
Electrophysiological Properties of Endogenous Single Ca2+ Activated Cl− Channels Induced by Local Ca2+ Entry in HEK293

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
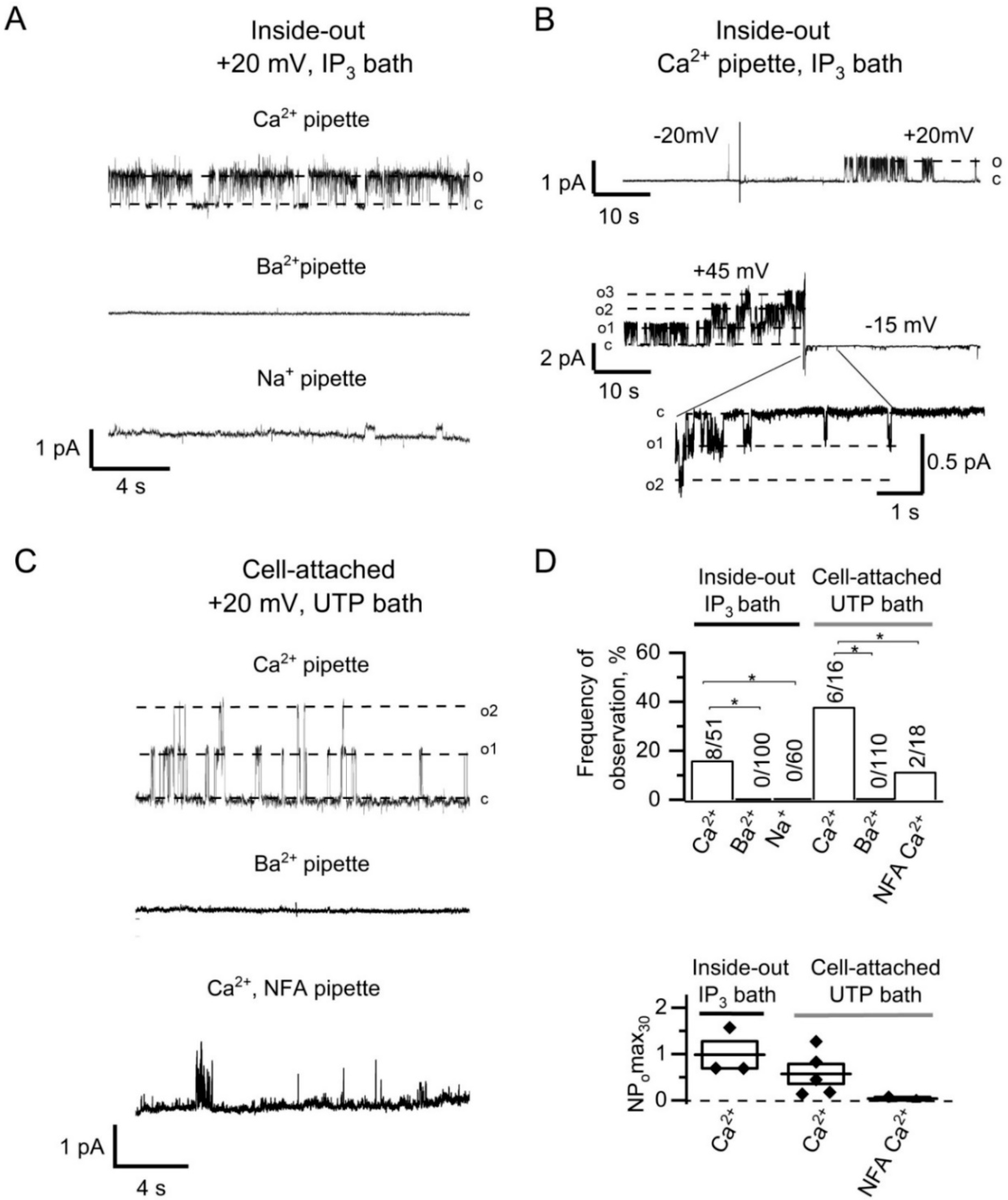
2.1. IP3-Induced Calcium Entry through TRPC1 Activates Outwardly Rectifying CaCCs in HEK293 Cells
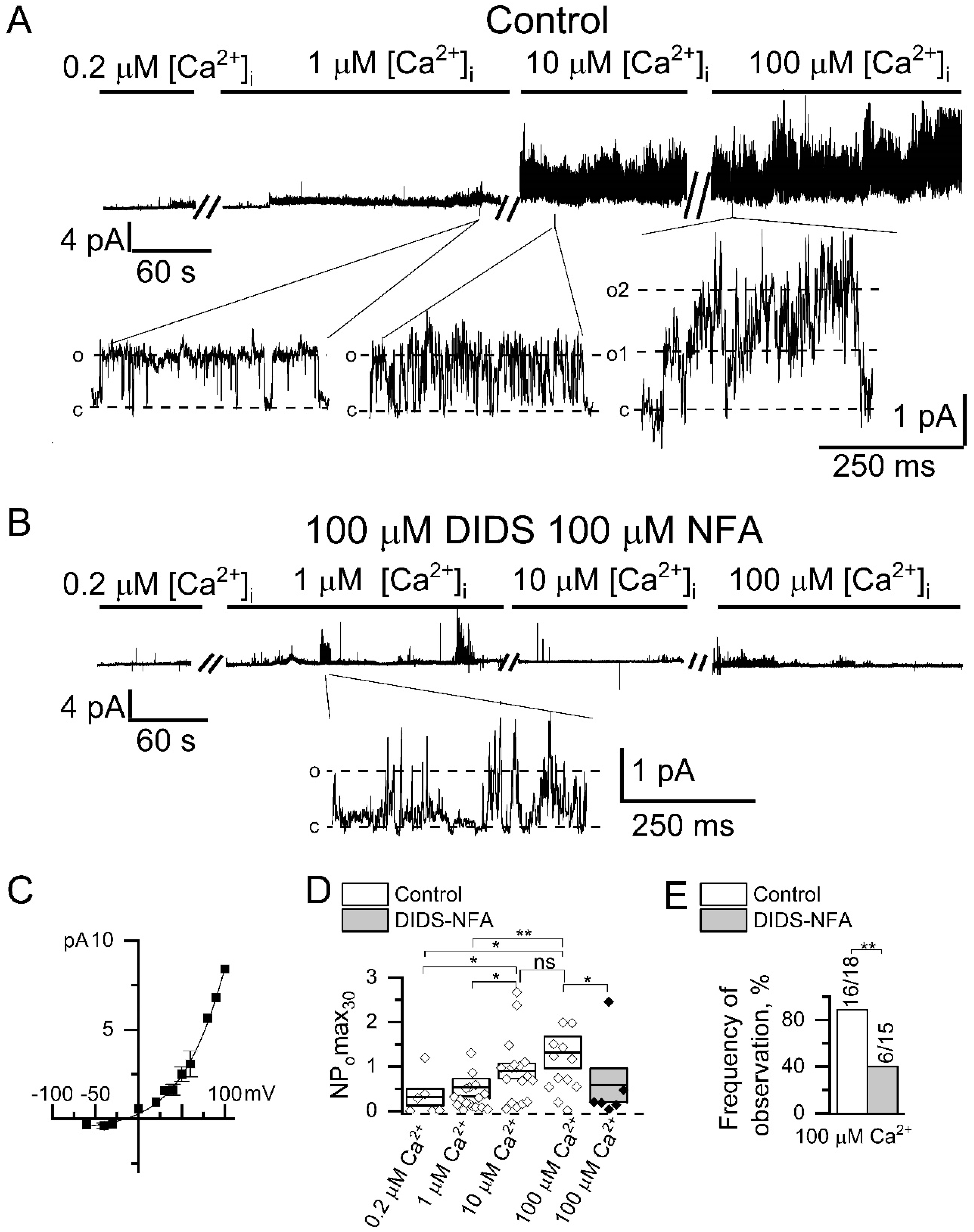
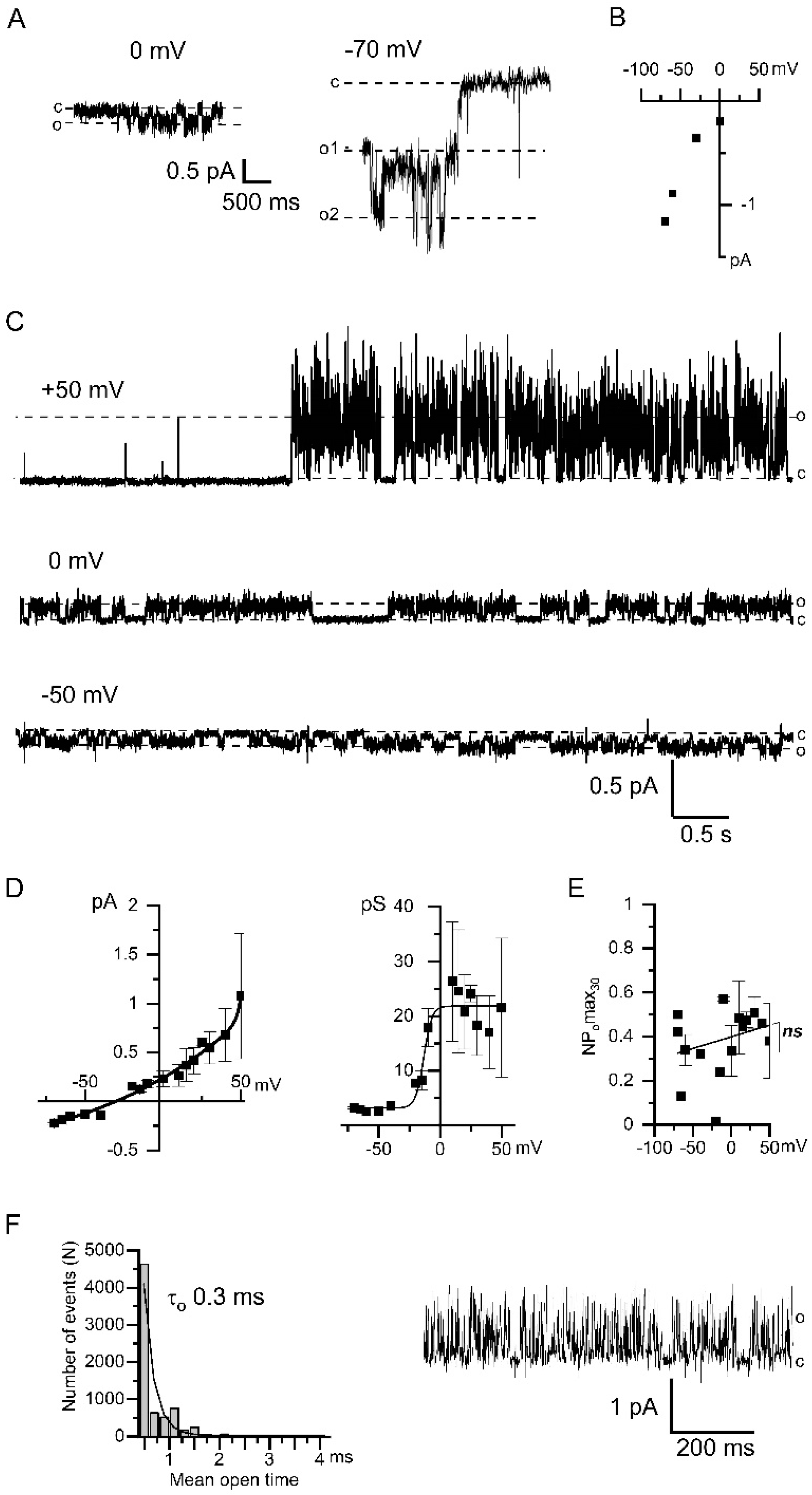
2.2. Direct Activation of Endogenous CaCCs in HEK293 by High [Ca2+]i
2.3. Functional Coupling and Close Arrangement of Endogenous TRPC1 Channels and CaCCs
3. Discussion
4. Materials and Methods
4.1. Cells
4.2. Electrophysiological Analysis
4.3. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barak, P.; Parekh, A.B. Signaling through Ca2+ Microdomains from Store-Operated CRAC Channels. Cold Spring Harb. Perspect. Biol. 2020, 12, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Parekh, A.B.; Putney, J.W. Store-Operated Calcium Channels. Physiol. Rev. 2005, 85, 757–810. [Google Scholar] [CrossRef] [Green Version]
- Gudlur, A.; Quintana, A.; Zhou, Y.; Hirve, N.; Mahapatra, S.; Hogan, P.G. STIM1 triggers a gating rearrangement at the extracellular mouth of the ORAI1 channel. Nat. Commun. 2014, 5, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saul, S.; Stanisz, H.; Backes, C.S.; Schwarz, E.C.; Hoth, M. How ORAI and TRP channels interfere with each other: Interaction models and examples from the immune system and the skin. Eur. J. Pharmacol. 2014, 739, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Ong, H.L.; Jang, S.-I.; Ambudkar, I.S. Distinct Contributions of Orai1 and TRPC1 to Agonist-Induced [Ca2+]i Signals Determine Specificity of Ca2+-Dependent Gene Expression. PLoS ONE 2012, 7, e47146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundivakkam, P.C.; Freichel, M.; Singh, V.; Yuan, J.P.; Vogel, S.M.; Flockerzi, V.; Malik, A.B.; Tiruppathi, C. The Ca2+ Sensor Stromal Interaction Molecule 1 (STIM1) Is Necessary and Sufficient for the Store-Operated Ca2+ Entry Function of Transient Receptor Potential Canonical (TRPC) 1 and 4 Channels in Endothelial Cells. Mol. Pharmacol. 2011, 81, 510–526. [Google Scholar] [CrossRef] [Green Version]
- Asanov, A.; Sampieri, A.; Moreno, C.; Pacheco, J.; Salgado, A.; Sherry, R.; Vaca, L. Combined single channel and single molecule detection identifies subunit composition of STIM1-activated transient receptor potential canonical (TRPC) channels. Cell Calcium 2015, 57, 1–13. [Google Scholar] [CrossRef]
- Cheng, K.T.; Liu, X.; Ong, H.L.; Swaim, W.; Ambudkar, I.S. Local Ca2+ Entry Via Orai1 Regulates Plasma Membrane Recruitment of TRPC1 and Controls Cytosolic Ca2+ Signals Required for Specific Cell Functions. PLoS Biol. 2011, 9, e1001025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.L.; Qian, Y.; Qiu, Q.Y.; Lan, X.J.; He, H.; Guan, Y.Y. Interaction between Cl− channels and CRAC-related Ca2+ signaling during T lymphocyte activation and proliferation1. Acta Pharmacol. Sin. 2006, 27, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Concepcion, A.R.; Vaeth, M.; Wagner, L.E.; Eckstein, M.; Hecht, L.; Yang, J.; Crottes, D.; Seidl, M.; Shin, H.P.; Weidinger, C.; et al. Store-operated Ca2+ entry regulates Ca2+-activated chloride channels and eccrine sweat gland function. J. Clin. Investig. 2016, 126, 4303–4318. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Drumm, B.T.; Earley, S.; Sung, T.S.; Koh, S.D.; Sanders, K.M. SOCE mediated by STIM and Orai is essential for pacemaker activity in the interstitial cells of Cajal in the gastrointestinal tract. Sci. Signal. 2018, 11, eaaq0918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Birnbaumer, L.; Singh, B.B. TRPC1 regulates calcium-activated chloride channels in salivary gland cells. J. Cell. Physiol. 2015, 230, 2848–2856. [Google Scholar] [CrossRef]
- Yamamura, A.; Yamamura, H.; Zeifman, A.; Yuan, J.X.-J. Activity of Ca2+-Activated Cl− Channels Contributes to Regulating Receptor- and Store-Operated Ca2+ Entry in Human Pulmonary Artery Smooth Muscle Cells. Pulm. Circ. 2011, 1, 269–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez, N.M.; Tamm, E.R.; Strauβ, O. Role of bestrophin-1 in store-operated calcium entry in retinal pigment epithelium. Pflügers Arch. Eur. J. Physiol. 2012, 465, 481–495. [Google Scholar] [CrossRef]
- Sharma, A.; Ramena, G.; Yin, Y.; Premkumar, L.; Elble, R.C. CLCA2 is a positive regulator of store-operated calcium entry and TMEM16A. PLoS ONE 2018, 13, e0196512. [Google Scholar] [CrossRef]
- Jha, A.; Chung, W.Y.; Vachel, L.; Maleth, J.; Lake, S.; Zhang, G.; Ahuja, M.; Muallem, S. Anoctamin 8 tethers endoplasmic reticulum and plasma membrane for assembly of Ca2+ signaling complexes at the ER/PM compartment. EMBO J. 2019, 38, e101452. [Google Scholar] [CrossRef]
- Cabrita, I.; Benedetto, R.; Fonseca, A.; Wanitchakool, P.; Sirianant, L.; Skryabin, B.V.; Schenk, L.K.; Pavenstädt, H.; Schreiber, R.; Kunzelmann, K. Differential effects of anoctamins on intracellular calcium signals. FASEB J. 2017, 31, 2123–2134. [Google Scholar] [CrossRef]
- Forrest, A.S.; Angermann, J.E.; Raghunathan, R.; Lachendro, C.; Greenwood, I.A.; Leblanc, N. Intricate interaction between store-operated calcium entry and calcium-activated chloride channels in pulmonary artery smooth muscle cells. In Membrane Receptors, Channels and Transporters in Pulmonary Circulation; Humana Press: Totowa, NJ, USA, 2010; pp. 31–55. [Google Scholar] [CrossRef]
- Sivilotti, L.G. What single-channel analysis tells us of the activation mechanism of ligand-gated channels: The case of the glycine receptor. J. Physiol. 2010, 588, 45–58. [Google Scholar] [CrossRef]
- Bugaj, V.; Alexeenko, V.; Zubov, A.; Glushankova, L.; Nikolaev, A.; Wang, Z.; Kaznacheyeva, E.; Bezprozvanny, I.; Mozhayeva, G.N. Functional Properties of Endogenous Receptor- and Store-operated Calcium Influx Channels in HEK293 Cells. J. Biol. Chem. 2005, 280, 16790–16797. [Google Scholar] [CrossRef] [Green Version]
- Skopin, A.; Shalygin, A.; Vigont, V.; Zimina, O.; Glushankova, L.; Mozhayeva, G.N.; Kaznacheyeva, E. TRPC1 protein forms only one type of native store-operated channels in HEK293 cells. Biochimie 2013, 95, 347–353. [Google Scholar] [CrossRef]
- Shalygin, A.; Skopin, A.; Kalinina, V.; Zimina, O.; Glushankova, L.; Mozhayeva, G.N.; Kaznacheyeva, E. STIM1 and STIM2 Proteins Differently Regulate Endogenous Store-operated Channels in HEK293 Cells. J. Biol. Chem. 2015, 290, 4717–4727. [Google Scholar] [CrossRef] [Green Version]
- Almaça, J.; Tian, Y.; Aldehni, F.; Ousingsawat, J.; Kongsuphol, P.; Rock, J.R.; Harfe, B.D.; Schreiber, R.; Kunzelmann, K. TMEM16 Proteins Produce Volume-regulated Chloride Currents that Are Reduced in Mice Lacking TMEM16A. J. Biol. Chem. 2009, 284, 28571–28578. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, T.; Iehara, T.; Sato, K.; Fujii, T.; Sakai, H.; Okada, Y. TMEM16F is a component of a Ca2+-activated Cl− channel but not a volume-sensitive outwardly rectifying Cl− channel. Am. J. Physiol. Cell Physiol. 2013, 304, C748–C759. [Google Scholar] [CrossRef] [Green Version]
- Scudieri, P.; Caci, E.; Venturini, A.; Sondo, E.; Pianigiani, G.; Marchetti, C.; Ravazzolo, R.; Pagani, F.; Galietta, L.J.V. Ion channel and lipid scramblase activity associated with expression of TMEM16F/ANO6 isoforms. J. Physiol. 2015, 593, 3829–3848. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Schreiber, R.; Kunzelmann, K. Anoctamins are a family of Ca2+-activated Cl− channels. J. Cell Sci. 2012, 125, 4991–4998. [Google Scholar] [CrossRef] [Green Version]
- Le, T.; Le, S.C.; Yang, H. Drosophila Subdued is a moonlighting transmembrane protein 16 (TMEM16) that transports ions and phospholipids. J. Biol. Chem. 2019, 294, 4529–4537. [Google Scholar] [CrossRef] [Green Version]
- Ousingsawat, J.; Wanitchakool, P.; Schreiber, R.; Kunzelmann, K. Contribution of TMEM16F to pyroptotic cell death. Cell Death Dis. 2018, 9, 1–11. [Google Scholar] [CrossRef]
- Schenk, L.K.; Schulze, U.; Henke, S.; Weide, T.; Pavenstädt, H. TMEM16F Regulates Baseline Phosphatidylserine Exposure and Cell Viability in Human Embryonic Kidney Cells. Cell. Physiol. Biochem. 2016, 38, 2452–2463. [Google Scholar] [CrossRef] [Green Version]
- Cruz-Rangel, S.; de Jesús-Pérez, J.J.; Contreras-Vite, J.A.; Pérez-Cornejo, P.; Hartzell, H.C.; Arreola, J. Gating modes of calcium-activated chloride channels TMEM16A and TMEM16B. J. Physiol. 2015, 593, 5283–5298. [Google Scholar] [CrossRef] [Green Version]
- Hartzell, C.; Putzier, I.; Arreola, J. Calcium-activated chloride channels. Annu. Rev. Physiol. 2005, 67, 719–758. [Google Scholar] [CrossRef] [Green Version]
- Martins, J.R.; Faria, D.; Kongsuphol, P.; Reisch, B.; Schreiber, R.; Kunzelmann, K. Anoctamin 6 is an essential component of the outwardly rectifying chloride channel. Proc. Natl. Acad. Sci. USA 2011, 108, 18168–18172. [Google Scholar] [CrossRef] [Green Version]
- Ye, W.; Han, T.W.; He, M.; Jan, Y.N.; Jan, L.Y. Dynamic change of electrostatic field in TMEM16F permeation pathway shifts its ion selectivity. eLife 2019, 8, e45187. [Google Scholar] [CrossRef]
- Reisert, J.; Bauer, P.J.; Yau, K.-W.; Frings, S. The Ca-activated Cl Channel and its Control in Rat Olfactory Receptor Neurons. J. Gen. Physiol. 2003, 122, 349–364. [Google Scholar] [CrossRef] [Green Version]
- Parekh, A.B. Ca2+ microdomains near plasma membrane Ca2+ channels: Impact on cell function. J. Physiol. 2008, 586, 3043–3054. [Google Scholar] [CrossRef]
- Fakler, B.; Adelman, J.P. Control of KCa Channels by Calcium Nano/Microdomains. Neuron 2008, 59, 873–881. [Google Scholar] [CrossRef] [Green Version]
- Pedemonte, N.; Galietta, L.J.V. Structure and Function of TMEM16 Proteins (Anoctamins). Physiol. Rev. 2014, 94, 419–459. [Google Scholar] [CrossRef] [Green Version]
- Hartzell, H.C.; Whitlock, J.M. TMEM16 chloride channels are two-faced. J. Gen. Physiol. 2016, 148, 367–373. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.D.; Cho, H.; Koo, J.Y.; Tak, M.H.; Cho, Y.; Shim, W.-S.; Park, S.P.; Lee, J.; Lee, B.; Kim, B.-M.; et al. TMEM16A confers receptor-activated calcium-dependent chloride conductance. Nat. Cell Biol. 2008, 455, 1210–1215. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Yu, K.; Perez-Cornejo, P.; Cui, Y.; Arreola, J.; Hartzell, H.C. Voltage- and calcium-dependent gating of TMEM16A/Ano1 chloride channels are physically coupled by the first intracellular loop. Proc. Natl. Acad. Sci. USA 2011, 108, 8891–8896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, J.; Nam, J.H.; Park, H.W.; Oh, U.; Yoon, J.-H.; Lee, M.G. Dynamic modulation of ANO1/TMEM16A HCO3- permeability by Ca2+/calmodulin. Proc. Natl. Acad. Sci. USA 2013, 110, 360–365. [Google Scholar] [CrossRef] [Green Version]
- Pifferi, S.; Dibattista, M.; Menini, A. TMEM16B induces chloride currents activated by calcium in mammalian cells. Pflügers Arch. Eur. J. Physiol. 2009, 458, 1023–1038. [Google Scholar] [CrossRef]
- Lin, H.; Jun, I.; Woo, J.H.; Lee, M.G.; Kim, S.J.; Nam, J.H. Temperature-dependent increase in the calcium sensitivity and acceleration of activation of ANO6 chloride channel variants. Sci. Rep. 2019, 9, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Ye, W.; Han, T.W.; Nassar, L.M.; Zubia, M.; Jan, Y.N.; Jan, L.Y. Phosphatidylinositol-(4, 5)-bisphosphate regulates calcium gating of small-conductance cation channel TMEM16F. Proc. Natl. Acad. Sci. USA 2018, 115, E1667–E1674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvadia, C.; Lim, N.K.; Mosina, V.C.; Oostergetel, G.T.; Dutzler, R.; Paulino, C. Cryo-EM structures and functional characterization of the murine lipid scramblase TMEM16F. eLife 2019, 8, e44365. [Google Scholar] [CrossRef]
- Grubb, S.; Poulsen, K.A.; Juul, C.A.; Kyed, T.; Klausen, T.K.; Larsen, E.H.; Hoffmann, E.K. TMEM16F (Anoctamin 6), an anion channel of delayed Ca2+ activation. J. Gen. Physiol. 2013, 141, 585–600. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Kim, A.; David, T.; Palmer, D.; Jin, T.; Tien, J.; Huang, F.; Cheng, T.; Coughlin, S.R.; Jan, Y.N.; et al. TMEM16F Forms a Ca2+-Activated Cation Channel Required for Lipid Scrambling in Platelets during Blood Coagulation. Cell 2012, 151, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Roh, J.; Woo, J.H.; Kim, S.J.; Nam, J.H. TMEM16F/ANO6, a Ca2+-activated anion channel, is negatively regulated by the actin cytoskeleton and intracellular MgATP. Biochem. Biophys. Res. Commun. 2018, 503, 2348–2354. [Google Scholar] [CrossRef]
- Wang, M.; Yang, H.; Zheng, L.-Y.; Zhang, Z.; Tang, Y.-B.; Wang, G.-L.; Du, Y.-H.; Lv, X.-F.; Liu, J.; Zhou, J.-G.; et al. Downregulation of TMEM16A Calcium-Activated Chloride Channel Contributes to Cerebrovascular Remodeling During Hypertension by Promoting Basilar Smooth Muscle Cell Proliferation. Circulation 2012, 125, 697–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dayal, A.; Ng, S.F.J.; Grabner, M. Ca2+-activated Cl− channel TMEM16A/ANO1 identified in zebrafish skeletal muscle is crucial for action potential acceleration. Nat. Commun. 2019, 10, 115. [Google Scholar] [CrossRef]
- Peters, C.J.; Gilchrist, J.M.; Tien, J.; Bethel, N.P.; Qi, L.; Chen, T.; Wang, L.; Jan, Y.N.; Grabe, M.; Jan, L.Y. The Sixth Transmembrane Segment Is a Major Gating Component of the TMEM16A Calcium-Activated Chloride Channel. Neuron 2018, 97, 1063–1077. [Google Scholar] [CrossRef] [Green Version]
- Nestorovich, E.M.; Rostovtseva, T.K.; Bezrukov, S.M. Residue Ionization and Ion Transport through OmpF Channels. Biophys. J. 2003, 85, 3718–3729. [Google Scholar] [CrossRef] [Green Version]
- Liang, P.; Yang, H. Molecular underpinning of intracellular pH regulation on TMEM16F. J. Gen. Physiol. 2021, 153, e202012704. [Google Scholar] [CrossRef] [PubMed]
- Naraghi, M.; Neher, E. Linearized Buffered Ca2+ Diffusion in Microdomains and Its Implications for Calculation of [Ca2+] at the Mouth of a Calcium Channel. J. Neurosci. 1997, 17, 6961–6973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Courjaret, R.; Machaca, K. Mid-range Ca2+ signalling mediated by functional coupling between store-operated Ca2+ entry and IP3-dependent Ca2+ release. Nat. Commun. 2014, 5, 3916. [Google Scholar] [CrossRef] [Green Version]
- Rizzuto, R.; Pozzan, T. Microdomains of Intracellular Ca2+: Molecular Determinants and Functional Consequences. Physiol. Rev. 2006, 86, 369–408. [Google Scholar] [CrossRef]
- Ehlen, H.W.; Chinenkova, M.; Moser, M.; Munter, H.-M.; Krause, Y.; Gross, S.; Brachvogel, B.; Wuelling, M.; Kornak, U.; Vortkamp, A. Inactivation of anoctamin-6/Tmem16f, a regulator of phosphatidylserine scrambling in osteoblasts, leads to decreased mineral deposition in skeletal tissues. J. Bone Miner. Res. 2012, 28, 246–259. [Google Scholar] [CrossRef]
- Ousingsawat, J.; Wanitchakool, P.; Schreiber, R.; Wuelling, M.; Vortkamp, A.; Kunzelmann, K. Anoctamin-6 Controls Bone Mineralization by Activating the Calcium Transporter NCX1. J. Biol. Chem. 2015, 290, 6270–6280. [Google Scholar] [CrossRef] [Green Version]
- Ousingsawat, J.; Wanitchakool, P.; Kmit, A.; Romao, A.M.; Jantarajit, W.; Schreiber, R.; Kunzelmann, K. Anoctamin 6 mediates effects essential for innate immunity downstream of P2X7 receptors in macrophages. Nat. Commun. 2015, 6, 6245. [Google Scholar] [CrossRef]
- Lacruz, R.S.; Feske, S. Diseases caused by mutations in ORAI1 and STIM1. Ann. N. Y. Acad. Sci. 2015, 1356, 45–79. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kolesnikov, D.; Perevoznikova, A.; Gusev, K.; Glushankova, L.; Kaznacheyeva, E.; Shalygin, A. Electrophysiological Properties of Endogenous Single Ca2+ Activated Cl− Channels Induced by Local Ca2+ Entry in HEK293. Int. J. Mol. Sci. 2021, 22, 4767. https://doi.org/10.3390/ijms22094767
Kolesnikov D, Perevoznikova A, Gusev K, Glushankova L, Kaznacheyeva E, Shalygin A. Electrophysiological Properties of Endogenous Single Ca2+ Activated Cl− Channels Induced by Local Ca2+ Entry in HEK293. International Journal of Molecular Sciences. 2021; 22(9):4767. https://doi.org/10.3390/ijms22094767
Chicago/Turabian StyleKolesnikov, Dmitrii, Anastasiia Perevoznikova, Konstantin Gusev, Lyubov Glushankova, Elena Kaznacheyeva, and Alexey Shalygin. 2021. "Electrophysiological Properties of Endogenous Single Ca2+ Activated Cl− Channels Induced by Local Ca2+ Entry in HEK293" International Journal of Molecular Sciences 22, no. 9: 4767. https://doi.org/10.3390/ijms22094767
APA StyleKolesnikov, D., Perevoznikova, A., Gusev, K., Glushankova, L., Kaznacheyeva, E., & Shalygin, A. (2021). Electrophysiological Properties of Endogenous Single Ca2+ Activated Cl− Channels Induced by Local Ca2+ Entry in HEK293. International Journal of Molecular Sciences, 22(9), 4767. https://doi.org/10.3390/ijms22094767