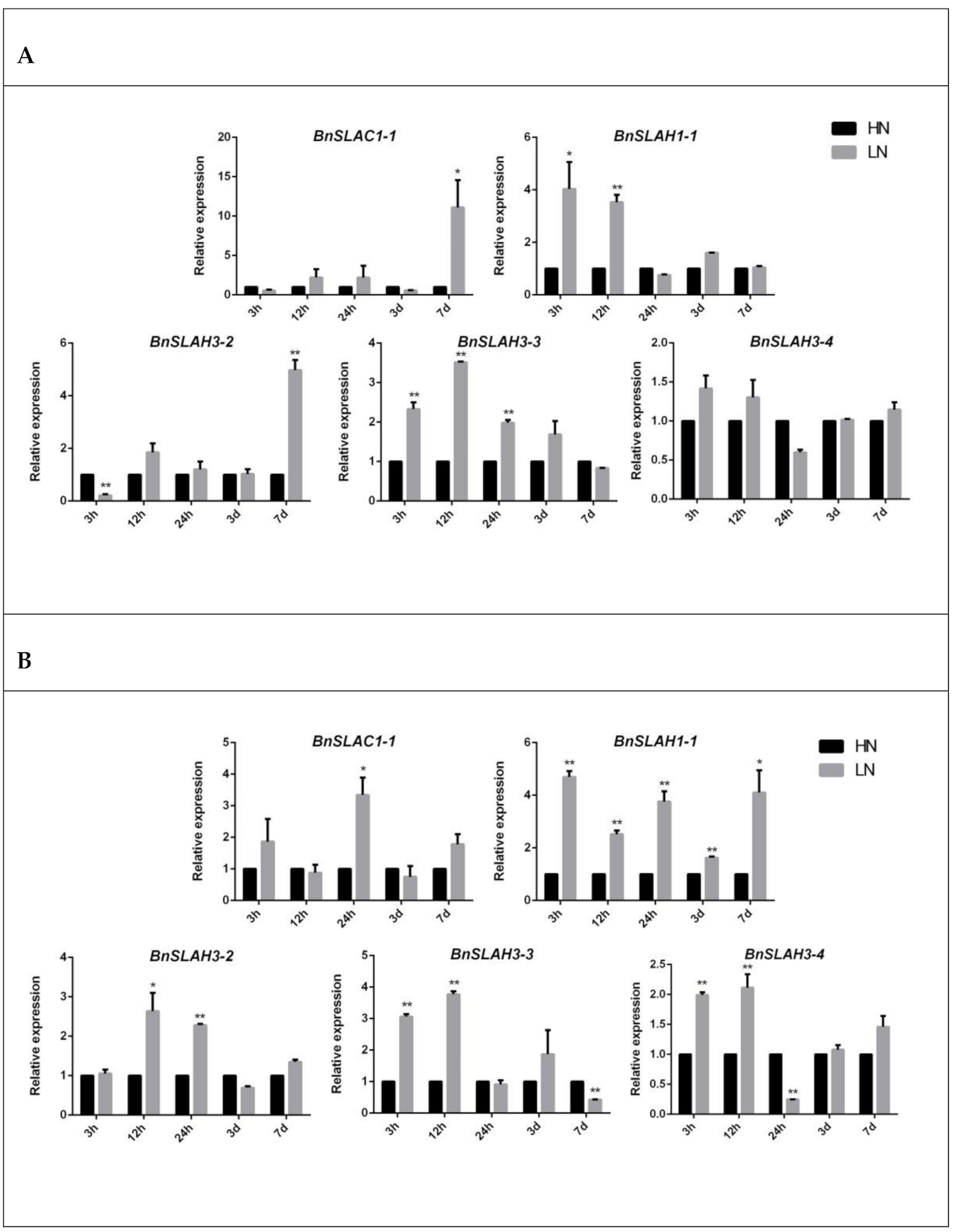
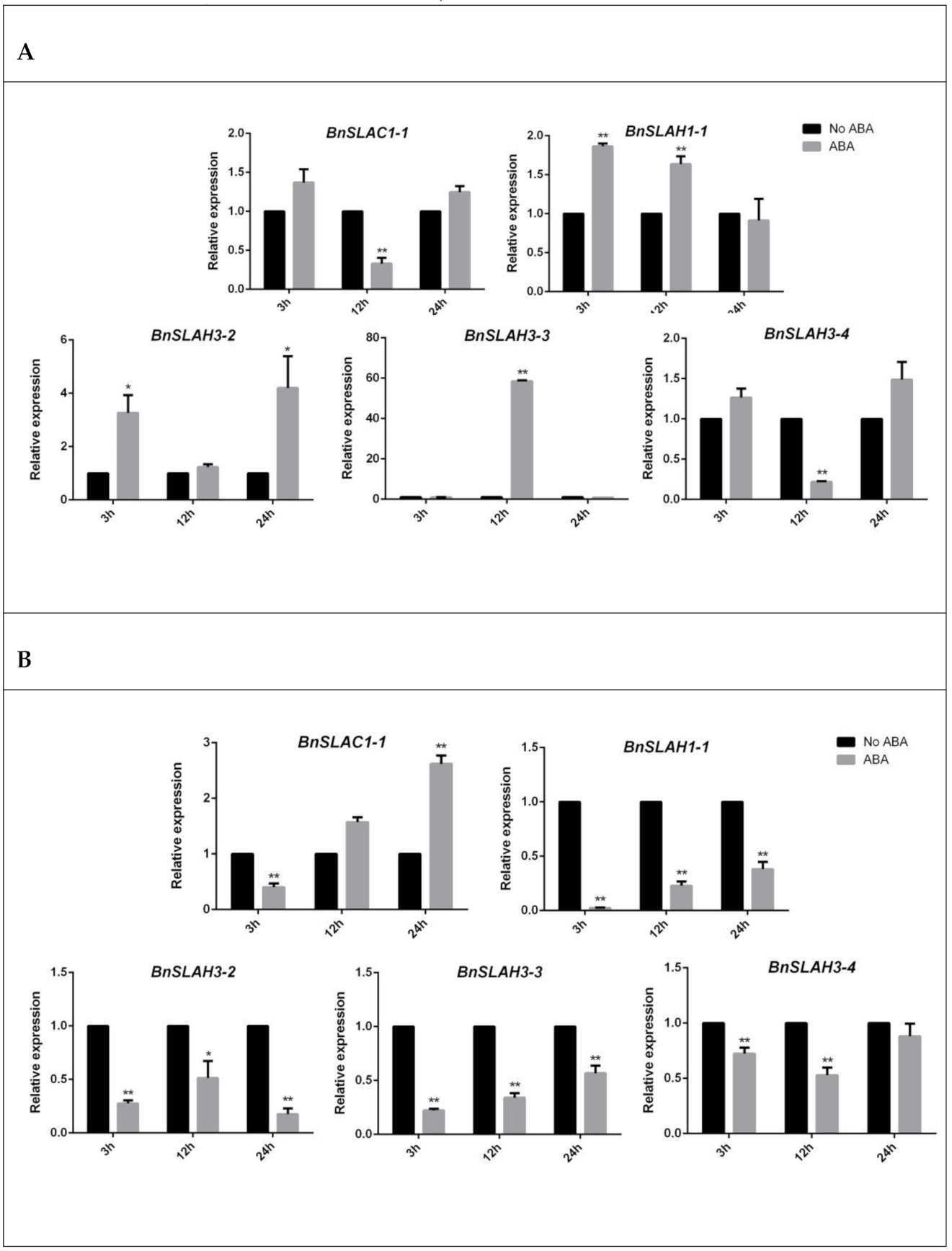
2.8. Expression Level of the SLAC/SLAH Genes in B. napus
3. Discussion
4. Materials and Methods
4.1. Identification of SLAC/SLAH Gene Family Members in B. napus
4.2. Phylogenetic and Synteny Analysis of SLAC/SLAH
4.3. Sequence Analysis of SLAC/SLAH Genes/Proteins in B. napus
4.4. Chromosomal Location and Duplication of SLAC/SLAH Genes in B. napus
4.5. Promoter cis-Acting Element Analysis and Interaction Protein Prediction
4.6. Subcellular Localization of the SLAC/SLAHs in B. napus
4.7. Plant Material and Stress Treatment
4.8. Gene Expression Analysis of SLAC/SLAHs in B. napus
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Roelfsema, M.R.G.; Hedrich, R.; Geiger, D. Anion channels: Master switches of stress responses. Trends Plant Sci. 2012, 17, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Vahisalu, T.; Kollist, H.; Wang, Y.F.; Nishimura, N.; Chan, W.Y.; Valerio, G.; Lamminmaki, A.; Brosche, M.; Moldau, H.; Desikan, R.; et al. SLAC1 is required for plant guard cell S-type anion channel function in stomatal signalling. Nature 2008, 452, 487-U415. [Google Scholar] [CrossRef]
- Brandt, B.; Brodsky, D.E.; Xue, S.W.; Negi, J.; Iba, K.; Kangasjarvi, J.; Ghassemian, M.; Stephan, A.B.; Hu, H.H.; Schroeder, J.I. Reconstitution of abscisic acid activation of SLAC1 anion channel by CPK6 and OST1 kinases and branched ABI1 PP2C phosphatase action. Proc. Natl. Acad. Sci. USA 2012, 109, 10593–10598. [Google Scholar] [CrossRef] [Green Version]
- Geiger, D.; Scherzer, S.; Mumm, P.; Marten, I.; Ache, P.; Matschi, S.; Liese, A.; Wellmann, C.; Al-Rasheid, K.A.S.; Grill, E.; et al. Guard cell anion channel SLAC1 is regulated by CDPK protein kinases with distinct Ca2+ affinities. Proc. Natl. Acad. Sci. USA 2010, 107, 8023–8028. [Google Scholar] [CrossRef] [Green Version]
- Geiger, D.; Maierhofer, T.; AL-Rasheid, K.A.S.; Scherzer, S.; Mumm, P.; Liese, A.; Ache, P.; Wellmann, C.; Marten, I.; Grill, E.; et al. Stomatal Closure by Fast Abscisic Acid Signaling Is Mediated by the Guard Cell Anion Channel SLAH3 and the Receptor RCAR1. Sci. Signal 2011, 4, ra32. [Google Scholar] [CrossRef] [PubMed]
- Vahisalu, T.; Puzorjova, I.; Brosche, M.; Valk, E.; Lepiku, M.; Moldau, H.; Pechter, P.; Wang, Y.S.; Lindgren, O.; Salojarvi, J.; et al. Ozone-triggered rapid stomatal response involves the production of reactive oxygen species, and is controlled by SLAC1 and OST1. Plant J. 2010, 62, 442–453. [Google Scholar] [CrossRef] [PubMed]
- Schafer, N.; Maierhofer, T.; Herrmann, J.; Jorgensen, M.E.; Lind, C.; von Meyer, K.; Lautner, S.; Fromm, J.; Felder, M.; Hetherington, A.M.; et al. A Tandem Amino Acid Residue Motif in Guard Cell SLAC1 Anion Channel of Grasses Allows for the Control of Stomatal Aperture by Nitrate. Curr. Biol. 2018, 28, 1370-U1145. [Google Scholar] [CrossRef] [Green Version]
- Gutermuth, T.; Lassig, R.; Portes, M.T.; Maierhofer, T.; Romeis, T.; Borst, J.W.; Hedrich, R.; Feijo, J.A.; Konrad, K.R. Pollen Tube Growth Regulation by Free Anions Depends on the Interaction between the Anion Channel SLAH3 and Calcium-Dependent Protein Kinases CPK2 and CPK20. Plant Cell 2013, 25, 4525–4543. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.J.; He, K.; Kleist, T.; Chen, F.; Luan, S. Anion channel SLAH3 functions in nitrate-dependent alleviation of ammonium toxicity in Arabidopsis. Plant Cell Environ. 2015, 38, 474–486. [Google Scholar] [CrossRef] [PubMed]
- Maierhofer, T.; Lind, C.; Huttl, S.; Scherzer, S.; Papenfuss, M.; Simon, J.; Al-Rasheid, K.A.S.; Ache, P.; Rennenberg, H.; Hedrich, R.; et al. A Single-Pore Residue Renders the Arabidopsis Root Anion Channel SLAH2 Highly Nitrate Selective. Plant Cell 2014, 26, 2554–2567. [Google Scholar] [CrossRef] [Green Version]
- Qiu, J.E.; Henderson, S.W.; Tester, M.; Roy, S.J.; Gilliham, M. SLAH1, a homologue of the slow type anion channel SLAC1, modulates shoot Cl- accumulation and salt tolerance in Arabidopsis thaliana. J. Exp. Bot. 2016, 67, 4495–4505. [Google Scholar] [CrossRef] [Green Version]
- Cubero-Font, P.; Maierhofer, T.; Jaslan, J.; Rosales, M.A.; Espartero, J.; Diaz-Rueda, P.; Muller, H.M.; Hurter, A.L.; AL-Rasheid, K.A.S.; Marten, I.; et al. Silent S-Type Anion Channel Subunit SLAH1 Gates SLAH3 Open for Chloride Root-to-Shoot Translocation. Curr. Biol. 2016, 26, 2213–2220. [Google Scholar] [CrossRef] [Green Version]
- Geiger, D.; Scherzer, S.; Mumm, P.; Stange, A.; Marten, I.; Bauer, H.; Ache, P.; Matschi, S.; Liese, A.; Al-Rasheid, K.A.S.; et al. Activity of guard cell anion channel SLAC1 is controlled by drought-stress signaling kinase-phosphatase pair. Proc. Natl. Acad. Sci. USA 2009, 106, 21425–21430. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.; Fang, X.; Xiao, C.; Ma, Z.; Huang, X.; Su, J.; Li, J.; Wang, J.; Wang, S.; Luan, S.; et al. Kinase SnRK1.1 Regulates nitrate channel SLAH3 Engaged in Nitrate-Dependent Alleviation of Ammonium Toxicity. Plant Physiol. 2021. [Google Scholar] [CrossRef]
- Sun, S.J.; Qi, G.N.; Gao, Q.F.; Wang, H.Q.; Yao, F.Y.; Hussain, J.; Wang, Y.F. Protein kinase OsSAPK8 functions as an essential activator of S-type anion channel OsSLAC1, which is nitrate-selective in rice. Planta 2016, 243, 489–500. [Google Scholar] [CrossRef]
- Liu, X.H.; Mak, M.; Babla, M.; Wang, F.F.; Chen, G.; Veljanoski, F.; Wang, G.; Shabala, S.; Zhou, M.X.; Chen, Z.H. Linking stomatal traits and expression of slow anion channel genes HvSLAH1 and HvSLAC1 with grain yield for increasing salinity tolerance in barley. Front. Plant Sci. 2014, 5, 634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, G.N.; Yao, F.Y.; Ren, H.M.; Sun, S.J.; Tan, Y.Q.; Zhang, Z.C.; Qiu, B.S.; Wang, Y.F. The S-Type Anion Channel ZmSLAC1 Plays Essential Roles in Stomatal Closure by Mediating Nitrate Efflux in Maize. Plant Cell Physiol. 2018, 59, 614–623. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.D.; Wang, L.; Chen, Q.; Qi, K.J.; Yin, H.; Cao, P.; Tang, C.; Wu, X.; Zhang, S.L.; Wang, P.; et al. PbrSLAH3 is a nitrate-selective anion channel which is modulated by calcium-dependent protein kinase 32 in pear. BMC Plant Biol. 2019, 19, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaborsky, M.; Maierhofer, T.; Olbrich, A.; Escalante-Perez, M.; Muller, H.M.; Simon, J.; Krol, E.; Cuin, T.A.; Fromm, J.; Ache, P.; et al. SLAH3-type anion channel expressed in poplar secretory epithelia operates in calcium kinase CPK-autonomous manner. New Phytol. 2016, 210, 922–933. [Google Scholar] [CrossRef] [Green Version]
- Kurusu, T.; Saito, K.; Horikoshi, S.; Hanamata, S.; Negi, J.; Yagi, C.; Kitahata, N.; Iba, K.; Kuchitsu, K. An S-Type Anion Channel SLAC1 Is Involved in Cryptogein-Induced Ion Fluxes and Modulates Hypersensitive Responses in Tobacco BY-2 Cells. PLoS ONE 2013, 8, e70623. [Google Scholar] [CrossRef]
- Chen, G.D.; Li, X.L.; Qiao, X.; Li, J.M.; Wang, L.; Kou, X.B.; Wu, X.; Wang, G.M.; Yin, H.; Wang, P.; et al. Genome-wide survey and expression analysis of the SLAC/SLAH gene family in pear (Pyrus bretschneideri) and other members of the Rosaceae. Genomics 2019, 111, 1097–1107. [Google Scholar] [CrossRef]
- Bouchet, A.S.; Laperche, A.; Bissuel-Belaygue, C.; Baron, C.; Morice, J.; Rousseau-Gueutin, M.; Dheu, J.E.; George, P.; Pinochet, X.; Foubert, T.; et al. Genetic basis of nitrogen use efficiency and yield stability across environments in winter rapeseed. BMC Genet. 2016, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chalhoub, B.; Denoeud, F.; Liu, S.Y.; Parkin, I.A.P.; Tang, H.B.; Wang, X.Y.; Chiquet, J.; Belcram, H.; Tong, C.B.; Samans, B.; et al. Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science 2014, 345, 950–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedrich, R.; Geiger, D. Biology of SLAC1-type anion channels—From nutrient uptake to stomatal closure. New Phytol. 2017, 216, 46–61. [Google Scholar] [CrossRef] [Green Version]
- Negi, J.; Matsuda, O.; Nagasawa, T.; Oba, Y.; Takahashi, H.; Kawai-Yamada, M.; Uchimiya, H.; Hashimoto, M.; Iba, K. CO2 regulator SLAC1 and its homologues are essential for anion homeostasis in plant cells. Nature 2008, 452, 483-U413. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y.; Hsu, P.K.; Tsay, Y.F. Uptake, allocation and signaling of nitrate. Trends Plant Sci. 2012, 17, 458–467. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, P.L.; Liu, Q.; Li, G.J.; Di, D.W.; Xia, G.M.; Kronzucker, H.J.; Fang, S.; Chu, J.F.; Shi, W.M. TaANR1-TaBG1 and TaWabi5-TaNRT2s/NARs Link ABA Metabolism and Nitrate Acquisition in Wheat Roots(1) ([OPEN]). Plant Physiol. 2020, 182, 1440–1453. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.L.; Liao, J.Y.; Yu, Y.; Song, H.X.; Rong, N.; Guan, C.Y.; Lepo, J.E.; Ismail, A.M.; Zhang, Z.H. Exogenous abscisic acid promotes the nitrogen use efficiency of Brassica napus by increasing nitrogen remobilization in the leaves. J. Plant Nutr. 2017, 40, 2540–2549. [Google Scholar] [CrossRef]
- Lee, T.H.; Tang, H.B.; Wang, X.Y.; Paterson, A.H. PGDD: A database of gene and genome duplication in plants. Nucleic Acids Res. 2013, 41, D1152–D1158. [Google Scholar] [CrossRef] [PubMed]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [Green Version]
- Drechsel, G.; Bergler, J.; Wippel, K.; Sauer, N.; Vogelmann, K.; Hoth, S. C-terminal armadillo repeats are essential and sufficient for association of the plant U-box armadillo E3 ubiquitin ligase SAUL1 with the plasma membrane. J. Exp. Bot. 2011, 62, 775–785. [Google Scholar] [CrossRef] [Green Version]
- Allender, C.J.; King, G.J. Origins of the amphiploid species Brassica napus L. investigated by chloroplast and nuclear molecular markers. BMC Plant Biol. 2010, 10, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Aufhammer, W.; Kubler, E.; Bury, M. Nitrogen Uptake and Nitrogen Residuals of Winter Oilseed Rape and Fallout Rape. J. Agron. Crop Sci. 1994, 172, 255–264. [Google Scholar] [CrossRef]
- Rathke, G.W.; Christen, O.; Diepenbrock, W. Effects of nitrogen source and rate on productivity and quality of winter oilseed rape (Brassica napus L.) grown in different crop rotations. Field Crop. Res. 2005, 94, 103–113. [Google Scholar] [CrossRef]
- Swarbreck, D.; Wilks, C.; Lamesch, P.; Berardini, T.Z.; Garcia-Hernandez, M.; Foerster, H.; Li, D.; Meyer, T.; Muller, R.; Ploetz, L.; et al. The Arabidopsis Information Resource (TAIR): Gene structure and function annotation. Nucleic Acids Res. 2008, 36, D1009–D1014. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.B.; Wang, X.Y.; Bowers, J.E.; Ming, R.; Alam, M.; Paterson, A.H. Unraveling ancient hexaploidy through multiply-aligned angiosperm gene maps. Genome Res. 2008, 18, 1944–1954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leblanc, A.; Renault, H.; Lecourt, J.; Etienne, P.; Deleu, C.; Le Deunff, E. Elongation changes of exploratory and root hair systems induced by aminocyclopropane carboxylic acid and aminoethoxyvinylglycine affect nitrate uptake and BnNrt2.1 and BnNrt1.1 transporter gene expression in oilseed rape. Plant Physiol. 2008, 146, 1928–1940. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Jin, J.P.; Guo, A.Y.; Zhang, H.; Luo, J.C.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.M.; Ma, H.L.; Yu, L.; Wang, X.; Zhao, J. Genome-Wide Survey and Expression Analysis of Amino Acid Transporter Gene Family in Rice (Oryza sativa L.). PLoS ONE 2012, 7, e49210. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.Y.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Maher, C.; Stein, L.; Ware, D. Evolution of Arabidopsis microRNA families through duplication events. Genome Res. 2006, 16, 510–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Qin, L.; Walk, T.C.; Han, P.P.; Chen, L.Y.; Zhang, S.; Li, Y.S.; Hu, X.J.; Xie, L.H.; Yang, Y.; Liu, J.P.; et al. Adaption of Roots to Nitrogen Deficiency Revealed by 3D Quantification and Proteomic Analysis. Plant Physiol. 2019, 179, 329–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Features | Protein Features | |||||||
|---|---|---|---|---|---|---|---|---|
| Putative Gene Name | Gene ID | Exon No. | Chr. Location | Protein Length (aa) | MW (KDa) | pI | GRAVY | TMD |
| BnSLAC1-1 | LOC106445577 | 3 | A06 (5554449..5557045) | 588 | 66.64 | 9.22 | −0.046 | 8 |
| BnSLAC1-2 | LOC111206299 | 3 | C05(7993671..7996097) | 586 | 66.18 | 9.3 | 0.03 | 8 |
| BnSLAH1-1 | LOC106365268 | 2 | A09(7429214..7430811) | 387 | 43.40 | 9.28 | 0.5 | 8 |
| BnSLAH1-2 | LOC106373608 | 2 | C04(33942406..33943955) | 387 | 43.81 | 9.29 | 0.455 | 8 |
| BnSLAH1-3 | LOC106373008 | 2 | A09 (9740564..9742068) | 381 | 42.99 | 9.27 | 0.566 | 8 |
| BnSLAH1-4 | LOC106400098 | 2 | C09 (12333910..12335636) | 381 | 43.04 | 9.11 | 0.551 | 8 |
| BnSLAH2-1 | LOC106409845 | 4 | C07 (54435207..54437674) | 498 | 56.26 | 9.3 | 0.231 | 10 |
| BnSLAH2-2 | LOC106359835 | 4 | A07(11635779..11638473) | 515 | 57.78 | 9.65 | 0.234 | 10 |
| BnSLAH2-3 | LOC106413896 | 4 | C03 (16311421..16314068) | 515 | 57.80 | 9.54 | 1.235 | 10 |
| BnSLAH2-4 | LOC106452714 | 3 | A01 (12909477..12912194) | 359 | 40.31 | 9.44 | 0.626 | 9 |
| BnSLAH2-5 | LOC106376145 | 3 | C01 (21092721..21095213) | 359 | 40.36 | 9.44 | 0.613 | 9 |
| BnSLAH3-1 | LOC106348290 | 5 | A06 (25034448..25039244) | 636 | 72.42 | 8.34 | −0.092 | 9 |
| BnSLAH3-2 | LOC106436465 | 5 | C07 (46161649..46166453) | 636 | 72.41 | 8.39 | −0.111 | 10 |
| BnSLAH3-3 | LOC106428459 | 5 | C09 (3799924..3805933) | 623 | 71.08 | 6.88 | −0.105 | 9 |
| BnSLAH3-4 | LOC106366019 | 5 | A01(11440762..11445373) | 623 | 71.07 | 7.07 | −0.11 | 9 |
| BnSLAH3-5 | LOC106415253 | 5 | A02 (32761829..32765883) | 640 | 72.59 | 8.95 | −0.07 | 9 |
| BnSLAH3-6 | LOC106378992 | 5 | C02 (66036653..66039643) | 636 | 72.24 | 8.88 | −0.074 | 9 |
| BnSLAH4-1 | LOC106417979 | 3 | A09 (9746804..9754357) | 476 | 54.27 | 9.26 | 0.308 | 10 |
| BnSLAH4-2 | LOC106397706 | 3 | C09 (12312761..12318196) | 470 | 53.58 | 9.22 | 0.32 | 10 |
| BnSLAH4-3 | LOC106412201 | 3 | C08(29119049..29124524) | 470 | 53.60 | 9.21 | 0.324 | 10 |
| BnSLAH4-4 | LOC106395319 | 2 | C04 (33847294..33848994) | 329 | 37.57 | 8.97 | 0.649 | 6 |
| BnSLAH4-5 | LOC106378846 | 6 | C01(36168496..36175425) | 533 | 60.56 | 6.04 | 0.19 | 8 |
| BnSLAH4-6 | LOC106378847 | 4 | C01(36145965..36151210) | 447 | 51.01 | 6.43 | 0.277 | 8 |
| Gene Name in A. thaliana | Gene Name in B. napus | Ka | Ks | Ka/Ks |
|---|---|---|---|---|
| AtSLAC1 | BnSLAC1-1 | 0.0616 | 0.4541 | 0.1356529 |
| AtSLAC1 | BnSLAC1-2 | 0.0634 | 0.4292 | 0.1477167 |
| AtSLAH1 | BnSLAH1-1 | 0.0867 | 0.4671 | 0.1856134 |
| AtSLAH1 | BnSLAH1-2 | 0.0958 | 0.4702 | 0.2037431 |
| AtSLAH1 | BnSLAH1-3 | 0.0929 | 0.4315 | 0.2152955 |
| AtSLAH1 | BnSLAH1-4 | 0.0955 | 0.4182 | 0.2283596 |
| AtSLAH2 | BnSLAH2-1 | 0.0567 | 0.3762 | 0.1507177 |
| AtSLAH2 | BnSLAH2-2 | 0.0686 | 0.4021 | 0.1706043 |
| AtSLAH2 | BnSLAH2-3 | 0.0706 | 0.4137 | 0.1706551 |
| AtSLAH2 | BnSLAH2-4 | 0.0762 | 0.4482 | 0.1700134 |
| AtSLAH2 | BnSLAH2-5 | 0.0741 | 0.46 | 0.161087 |
| AtSLAH3 | BnSLAH3-1 | 0.0694 | 0.4353 | 0.1594303 |
| AtSLAH3 | BnSLAH3-2 | 0.0686 | 0.3896 | 0.176078 |
| AtSLAH3 | BnSLAH3-3 | 0.0638 | 0.4419 | 0.1443766 |
| AtSLAH3 | BnSLAH3-4 | 0.0635 | 0.4699 | 0.1351351 |
| AtSLAH3 | BnSLAH3-5 | 0.1084 | 0.4767 | 0.2273967 |
| AtSLAH3 | BnSLAH3-6 | 0.1092 | 0.491 | 0.2224033 |
| AtSLAH4 | BnSLAH4-1 | 0.172 | 0.0677 | 2.5406204 |
| AtSLAH4 | BnSLAH4-2 | 0.1757 | 0.0679 | 2.5876289 |
| AtSLAH4 | BnSLAH4-3 | 0.1756 | 0.068 | 2.5823529 |
| AtSLAH4 | BnSLAH4-4 | 0.1595 | 0.0989 | 1.6127401 |
| AtSLAH4 | BnSLAH4-5 | 0.2183 | 0.1575 | 1.3860317 |
| AtSLAH4 | BnSLAH4-6 | 0.2183 | 0.1575 | 1.3860317 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nan, Y.; Xie, Y.; Atif, A.; Wang, X.; Zhang, Y.; Tian, H.; Gao, Y. Identification and Expression Analysis of SLAC/SLAH Gene Family in Brassica napus L. Int. J. Mol. Sci. 2021, 22, 4671. https://doi.org/10.3390/ijms22094671
Nan Y, Xie Y, Atif A, Wang X, Zhang Y, Tian H, Gao Y. Identification and Expression Analysis of SLAC/SLAH Gene Family in Brassica napus L. International Journal of Molecular Sciences. 2021; 22(9):4671. https://doi.org/10.3390/ijms22094671
Chicago/Turabian StyleNan, Yunyou, Yuyu Xie, Ayub Atif, Xiaojun Wang, Yanfeng Zhang, Hui Tian, and Yajun Gao. 2021. "Identification and Expression Analysis of SLAC/SLAH Gene Family in Brassica napus L." International Journal of Molecular Sciences 22, no. 9: 4671. https://doi.org/10.3390/ijms22094671
APA StyleNan, Y., Xie, Y., Atif, A., Wang, X., Zhang, Y., Tian, H., & Gao, Y. (2021). Identification and Expression Analysis of SLAC/SLAH Gene Family in Brassica napus L. International Journal of Molecular Sciences, 22(9), 4671. https://doi.org/10.3390/ijms22094671