
The Interference between SARS-CoV-2 and Tyrosine Kinase Receptor Signaling in Cancer

 , , and
, , and 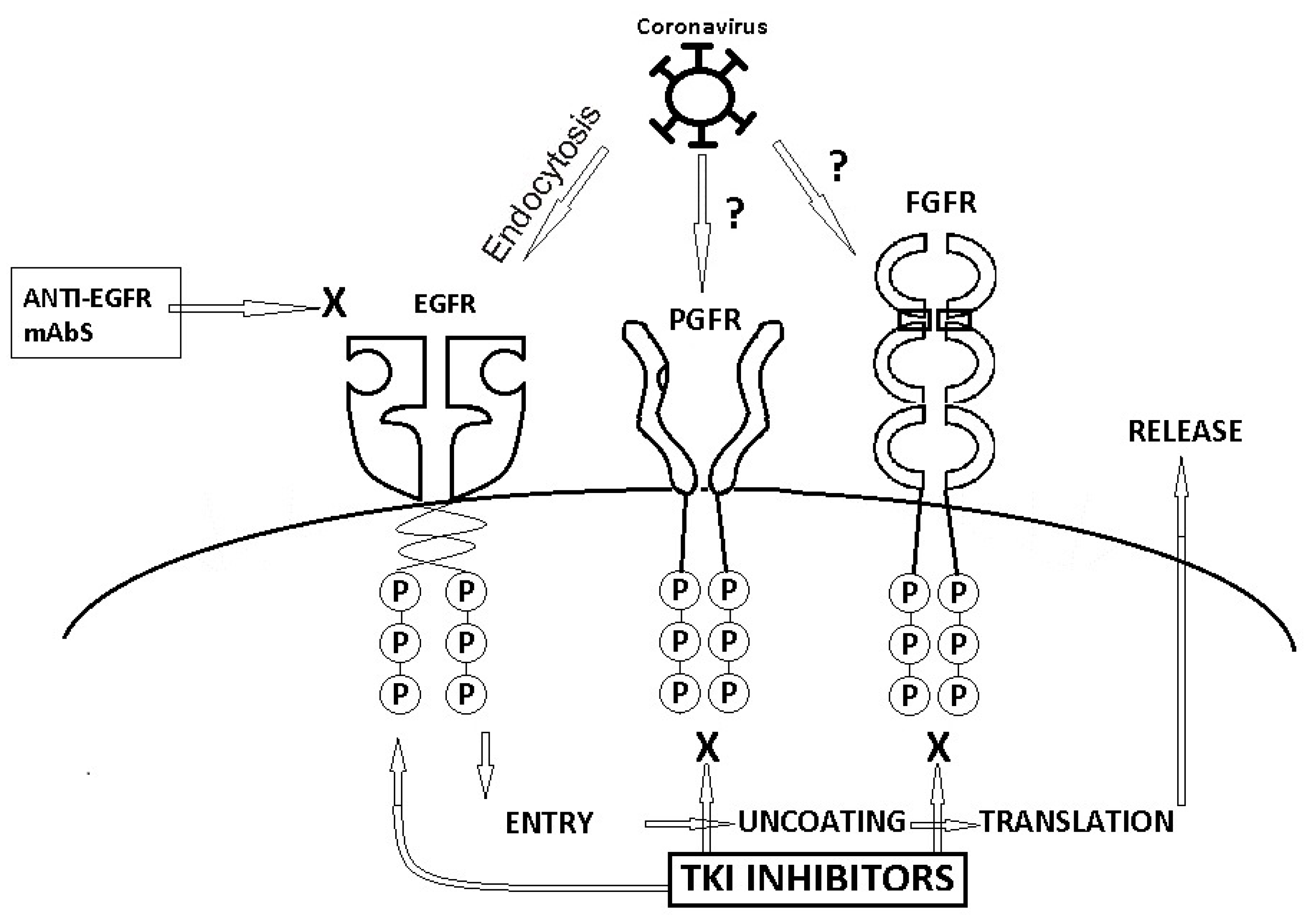{kind=link}
Abstract
:1. Introduction
2. Growth Factors, Tyrosine Kinase Receptors and SARS-CoV-2: A Complex Equation
2.1. The Epithelial Growth Factor Receptor
2.2. The Fibroblast Growth Factor Receptor
2.3. The Platelet-Derived Growth Factor Receptor
3. The Link between Antiviral and Anticancer Drugs
4. SARS-CoV-2 and Viral Tumorigenicity: A Tale of a Two-Edged Sword
4.1. SARS-CoV-2-Induced Carcinogenesis via Tyrosine Kinase Receptors
4.2. Oncolytic Virotherapy Potential of the Coronavirus
5. Discussion
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| TKR | Tyrosine kinase receptor |
| SARS-CoV | Severe acute respiratory syndrome coronavirus |
| MERS-CoV | Middle East respiratory syndrome coronavirus |
| ACE2 | Angiotensin conversion enzyme 2 |
| RBD | Receptor-binding domain |
| HR | Heptane repeats |
| PRRs | Pattern recognition receptors |
| PAMP | Pathogen-associated molecular patterns |
| TLR | Toll-like receptor |
| RLR-RIG | I-like receptor |
| NLR-NOD | Like receptor |
| CLmin-C | Type lectin-like receptors |
| MDA5 | Cytosolic receptor melanoma differentiation-associated gene 5 |
| cGAS | Nucleotidyltransferase cyclic GMP-AMP synthase |
| NF-κB | Transcription factor nuclear factor-κB |
| IRF3 | Interferon regulatory factor 3 |
| IFN | Interferon |
| GFR | Growth factor receptors |
| EGFR | Epithelial growth factor receptor |
| NSCLC | Non-small-cell lung cancer |
| HMCV | Human cytomegalovirus |
| TKI | Tyrosine kinase inhibitors |
| PI3K | Phosphoinositide 3 kinase |
| TGEV | Transmissible gastroenteritis virus |
| FGFR | Fibroblast growth factor receptor |
| EBV | Epstein–Barr virus |
| NPC | Non-keratinizing nasopharyngeal cancer |
| PDGFR | Platelet-derived growth factor receptors |
| BTK | Bruton tyrosine kinase |
| SINE | Selective inhibitor of nuclear export |
| MHCV | Mouse hepatitis coronavirus |
| CEACAM1a | Carcinoembryonic antigen-related cell adhesion molecule 1 |
| JAKi | JAK inhibitors |
| BBB | Blood–brain barrier |
References
- Li, W.; Shi, Z.; Yu, M.; Ren, W.; Smith, C.; Epstein, J.H.; Wang, H.; Crameri, G.; Hu, Z.; Zhang, H.; et al. Bats Are Natural Reservoirs of SARS-Like Coronaviruses. Science 2005, 310, 676–679. [Google Scholar] [CrossRef]
- Banerjee, A.; Kulcsar, K.; Misra, V.; Frieman, M.; Mossman, K. Bats and Coronaviruses. Viruses 2019, 11, 41. [Google Scholar] [CrossRef] [Green Version]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Rothan, H.A.; Byrareddy, S.N. The epidemiology and pathogenesis of coronavirus disease (COVID-19) outbreak. J. Autoimmun. 2020, 109, 102433. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Guan, X.; Wu, P.; Wang, X.; Zhou, L.; Tong, Y.; Ren, R.; Leung, K.S.M.; Lau, E.H.Y.; Wong, J.Y.; et al. Early transmission dynamics in Wuhan, China, of Novel Coronavirus—Infected pneumonia. N. Engl. J. Med. 2020, 382, 1199–1207. [Google Scholar] [CrossRef]
- Bai, Y.; Yao, L.; Wei, T.; Tian, F.; Jin, D.Y.; Chen, L.; Wang, M. Presumed Asymptomatic Carrier Transmission of COVID-19. JAMA 2020, 323, 1406–1407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Hu, B.; Hu, C.; Zhu, F.; Liu, X.; Zhang, J.; Wang, B.; Xiang, H.; Cheng, Z.; Xiong, Y.; et al. Clinical Characteristics of 138 Hospitalized Patients With 2019 Novel Coronavirus–Infected Pneumonia in Wuhan, China. JAMA 2020, 323, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Guan, W.J.; Ni, Z.Y.; Hu, Y.; Liang, W.H.; Ou, C.Q.; He, J.X.; Liu, L.; Shan, H.; Lei, C.L.; Hui, D.S.C.; et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef] [PubMed]
- Tortorici, M.A.; Veesler, D. Structural Insights into Coronavirus Entry. In Advances in Virus Research; Academic Press: Cambridge, MA, USA, 2019; pp. 93–116. [Google Scholar]
- Guo, Y.R.; Cao, Q.D.; Hong, Z.S.; Tan, Y.Y.; Chen, S.D.; Jin, H.J.; Tan, K.; Wang, D.; Yan, Y. The origin, transmission and clinical therapies on coronavirus disease 2019 (COVID-19) outbreak–an update on the status. Mil. Med Res. 2020, 7, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vicenzi, E.; Canducci, F.; Pinna, D.; Mancini, N.; Carletti, S.; Lazzarin, A.; Bordignon, C.; Poli, G.; Clementi, M. Coronaviridae and SARS-associated coronavirus strain HSR1. Emerg. Infect. Dis. 2004, 10, 413. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Chu, H.; Wong, B.H.-Y.; Chiu, M.C.; Wang, D.; Li, C.; Liu, X.; Yang, D.; Poon, V.K.-M.; Cai, J.; et al. Activation of C-Type Lectin Receptor and (RIG)-I-Like Receptors Contributes to Proinflammatory Response in Middle East Respiratory Syndrome Coronavirus-Infected Macrophages. J. Infect. Dis. 2019, 221, 647–659. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Fan, Y.; Lai, Y.; Han, T.; Li, Z.; Zhou, P.; Pan, P.; Wang, W.; Hu, D.; Liu, X.; et al. Coronavirus infections and immune responses. J. Med. Virol. 2020, 92, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Sun, L.; Chen, X.; Du, F.; Shi, H.; Chen, C.; Chen, Z.J. Cyclic GMP-AMP Is an Endogenous Second Messenger in Innate Immune Signaling by Cytosolic DNA. Science 2013, 339, 826–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeuchi, O.; Akira, S. Innate immunity to virus infection. Immunol. Rev. 2008, 227, 75–86. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Thorne, S.H.; Hermiston, T.; Kirn, D. Oncolytic Virotherapy: Approaches to Tumor Targeting and Enhancing Antitumor Effects. In Seminars in Oncology; W.B. Saunders Ltd.: Philadelphia, PA, USA, 2005. [Google Scholar]
- Liu, T.-C.; Galanis, E.; Kirn, D.H. Clinical trial results with oncolytic virotherapy: A century of promise, a decade of progress. Nat. Clin. Pract. Oncol. 2007, 4, 101–117. [Google Scholar] [CrossRef]
- Carapancea, M.; Alexandru, O.; Fetea, A.S.; Dragutescu, L.; Castro, J.; Georgescu, A.; Popa-Wagner, A.; Bäcklund, M.L.; Lewensohn, R.; Dricu, A. Growth factor receptors signaling in glioblastoma cells: Therapeutic implications. J. Neuro-Oncology 2008, 92, 137–147. [Google Scholar] [CrossRef]
- Popescu, A.M.; Alexandru, O.; Brindusa, C.; Purcaru, S.O.; Tache, D.E.; Tataranu, L.G.; Taisescu, C.; Dricu, A. Targeting the VEGF and PDGF signaling pathway in glioblastoma treatment. Int. J. Clin. Exp. Pathol. 2015, 8, 7825–7837. [Google Scholar]
- Hondermarck, H.; Bartlett, N.W.; Nurcombe, V. The role of growth factor receptors in viral infections: An opportunity for drug repurposing against emerging viral diseases such as COVID-19? FASEB BioAdvances 2020, 2, 296–303. [Google Scholar] [CrossRef]
- Carapancea, M.; Cosaceanu, D.; Budiu, R.; Kwiecinska, A.; Tataranu, L.G.; Ciubotaru, V.; Alexandru, O.; Banita, M.; Pisoschi, C.; Bäcklund, M.L.; et al. Dual targeting of IGF-1R and PDGFR inhibits proliferation in high-grade gliomas cells and induces radiosensitivity in JNK-1 expressing cells. J. Neuro-Oncology 2007, 85, 245–254. [Google Scholar] [CrossRef]
- Schlessinger, J. Receptor Tyrosine Kinases: Legacy of the First Two Decades. Cold Spring Harb. Perspect. Biol. 2014, 6, a008912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yarden, Y.; Pines, G. The ERBB network: At last, cancer therapy meets systems biology. Nat. Rev. Cancer 2012, 12, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, P.; Zhang, C.; Ma, Z. Epidermal growth factor receptor (EGFR): A rising star in the era of precision medicine of lung cancer. Oncotarget 2017, 8, 50209–50220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, W.; Zhang, S.; Shen, Y.; Yang, Q. Epidermal growth factor receptor is a co-factor for transmissible gastroenteritis virus entry. Virology 2018, 521, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Zhu, L.; Yang, X.; Lin, J.; Yang, Q. The epidermal growth factor receptor regulates cofilin activity and promotes transmissible gastroenteritis virus entry into intestinal epithelial cells. Oncotarget 2016, 7, 12206–12221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, W.; Xie, W.; Liu, Y.; Sui, B.; Zhang, H.; Liu, L.; Tan, Y.; Tong, X.; Fu, Z.F.; Yin, P.; et al. Receptor tyrosine kinase inhibitors block proliferation of TGEV mainly through p38 mitogen-activated protein kinase pathways. Antivir. Res. 2020, 173, 104651. [Google Scholar] [CrossRef] [PubMed]
- Venkataraman, T.; Coleman, C.M.; Frieman, M.B. Overactive Epidermal Growth Factor Receptor Signaling Leads to Increased Fibrosis after Severe Acute Respiratory Syndrome Coronavirus Infection. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [Green Version]
- Shah, R.R. Tyrosine Kinase Inhibitor-Induced Interstitial Lung Disease: Clinical Features, Diagnostic Challenges, and Therapeutic Dilemmas. Drug Saf. 2016, 39, 1073–1091. [Google Scholar] [CrossRef]
- Chang, H.-L.; Chen, Y.-H.; Taiwan, H.-C.; Yang, C.-J. EGFR Tyrosine Kinase Inhibitor–Associated Interstitial Lung Disease during the Coronavirus Disease 2019 Pandemic. J. Thorac. Oncol. 2020, 15, e129–e131. [Google Scholar] [CrossRef] [PubMed]
- Kato, T.; Nishio, K. Clinical aspects of epidermal growth factor receptor inhibitors: Benefit and risk. Respirology 2006, 11, 693–698. [Google Scholar] [CrossRef]
- Totura, A.L.; Whitmore, A.C.; Agnihothram, S.; Schäfer, A.; Katze, M.G.; Heise, M.T.; Baric, R.S. Toll-Like Receptor 3 Signaling via TRIF Contributes to a Protective Innate Immune Response to Severe Acute Respiratory Syndrome Coronavirus Infection. MBio 2015, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, H.; Aoshiba, K.; Yokohori, N.; Nagai, A. Epidermal growth factor receptor tyrosine kinase inhibition augments a murine model of pulmonary fibrosis. Cancer Res. 2003, 63, 5054–5059. [Google Scholar] [PubMed]
- Zhou, Y.; Lee, J.-Y.; Lee, C.-M.; Cho, W.-K.; Kang, M.-J.; Koff, J.L.; Yoon, P.-O.; Chae, J.; Park, H.-O.; Elias, J.A.; et al. Amphiregulin, an Epidermal Growth Factor Receptor Ligand, Plays an Essential Role in the Pathogenesis of Transforming Growth Factor-β-induced Pulmonary Fibrosis. J. Biol. Chem. 2012, 287, 41991–42000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardie, W.D.; Davidson, C.; Ikegami, M.; Leikauf, G.D.; Le Cras, T.D.; Prestridge, A.; Whitsett, J.A.; Korfhagen, T.R. EGF receptor tyrosine kinase inhibitors diminish transforming growth factor-α-induced pulmonary fibrosis. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2008, 294, L1217–L1225. [Google Scholar] [CrossRef] [PubMed]
- Venkataraman, T.; Frieman, M.B. The role of epidermal growth factor receptor (EGFR) signaling in SARS coronavirus-induced pulmonary fibrosis. Antivir. Res. 2017, 143, 142–150. [Google Scholar] [CrossRef]
- Ornitz, D.M.; Itoh, N. Fibroblast growth factors. Genome Biol. 2001, 2, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Itoh, N.; Ornitz, D.M. Evolution of the Fgf and Fgfr gene families. Trends Genet. 2004, 20, 563–569. [Google Scholar] [CrossRef]
- Porta, R.; Borea, R.; Coelho, A.; Khan, S.; Araújo, A.; Reclusa, P.; Franchina, T.; Van Der Steen, N.; Van Dam, P.; Ferri, J.; et al. FGFR a promising druggable target in cancer: Molecular biology and new drugs. Crit. Rev. Oncol. Hematol. 2017, 113, 256–267. [Google Scholar] [CrossRef] [Green Version]
- Ornitz, D.M. FGFs, heparan sulfate and FGFRs: Complex interactions essential for development. BioEssays 2000, 22, 108–112. [Google Scholar] [CrossRef]
- Qing, K.; Mah, C.; Hansen, J.; Zhou, S.; Dwarki, V.; Srivastava, A. Human fibroblast growth factor receptor 1 is a co-receptor for infection by adeno-associated virus 2. Nat. Med. 1999, 5, 71–77. [Google Scholar] [CrossRef]
- König, R.; Stertz, S.; Zhou, Y.; Inoue, A.; Hoffmann, H.-H.; Bhattacharyya, S.; Alamares, J.G.; Tscherne, D.M.; Ortigoza, M.B.; Liang, Y.; et al. Human host factors required for influenza virus replication. Nature 2010, 463, 813–817. [Google Scholar] [CrossRef] [PubMed]
- Cortese, M.; Kumar, A.; Matula, P.; Kaderali, L.; Scaturro, P.; Erfle, H.; Acosta, E.G.; Buehler, S.; Ruggieri, A.; Chatel-Chaix, L.; et al. Reciprocal Effects of Fibroblast Growth Factor Receptor Signaling on Dengue Virus Replication and Virion Production. Cell Rep. 2019, 27, 2579–2592.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limonta, D.; Jovel, J.; Kumar, A.; Lu, J.; Hou, S.; Airo, A.M.; Lopez-Orozco, J.; Wong, C.P.; Saito, L.; Branton, W.; et al. Fibroblast Growth Factor 2 Enhances Zika Virus Infection in Human Fetal Brain. J. Infect. Dis. 2019, 220, 1377–1387. [Google Scholar] [CrossRef] [PubMed]
- Lo, A.K.-F.; Dawson, C.W.; Young, L.S.; Ko, C.-W.; Hau, P.-M.; Lo, K.-W. Activation of the FGFR1 signalling pathway by the Epstein-Barr virus-encoded LMP1 promotes aerobic glycolysis and transformation of human nasopharyngeal epithelial cells. J. Pathol. 2015, 237, 238–248. [Google Scholar] [CrossRef] [Green Version]
- Yeung, M.-L.; Yao, Y.; Jia, L.; Chan, J.F.W.; Chan, K.-H.; Cheung, K.-F.; Chen, H.; Poon, V.K.M.; Tsang, A.K.L.; To, K.K.; et al. MERS coronavirus induces apoptosis in kidney and lung by upregulating Smad7 and FGF2. Nat. Microbiol. 2016, 1, 16004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.-H.; Chen, X.; He, X. Platelet-derived growth factors and their receptors: Structural and functional perspectives. Biochim. Biophys. Acta—Proteins Proteom. 2013, 1834, 2176–2186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levitzki, A. PDGF receptor kinase inhibitors for the treatment of PDGF driven diseases. Cytokine Growth Factor Rev. 2004, 15, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Vrijens, P.; Noppen, S.; Boogaerts, T.; Vanstreels, E.; Ronca, R.; Chiodelli, P.; Laporte, M.; Vanderlinden, E.; Liekens, S.; Stevaert, A.; et al. Influenza virus entry via the GM3 ganglioside-mediated platelet-derived growth factor receptor β signalling pathway. J. Gen. Virol. 2019, 100, 583–601. [Google Scholar] [CrossRef]
- Wu, K.; Oberstein, A.; Wang, W.; Shenk, T. Role of PDGF receptor-α during human cytomegalovirus entry into fibroblasts. Proc. Natl. Acad. Sci. USA 2018, 115, E9889–E9898. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Prager, A.; Boos, S.; Resch, M.; Brizic, I.; Mach, M.; Wildner, S.; Scrivano, L.; Adler, B. Human cytomegalovirus glycoprotein complex gH/gL/gO uses PDGFR-α as a key for entry. PLoS Pathog. 2017, 13, e1006281. [Google Scholar] [CrossRef]
- Liu, J.; Jardetzky, T.S.; Chin, A.L.; Johnson, D.C.; Vanarsdall, A.L. The Human Cytomegalovirus Trimer and Pentamer Promote Sequential Steps in Entry into Epithelial and Endothelial Cells at Cell Surfaces and Endosomes. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [Green Version]
- Allegra, A.; Pioggia, G.; Tonacci, A.; Musolino, C.; Gangemi, S. Cancer and SARS-CoV-2 Infection: Diagnostic and Therapeutic Challenges. Cancers 2020, 12, 1581. [Google Scholar] [CrossRef] [PubMed]
- Anil, I.; Arnold, R.; Benkwitz-Beford, S.; Branford, S.; Campton, N.; Cazier, J.-B.; Cheng, V.; Curley, H.; D’Costa, J.; Edmondson, A.; et al. The UK Coronavirus Cancer Monitoring Project: Protecting patients with cancer in the era of COVID-19. Lancet Oncol. 2020, 21, 622–624. [Google Scholar] [CrossRef]
- Florence, J.M.; Krupa, A.; Booshehri, L.M.; Davis, S.A.; Matthay, M.A.; Kurdowska, A.K. Inhibiting Bruton’s tyrosine kinase rescues mice from lethal influenza-induced acute lung injury. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2018, 315, L52–L58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treon, S.P.; Castillo, J.J.; Skarbnik, A.P.; Soumerai, J.D.; Ghobrial, I.M.; Guerrera, M.L.; Meid, K.E.; Yang, G. The BTK inhibitor ibrutinib may protect against pulmonary injury in COVID-19–infected patients. Blood 2020, 135, 1912–1915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roschewski, M.; Lionakis, M.S.; Sharman, J.P.; Roswarski, J.; Goy, A.; Monticelli, M.A.; Roshon, M.; Wrzesinski, S.H.; Desai, J.V.; Zarakas, M.A.; et al. Inhibition of Bruton tyrosine kinase in patients with severe COVID-19. Sci. Immunol. 2020, 5, eabd0110. [Google Scholar] [CrossRef] [PubMed]
- Allegra, A.; Innao, V.; Allegra, A.G.; Leanza, R.; Musolino, C. Selective inhibitors of nuclear export in the treatment of hematologic malignancies. Clin. Lymphoma Myeloma Leuk. 2019, 19, 689–698. [Google Scholar] [CrossRef] [PubMed]
- Agree, I. Karyopharm to Evaluate Low Dose Selinexor as a Potential Treatment for Hospitalized Patients with COVID-19. Available online: https://www.globenewswire.com/news-release/2020/04/07/2012711/0/en/Karyopharm-to-Evaluate-Low-Dose-Selinexor-as-a-Potential-Treatment-for-Hospitalized-Patients-with-COVID-19.html (accessed on 1 May 2021).
- Schor, S.; Einav, S. Repurposing of Kinase Inhibitors as Broad-Spectrum Antiviral Drugs. DNA Cell Biol. 2018, 37, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Kindrachuk, J.; Ork, B.; Hart, B.J.; Mazur, S.; Holbrook, M.R.; Frieman, M.B.; Traynor, D.; Johnson, R.F.; Dyall, J.; Kuhn, J.H.; et al. Antiviral Potential of ERK/MAPK and PI3K/AKT/mTOR Signaling Modulation for Middle East Respiratory Syndrome Coronavirus Infection as Identified by Temporal Kinome Analysis. Antimicrob. Agents Chemother. 2015, 59, 1088–1099. [Google Scholar] [CrossRef] [Green Version]
- Coleman, C.M.; Sisk, J.M.; Mingo, R.M.; Nelson, E.A.; White, J.M.; Frieman, M.B. Abelson Kinase Inhibitors Are Potent Inhibitors of Severe Acute Respiratory Syndrome Coronavirus and Middle East Respiratory Syndrome Coronavirus Fusion. J. Virol. 2016, 90, 8924–8933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assaad, H.S.; Assaad-Khalil, S. Imatinib a Tyrosine Kinase Inhibitor: A potential treatment for SARS- COV-2 induced pneumonia. Alex. J. Med. 2020, 56, 68–72. [Google Scholar] [CrossRef]
- Overbeek, M.J.; Amerongen, G.P.V.N.; Boonstra, A.; Smit, E.F.; Vonk-Noordegraaf, A. Possible role of imatinib in clinical pulmonary veno-occlusive disease. Eur. Respir. J. 2008, 32, 232–235. [Google Scholar] [CrossRef] [Green Version]
- Cahill, K.N.; Katz, H.; Cui, J.; Lai, J.; Kazani, S.; Crosby-Thompson, A.; Garofalo, D.; Castro, M.; Jarjour, N.N.; DiMango, E.; et al. Effect of KIT Inhibition by Imatinib on Airway Mast Cells in Patients with Severe Refractory Asthma (KIA). J. Allergy Clin. Immunol. 2017, 139, AB169. [Google Scholar] [CrossRef]
- Hoeper, M.M.; Barst, R.J.; Bourge, R.C.; Feldman, J.; Frost, A.E.; Galié, N.; Gómez-Sánchez, M.A.; Grimminger, F.; Grünig, E.; Hassoun, P.M.; et al. Imatinib mesylate as add-on therapy for pulmonary arterial hypertension: Results of the randomized IMPRES study. Circulation 2013, 127, 1128–1138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daniels, C.E.; Lasky, J.A.; Limper, A.H.; Mieras, K.; Gabor, E.; Schroeder, D.R. Imatinib treatment for idiopathic pulmonary fibrosis: Randomized placebo-controlled trial results. Am. J. Respir. Crit. Care Med. 2010, 181, 604–610. [Google Scholar] [CrossRef] [PubMed]
- Mulgaonkar, N.S.; Wang, H.; Mallawarachchi, S.; Ruzek, D.; Martina, B.; Fernando, S. Bcr-Abl tyrosine kinase inhibitor imatinib as a potential drug for COVID-19. bioRxiv 2020. [Google Scholar] [CrossRef]
- Rizzo, A.N.; Sammani, S.; Esquinca, A.E.; Jacobson, J.R.; Garcia, J.G.N.; Letsiou, E.; Dudek, S.M. Imatinib attenuates inflammation and vascular leak in a clinically relevant two-hit model of acute lung injury. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2015, 309, L1294–L1304. [Google Scholar] [CrossRef] [Green Version]
- Dosch, S.F.; Mahajan, S.D.; Collins, A.R. Collins, SARS coronavirus spike protein-induced innate immune response occurs via activation of the NF-κB pathway in human monocyte macrophages in vitro. Virus Res. 2009, 142, 19–27. [Google Scholar] [CrossRef]
- Morales-Ortega, A.; Bernal-Bello, D.; Llarena-Barroso, C.; Frutos-Pérez, B.; Duarte-Millán, M.Á.; de Viedma-García, V.G.; Farfán-Sedano, A.I.; Canalejo-Castrillero, E.; Ruiz-Giardín, J.M.; Ruiz-Ruiz, J.; et al. Imatinib for COVID-19: A case report. Clin. Immunol. 2020, 218, 108518. [Google Scholar] [CrossRef]
- Peterson, D.; Damsky, W.; King, B. The use of Janus kinase inhibitors in the time of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). J. Am. Acad. Dermatol. 2020, 82, e223–e226. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin-Drubin, M.E.; Munger, K. Viruses associated with human cancer. Biochim. Biophys. Acta—Mol. Basis Dis. 2008, 1782, 127–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, I.; Hatiboglu, M.A. Can COVID-19 induce glioma tumorogenesis through binding cell receptors? Med. Hypotheses 2020, 144, 110009. [Google Scholar] [CrossRef] [PubMed]
- Rhea, E.M.; Logsdon, A.F.; Hansen, K.M.; Williams, L.M.; Reed, M.J.; Baumann, K.K.; Holden, S.J.; Raber, J.; Banks, W.A.; Erickson, M.A.; et al. The S1 protein of SARS-CoV-2 crosses the blood-brain barrier in mice. Nat. Neurosci. 2021, 24, 368–378. [Google Scholar] [CrossRef]
- Pellegrini, L.; Albecka, A.; Mallery, D.L.; Kellner, M.J.; Paul, D.; Carter, A.P.; James, L.C.; Lancaster, M.A. SARS-CoV-2 Infects the Brain Choroid Plexus and Disrupts the Blood-CSF Barrier in Human Brain Organoids. Cell Stem Cell 2020, 27, 951–961. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhu, F.; Xie, L.; Wang, C.; Wang, J.; Chen, R.; Jia, P.; Guan, H.Q.; Peng, L.; Chen, Y.; et al. Clinical characteristics of COVID-19-infected cancer patients: A retrospective case study in three hospitals within Wuhan, China. Ann. Oncol. 2020, 31, 894–901. [Google Scholar] [CrossRef] [PubMed]
- Goulet, C.R.; Champagne, A.; Bernard, G.; Vandal, D.; Chabaud, S.; Pouliot, F.; Bolduc, S. Cancer-associated fibroblasts induce epithelial–mesenchymal transition of bladder cancer cells through paracrine IL-6 signalling. BMC Cancer 2019, 19, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, C.-W.; Chen, M.-W.; Hsiao, M.; Wang, S.; Chen, C.-A.; Hsiao, S.-M.; Chang, J.-S.; Lai, T.-C.; Rose-John, S.; Kuo, M.-L.; et al. IL-6 Trans-Signaling in Formation and Progression of Malignant Ascites in Ovarian Cancer. Cancer Res. 2010, 71, 424–434. [Google Scholar] [CrossRef] [Green Version]
- Ring, C.J.A. Cytolytic viruses as potential anti-cancer agents. J. Gen. Virol. 2002, 83, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Artene, S.-A.; Turcu-Stiolica, A.; Ciurea, M.E.; Folcuti, C.; Tataranu, L.G.; Alexandru, O.; Purcaru, O.S.; Tache, D.E.; Boldeanu, M.V.; Silosi, C.; et al. Comparative effect of immunotherapy and standard therapy in patients with high grade glioma: A meta-analysis of published clinical trials. Sci. Rep. 2018, 8, 11800. [Google Scholar] [CrossRef]
- Artene, S.-A.; Turcu-Stiolica, A.; Hartley, R.; Ciurea, M.E.; Daianu, O.; Brindusa, C.; Alexandru, O.; Tataranu, L.G.; Purcaru, Ş.O.; Dricu, A. Dendritic cell immunotherapy versus bevacizumab plus irinotecan in recurrent malignant glioma patients: A survival gain analysis. OncoTargets Ther. 2016, 9, 6669–6677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verheije, M.H.; Rottier, P.J.M. Retargeting of Viruses to Generate Oncolytic Agents. Adv. Virol. 2011, 2012, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Würdinger, T.; Verheije, M.H.; Broen, K.; Bosch, B.J.; Haijema, B.J.; De Haan, C.A.M.; Van Beusechem, V.W.; Gerritsen, W.R.; Rottier, P.J.M. Soluble Receptor-Mediated Targeting of Mouse Hepatitis Coronavirus to the Human Epidermal Growth Factor Receptor. J. Virol. 2005, 79, 15314–15322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verheije, M.H.; Lamfers, M.L.M.; Würdinger, T.; Grinwis, G.C.M.; Gerritsen, W.R.; Van Beusechem, V.W.; Rottier, P.J.M. Coronavirus Genetically Redirected to the Epidermal Growth Factor Receptor Exhibits Effective Antitumor Activity against a Malignant Glioblastoma. J. Virol. 2009, 83, 7507–7516. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Purcaru, O.-S.; Artene, S.-A.; Barcan, E.; Silosi, C.A.; Stanciu, I.; Danoiu, S.; Tudorache, S.; Tataranu, L.G.; Dricu, A. The Interference between SARS-CoV-2 and Tyrosine Kinase Receptor Signaling in Cancer. Int. J. Mol. Sci. 2021, 22, 4830. https://doi.org/10.3390/ijms22094830
Purcaru O-S, Artene S-A, Barcan E, Silosi CA, Stanciu I, Danoiu S, Tudorache S, Tataranu LG, Dricu A. The Interference between SARS-CoV-2 and Tyrosine Kinase Receptor Signaling in Cancer. International Journal of Molecular Sciences. 2021; 22(9):4830. https://doi.org/10.3390/ijms22094830
Chicago/Turabian StylePurcaru, Oana-Stefana, Stefan-Alexandru Artene, Edmond Barcan, Cristian Adrian Silosi, Ilona Stanciu, Suzana Danoiu, Stefania Tudorache, Ligia Gabriela Tataranu, and Anica Dricu. 2021. "The Interference between SARS-CoV-2 and Tyrosine Kinase Receptor Signaling in Cancer" International Journal of Molecular Sciences 22, no. 9: 4830. https://doi.org/10.3390/ijms22094830
APA StylePurcaru, O. -S., Artene, S. -A., Barcan, E., Silosi, C. A., Stanciu, I., Danoiu, S., Tudorache, S., Tataranu, L. G., & Dricu, A. (2021). The Interference between SARS-CoV-2 and Tyrosine Kinase Receptor Signaling in Cancer. International Journal of Molecular Sciences, 22(9), 4830. https://doi.org/10.3390/ijms22094830