
Developing a Riboswitch-Mediated Regulatory System for Metabolic Flux Control in Thermophilic Bacillus methanolicus



Abstract
:1. Introduction
2. Results
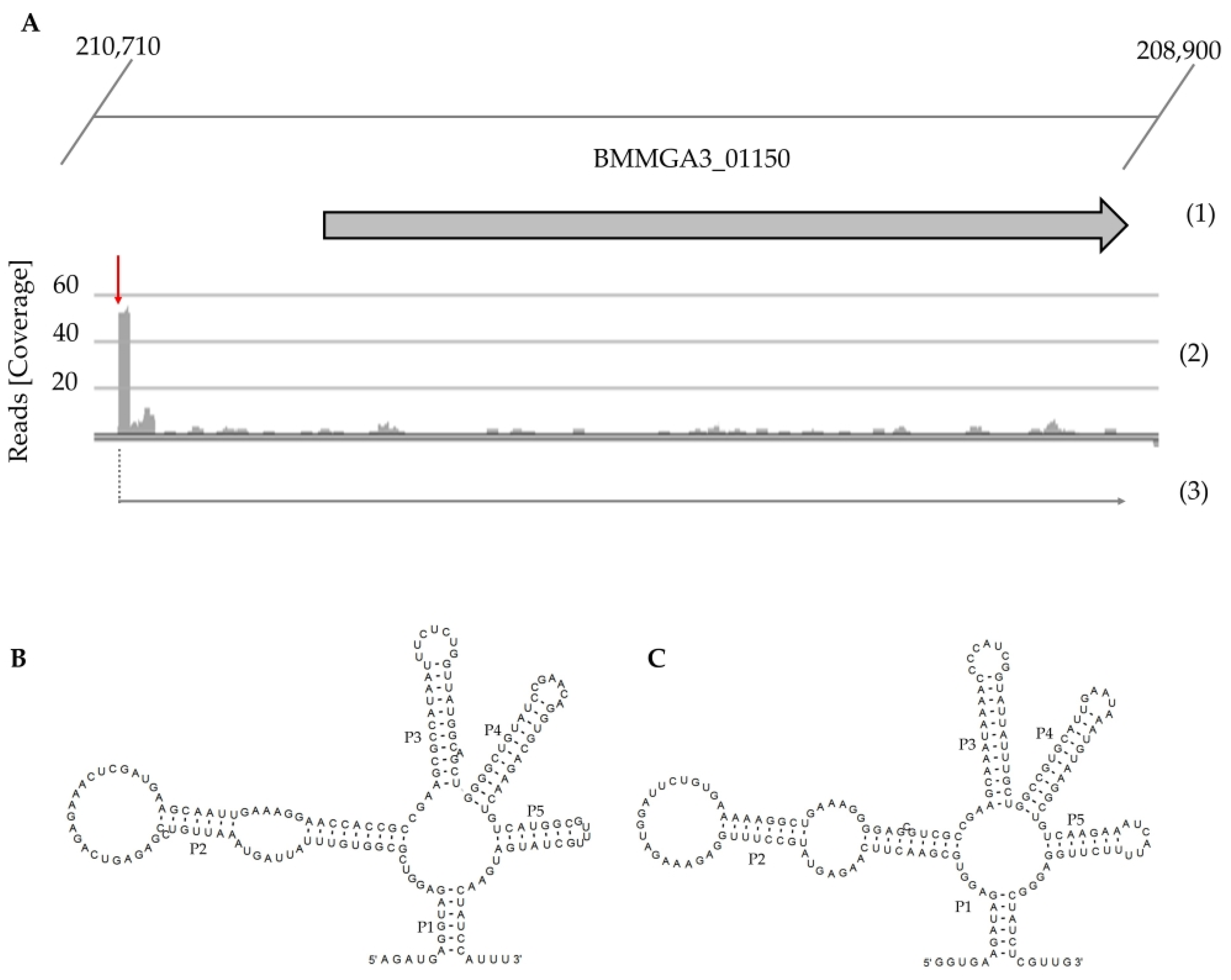
2.1. Transcriptome Analysis for Discovery of Novel Regulatory Elements
2.2. Detection of Putative Lysine Riboswitch in the Genome of B. methanolicus
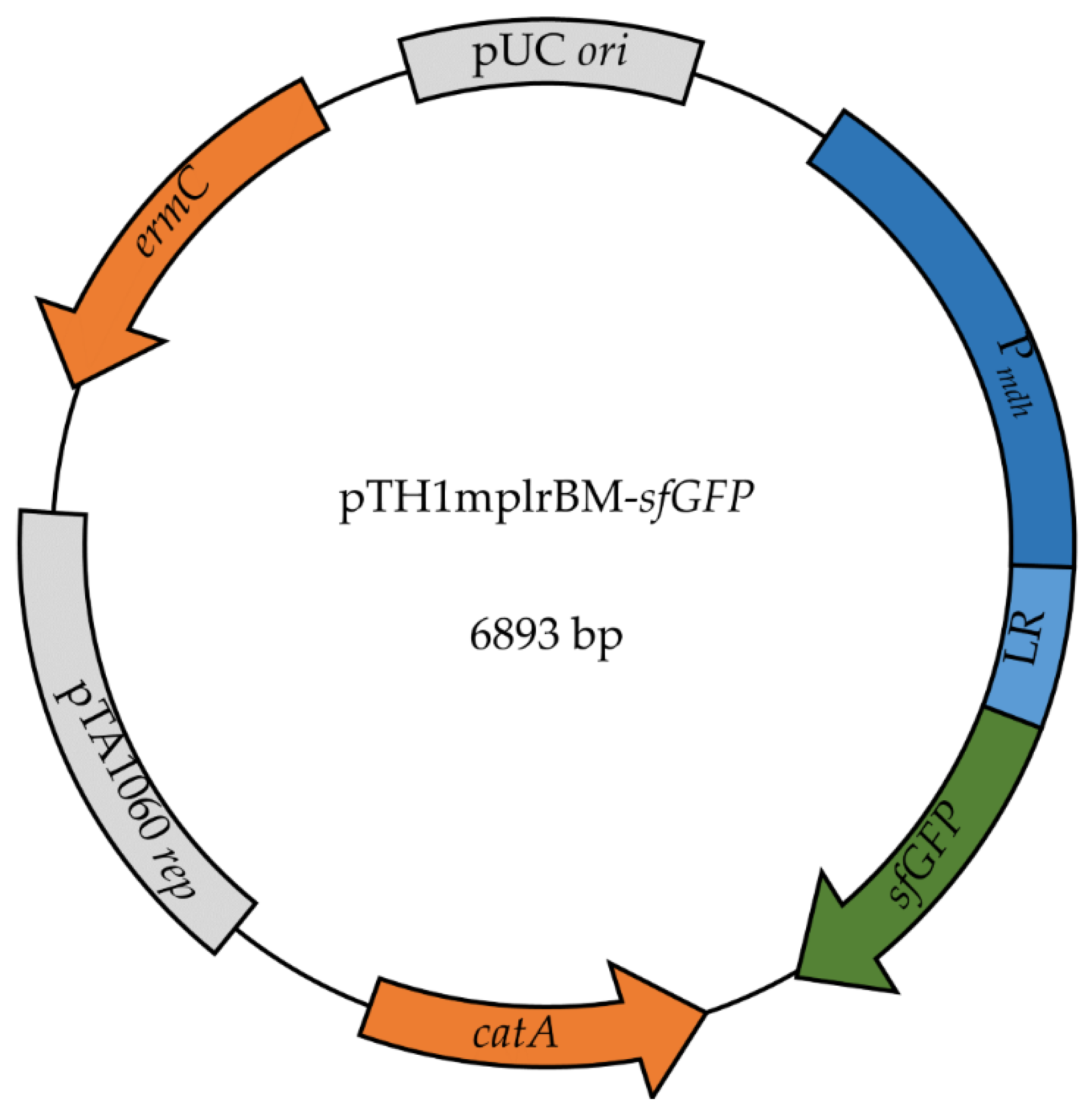
2.3. Construction of Model System to Characterize Riboswitches in B. methanolicus
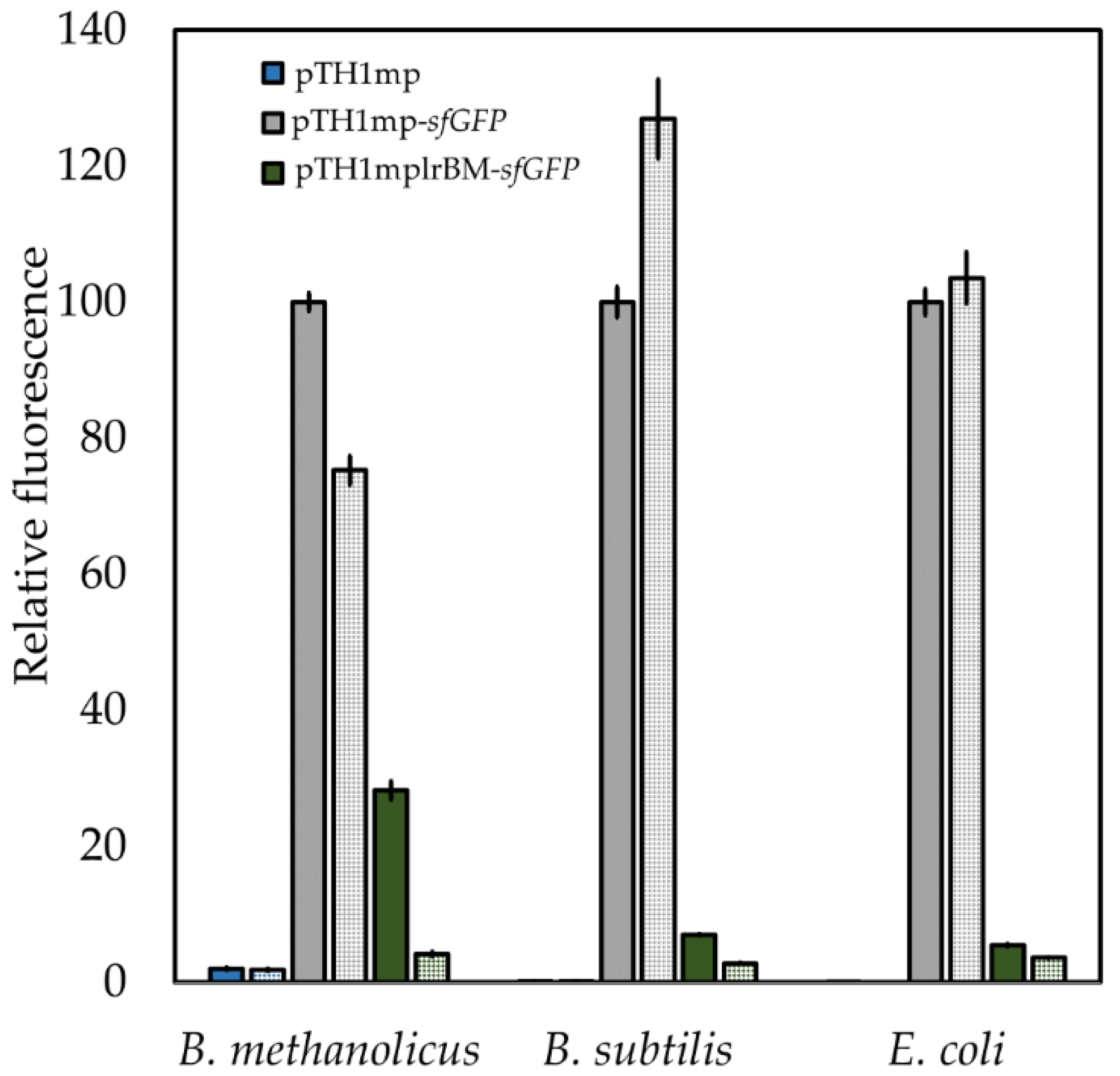
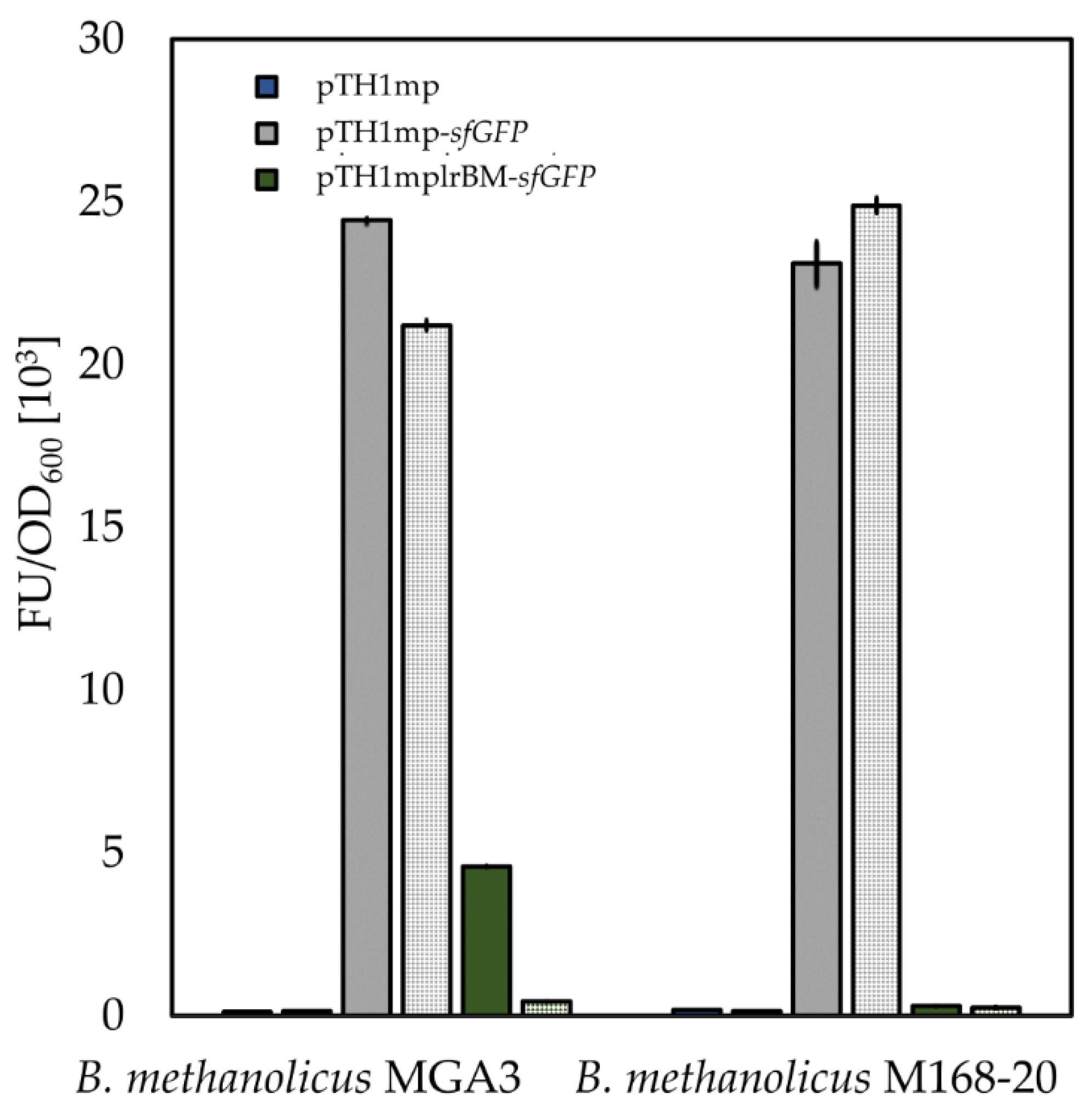
2.4. Characterization of Functionality of Putative Lysine Riboswitch from B. methanolicus
2.5. Sensitivity of B. methanolicus-Derived Lysine Riboswitch in Its Native Host to Intracellular l-lysine Concentration
2.6. Sensitivity of B. methanolicus-Derived Lysine Riboswitch in Its Native Host Extracellular l-lysine Concentration
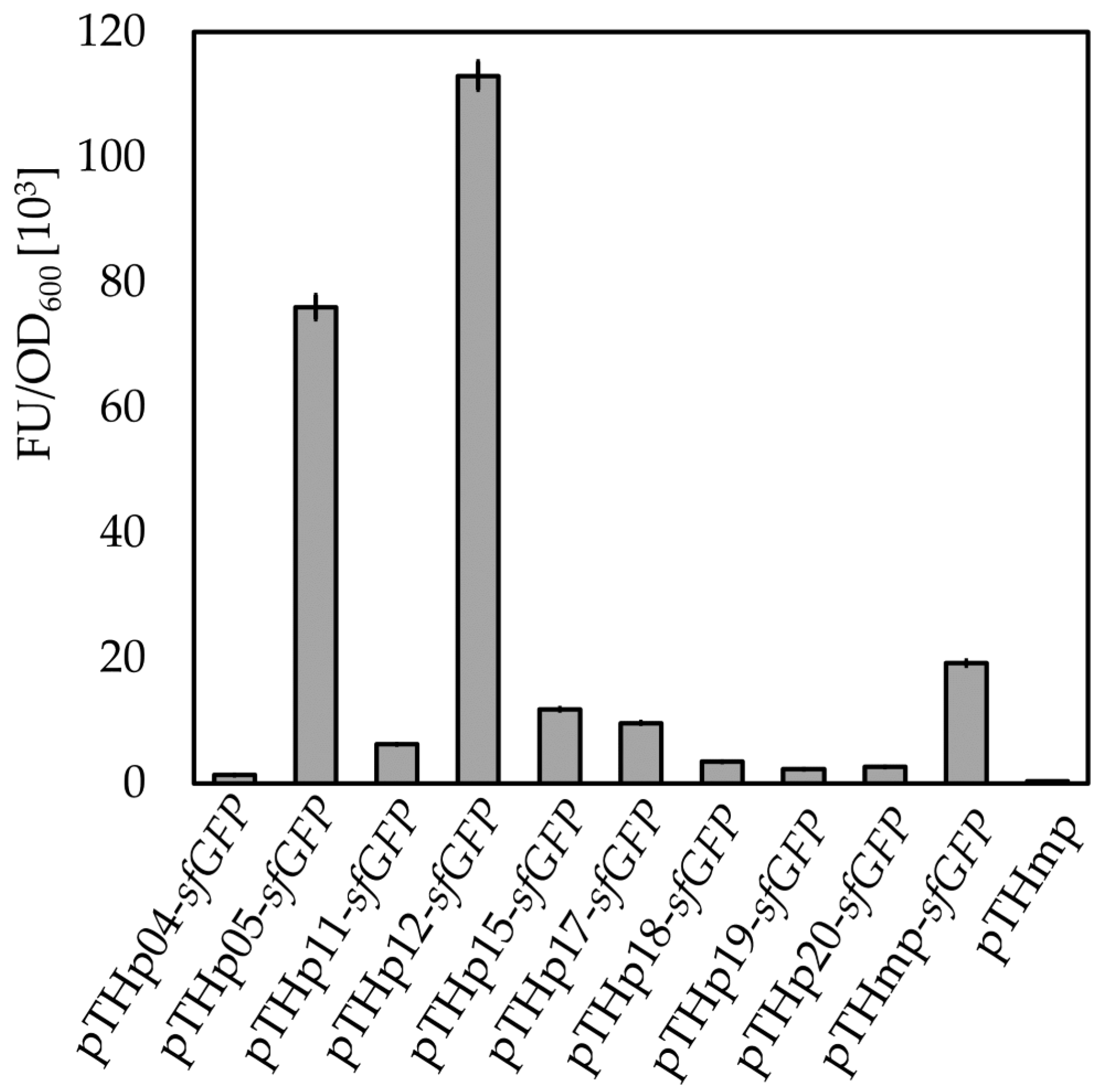
2.7. Characterization of Novel Promoters in B. methanolicus
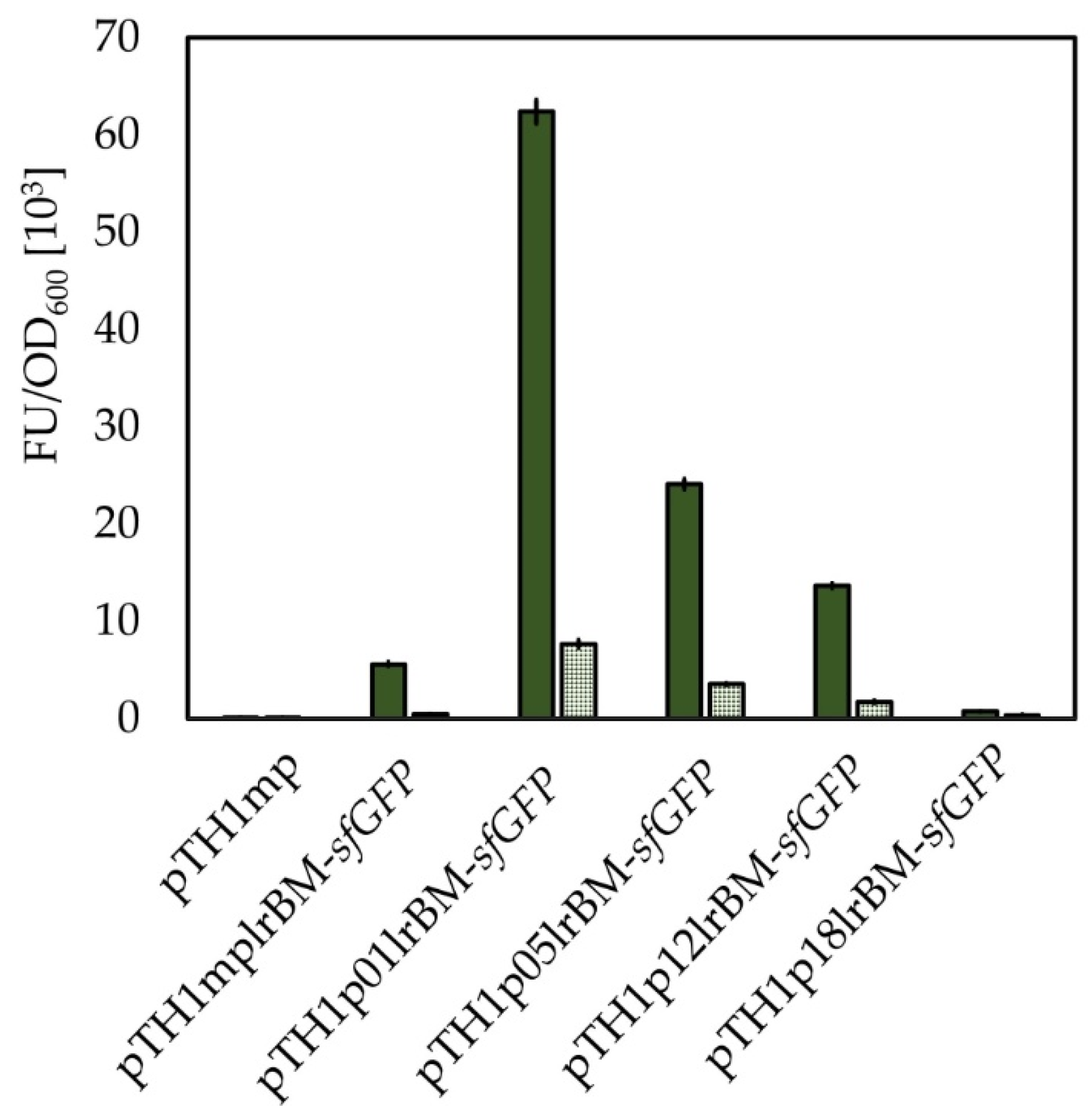
2.8. Consolidation of Identified Promoters and Lysine Riboswitch to Create Novel Tools for Gene Expression Regulation
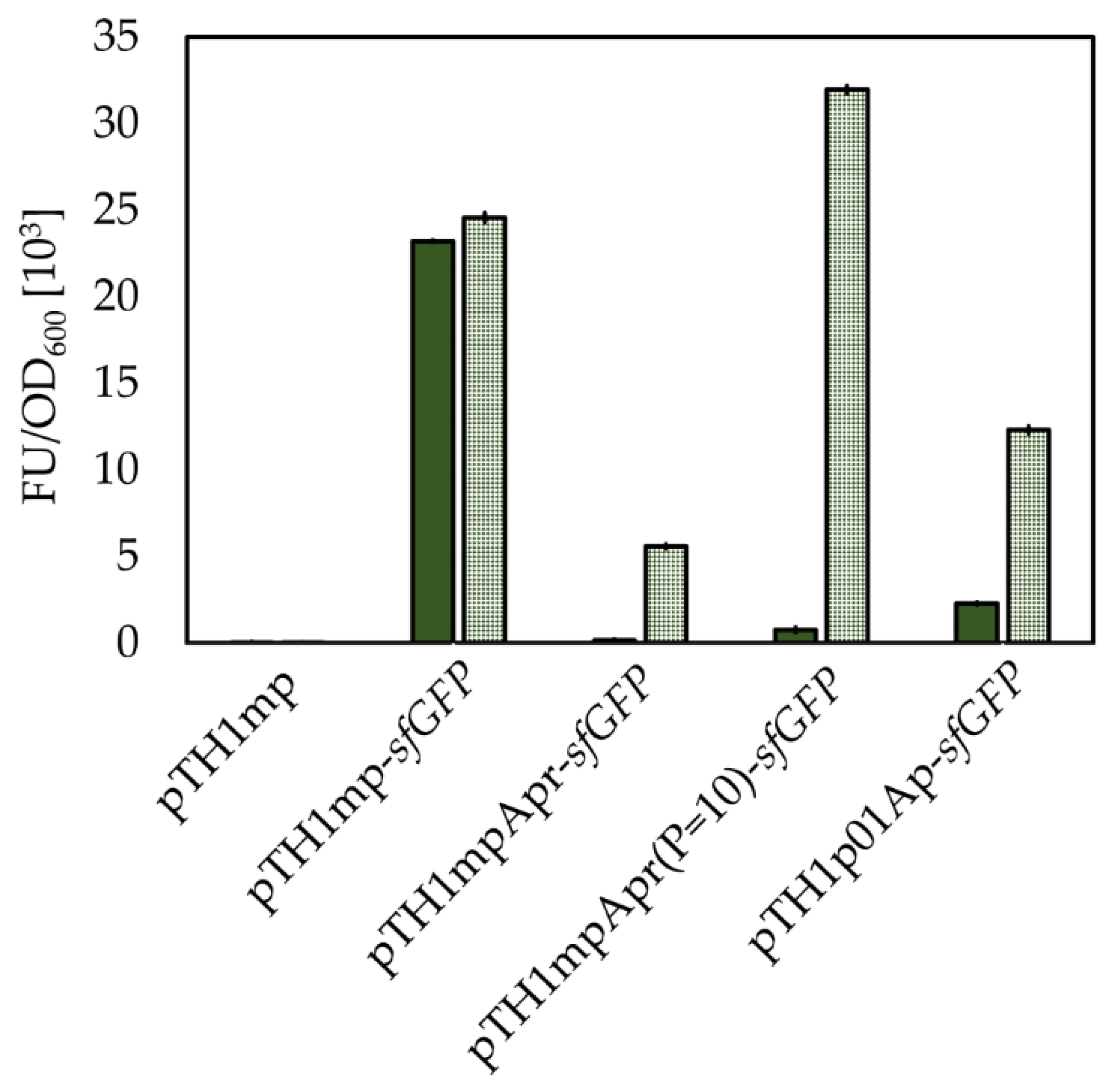
2.9. Transfer of Mesophilic Riboswitches to Thermophilic B. methanolicus
3. Discussion
4. Materials and Methods
4.1. Strains, Plasmids, and Primers
4.2. Molecular Cloning
4.3. Media and Conditions for Shake Flask Cultivations
4.4. Fluorescence Microplate Assay
4.5. Determination of Amino Acid Concentration
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pavlova, N.; Kaloudas, D.; Penchovsky, R. Riboswitch distribution, structure, and function in bacteria. Gene 2019, 708, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Xiu, Y.; Jang, S.; Jones, J.A.; Zill, N.A.; Linhardt, R.J.; Yuan, Q.; Jung, G.Y.; Koffas, M.A.G. Naringenin-responsive riboswitch-based fluorescent biosensor module for Escherichia coli co-cultures. Biotechnol. Bioeng. 2017, 114, 2235–2244. [Google Scholar] [CrossRef] [PubMed]
- Vitreschak, A.G. Riboswitches: The oldest mechanism for the regulation of gene expression? Trends Genet. 2004, 20, 44–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machtel, P.; Bąkowska-Żywicka, K.; Żywicki, M. Emerging applications of riboswitches—From antibacterial targets to molecular tools. J. Appl. Genet. 2016, 57, 531–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinumvayo, J.P.; Zhao, C.; Tuyishime, P. Recent advances and future trends of riboswitches: Attractive regulatory tools. World J. Microbiol. Biotechnol. 2018, 34, 171. [Google Scholar] [CrossRef] [PubMed]
- Blount, K.F.; Wang, J.X.; Lim, J.; Sudarsan, N.; Breaker, R.R. Antibacterial lysine analogs that target lysine riboswitches. Nat. Chem. Biol. 2006, 3, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Grundy, F.J.; Lehman, S.C.; Henkin, T.M. The L box regulon: Lysine sensing by leader RNAs of bacterial lysine biosynthesis genes. Proc. Natl. Acad. Sci. USA 2003, 100, 12057–12062. [Google Scholar] [CrossRef] [Green Version]
- Sudarsan, N.; Wickiser, J.K.; Nakamura, S.; Ebert, M.S.; Breaker, R.R. An mRNA structure in bacteria that controls gene expression by binding lysine. Genes Dev. 2003, 17, 2688–2697. [Google Scholar] [CrossRef] [Green Version]
- Caron, M.-P.; Bastet, L.; Lussier, A.; Simoneau-Roy, M.; Massé, E.; Lafontaine, D.A. Dual-acting riboswitch control of translation initiation and mRNA decay. Proc. Natl. Acad. Sci. USA 2012, 109, E3444–E3453. [Google Scholar] [CrossRef] [Green Version]
- Mandal, M.; Breaker, R.R. Adenine riboswitches and gene activation by disruption of a transcription terminator. Nat. Struct. Mol. Biol. 2003, 11, 29–35. [Google Scholar] [CrossRef]
- Zhou, L.; Ren, J.; Li, Z.; Nie, J.; Wang, C.; Zeng, A.-P. Characterization and engineering of a Clostridium glycine rboswitch and its use to control a novel metabolic pathway for 5-aminolevulinic acid production in Escherichia coli. ACS Synth. Biol. 2019, 8, 2327–2335. [Google Scholar] [CrossRef]
- Zhou, L.-B.; Zeng, A.-P. Exploring lysine riboswitch for metabolic flux control and improvement of l-lysine synthesis in Corynebacterium glutamicum. ACS Synth. Biol. 2014, 4, 729–734. [Google Scholar] [CrossRef] [PubMed]
- Boumezbeur, A.-H.; Bruer, M.; Stoecklin, G.; Mack, M. Rational engineering of transcriptional riboswitches leads to enhanced metabolite levels in Bacillus subtilis. Metab. Eng. 2020, 61, 58–68. [Google Scholar] [CrossRef]
- Zhou, L.-B.; Zeng, A.-P. Engineering a lysine-ON riboswitch for metabolic control of lysine production in Corynebacterium glutamicum. ACS Synth. Biol. 2015, 4, 1335–1340. [Google Scholar] [CrossRef]
- Cai, Y.; Xia, M.; Dong, H.; Qian, Y.; Zhang, T.; Zhu, B.; Wu, J.; Zhang, D. Engineering a vitamin B12 high-throughput screening system by riboswitch sensor in Sinorhizobium meliloti. BMC Biotechnol. 2018, 18, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, S.; Jang, S.; Im, D.-K.; Kang, T.J.; Oh, M.-K.; Jung, G.Y. Artificial caprolactam-specific riboswitch as an intracellular metabolite sensor. ACS Synth. Biol. 2019, 8, 1276–1283. [Google Scholar] [CrossRef]
- Pham, H.L.; Wong, A.; Chua, N.; Teo, W.S.; Yew, W.S.; Chang, M.W. Engineering a riboswitch-based genetic platform for the self-directed evolution of acid-tolerant phenotypes. Nat. Commun. 2017, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Zhang, L.; Barton, K.W.; Zhao, H. Systematic identification of a panel of strong constitutive promoters from Streptomyces albus. ACS Synth. Biol. 2015, 4, 1001–1010. [Google Scholar] [CrossRef] [PubMed]
- Siegl, T.; Tokovenko, B.; Myronovskyi, M.; Luzhetskyy, A. Design, construction and characterisation of a synthetic promoter library for fine-tuned gene expression in actinomycetes. Metab. Eng. 2013, 19, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Seghezzi, N.; Amar, P.; Koebmann, B.; Jensen, P.R.; Virolle, M.-J. The construction of a library of synthetic promoters revealed some specific features of strong Streptomyces promoters. Appl. Microbiol. Biotechnol. 2011, 90, 615–623. [Google Scholar] [CrossRef]
- Liu, D.; Mao, Z.; Guo, J.; Wei, L.; Ma, H.; Tang, Y.-J.; Chen, T.; Wang, Z.; Zhao, X. Construction, model-based analysis, and characterization of a promoter library for fine-tuned gene expression in Bacillus subtilis. ACS Synth. Biol. 2018, 7, 1785–1797. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, J.; Li, X.; Yin, S.; Wang, W.; Yang, K. Genome-wide identification and evaluation of constitutive promoters in streptomycetes. Microb. Cell Factories 2015, 14, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, S.; Ding, R.; Chen, J.; Du, G.; Li, H.; Zhou, J. Obtaining a panel of cascade promoter-5′-UTR complexes in Escherichia coli. ACS Synth. Biol. 2017, 6, 1065–1075. [Google Scholar] [CrossRef]
- Babski, J.; Haas, K.A.; Näther-Schindler, D.; Pfeiffer, F.; Förstner, K.U.; Hammelmann, M.; Hilker, R.; Becker, A.; Sharma, C.M.; Marchfelder, A.; et al. Genome-wide identification of transcriptional start sites in the haloarchaeon Haloferax volcanii based on differential RNA-Seq (dRNA-Seq). BMC Genom. 2016, 17, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Shen, W.; Huang, J.; Li, R.; Xiao, Y.; Wei, H.; Chou, Y.-C.; Zhang, M.; Himmel, M.E.; Chen, S.; et al. Prediction and characterization of promoters and ribosomal binding sites of Zymomonas mobilis in system biology era. Biotechnol. Biofuels 2019, 12, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Yang, H.; Zheng, J.; Ye, Y.; Pan, L. Identification of strong promoters based on the transcriptome of Bacillus licheniformis. Biotechnol. Lett. 2017, 39, 873–881. [Google Scholar] [CrossRef] [PubMed]
- Brautaset, T.; Jakobsen, Ø.M.; Josefsen, K.D.; Flickinger, M.C.; Ellingsen, T.E. Bacillus methanolicus: A candidate for industrial production of amino acids from methanol at 50 °C. Appl. Microbiol. Biotechnol. 2007, 74, 22–34. [Google Scholar] [CrossRef]
- Heggeset, T.M.B.; Krog, A.; Balzer, S.; Wentzel, A.; Ellingsen, T.E.; Brautaset, T. Genome sequence of thermotolerant Bacillus methanolicus: Features and regulation related to methylotrophy and production of l-lysine and l-glutamate from methanol. Appl. Environ. Microbiol. 2012, 78, 5170–5181. [Google Scholar] [CrossRef] [Green Version]
- Brautaset, T.; Williams, M.D.; Dillingham, R.D.; Kaufmann, C.; Bennaars, A.; Crabbe, E.; Flickinger, M.C. Role of the Bacillus methanolicus citrate synthase II gene, citY, in regulating the secretion of glutamate in l-lysine-secreting mutants. Appl. Environ. Microbiol. 2003, 69, 3986–3995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wendisch, V.F. Metabolic engineering advances and prospects for amino acid production. Metab. Eng. 2020, 58, 17–34. [Google Scholar] [CrossRef]
- Ochsner, A.M.; Müller, J.E.; Mora, C.A.; Vorholt, J.A. In vitro activation of NAD-dependent alcohol dehydrogenases by Nudix hydrolases is more widespread than assumed. FEBS Lett. 2014, 588, 2993–2999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irla, M.; Neshat, A.; Brautaset, T.; Rückert, C.; Kalinowski, J.; Wendisch, V.F. Transcriptome analysis of thermophilic methylotrophic Bacillus methanolicus MGA3 using RNA-sequencing provides detailed insights into its previously uncharted transcriptional landscape. BMC Genom. 2015, 16, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, J.E.N.; Litsanov, B.; Bortfeld-Miller, M.; Trachsel, C.; Grossmann, J.; Brautaset, T.; Vorholt, J.A. Proteomic analysis of the thermophilic methylotroph Bacillus methanolicus MGA. Proteomics 2014, 14, 725–737. [Google Scholar] [CrossRef] [PubMed]
- Carnicer, M.; Vieira, G.; Brautaset, T.; Portais, J.-C.; Heux, S. Quantitative metabolomics of the thermophilic methylotroph Bacillus methanolicus. Microb. Cell Factories 2016, 15, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delépine, B.; Gil López, M.; Carnicer, M.; Vicente, C.M.; Wendisch, V.F.; Heux, S. Charting the metabolic landscape of the facultative methylotroph Bacillus methanolicus. mSystems 2020, 5, e00745-20. [Google Scholar] [CrossRef]
- Irla, M.; Heggeset, T.M.B.; Nærdal, I.; Paul, L.; Haugen, T.; Le, S.B.; Brautaset, T.; Wendisch, V.F. Genome-based genetic tool development for Bacillus methanolicus: Theta- and rolling circle-replicating plasmids for inducible gene expression and application to methanol-based cadaverine production. Front. Microbiol. 2016, 7, 1481. [Google Scholar] [CrossRef] [Green Version]
- Rodionov, D.A.; Vitreschak, A.G.; Mironov, A.A.; Gelfand, M.S. Regulation of lysine biosynthesis and transport genes in bacteria: Yet another RNA riboswitch? Nucleic Acids Res. 2003, 31, 6748–6757. [Google Scholar] [CrossRef] [Green Version]
- Hilker, R.; Stadermann, K.B.; Doppmeier, D.; Kalinowski, J.; Stoye, J.; Straube, J.; Winnebald, J.; Goesmann, A. ReadXplorer—visualization and analysis of mapped sequences. Bioinformatics 2014, 30, 2247–2254. [Google Scholar] [CrossRef] [Green Version]
- Griffiths-Jones, S.; Bateman, A.; Marshall, M.; Khanna, A.; Eddy, S.R. Rfam: An RNA family database. Nucleic Acids Res. 2003, 31, 439–441. [Google Scholar] [CrossRef] [Green Version]
- Sweeney, B.A.; Hoksza, D.; Nawrocki, E.P.; Ribas, C.E.; Madeira, F.; Cannone, J.J.; Gutell, R.R.; Maddala, A.; Meade, C.; Williams, L.D.; et al. R2DT: Computational framework for template-based RNA secondary structure visualisation across non-coding RNA types. bioRxiv 2020. [Google Scholar] [CrossRef]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, T.J.; Eddy, S.R. nhmmer: DNA homology search with profile HMMs. Bioinformatics 2013, 29, 2487–2489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brautaset, T.; Jakobsen, Ø.M.; Degnes, K.F.; Netzer, R.; Naerdal, I.; Krog, A.; Dillingham, R.; Flickinger, M.C.; Ellingsen, T.E. Bacillus methanolicus pyruvate carboxylase and homoserine dehydrogenase I and II and their roles for l-lysine production from methanol at 50 °C. Appl. Microbiol. Biotechnol. 2010, 87, 951–964. [Google Scholar] [CrossRef]
- Nærdal, I.; Netzer, R.; Ellingsen, T.E.; Brautaset, T. Analysis and manipulation of aspartate pathway genes for l-lysine overproduction from methanol by Bacillus methanolicus. Appl. Environ. Microbiol. 2011, 77, 6020–6026. [Google Scholar] [CrossRef] [Green Version]
- Nærdal, I.; Pfeifenschneider, J.; Brautaset, T.; Wendisch, V.F. Methanol-based cadaverine production by genetically engineered Bacillus methanolicus strains. Microb. Biotechnol. 2015, 8, 342–350. [Google Scholar] [CrossRef]
- Brautaset, T.; Øyvind, M.J.M.; Flickinger, M.C.; Valla, S.; Ellingsen, T.E. Plasmid-dependent methylotrophy in thermotolerant Bacillus methanolicus. J. Bacteriol. 2004, 186, 1229–1238. [Google Scholar] [CrossRef] [Green Version]
- Priyakumar, U.D.; MacKerell, A.D. Role of the adenine ligand on the stabilization of the secondary and tertiary interactions in the adenine riboswitch. J. Mol. Biol. 2010, 396, 1422–1438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nærdal, I.; Netzer, R.; Irla, M.; Krog, A.; Heggeset, T.M.B.; Wendisch, V.F.; Brautaset, T. l-Lysine production by Bacillus methanolicus: Genome-based mutational analysis and l-lysine secretion engineering. J. Biotechnol. 2017, 244, 25–33. [Google Scholar] [CrossRef]
- Tempest, D.W.; Meers, J.L.; Brown, C.M. Influence of environment on the content and composition of microbial free amino acid pools. J. Gen. Microbiol. 1970, 64, 171–185. [Google Scholar] [CrossRef] [Green Version]
- Arfman, N.; Watling, E.M.; Clement, W.; Van Oosterwijk, R.J.; De Vries, G.E.; Harder, W.; Attwood, M.M.; Dijkhuizen, L. Methanol metabolism in thermotolerant methylotrophic Bacillus strains involving a novel catabolic NAD-dependent methanol dehydrogenase as a key enzyme. Arch. Microbiol. 1989, 152, 280–288. [Google Scholar] [CrossRef] [Green Version]
- Yasueda, H.; Kawahara, Y.; Sugimoto, S.-I. Bacillus subtilis yckG and yckF encode two key enzymes of the ribulose monophosphate pathway used by methylotrophs, and yckH is required for their expression. J. Bacteriol. 1999, 181, 7154–7160. [Google Scholar] [CrossRef] [Green Version]
- Schendel, F.J.; Bremmon, C.E.; Flickinger, M.C.; Guettler, M.; Hanson, R.S. l-Lysine production at 50 degrees C by mutants of a newly isolated and characterized methylotrophic Bacillus sp. Appl. Environ. Microbiol. 1990, 56, 963–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frieda, K.L.; Block, S.M. Direct observation of cotranscriptional folding in an adenine riboswitch. Science 2012, 338, 397–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcano-Velázquez, J.G.; Batey, R.T. Structure-guided mutational analysis of gene regulation by the Bacillus subtilis pbuE adenine-responsive riboswitch in a cellular context. J. Biol. Chem. 2015, 290, 4464–4475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcano-Velazquez, J.G.; Lo, J.; Nag, A.; Maness, P.-C.; Chou, K.J.; Marcano, J. Developing riboswitch-mediated gene regulatory controls in thermophilic bacteria. ACS Synth. Biol. 2019, 8, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Irla, M.; Drejer, E.B.; Brautaset, T.; Hakvåg, S. Establishment of a functional system for recombinant production of secreted proteins at 50 °C in the thermophilic Bacillus methanolicus. Microb. Cell Factories 2020, 19, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Saida, F.; Uzan, M.; Odaert, B.; Bontems, F. Expression of highly toxic genes in E. coli: Special strategies and genetic tools. Curr. Protein Pept. Sci. 2006, 7, 47–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowers, L.M.; LaPoint, K.; Anthony, L.; Pluciennik, A.; Filutowicz, M. Bacterial expression system with tightly regulated gene expression and plasmid copy number. Gene 2004, 340, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Mardanov, A.V.; Strakhova, T.S.; Smagin, V.A.; Ravin, N.V. Tightly regulated, high-level expression from controlled copy number vectors based on the replicon of temperate phage N. Gene 2007, 395, 15–21. [Google Scholar] [CrossRef]
- Sletta, H.; Nedal, A.; Aune, T.E.V.; Hellebust, H.; Hakvåg, S.; Aune, R.; Ellingsen, T.E.; Valla, S.; Brautaset, T. Broad-host-range plasmid pJB658 can be used for industrial-level production of a secreted host-toxic single-chain antibody fragment in Escherichia coli. Appl. Environ. Microbiol. 2004, 70, 7033–7039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pédelacq, J.-D.; Cabantous, S.; Tran, T.; Terwilliger, T.C.; Waldo, G.S. Engineering and characterization of a superfolder green fluorescent protein. Nat. Biotechnol. 2005, 24, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Green, R.; Rogers, E.J. Transformation of chemically competent E. coli. Methods Enzymol. 2013, 529, 329–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Eikmanns, B.J.; Thum-Schmitz, N.; Eggeling, L.; Lüdtke, K.-U.; Sahm, H. Nucleotide sequence, expression and transcriptional analysis of the Corynebacterium glutamicum gltA gene encoding citrate synthase. Microbiology 1994, 140, 1817–1828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, D.G.; Young, L.; Chuang, R.; Venter, J.C.; Hutchison, C.A., III; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods. 2009, 6, 343–345. [Google Scholar] [CrossRef] [PubMed]
- Jakobsen, Ø.M.; Benichou, A.; Flickinger, M.C.; Valla, S.; Ellingsen, T.E.; Brautaset, T. Upregulated transcription of plasmid and chromosomal ribulose monophosphate pathway genes is critical for methanol assimilation rate and methanol tolerance in the methylotrophic bacterium Bacillus methanolicus. J. Bacteriol. 2006, 188, 3063–3072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertani, G. Studies on lysogenesis. I. The mode of phage liberation by lysogenic Escherichia coli. J. Bacteriol. 1951, 62, 293–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, T.; Poetter, M.; Pfeffer, J.C.; Kroutil, W.; Skerra, A.; Lerchner, A.; Tauber, K.C.; Sattler, J.H.; Schaffer, S. Oxidation and Amination of Secondary Alcohols. U.S. Patent Application 14/237,121, 16 October 2014. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Promoter Designation | Locus Tag | Encoded Protein | LogRPKM |
|---|---|---|---|
| p01 (Phps-phi) | BMMGA3_06845- BMMGA3_06840 | 3-Hexulose-6-phosphate synthase and 3-hexulose-6-phosphate isomerase | 12307.54 |
| p04 | BMMGA3_16050 | Putative sugar phosphate isomerase, RpiB | 3472.63 |
| p05 | BMMGA3_01940 | Glutamate synthase, large subunit, GltA, small subunit, GltB | 1779.97 |
| p11 | BMMGA3_14565 | NH3-dependent NAD+ synthetase, NadE | 1598.52 |
| p12 | BMMGA3_10275 | Dihydroxy-acid dehydratase, IlvD | 758.16 |
| p15 | BMMGA3_16310 | Pyridoxine kinase, PdxK | 231.45 |
| p17 | BMMGA3_11740 | Hypothetical protein | 160.00 |
| p18 | BMMGA3_02750 | d-Isomer specific 2-hydroxyacid dehydrogenase NAD-binding protein | 107.14 |
| p19 | BMMGA3_09580 | Putative membrane protein | 66.73 |
| p20 | BMMGA3_09845 | Hypothetical protein | 39.59 |
| Strain Name | Relevant Characteristics | Reference |
| E. coli DH5α | General cloning host, F-thi-1 endA1 hsdR17(r-,m-) supE44 _lacU169 (_80lacZ_M15) recA1 gyrA96 relA1 | Stratagene |
| E. coli MG1655 | Wild-type strain; F- λ- ilvG- rfb-50 rph-1 | ATCC 47076 |
| B. methanolicus MGA3 | Wild-type strain | ATCC 53907 |
| B. methanolicus M168-20 | 1st generation S-(2-aminoethyl) cysteine-resistant mutant of MGA3; l-lysine overproducer | [43] |
| B. subtilis 168 | Wild-type strain | ATCC 23857 |
| Plasmid Name | Relevant Characteristics | Reference |
| pTH1mp | CmR; derivative of pTH1mp-lysC for gene expression under control of the mdh promoter. The lysC gene was replaced with multiple cloning site. | [36] |
| sfGFP-pBAD | AmR; pBAD/His derivative for expression of sfGFP | Addgene # 54519 [61] |
| pTH1mp-sfGFP | CmR; pTH1mp derivative for expression of sfGFP from sfGFP-pBAD under control of the mdh promoter | This study |
| pTH1mplrBM-sfGFP | CmR; pTH1mp derivative for expression of sfGFP from sfGFP-pBAD under control of the mdh promoter and B. methanolicus-derived lysine riboswitch | This study |
| pTH1p04-sfGFP | CmR; pTH1mp derivative for expression of sfGFP from sfGFP-pBAD under control of the promoter p04 (Table 1) | This study |
| pTH1p05-sfGFP | CmR; pTH1mp derivative for expression of sfGFP from sfGFP-pBAD under control of the promoter p05 (Table 1) | This study |
| pTH1p11-sfGFP | CmR; pTH1mp derivative for expression of sfGFP from sfGFP-pBAD under control of the promoter p11 (Table 1) | This study |
| pTH1p12-sfGFP | CmR; pTH1mp derivative for expression of sfGFP from sfGFP-pBAD under control of the promoter p12 (Table 1) | This study |
| pTH1p15-sfGFP | CmR; pTH1mp derivative for expression of sfGFP from sfGFP-pBAD under control of the promoter p15 (Table 1) | This study |
| pTH1p17-sfGFP | CmR; pTH1mp derivative for expression of sfGFP from sfGFP-pBAD under control of the promoter p17 (Table 1) | This study |
| pTH1p18-sfGFP | CmR; pTH1mp derivative for expression of sfGFP from sfGFP-pBAD under control of the promoter p18 (Table 1) | This study |
| pTH1p19-sfGFP | CmR; pTH1mp derivative for expression of sfGFP from sfGFP-pBAD under control of the promoter p19 (Table 1) | This study |
| pTH1p20-sfGFP | CmR; pTH1mp derivative for expression of sfGFP from sfGFP-pBAD under control of the promoter p20 (Table 1) | This study |
| pTH1p01lrBM-sfGFP | CmR; pTH1mp derivative for expression of sfGFP from sfGFP-pBAD under control of the promoter p01 (Table 1) and B. methanolicus-derived lysine riboswitch | This study |
| pTH1p05lrBM-sfGFP | CmR; pTH1mp derivative for expression of sfGFP from sfGFP-pBAD under control of the promoter p05 (Table 1) and B. methanolicus-derived lysine riboswitch | This study |
| pTH1p12lrBM-sfGFP | CmR; pTH1mp derivative for expression of sfGFP from sfGFP-pBAD under control of the promoter p12 (Table 1) and B. methanolicus derived-lysine riboswitch | This study |
| pTH1p18lrBM-sfGFP | CmR; pTH1mp derivative for expression of sfGFP from sfGFP-pBAD under control of the promoter p18 (Table 1) and B. methanolicus-derived lysine riboswitch | This study |
| pTH1mpApr-sfGFP | CmR; pTH1mp derivative for expression of sfGFP from sfGFP-pBAD under control of the mdh promoter and B. subtilis-derived pbuE riboswitch | This study |
| pTH1mpApr(P1 = 10)-sfGFP | CmR; pTH1mp derivative for expression of sfGFP from sfGFP-pBAD under control of the mdh promoter and modified B. subtilis-derived pbuE riboswitch | This study |
| pTH1p01Apr-sfGFP | CmR; pTH1mp derivative for expression of sfGFP from sfGFP-pBAD under control of the promoter p01 (Table 1) and B. subtilis-derived pbuE riboswitch | This study |
| Plasmid Name | Vector Backbone and Method of Linearization | Insert and Primers Used for Amplification |
|---|---|---|
| pTH1mp-sfGFP | pTH1mp digested with XbaI and AflIII | sfGFP gene PCR-amplified with sfGFP-pTH1mp_FW and sfGFP-pTH1mp_RW |
| pTH1mplrBM-sfGFP | pTHmp-sfGFP PCR-amplified with PSGF and PSGR | Lysine riboswitch derived from B. methanolicus PCR-amplified with LRIF and LRIR |
| pTH1p04-sfGFP | pTHmp-sfGFP PCR-amplified with PROM01 and PROM02 | Promoter p04 PCR-amplified with PROM09 and PROM10 |
| pTH1p05-sfGFP | pTHmp-sfGFP PCR-amplified with PROM01 and PROM02 | Promoter p05 PCR-amplified with PROM11 and PROM12 |
| pTH1p11-sfGFP | pTHmp-sfGFP PCR-amplified with PROM01 and PROM02 | Promoter p11 PCR-amplified with PROM23 and PROM24 |
| pTH1p12-sfGFP | pTHmp-sfGFP PCR-amplified with PROM01 and PROM02 | Promoter p12 PCR-amplified with PROM25 and PROM26 |
| pTH1p15-sfGFP | pTHmp-sfGFP PCR-amplified with PROM01 and PROM02 | Promoter p15 PCR-amplified with PROM31 and PROM32 |
| pTH1p17-sfGFP | pTHmp-sfGFP PCR-amplified with PROM01 and PROM02 | Promoter p17 PCR-amplified with PROM35 and PROM36 |
| pTH1p18-sfGFP | pTHmp-sfGFP PCR-amplified with PROM01 and PROM02 | Promoter p18 PCR-amplified with PROM37 and PROM38 |
| pTH1p19-sfGFP | pTHmp-sfGFP PCR-amplified with PROM01 and PROM02 | Promoter p19 PCR-amplified with PROM39 and PROM40 |
| pTH1p20-sfGFP | pTHmp-sfGFP PCR-amplified with PROM01 and PROM02 | Promoter p20 PCR-amplified with PROM41 and PROM42 |
| pTH1p01lrBM-sfGFP | pTHmplrBM-sfGFP PCR-amplified with LR11 and PROM02 | Promoter p01 PCR-amplified with PROM03 and LR12 |
| pTH1p05lrBM-sfGFP | pTHmplrBM-sfGFP PCR-amplified with LR11 and PROM02 | Promoter p05 PCR-amplified with PROM11 and LR20 |
| pTH1p12lrBM-sfGFP | pTHmplrBM-sfGFP PCR-amplified with LR11 and PROM02 | Promoter p12 PCR-amplified with PROM25 and LR14 |
| pTH1p18lrBM-sfGFP | pTHmplrBM-sfGFP PCR-amplified with LR11 and PROM02 | Promoter p18 PCR-amplified with PROM37 and LR15 |
| pTH1mpApr-sfGFP | pTH1mp-sfGFP PCR-amplified with PGF2 and PSGR | B. subtilis-derived pbuE riboswitch amplified with APR01 and APR02 |
| pTH1mpApr(P1 = 10)-sfGFP | pTH1mpApr-sfGFP PCR-amplified in site directed mutagenesis approach with APR05 and APR06 | |
| pTH1p01Apr-sfGFP | pTH1mp-sfGFP PCR-amplified with PROM01 and PROM02 | Promoter p01 PCR-amplified with PROM03 and APR03; B. subtilis-derived pbuE riboswitch amplified with APR04 and APR02 |
| Time (min) | Total Flow | %A | %B |
|---|---|---|---|
| 1.3 | 62.0 | 38.0 | |
| 5 | 1.3 | 62.0 | 38.0 |
| 12 | 1.3 | 43.0 | 57.0 |
| 14 | 1.3 | 24.0 | 76.0 |
| 15 | 1.3 | 43.0 | 57.0 |
| 18 | 1.3 | 620 | 38.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Irla, M.; Hakvåg, S.; Brautaset, T. Developing a Riboswitch-Mediated Regulatory System for Metabolic Flux Control in Thermophilic Bacillus methanolicus. Int. J. Mol. Sci. 2021, 22, 4686. https://doi.org/10.3390/ijms22094686
Irla M, Hakvåg S, Brautaset T. Developing a Riboswitch-Mediated Regulatory System for Metabolic Flux Control in Thermophilic Bacillus methanolicus. International Journal of Molecular Sciences. 2021; 22(9):4686. https://doi.org/10.3390/ijms22094686
Chicago/Turabian StyleIrla, Marta, Sigrid Hakvåg, and Trygve Brautaset. 2021. "Developing a Riboswitch-Mediated Regulatory System for Metabolic Flux Control in Thermophilic Bacillus methanolicus" International Journal of Molecular Sciences 22, no. 9: 4686. https://doi.org/10.3390/ijms22094686
APA StyleIrla, M., Hakvåg, S., & Brautaset, T. (2021). Developing a Riboswitch-Mediated Regulatory System for Metabolic Flux Control in Thermophilic Bacillus methanolicus. International Journal of Molecular Sciences, 22(9), 4686. https://doi.org/10.3390/ijms22094686