
Age-Dependent Control of Collagen-Dependent Platelet Responses by Thrombospondin-1—Comparative Analysis of Platelets from Neonates, Children, Adolescents, and Adults

 ,
,  and
and
Abstract
:1. Introduction
2. Results
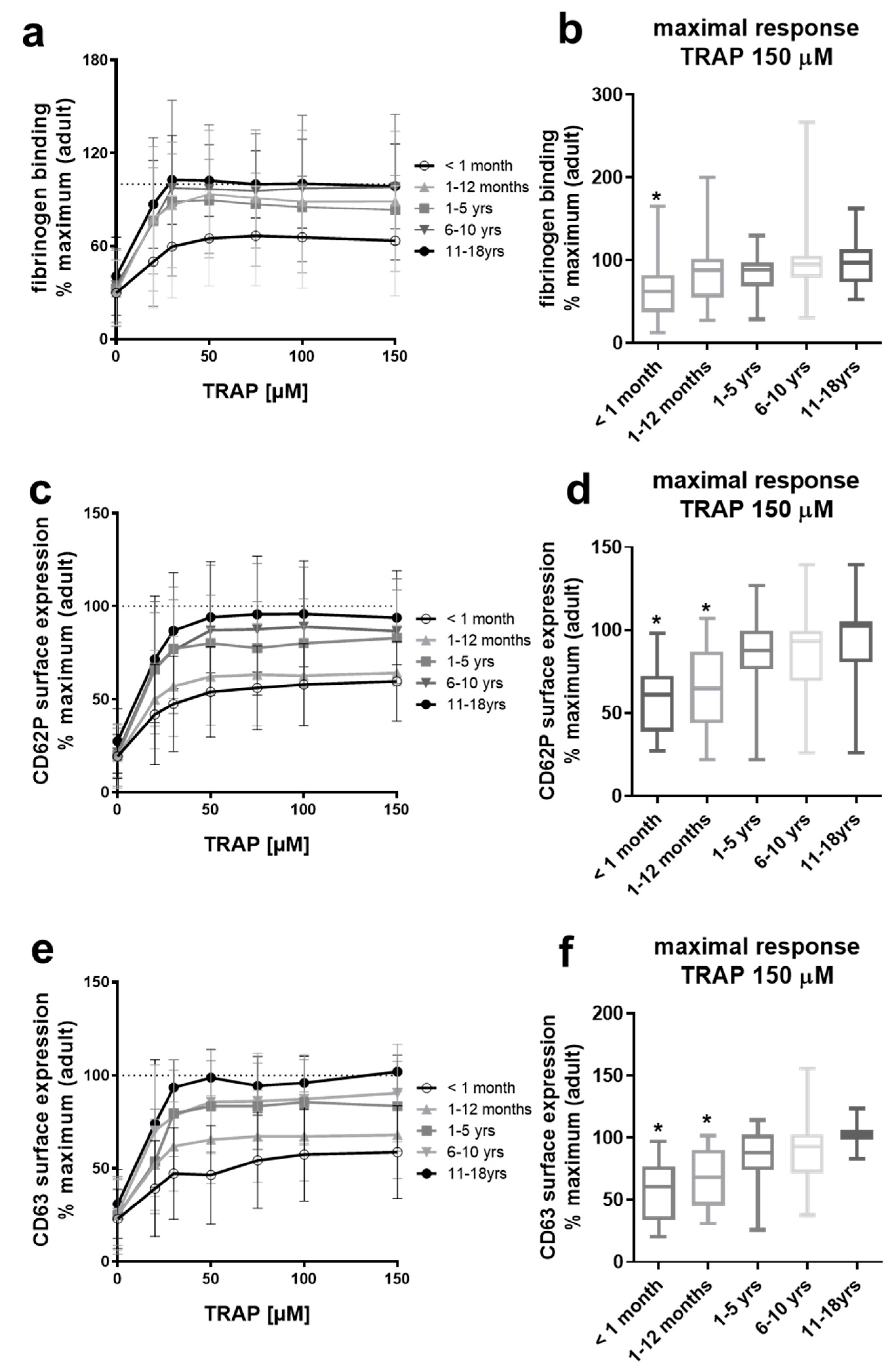
2.1. Platelets from Neonates and Infants Are Differentially Hypo-Reactive After PAR-1 Stimulation
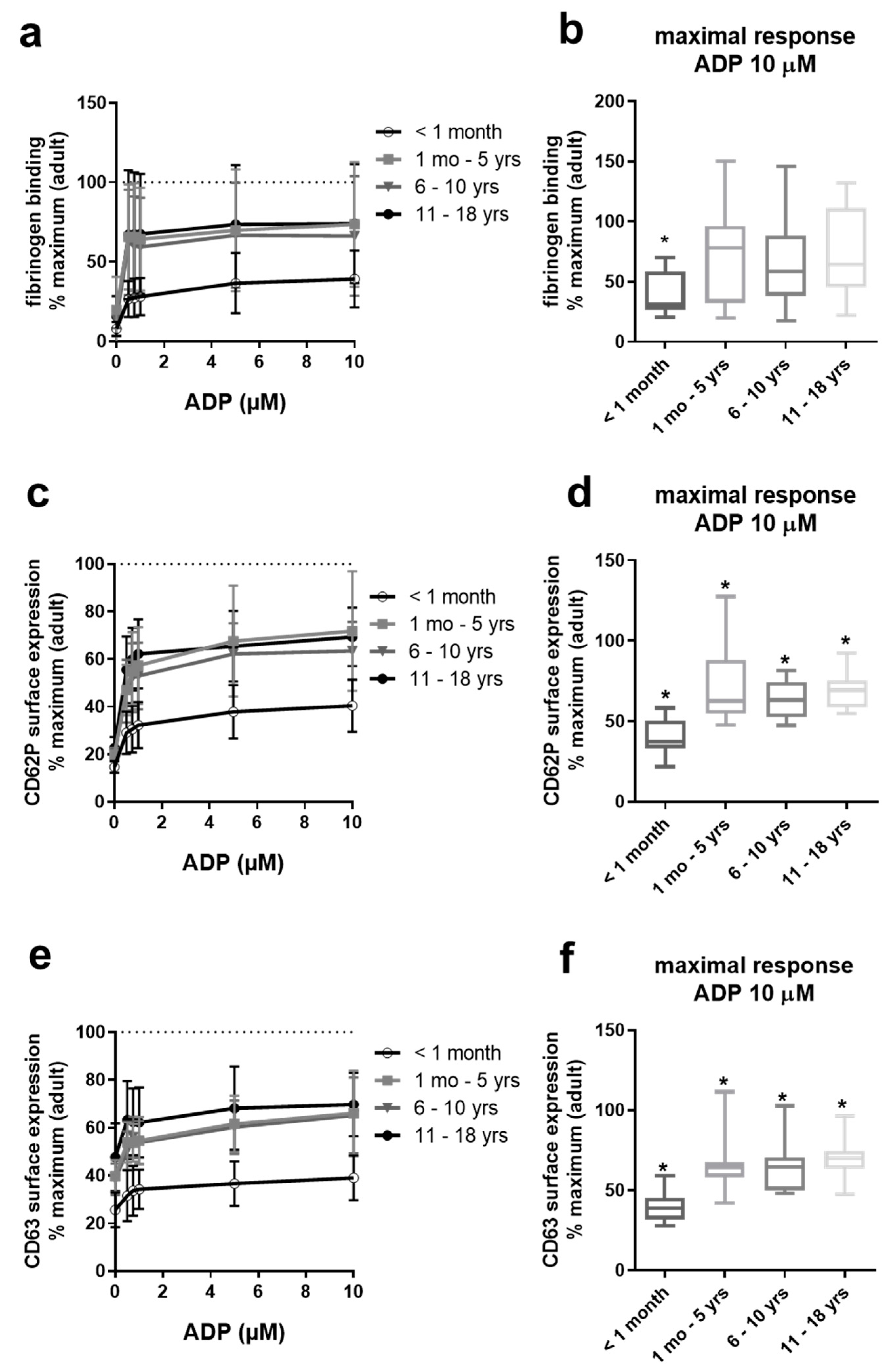
2.2. Gradual Hypo-Reactivity of Platelets from Pediatric Populations in Response to ADP
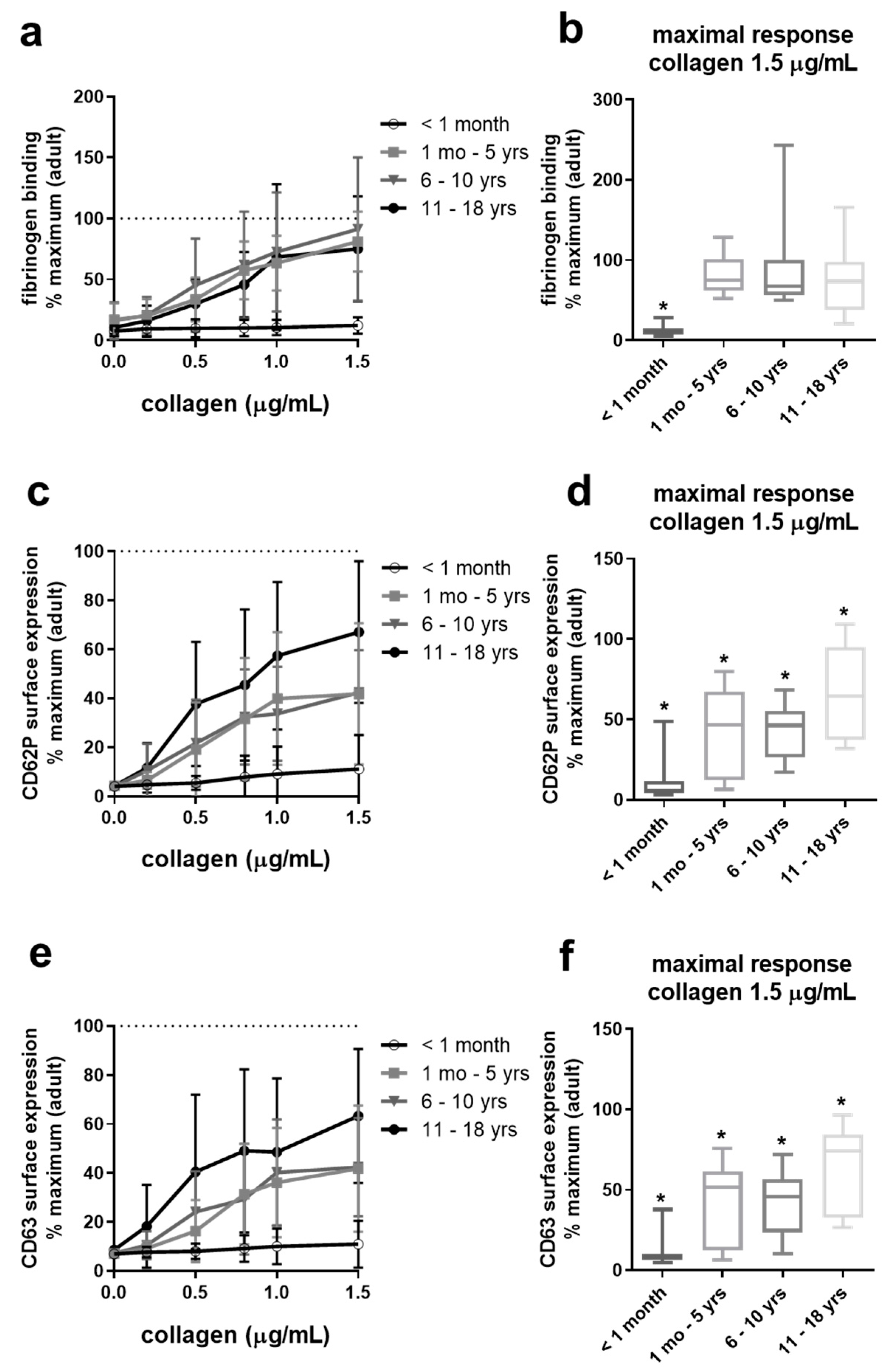
2.3. Collagen-Induced Platelet Activation Is Slightly Reduced in Adolescents, Differentially Reduced in Infants and Children, But Severely Impaired in Neonates
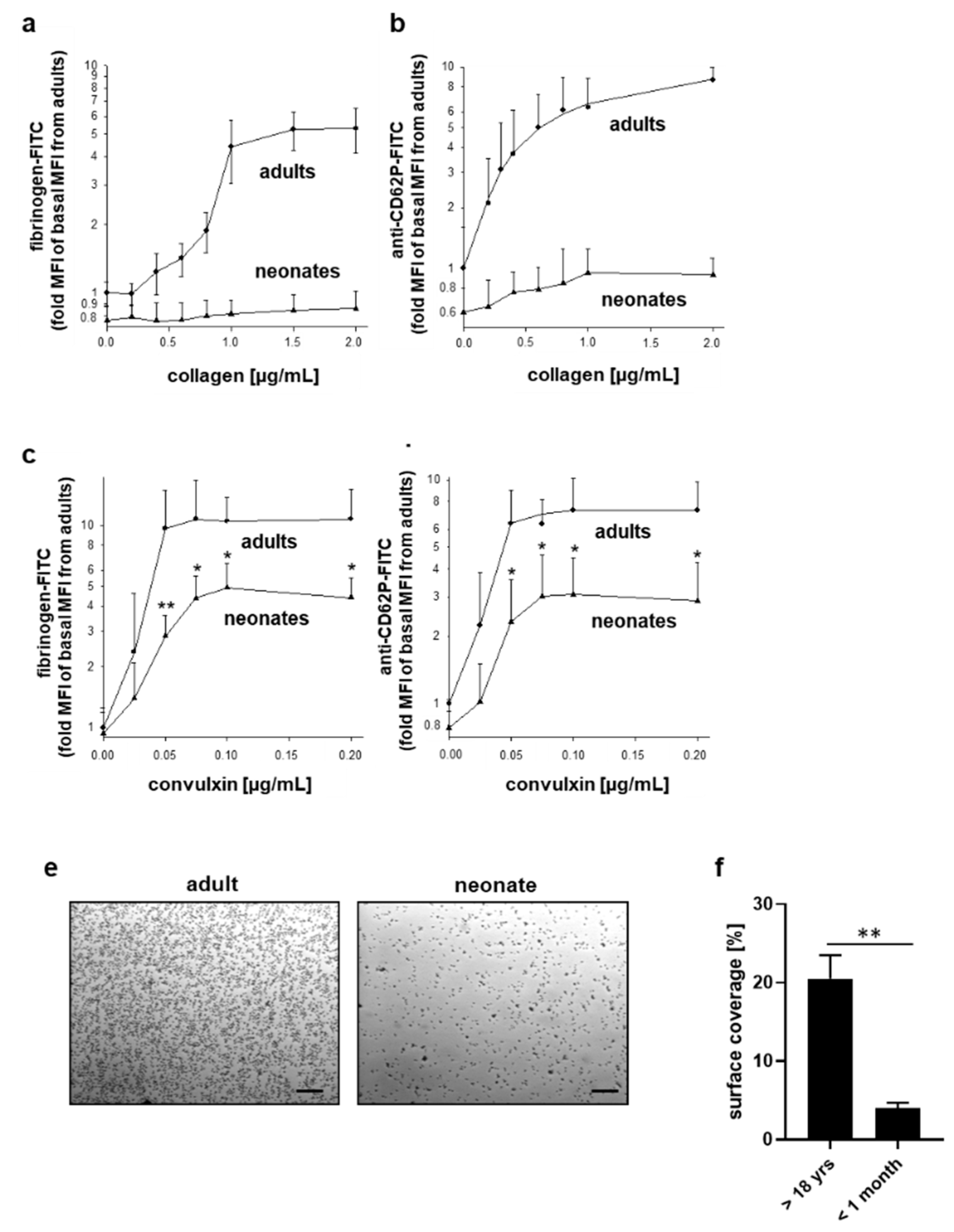
2.4. Differentially Impaired Responsiveness of Neonatal Platelets to Collagen and Convulxin
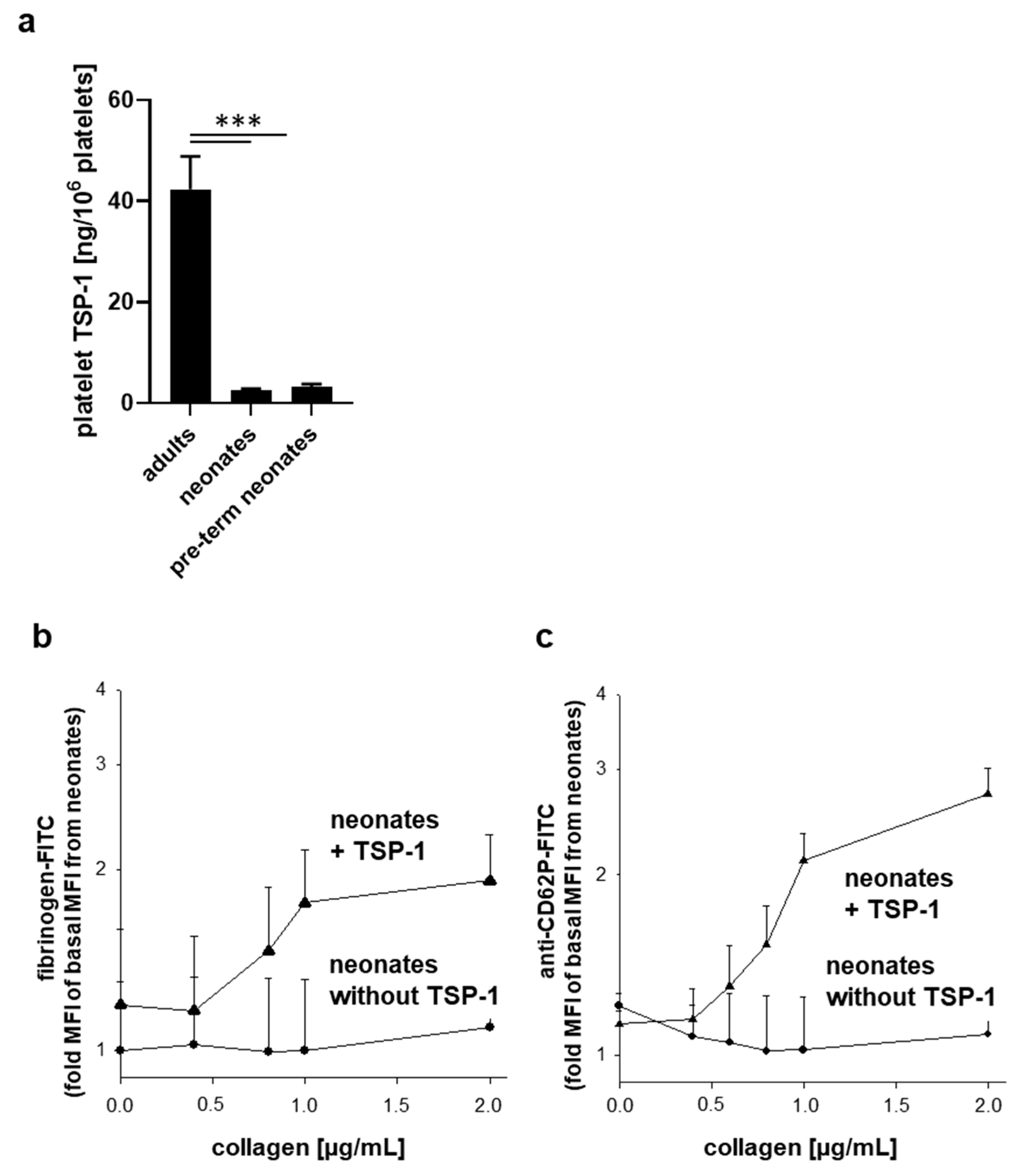
2.5. Neonatal Platelets Exhibit TSP-1 Deficiency and Exogenous TSP-1 Rescues the Platelet Activation Defect in Response to Collagen
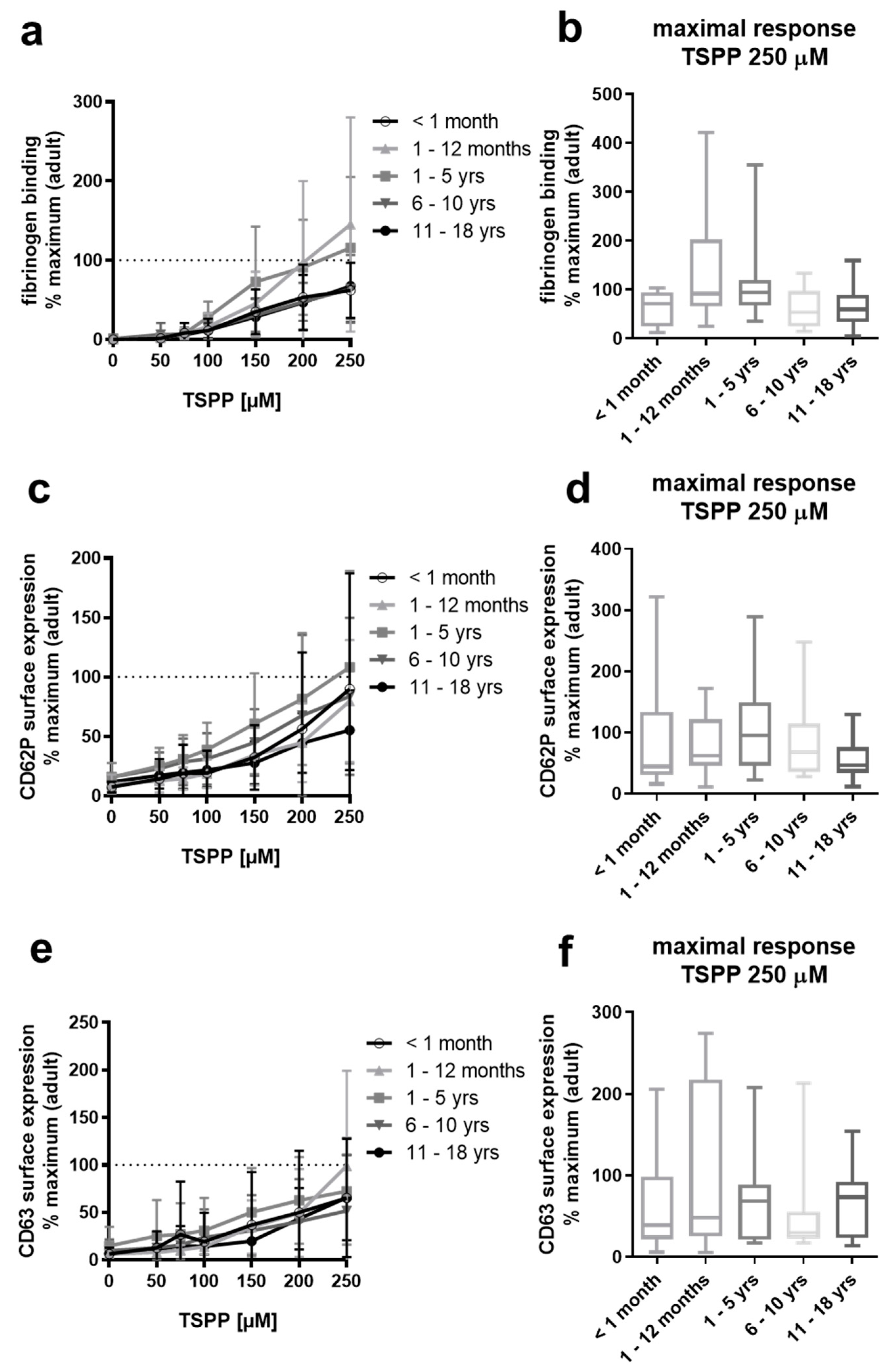
2.6. Platelets from Neonates, Infants, Children, and Adolescents Respond Normally to the TSP-1-C-Terminus-Derived Peptide RFYVVMWK
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Study Participants and Blood Collection
4.3. Preparation of Platelet-Rich Plasma and Washed Platelets
4.4. Flow Cytometric Analysis of Platelet Surface Receptors
4.5. Flow Cytometric Analysis of Agonist-Induced Platelet Activation
4.6. Platelet Adhesion Analysis under Flow
4.7. Quantification of Platelet TSP-1
4.8. SDS-PAGE and Protein Silver Staining
4.9. Data Analysis and Statistical Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Christensen, R.D.; Henry, E.; Wiedmeier, S.E.; Stoddard, R.A.; Sola-Visner, M.C.; Lambert, D.K.; Kiehn, T.I.; Ainsworth, S. Thrombocytopenia among extremely low birth weight neonates: Data from a multihospital healthcare system. J. Perinatol. Off. J. Calif. Perinat. Assoc. 2006, 26, 348–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cremer, M.; Weimann, A.; Schmalisch, G.; Hammer, H.; Buhrer, C.; Dame, C. Immature platelet values indicate impaired megakaryopoietic activity in neonatal early-onset thrombocytopenia. Thromb. Haemost. 2010, 103, 1016–1021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattia, G.; Vulcano, F.; Milazzo, L.; Barca, A.; Macioce, G.; Giampaolo, A.; Hassan, H.J. Different ploidy levels of megakaryocytes generated from peripheral or cord blood CD34+ cells are correlated with different levels of platelet release. Blood 2002, 99, 888–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tesfamariam, B. Distinct characteristics of neonatal platelet reactivity. Pharmacol. Res. 2017, 123, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Israels, S.J.; Rand, M.L.; Michelson, A.D. Neonatal platelet function. Semin. Thromb. Hemost. 2003, 29, 363–372. [Google Scholar] [CrossRef]
- Sola-Visner, M. Platelets in the neonatal period: Developmental differences in platelet production, function, and hemostasis and the potential impact of therapies. Hematol. Am. Soc. Hematol. Educ. Program 2012, 2012, 506–511. [Google Scholar] [CrossRef] [Green Version]
- Andres, O.; Schulze, H.; Speer, C.P. Platelets in neonates: Central mediators in haemostasis, antimicrobial defence and inflammation. Thromb. Haemost. 2015, 113, 3–12. [Google Scholar] [CrossRef]
- Margraf, A.; Nussbaum, C.; Sperandio, M. Ontogeny of platelet function. Blood Adv. 2019, 3, 692–703. [Google Scholar] [CrossRef] [Green Version]
- Urban, D.; Pluthero, F.G.; Christensen, H.; Baidya, S.; Rand, M.L.; Das, A.; Shah, P.S.; Chitayat, D.; Blanchette, V.S.; Kahr, W.H. Decreased numbers of dense granules in fetal and neonatal platelets. Haematologica 2017, 102, e36–e38. [Google Scholar] [CrossRef] [Green Version]
- Anderson, G.M.; Czarkowski, K.; Ravski, N.; Epperson, C.N. Platelet serotonin in newborns and infants: Ontogeny, heritability, and effect of in utero exposure to selective serotonin reuptake inhibitors. Pediatr. Res. 2004, 56, 418–422. [Google Scholar] [CrossRef] [Green Version]
- Mankin, P.; Maragos, J.; Akhand, M.; Saving, K.L. Imparied platelet-dense granule release in neonates. J. Pediatr. Hematol. Oncol. 2000, 22, 143–147. [Google Scholar] [CrossRef]
- Bluteau, O.; Langlois, T.; Rivera-Munoz, P.; Favale, F.; Rameau, P.; Meurice, G.; Dessen, P.; Solary, E.; Raslova, H.; Mercher, T.; et al. Developmental changes in human megakaryopoiesis. J. Thromb. Haemost. JTH 2013, 11, 1730–1741. [Google Scholar] [CrossRef]
- Ghevaert, C. Megakaryopoiesis through the ages: From the twinkle in the eye to the fully grown adult. J. Thromb. Haemost. JTH 2013, 11, 1727–1729. [Google Scholar] [CrossRef]
- Liu, Z.J.; Sola-Visner, M. Neonatal and adult megakaryopoiesis. Curr. Opin. Hematol. 2011, 18, 330–337. [Google Scholar] [CrossRef]
- Setzer, E.S.; Webb, I.B.; Wassenaar, J.W.; Reeder, J.D.; Mehta, P.S.; Eitzman, D.V. Platelet dysfunction and coagulopathy in intraventricular hemorrhage in the premature infant. J. Pediatr. 1982, 100, 599–605. [Google Scholar]
- Ferrer-Marin, F.; Stanworth, S.; Josephson, C.; Sola-Visner, M. Distinct differences in platelet production and function between neonates and adults: Implications for platelet transfusion practice. Transfusion 2013, 53, 2814–2821. [Google Scholar] [CrossRef]
- Bonduel, M.; Frontroth, J.P.; Hepner, M.; Sciuccati, G.; Feliu-Torres, A. Platelet aggregation and adenosine triphosphate release values in children and adults. J. Thromb. Haemost. JTH 2007, 5, 1782–1783. [Google Scholar] [CrossRef]
- Ravn, H.B.; Andreasen, J.B.; Hvas, A.M. Does whole blood coagulation analysis reflect developmental haemostasis? Blood Coagul. Fibrinolysis Int. J. Haemost. Thromb. 2017, 28, 218–223. [Google Scholar] [CrossRef]
- Knofler, R.; Weissbach, G.; Kuhlisch, E. Platelet function tests in childhood. Measuring aggregation and release reaction in whole blood. Semin. Thromb. Hemost. 1998, 24, 513–521. [Google Scholar] [CrossRef]
- Pietrucha, T.; Wojciechowski, T.; Greger, J.; Jecçdrzejewska, E.; Nowak, S.; Chrul, S.; Golanski, J.; Watala, C. Differentiated reactivity of whole blood neonatal platelets to various agonists. Platelets 2001, 12, 99–107. [Google Scholar] [CrossRef]
- Hezard, N.; Potron, G.; Schlegel, N.; Amory, C.; Leroux, B.; Nguyen, P. Unexpected persistence of platelet hyporeactivity beyond the neonatal period: A flow cytometric study in neonates, infants and older children. Thromb. Haemost. 2003, 90, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Hvas, A.M.; Favaloro, E.J. Platelet function testing in pediatric patients. Expert Rev. Hematol. 2017, 10, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Versteeg, H.H.; Heemskerk, J.W.; Levi, M.; Reitsma, P.H. New fundamentals in hemostasis. Physiol. Rev. 2013, 93, 327–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurk, K.; Kehrel, B.E. Platelets: Physiology and biochemistry. Semin. Thromb. Hemost. 2005, 31, 381–392. [Google Scholar] [CrossRef] [Green Version]
- Israels, S.J.; Daniels, M.; McMillan, E.M. Deficient collagen-induced activation in the newborn platelet. Pediatr. Res 1990, 27, 337–343. [Google Scholar] [CrossRef] [Green Version]
- Ucar, T.; Gurman, C.; Arsan, S.; Kemahli, S. Platelet aggregation in term and preterm newborns. Pediatr. Hematol. Oncol. 2005, 22, 139–145. [Google Scholar] [CrossRef]
- Gelman, B.; Setty, B.N.; Chen, D.; Amin-Hanjani, S.; Stuart, M.J. Impaired mobilization of intracellular calcium in neonatal platelets. Pediatr. Res. 1996, 39, 692–696. [Google Scholar] [CrossRef]
- Kehrel, B.; Wierwille, S.; Clemetson, K.J.; Anders, O.; Steiner, M.; Knight, C.G.; Farndale, R.W.; Okuma, M.; Barnes, M.J. Glycoprotein VI is a major collagen receptor for platelet activation: It recognizes the platelet-activating quaternary structure of collagen, whereas CD36, glycoprotein IIb/IIIa, and von Willebrand factor do not. Blood 1998, 91, 491–499. [Google Scholar] [CrossRef]
- Nieswandt, B.; Watson, S.P. Platelet-collagen interaction: Is GPVI the central receptor? Blood 2003, 102, 449–461. [Google Scholar] [CrossRef]
- Baker-Groberg, S.M.; Lattimore, S.; Recht, M.; McCarty, O.J.; Haley, K.M. Assessment of neonatal platelet adhesion, activation, and aggregation. J. Thromb. Haemost. JTH 2016, 14, 815–827. [Google Scholar] [CrossRef] [Green Version]
- Hardy, A.T.; Palma-Barqueros, V.; Watson, S.K.; Malcor, J.D.; Eble, J.A.; Gardiner, E.E.; Blanco, J.E.; Guijarro-Campillo, R.; Delgado, J.L.; Lozano, M.L.; et al. Significant Hypo-Responsiveness to GPVI and CLEC-2 Agonists in Pre-Term and Full-Term Neonatal Platelets and following Immune Thrombocytopenia. Thromb. Haemost. 2018, 118, 1009–1020. [Google Scholar] [CrossRef] [Green Version]
- Shenkman, B.; Linder, N.; Savion, N.; Tamarin, I.; Dardik, R.; Kennet, G.; German, B.; Varon, D. Increased neonatal platelet deposition on subendothelium under flow conditions: The role of plasma von Willebrand factor. Pediatr. Res. 1999, 45, 270–275. [Google Scholar] [CrossRef]
- Israels, S.J.; Cheang, T.; McMillan-Ward, E.M.; Cheang, M. Evaluation of primary hemostasis in neonates with a new in vitro platelet function analyzer. J. Pediatr. 2001, 138, 116–119. [Google Scholar] [CrossRef]
- Roschitz, B.; Sudi, K.; Kostenberger, M.; Muntean, W. Shorter PFA-100 closure times in neonates than in adults: Role of red cells, white cells, platelets and von Willebrand factor. Acta Paediatr. (Oslo, Norway: 1992) 2001, 90, 664–670. [Google Scholar] [CrossRef]
- Weinstein, M.J.; Blanchard, R.; Moake, J.L.; Vosburgh, E.; Moise, K. Fetal and neonatal von Willebrand factor (vWF) is unusually large and similar to the vWF in patients with thrombotic thrombocytopenic purpura. Br. J. Haematol. 1989, 72, 68–72. [Google Scholar] [CrossRef]
- Katz, J.A.; Moake, J.L.; McPherson, P.D.; Weinstein, M.J.; Moise, K.J.; Carpenter, R.J.; Sala, D.J. Relationship between human development and disappearance of unusually large von Willebrand factor multimers from plasma. Blood 1989, 73, 1851–1858. [Google Scholar] [CrossRef] [Green Version]
- Jurk, K.; Clemetson, K.J.; de Groot, P.G.; Brodde, M.F.; Steiner, M.; Savion, N.; Varon, D.; Sixma, J.J.; Van Aken, H.; Kehrel, B.E. Thrombospondin-1 mediates platelet adhesion at high shear via glycoprotein Ib (GPIb): An alternative/backup mechanism to von Willebrand factor. FASEB J. 2003, 17, 1490–1492. [Google Scholar] [CrossRef]
- Roberts, W.; Magwenzi, S.; Aburima, A.; Naseem, K.M. Thrombospondin-1 induces platelet activation through CD36-dependent inhibition of the cAMP/protein kinase A signaling cascade. Blood 2010, 116, 4297–4306. [Google Scholar] [CrossRef]
- Kehrel, B.; Balleisen, L.; Kokott, R.; Mesters, R.; Stenzinger, W.; Clemetson, K.J.; van de Loo, J. Deficiency of intact thrombospondin and membrane glycoprotein Ia in platelets with defective collagen-induced aggregation and spontaneous loss of disorder. Blood 1988, 71, 1074–1078. [Google Scholar] [CrossRef] [Green Version]
- Voit, S.; Udelhoven, M.; Lill, G.; Aktas, B.; Nieswandt, B.; Schror, K.; Weber, A.A. The C-terminal peptide of thrombospondin-1 stimulates distinct signaling pathways but induces an activation-independent agglutination of platelets and other cells. FEBS Lett. 2003, 544, 240–245. [Google Scholar] [CrossRef] [Green Version]
- Tulasne, D.; Judd, B.A.; Johansen, M.; Asazuma, N.; Best, D.; Brown, E.J.; Kahn, M.; Koretzky, G.A.; Watson, S.P. C-terminal peptide of thrombospondin-1 induces platelet aggregation through the Fc receptor gamma-chain-associated signaling pathway and by agglutination. Blood 2001, 98, 3346–3352. [Google Scholar] [CrossRef] [Green Version]
- Lagadec, P.; Dejoux, O.; Ticchioni, M.; Cottrez, F.; Johansen, M.; Brown, E.J.; Bernard, A. Involvement of a CD47-dependent pathway in platelet adhesion on inflamed vascular endothelium under flow. Blood 2003, 101, 4836–4843. [Google Scholar] [CrossRef] [Green Version]
- Jurk, K.; Ritter, M.A.; Schriek, C.; Van Aken, H.; Droste, D.W.; Ringelstein, E.B.; Kehrel, B.E. Activated monocytes capture platelets for heterotypic association in patients with severe carotid artery stenosis. Thromb. Haemost. 2010, 103, 1193–1202. [Google Scholar] [CrossRef]
- Michelson, A.D. Evaluation of platelet function by flow cytometry. Pathophysiol. Haemost. Thromb. 2006, 35, 67–82. [Google Scholar] [CrossRef]
- Kehrel, B.E.; Brodde, M.F. State of the art in platelet function testing. Transfus. Med. Hemother. 2013, 40, 73–86. [Google Scholar] [CrossRef] [Green Version]
- Bonnefoy, A.; Hoylaerts, M.F. Thrombospondin-1 in von Willebrand factor function. Curr. Drug Targets 2008, 9, 822–832. [Google Scholar] [CrossRef]
- Xie, L.; Chesterman, C.N.; Hogg, P.J. Control of von Willebrand factor multimer size by thrombospondin-1. J. Exp. Med. 2001, 193, 1341–1349. [Google Scholar] [CrossRef] [Green Version]
- Gatti, L.; Guarneri, D.; Caccamo, M.L.; Gianotti, G.A.; Marini, A. Platelet activation in newborns detected by flow-cytometry. Biol. Neonate 1996, 70, 322–327. [Google Scholar] [CrossRef]
- Ramstrom, S.; Sodergren, A.L.; Tynngard, N.; Lindahl, T.L. Platelet Function Determined by Flow Cytometry: New Perspectives? Semin. Thromb. Hemost. 2016, 42, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Jurk, K. Analysis of platelet function and dysfunction. Hamostaseologie 2015, 35, 60–72. [Google Scholar] [CrossRef]
- Bertling, A.; Fender, A.C.; Schungel, L.; Rumpf, M.; Mergemeier, K.; Geissler, G.; Sibrowski, W.; Kelsch, R.; Waltenberger, J.; Jakubowski, J.A.; et al. Reversibility of platelet P2Y12 inhibition by platelet supplementation: Ex vivo and in vitro comparisons of prasugrel, clopidogrel and ticagrelor. J. Thromb. Haemost. JTH 2018, 16, 1089–1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurk, K.; Shiravand, Y. Platelet Phenotyping and Function Testing in Thrombocytopenia. J. Clin. Med. 2021, 10, 1114. [Google Scholar] [CrossRef] [PubMed]
- Sitaru, A.G.; Holzhauer, S.; Speer, C.P.; Singer, D.; Obergfell, A.; Walter, U.; Grossmann, R. Neonatal platelets from cord blood and peripheral blood. Platelets 2005, 16, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Simak, J.; Holada, K.; Janota, J.; Stranak, Z. Surface expression of major membrane glycoproteins on resting and TRAP-activated neonatal platelets. Pediatr. Res. 1999, 46, 445–449. [Google Scholar] [CrossRef] [Green Version]
- Schlagenhauf, A.; Schweintzger, S.; Birner-Gruenberger, R.; Leschnik, B.; Muntean, W. Newborn platelets: Lower levels of protease-activated receptors cause hypoaggregability to thrombin. Platelets 2010, 21, 641–647. [Google Scholar] [CrossRef]
- Rehak, T.; Cvirn, G.; Gallistl, S.; Leschnik, B.; Kostenberger, M.; Katzer, H.; Ribitsch, V.; Muntean, W. Increased shear stress- and ristocetin-induced binding of von Willebrand factor to platelets in cord compared with adult plasma. Thromb. Haemost. 2004, 92, 682–687. [Google Scholar] [CrossRef]
- Kuijpers, M.J.; de Witt, S.; Nergiz-Unal, R.; van Kruchten, R.; Korporaal, S.J.; Verhamme, P.; Febbraio, M.; Tjwa, M.; Voshol, P.J.; Hoylaerts, M.F.; et al. Supporting roles of platelet thrombospondin-1 and CD36 in thrombus formation on collagen. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 1187–1192. [Google Scholar] [CrossRef] [Green Version]
- Lawler, J.; Sunday, M.; Thibert, V.; Duquette, M.; George, E.L.; Rayburn, H.; Hynes, R.O. Thrombospondin-1 is required for normal murine pulmonary homeostasis and its absence causes pneumonia. J. Clin. Investig. 1998, 101, 982–992. [Google Scholar] [CrossRef]
- Zhao, Y.; Olonisakin, T.F.; Xiong, Z.; Hulver, M.; Sayeed, S.; Yu, M.T.; Gregory, A.D.; Kochman, E.J.; Chen, B.B.; Mallampalli, R.K.; et al. Thrombospondin-1 restrains neutrophil granule serine protease function and regulates the innate immune response during Klebsiella pneumoniae infection. Mucosal Immunol. 2015, 8, 896–905. [Google Scholar] [CrossRef] [Green Version]
- Qu, Y.; Olonisakin, T.; Bain, W.; Zupetic, J.; Brown, R.; Hulver, M.; Xiong, Z.; Tejero, J.; Shanks, R.M.; Bomberger, J.M.; et al. Thrombospondin-1 protects against pathogen-induced lung injury by limiting extracellular matrix proteolysis. JCI Insight 2018, 3. [Google Scholar] [CrossRef] [Green Version]
- Margraf, A.; Nussbaum, C.; Rohwedder, I.; Klapproth, S.; Kurz, A.R.M.; Florian, A.; Wiebking, V.; Pircher, J.; Pruenster, M.; Immler, R.; et al. Maturation of Platelet Function During Murine Fetal Development In Vivo. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 1076–1086. [Google Scholar] [CrossRef] [Green Version]
- Davizon-Castillo, P.; McMahon, B.; Aguila, S.; Bark, D.; Ashworth, K.; Allawzi, A.; Campbell, R.A.; Montenont, E.; Nemkov, T.; D’Alessandro, A.; et al. TNF-alpha-driven inflammation and mitochondrial dysfunction define the platelet hyperreactivity of aging. Blood 2019, 134, 727–740. [Google Scholar] [CrossRef]
- Jain, K.; Tyagi, T.; Patell, K.; Xie, Y.; Kadado, A.J.; Lee, S.H.; Yarovinsky, T.; Du, J.; Hwang, J.; Martin, K.A.; et al. Age associated non-linear regulation of redox homeostasis in the anucleate platelet: Implications for CVD risk patients. EBioMedicine 2019, 44, 28–40. [Google Scholar] [CrossRef] [Green Version]
- Kehrel, B.; Kronenberg, A.; Schwippert, B.; Niesing-Bresch, D.; Niehues, U.; Tschope, D.; van de Loo, J.; Clemetson, K.J. Thrombospondin binds normally to glycoprotein IIIb deficient platelets. Biochem. Biophys. Res. Commun. 1991, 179, 985–991. [Google Scholar] [CrossRef]
- Rennemeier, C.; Hammerschmidt, S.; Niemann, S.; Inamura, S.; Zahringer, U.; Kehrel, B.E. Thrombospondin-1 promotes cellular adherence of gram-positive pathogens via recognition of peptidoglycan. FASEB J. 2007, 21, 3118–3132. [Google Scholar] [CrossRef]
- Knoppert, D.; Reed, M.; Benavides, S.; Totton, J.; Hoff, D.; Moffett, B.; Norris, K.; Vaillancout, R.; Aucoin, R.; Worthington, M. Paediatric age categories to be used in differentiating between listing on a model essential medicines list for children. WHO Postition Pap. 2007. [Google Scholar]
- Lahav, J.; Jurk, K.; Hess, O.; Barnes, M.J.; Farndale, R.W.; Luboshitz, J.; Kehrel, B.E. Sustained integrin ligation involves extracellular free sulfhydryls and enzymatically catalyzed disulfide exchange. Blood 2002, 100, 2472–2478. [Google Scholar] [CrossRef] [Green Version]
- Lahav, J.; Wijnen, E.M.; Hess, O.; Hamaia, S.W.; Griffiths, D.; Makris, M.; Knight, C.G.; Essex, D.W.; Farndale, R.W. Enzymatically catalyzed disulfide exchange is required for platelet adhesion to collagen via integrin alpha2beta1. Blood 2003, 102, 2085–2092. [Google Scholar] [CrossRef]
- Jurk, K.; Schulz, A.S.; Kehrel, B.E.; Rapple, D.; Schulze, H.; Mobest, D.; Friedrich, W.W.; Omran, H.; Deak, E.; Henschler, R.; et al. Novel integrin-dependent platelet malfunction in siblings with leukocyte adhesion deficiency-III (LAD-III) caused by a point mutation in FERMT3. Thromb. Haemost. 2010, 103, 1053–1064. [Google Scholar] [CrossRef]
- Loroch, S.; Trabold, K.; Gambaryan, S.; Reiss, C.; Schwierczek, K.; Fleming, I.; Sickmann, A.; Behnisch, W.; Zieger, B.; Zahedi, R.P.; et al. Alterations of the platelet proteome in type I Glanzmann thrombasthenia caused by different homozygous delG frameshift mutations in ITGA2B. Thromb. Haemost. 2017, 117, 556–569. [Google Scholar] [CrossRef]
- Shenkman, B.; Savion, N.; Dardik, R.; Tamarin, I.; Varon, D. Testing of platelet deposition on polystyrene surface under flow conditions by the cone and plate(let) analyzer: Role of platelet activation, fibrinogen and von Willebrand factor. Thromb. Res. 2000, 99, 353–361. [Google Scholar] [CrossRef]
- Blum, H.; Beier, H.; Gross, H.J. Improved Silver Staining of Plant-Proteins, Rna and DNA in Polyacrylamide Gels. Electrophoresis 1987, 8, 93–99. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Neonates (Full-Term ≥37 Weeks’ Gestation) | <1 Month (Total n = 46) |
|---|---|
| Infants | 1–12 months (total n = 51) |
| Young children | 1–5 years (total n = 56) |
| Children | 6–10 years (total n = 47) |
| Adolescents | 11–18 years (total n = 52) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herken, K.; Glauner, M.; Robert, S.C.; Maas, M.; Zippel, S.; Nowak-Göttl, U.; Zieger, B.; Lahav, J.; Fender, A.C.; Jurk, K.; et al. Age-Dependent Control of Collagen-Dependent Platelet Responses by Thrombospondin-1—Comparative Analysis of Platelets from Neonates, Children, Adolescents, and Adults. Int. J. Mol. Sci. 2021, 22, 4883. https://doi.org/10.3390/ijms22094883
Herken K, Glauner M, Robert SC, Maas M, Zippel S, Nowak-Göttl U, Zieger B, Lahav J, Fender AC, Jurk K, et al. Age-Dependent Control of Collagen-Dependent Platelet Responses by Thrombospondin-1—Comparative Analysis of Platelets from Neonates, Children, Adolescents, and Adults. International Journal of Molecular Sciences. 2021; 22(9):4883. https://doi.org/10.3390/ijms22094883
Chicago/Turabian StyleHerken, Katrin, Martin Glauner, Stefanie C. Robert, Matthias Maas, Sonja Zippel, Ulrike Nowak-Göttl, Barbara Zieger, Judith Lahav, Anke C. Fender, Kerstin Jurk, and et al. 2021. "Age-Dependent Control of Collagen-Dependent Platelet Responses by Thrombospondin-1—Comparative Analysis of Platelets from Neonates, Children, Adolescents, and Adults" International Journal of Molecular Sciences 22, no. 9: 4883. https://doi.org/10.3390/ijms22094883
APA StyleHerken, K., Glauner, M., Robert, S. C., Maas, M., Zippel, S., Nowak-Göttl, U., Zieger, B., Lahav, J., Fender, A. C., Jurk, K., & Kehrel, B. E. (2021). Age-Dependent Control of Collagen-Dependent Platelet Responses by Thrombospondin-1—Comparative Analysis of Platelets from Neonates, Children, Adolescents, and Adults. International Journal of Molecular Sciences, 22(9), 4883. https://doi.org/10.3390/ijms22094883