
Production of Aldehydes by Biocatalysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. One-Step Bioconversion—Enzymes That Produce Aldehydes
2.1. Aldehydes Produced as Primary Product
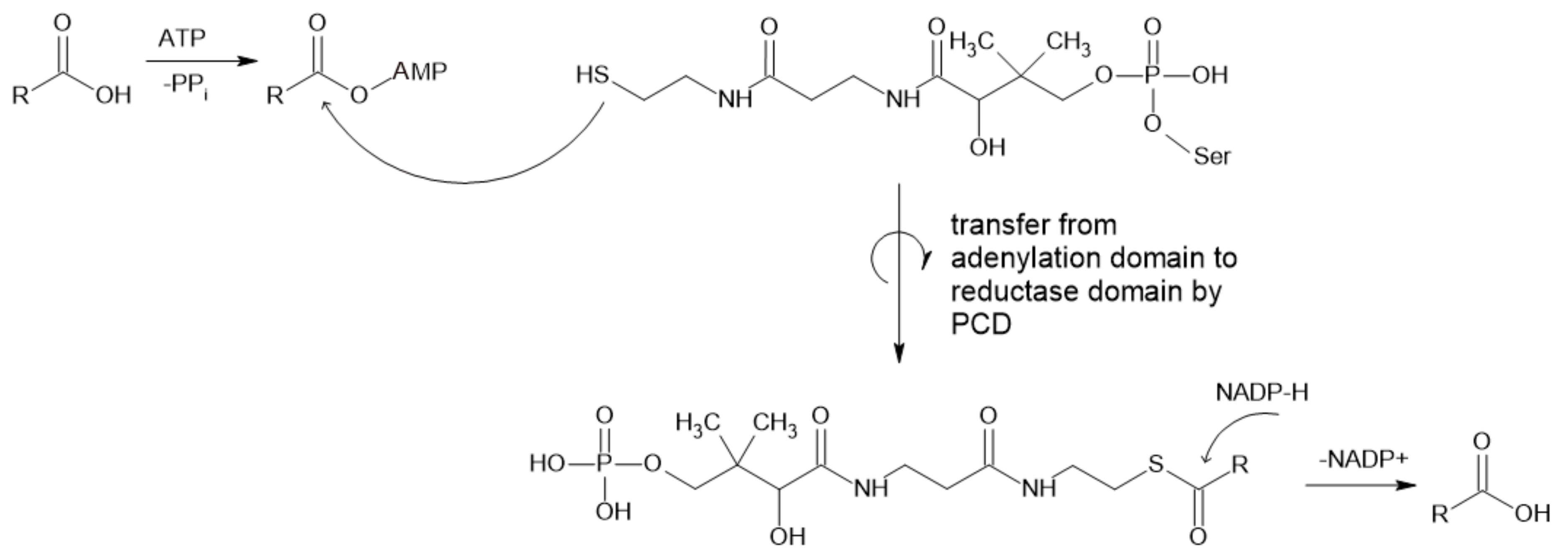
2.1.1. Carboxylic Acid Reductases
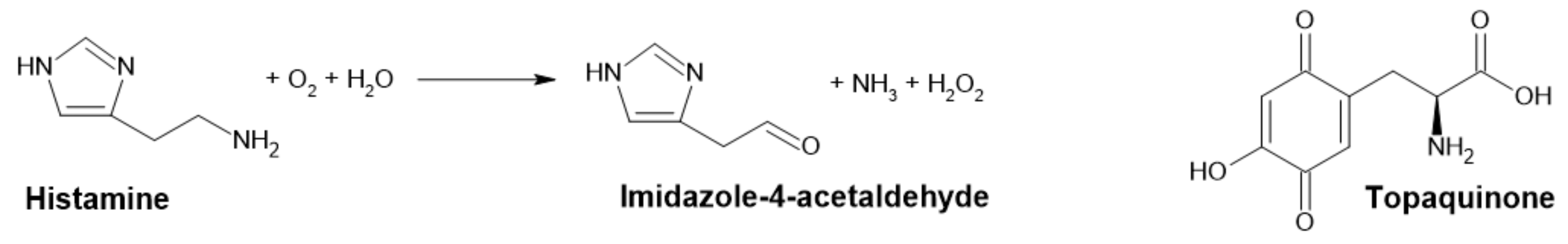
2.1.2. Diamine Oxidase
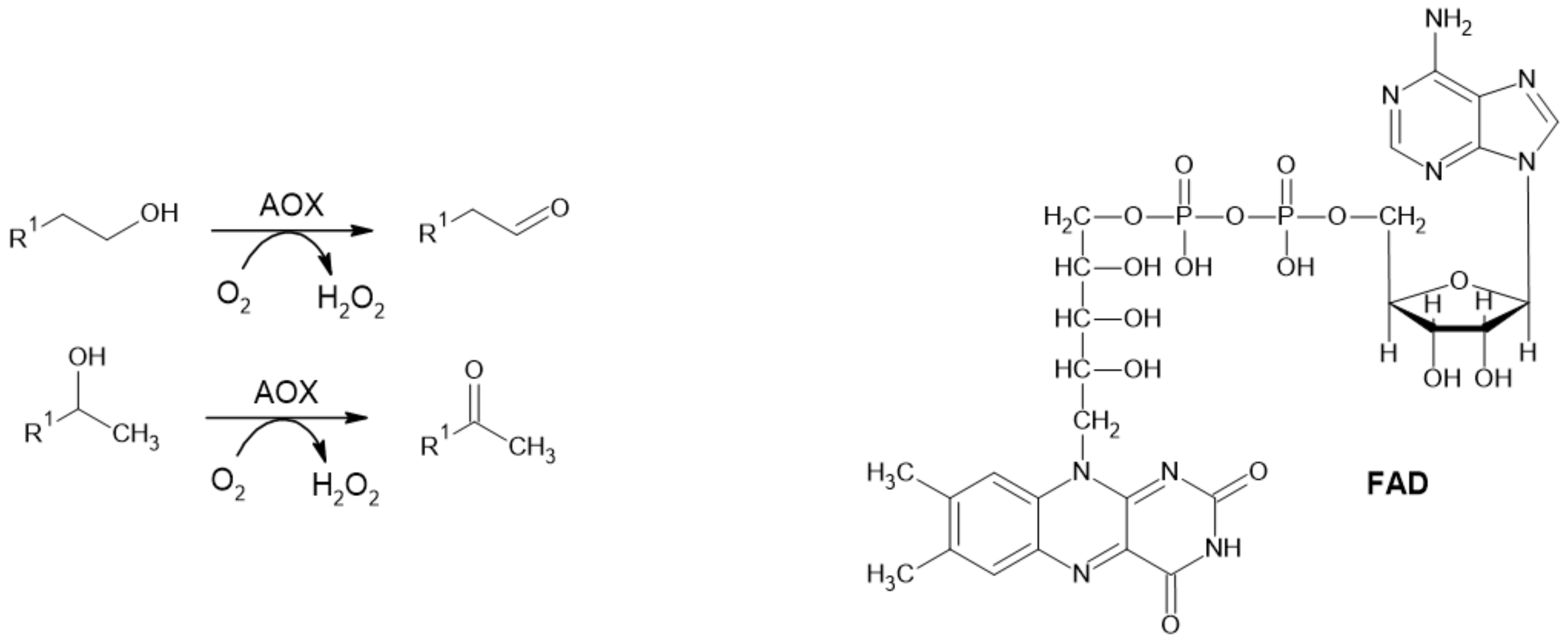
2.1.3. Alcohol Oxidase
2.2. Aldehydes Produced as Intermediates
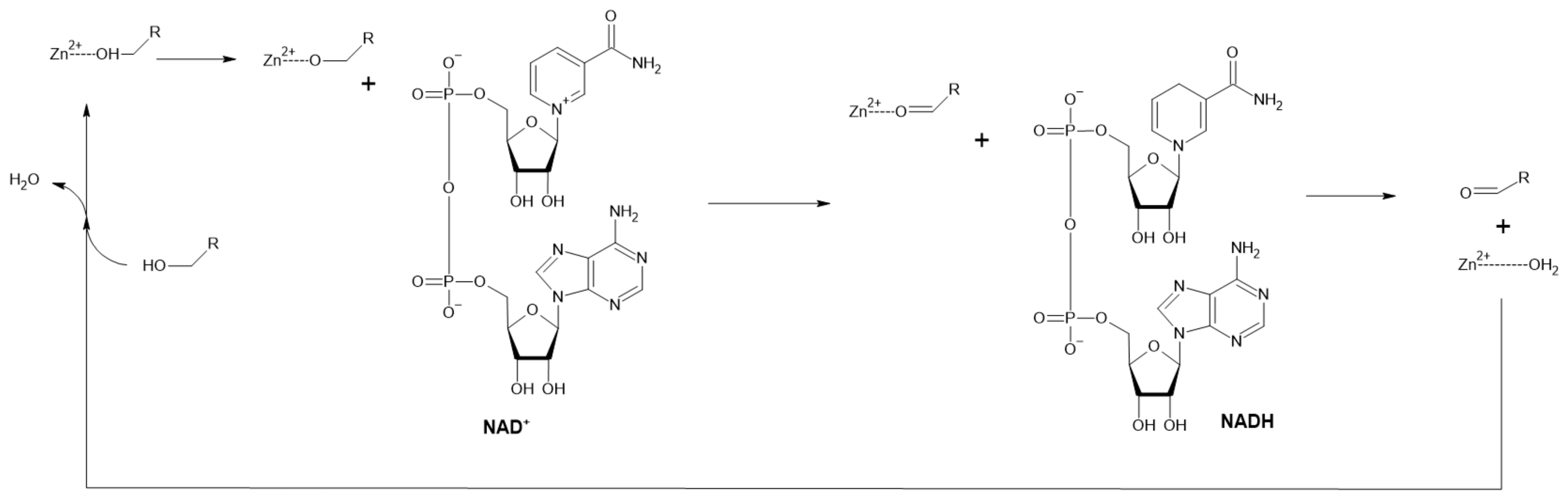
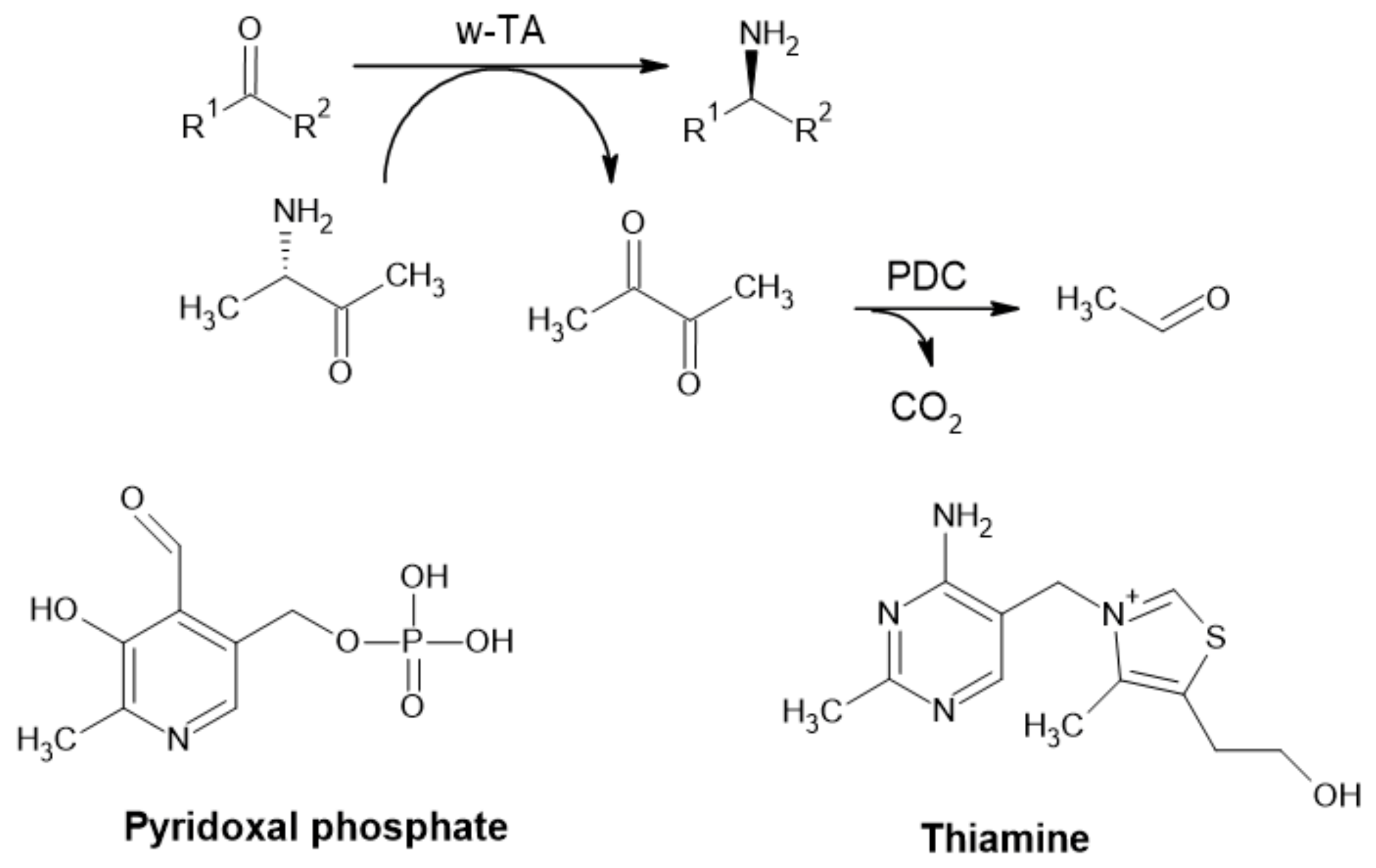
2.2.1. Alcohol Dehydrogenase
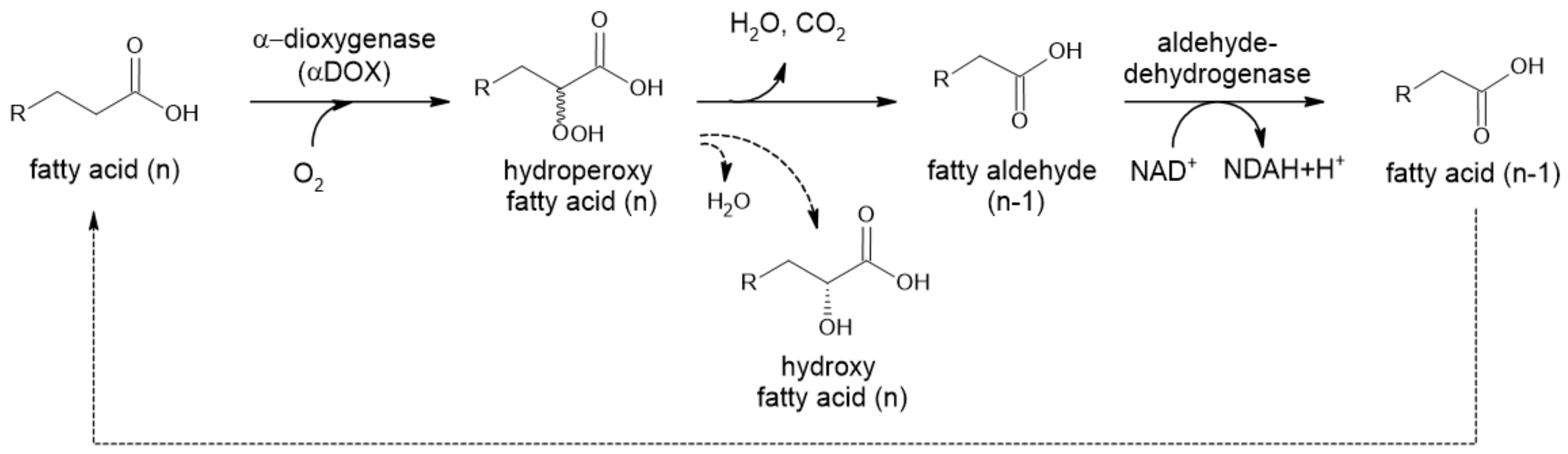
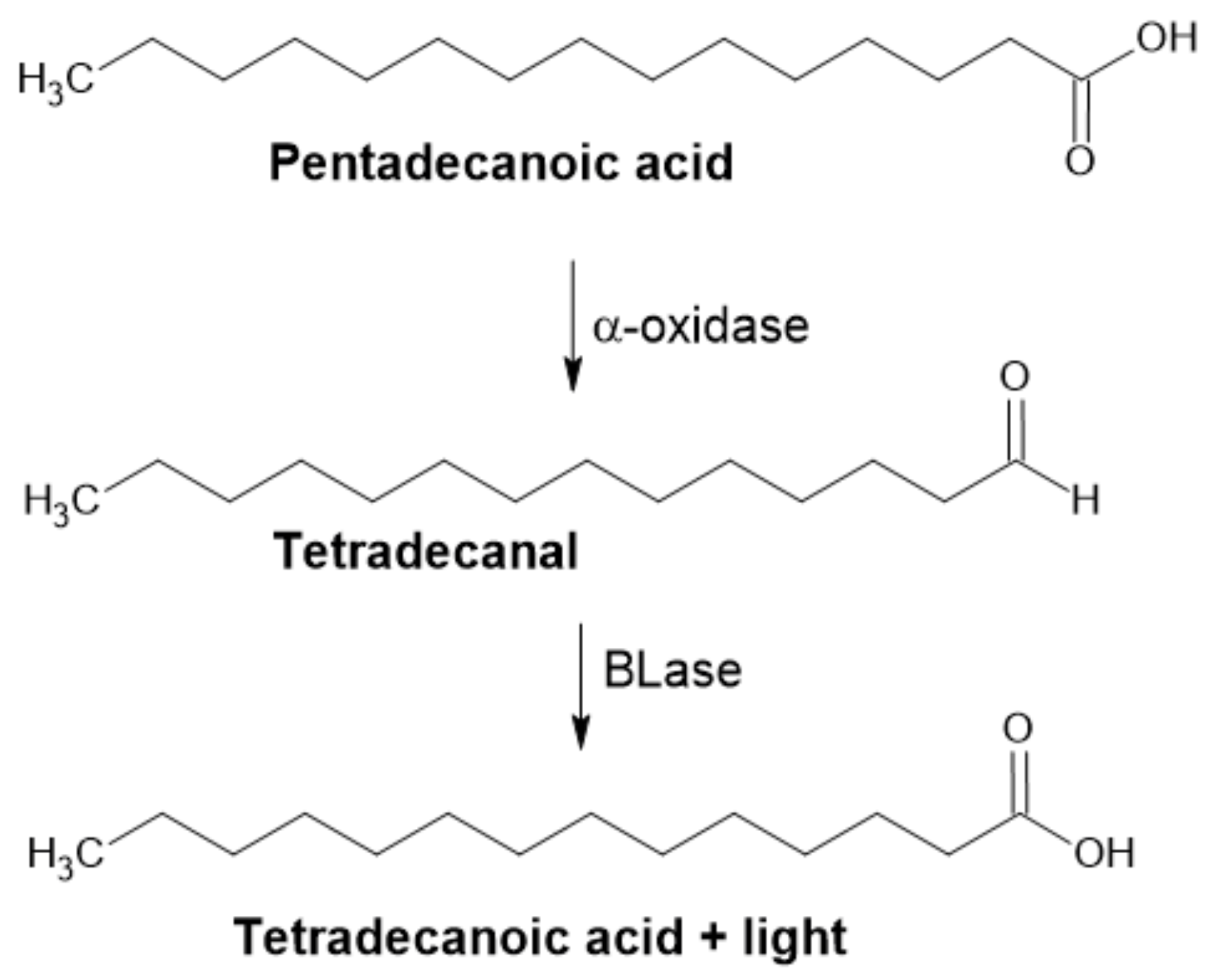
2.2.2. α-Dioxygenase
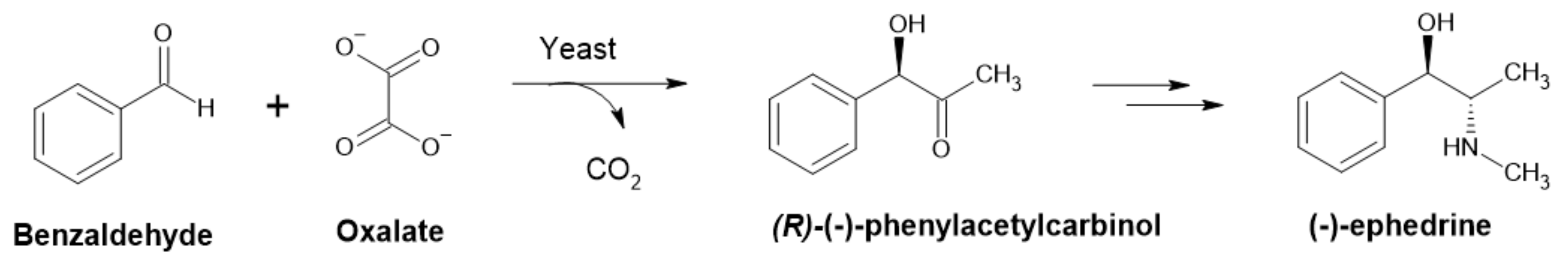
2.2.3. Pyruvate Decarboxylase
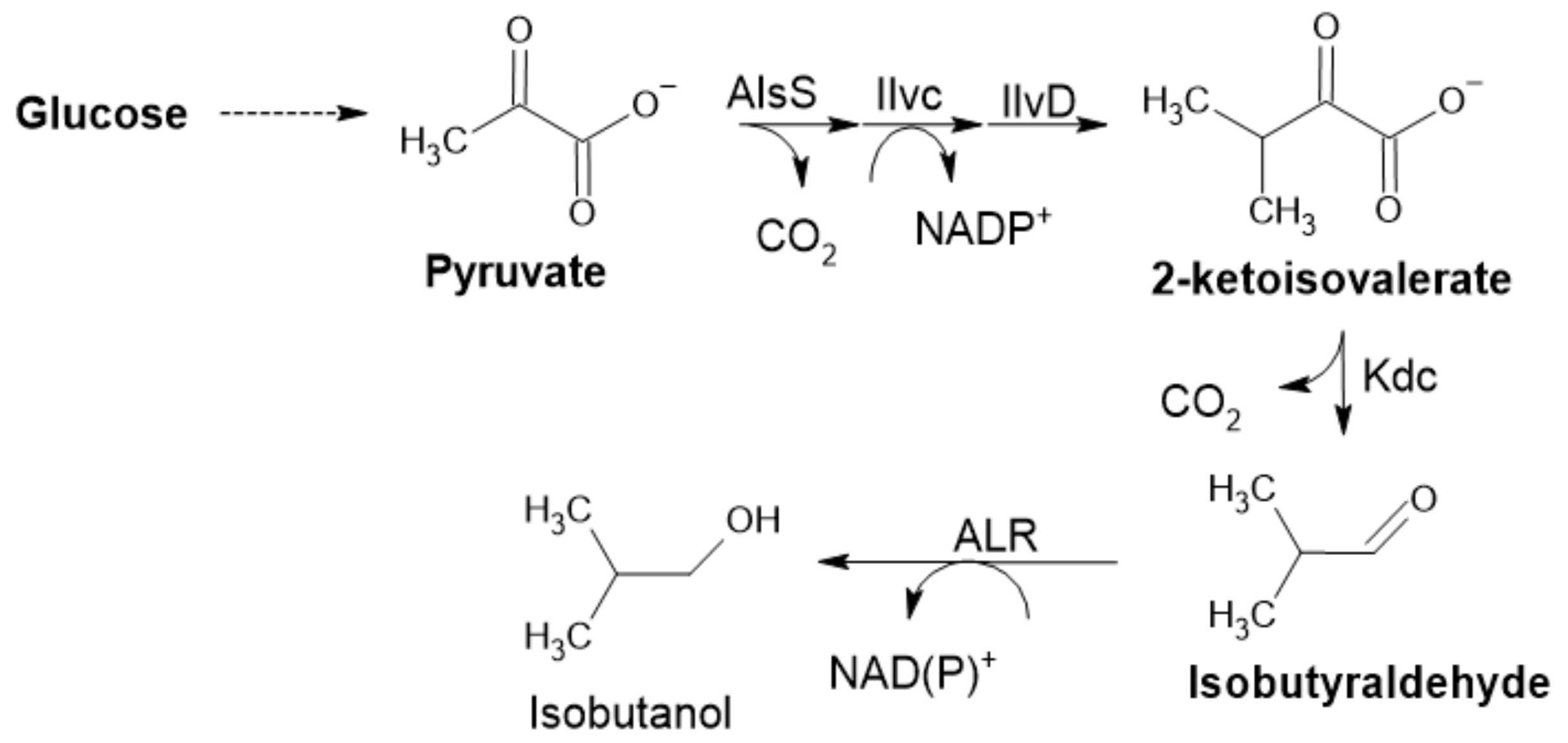
2.2.4. Acetohydroxyacid Synthase
2.2.5. α-Ketoisovalerate Decarboxylase
3. Aldehydes Produced by Enzymatic Cascades
Lipoxygenase Pathway
4. Engineering of Whole Cell Biocatalysts
4.1. Engineering of Microorganisms for the Accumulation of Aldehydes
4.2. Recombinant Biocatalysts
4.2.1. Biosynthesis and Regeneration of Cofactors
Heme Biosynthesis for HPL Expression
Post-Translational Modifications of 4-Phosphopantetheine (Ppant)
NAD(P)H Regeneration
ATP Regeneration
5. Analytics of Aldehydes Used in Biocatalysis
5.1. Photometric Methods
5.1.1. Detection of Aldehyde Products
Luciferase Assay
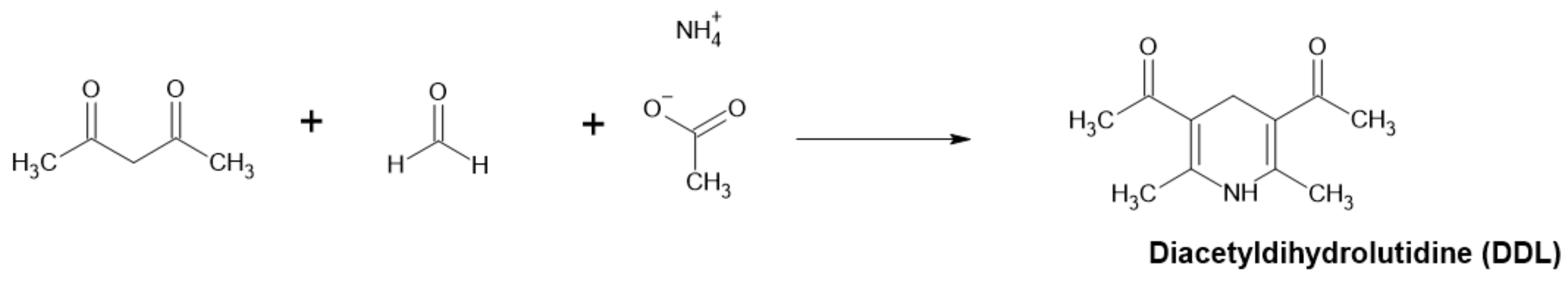
Detection of Formaldehyde by Acetylacetone and Ammonium Salt
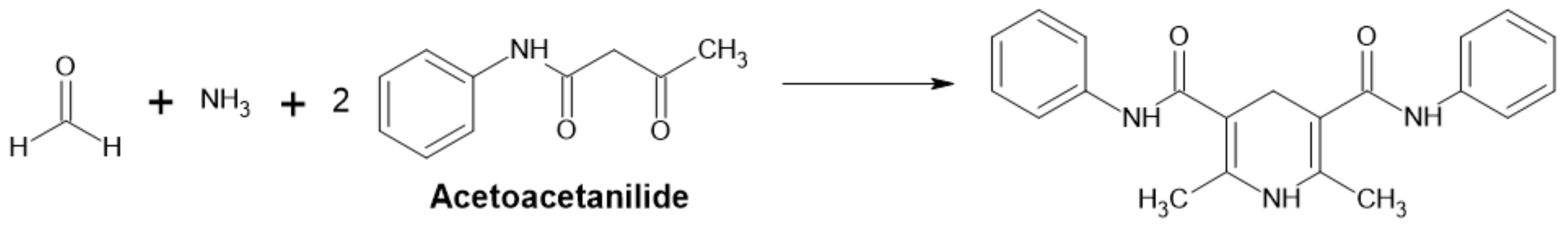
Acetylacetanilide Method
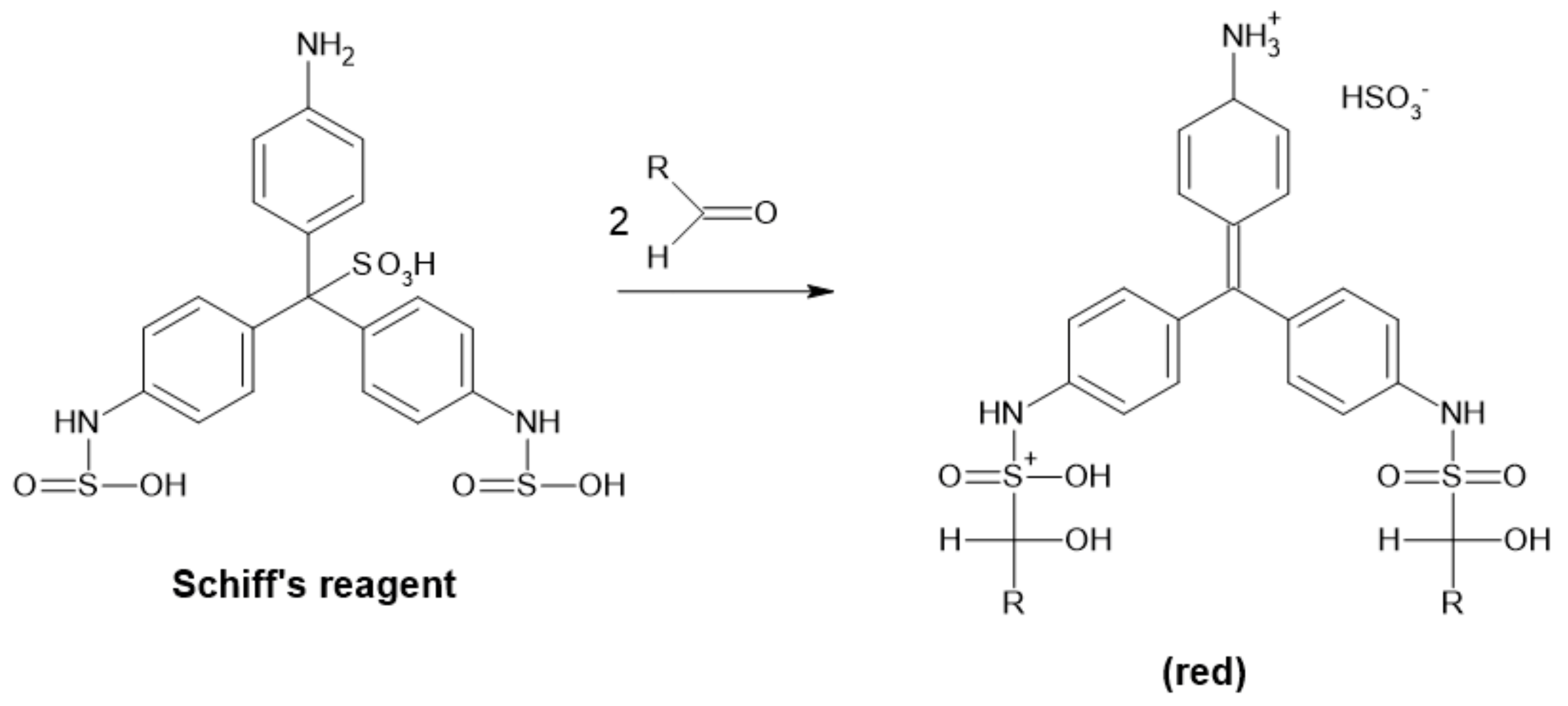
Basic Fuchsin Method
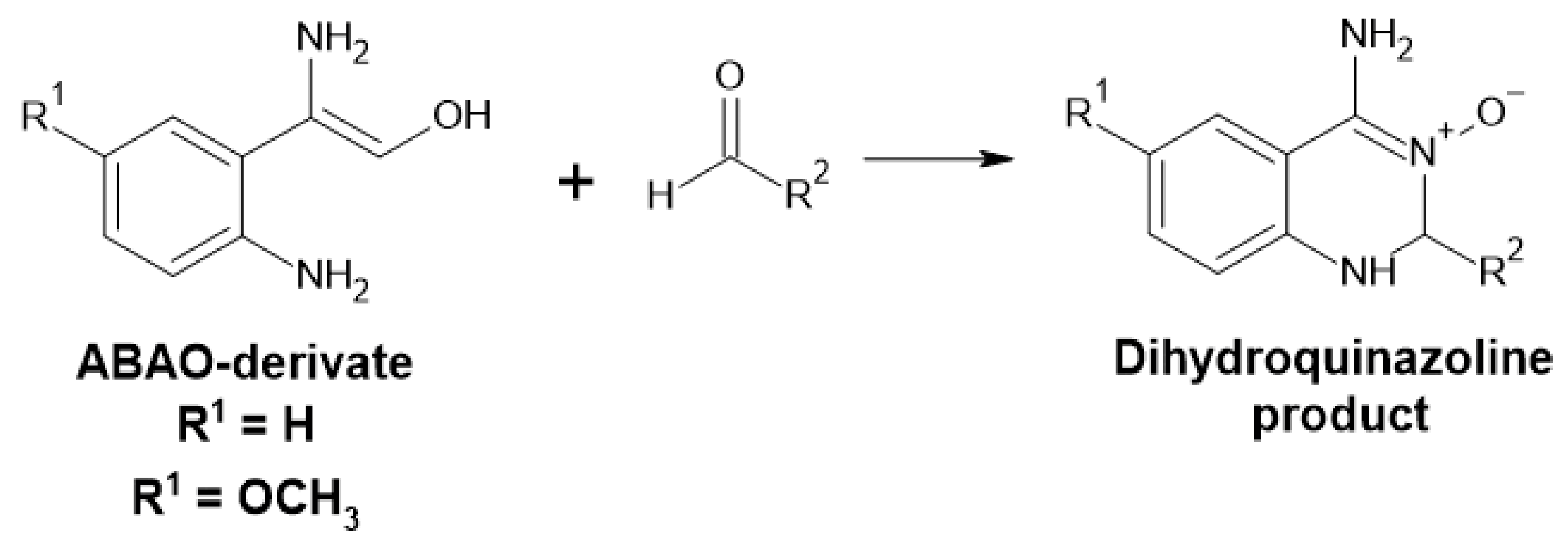
Amino Benzamidoxime Assay
5.1.2. Detection of Other Involved Chemicals
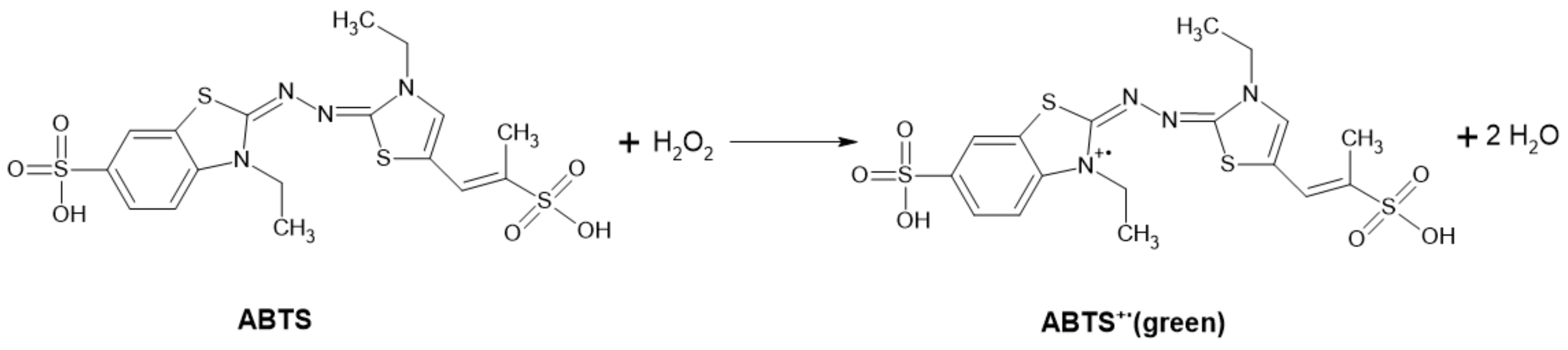
ABTS-POD Method
Disruption of the Conjugated Double Bond System—HPL Reaction
NAD(P)H Spectrophotometric Assay
5.2. Chromatographic Methods
5.3. Biosenors
5.3.1. Ligand-Responsive Transcription Factors
5.3.2. Luciferase-Based Biosensors
6. Products/Applications
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| (h)DAO | (human) Diamine oxidase |
| 13-HPOD | hydroperoxyoctadecadienoic acid |
| 4-HBA | 4-hydrazinobenzoic acid |
| ABTS | 2,2′-azino-bis-(3-ethylbenzothiazoline-6-sulfonic acid) |
| ADH | Alcohol dehydrogenases |
| AHAS | Acetohydroxyacid synthases |
| ALR | aldehyde reductase |
| AMP | Adenosine monophosphate |
| AOs | amine oxidases |
| AOX | Alcohol oxidase |
| ATP | Adenosine triphosphate |
| BLase | bacterial luciferase |
| CAR | Carboxylic acid reductase |
| CoA | coenzyme A |
| DOX | Dioxygenase |
| FAD | flavine adenine dinucleotide |
| GLVs | green leaf volatiles |
| HPL | hydroperoxide lyase |
| IPTG | Isopropyl-β-D-thiogalactopyranoside |
| KID | Ketoisovalerate decarboxylase |
| LOX | Lipoxygenase |
| NAD(P)H | Nicotinamide adenine dinucleotide (phosphate) |
| PCD | Peptidyl carrier domain |
| PDC | Pyruvate decarboxylase |
| Ppant | 4-phosphopantetheine |
| PPTase | Phosphopantetheine transferase |
| (R)-PAC | R-phenylacetylcarbinol |
| TAT | Twin-arginine translocation |
| ThPP | Thiamine pyrophosphate |
| δ-ALA | δ-aminolevulinic acid |
References
- Kunjapur, A.M.; Prather, K.L.J. Microbial Engineering for Aldehyde Synthesis. Appl. Environ. Microbiol. 2015, 81, 1892–1901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winkler, M. Carboxylic Acid Reductase Enzymes (CARs). Curr. Opin. Chem. Biol. 2018, 43, 23–29. [Google Scholar] [CrossRef] [PubMed]
- LoPachin, R.M.; Gavin, T. Molecular Mechanisms of Aldehyde Toxicity: A Chemical Perspective. Chem. Res. Toxicol. 2014, 27, 1081–1091. [Google Scholar] [CrossRef] [PubMed]
- Ricca, E.; Brucher, B.; Schrittwieser, J.H. Multi-Enzymatic Cascade Reactions: Overview and Perspectives. Adv. Synth. Catal. 2011, 353, 2239–2262. [Google Scholar] [CrossRef]
- Bayer, T.; Milker, S.; Wiesinger, T.; Winkler, M.; Mihovilovic, M.D.; Rudroff, F. In Vivo Synthesis of Polyhydroxylated Compounds from a “Hidden Reservoir” of Toxic Aldehyde Species. ChemCatChem 2017, 9, 2919–2923. [Google Scholar] [CrossRef]
- Biewenga, L.; Kunzendorf, A.; Poelarends, G.J. In Situ Acetaldehyde Synthesis for Carboligation Reactions. Chembiochem 2020, 21, 1505–1509. [Google Scholar] [CrossRef]
- Rodriguez, G.M.; Atsumi, S. Toward Aldehyde and Alkane Production by Removing Aldehyde Reductase Activity in Escherichia Coli. Metab. Eng. 2014, 25, 227–237. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, G.; Chattopadhyay, P. Vanillin Biotechnology: The Perspectives and Future. J. Sci. Food Agric. 2019, 99, 499–506. [Google Scholar] [CrossRef]
- Longo, M.A.; Sanromán, M.A. Production of Food Aroma Compounds: Microbial and Enzymatic Methodologies. Food Technol. Biotechnol. 2006, 44, 335–353. [Google Scholar]
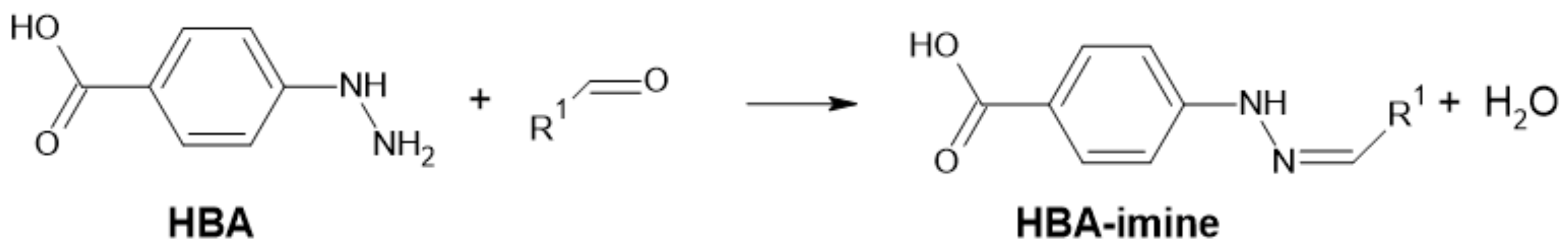
- de Lima, L.F.; Brandão, P.F.; Donegatti, T.A.; Ramos, R.M.; Gonçalves, L.M.; Cardoso, A.A.; Pereira, E.A.; Rodrigues, J.A. 4-Hydrazinobenzoic Acid as a Derivatizing Agent for Aldehyde Analysis by HPLC-UV and CE-DAD. Talanta 2018, 187, 113–119. [Google Scholar] [CrossRef] [Green Version]
- de Carvalho, C.C.C.R. Whole Cell Biocatalysts: Essential Workers from Nature to the Industry. Microb. Biotechnol. 2017, 10, 250–263. [Google Scholar] [CrossRef] [Green Version]
- Pickl, M.; Fuchs, M.; Glueck, S.M.; Faber, K. The Substrate Tolerance of Alcohol Oxidases. Appl. Microbiol. Biotechnol. 2015, 99, 6617–6642. [Google Scholar] [CrossRef] [Green Version]
- Rudroff, F. Whole-Cell Based Synthetic Enzyme Cascades—Light and Shadow of a Promising Technology. Curr. Opin. Chem. Biol. 2019, 49, 84–90. [Google Scholar] [CrossRef]
- Gahloth, D.; Aleku, G.A.; Leys, D. Carboxylic Acid Reductase: Structure and Mechanism. J. Biotechnol. 2020, 307, 107–113. [Google Scholar] [CrossRef]
- Park, J.; Lee, H.-S.; Oh, J.; Joo, J.C.; Yeon, Y.J. A Highly Active Carboxylic Acid Reductase from Mycobacterium Abscessus for Biocatalytic Reduction of Vanillic Acid to Vanillin. Biochem. Eng. J. 2020, 161, 107683. [Google Scholar] [CrossRef]
- He, A.; Li, T.; Daniels, L.; Fotheringham, I.; Rosazza, J.P.N. Nocardia sp. Carboxylic Acid Reductase: Cloning, Expression, and Characterization of a New Aldehyde Oxidoreductase Family. Appl. Environ. Microbiol. 2004, 70, 1874–1881. [Google Scholar] [CrossRef] [Green Version]
- Moura, M.; Pertusi, D.; Lenzini, S.; Bhan, N.; Broadbelt, L.J.; Tyo, K.E.J. Characterizing and Predicting Carboxylic Acid Reductase Activity for Diversifying Bioaldehyde Production. Biotechnol. Bioeng. 2016, 113, 944–952. [Google Scholar] [CrossRef]
- Stolterfoht, H.; Steinkellner, G.; Schwendenwein, D.; Pavkov-Keller, T.; Gruber, K.; Winkler, M. Identification of Key Residues for Enzymatic Carboxylate Reduction. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Venkitasubramanian, P.; Daniels, L.; Rosazza, J.P.N. Reduction of Carboxylic Acids by Nocardia Aldehyde Oxidoreductase Requires a Phosphopantetheinylated Enzyme. J. Biol. Chem. 2007, 282, 478–485. [Google Scholar] [CrossRef] [Green Version]
- Gahloth, D.; Dunstan, M.S.; Quaglia, D.; Klumbys, E.; Lockhart-Cairns, M.P.; Hill, A.M.; Derrington, S.R.; Scrutton, N.S.; Turner, N.J.; Leys, D. Structures of Carboxylic Acid Reductase Reveal Domain Dynamics Underlying Catalysis. Nat. Chem. Biol. 2017, 13, 975–981. [Google Scholar] [CrossRef] [Green Version]
- Finnigan, W.; Thomas, A.; Cromar, H.; Gough, B.; Snajdrova, R.; Adams, J.P.; Littlechild, J.A.; Harmer, N.J. Characterization of Carboxylic Acid Reductases as Enzymes in the Toolbox for Synthetic Chemistry. ChemCatChem 2017, 9, 1005–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosini, E.; Nossa, S.; Valentino, M.; D’Arrigo, P.; Marinesco, S.; Pollegioni, L. Expression of Rat Diamine Oxidase in Escherichia Coli. J. Mol. Catal. B Enzym. 2012, 82, 115–120. [Google Scholar] [CrossRef]
- Gludovacz, E.; Maresch, D.; Bonta, M.; Szöllösi, H.; Furtmüller, P.G.; Weik, R.; Altmann, F.; Limbeck, A.; Borth, N.; Jilma, B.; et al. Characterization of Recombinant Human Diamine Oxidase (RhDAO) Produced in Chinese Hamster Ovary (CHO) Cells. J. Biotechnol. 2016, 227, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Boehm, T.; Pils, S.; Gludovacz, E.; Szoelloesi, H.; Petroczi, K.; Majdic, O.; Quaroni, A.; Borth, N.; Valent, P.; Jilma, B. Quantification of Human Diamine Oxidase. Clin. Biochem. 2017, 50, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Mergemeier, K.; Lehr, M. HPLC-UV Assays for Evaluation of Inhibitors of Mono and Diamine Oxidases Using Novel Phenyltetrazolylalkanamine Substrates. Anal. Biochem. 2018, 549, 29–38. [Google Scholar] [CrossRef] [PubMed]
- McGrath, A.P.; Hilmer, K.M.; Collyer, C.A.; Shepard, E.M.; Elmore, B.O.; Brown, D.E.; Dooley, D.M.; Guss, J.M. Structure and Inhibition of Human Diamine Oxidase. Biochemistry 2009, 48, 9810–9822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartko, J.; Gludovacz, E.; Petroczi, K.; Borth, N.; Jilma, B.; Boehm, T. Recombinant Human Diamine Oxidase Activity Is Not Inhibited by Ethanol, Acetaldehyde, Disulfiram, Diethyldithiocarbamate or Cyanamide. Alcohol 2016, 54, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Elmore, B.O.; Bollinger, J.A.; Dooley, D.M. Human Kidney Diamine Oxidase: Heterologous Expression, Purification, and Characterization. J. Biol. Inorg. Chem. 2002, 7, 565–579. [Google Scholar] [CrossRef]
- Mangkorn, N.; Kanokratana, P.; Roongsawang, N.; Laosiripojana, N.; Champreda, V. Purification, Characterization, and Stabilization of Alcohol Oxidase from Ogataea Thermomethanolica. Protein Expr. Purif. 2018, 150, 26–32. [Google Scholar] [CrossRef]
- Buchholz-Afari, M.I.; Halalipour, A.; Yang, D.; Reyes-De-Corcuera, J.I. Increased Stability of Alcohol Oxidase under High Hydrostatic Pressure. J. Food Eng. 2019, 246, 95–101. [Google Scholar] [CrossRef]
- Venkatesagowda, B.; Dekker, R.F.H. A Rapid Method to Detect and Estimate the Activity of the Enzyme, Alcohol Oxidase by the Use of Two Chemical Complexes—Acetylacetone (3,5-Diacetyl-1,4-Dihydrolutidine) and Acetylacetanilide (3,5-Di-N-Phenylacetyl-1,4-Dihydrolutidine). J. Microbiol. Methods 2019, 158, 71–79. [Google Scholar] [CrossRef]
- Matsumura, K.; Yamada, M.; Yamashita, T.; Muto, H.; Nishiyama, K.-I.; Shimoi, H.; Isobe, K. Expression of Alcohol Oxidase Gene from Ochrobactrum sp. AIU 033 in Recombinant Escherichia Coli through the Twin-Arginine Translocation Pathway. J. Biosci. Bioeng. 2019, 128, 13–21. [Google Scholar] [CrossRef]
- Zhang, L.; Jiang, D.; Li, Y.; Wu, L.; Liu, Q.; Dong, K.; Oger, P. Characterization of a Novel Type III Alcohol Dehydrogenase from Thermococcus Barophilus Ch5. Int. J. Biol. Macromol. 2021, 171, 491–501. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, R.; Xu, Y. Structure-Based Mechanisms: On the Way to Apply Alcohol Dehydrogenases/Reductases to Organic-Aqueous Systems. Int. J. Biol. Macromol. 2021, 168, 412–427. [Google Scholar] [CrossRef]
- Crichton, R. Chapter 12—Zinc—Lewis Acid and Gene Regulator. In Biological Inorganic Chemistry, 3rd ed.; Crichton, R., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 339–362. ISBN 978-0-12-811741-5. [Google Scholar]
- Bornscheuer, U.T.; Huisman, G.W.; Kazlauskas, R.J.; Lutz, S.; Moore, J.C.; Robins, K. Engineering the Third Wave of Biocatalysis. Nature 2012, 485, 185–194. [Google Scholar] [CrossRef]
- Böttcher, D.; Bornscheuer, U.T. Protein Engineering of Microbial Enzymes. Curr. Opin. Microbiol. 2010, 13, 274–282. [Google Scholar] [CrossRef]
- Thompson, M.P.; Turner, N.J. Two-Enzyme Hydrogen-Borrowing Amination of Alcohols Enabled by a Cofactor-Switched Alcohol Dehydrogenase. ChemCatChem 2017, 9, 3833–3836. [Google Scholar] [CrossRef]
- Seo, E.-J.; Kim, H.-J.; Kim, M.-J.; Kim, J.-S.; Park, J.-B. Cofactor Specificity Engineering of a Long-Chain Secondary Alcohol Dehydrogenase from Micrococcus Luteus for Redox-Neutral Biotransformation of Fatty Acids. Chem. Commun. 2019, 55, 14462–14465. [Google Scholar] [CrossRef]
- Liu, J.; Wu, S.; Li, Z. Recent Advances in Enzymatic Oxidation of Alcohols. Curr. Opin. Chem. Biol. 2018, 43, 77–86. [Google Scholar] [CrossRef]
- Wu, S.; Zhou, Y.; Wang, T.; Too, H.-P.; Wang, D.I.C.; Li, Z. Highly Regio- and Enantioselective Multiple Oxy- and Amino-Functionalizations of Alkenes by Modular Cascade Biocatalysis. Nat. Commun. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Klatte, S.; Wendisch, V.F. Redox Self-Sufficient Whole Cell Biotransformation for Amination of Alcohols. Bioorg. Med. Chem. 2014, 22, 5578–5585. [Google Scholar] [CrossRef]
- Kaehne, F.; Buchhaupt, M.; Schrader, J. A Recombinant α-Dioxygenase from Rice to Produce Fatty Aldehydes Using E. Coli. Appl. Microbiol. Biotechnol. 2011, 90, 989–995. [Google Scholar] [CrossRef]
- Cao, Y.-X.; Xiao, W.-H.; Liu, D.; Zhang, J.-L.; Ding, M.-Z.; Yuan, Y.-J. Biosynthesis of Odd-Chain Fatty Alcohols in Escherichia Coli. Metab. Eng. 2015, 29, 113–123. [Google Scholar] [CrossRef]
- Buchhaupt, M.; Kähne, F.; Etschmann, M.M.W.; Schrader, J. Chapter 37—Biotechnological Production of Fatty Aldehydes. In Flavour Science; Ferreira, V., Lopez, R., Eds.; Academic Press: San Diego, CA, USA, 2014; pp. 195–199. ISBN 978-0-12-398549-1. [Google Scholar]
- Schaffer, S.; Gielen, J.; Wessel, M.; Hennemann, H.-G.; Haeger, H.; Haas, T.; Bluemke, W. Production of Omega-Amino Fatty Acids. U.S. Patent US20140178948A1, 26 July 2014. [Google Scholar]
- Somboon, K.; Doble, A.; Bulmer, D.; Baslé, A.; Khalid, S.; van den Berg, B. Uptake of Monoaromatic Hydrocarbons during Biodegradation by FadL Channel-Mediated Lateral Diffusion. Nat. Commun. 2020, 11, 6331. [Google Scholar] [CrossRef] [PubMed]
- Açar, Ş.; Yücel, M.; Hamamcı, H. Purification and Characterisation of Two Isozymes of Pyruvate Decarboxylase from Rhizopus Oryzae. Enzyme Microb. Technol. 2007, 40, 675–682. [Google Scholar] [CrossRef]
- Raj, K.C.; Talarico, L.A.; Ingram, L.O.; Maupin-Furlow, J.A. Cloning and Characterization of the Zymobacter Palmae Pyruvate Decarboxylase Gene (Pdc) and Comparison to Bacterial Homologues. Appl. Environ. Microbiol. 2002, 68, 2869–2876. [Google Scholar] [CrossRef] [Green Version]
- Bell, P.; Hoyt, K.; Shabangi, M. The Electrochemical Investigation of the Catalytic Power of Pyruvate Decarboxylase and Its Coenzyme. Bioelectrochemistry 2006, 68, 171–174. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, L.; Boo, K.H.; Park, E.; Drakakaki, G.; Zakharov, F. PDC1, a Pyruvate/α-Ketoacid Decarboxylase, Is Involved in Acetaldehyde, Propanal and Pentanal Biosynthesis in Melon (Cucumis Melo L.) Fruit. Plant J. 2019, 98, 112–125. [Google Scholar] [CrossRef] [PubMed]
- van Zyl, L.J.; Schubert, W.-D.; Tuffin, M.I.; Cowan, D.A. Structure and Functional Characterization of Pyruvate Decarboxylase from Gluconacetobacter Diazotrophicus. BMC Struct. Biol. 2014, 14, 21. [Google Scholar] [CrossRef] [Green Version]
- Sehl, T.; Walter, L.; Menyes, U.; Bock, S.; Marx, L.; Westphal, R.; Maugeri, Z.; Pohl, M.; Vogel, C.; Erhardt, M.; et al. Asymmetric Synthesis of (S)-Phenylacetylcarbinol—Closing a Gap in C–C Bond Formation; RSC: Berlin, Germany, 2017; Volume 19, p. 380. [Google Scholar]
- Sanchis, V.; Vinas, I.; Roberts, I.N.; Jeenes, D.J.; Watson, A.J.; Archer, D.B. A Pyruvate Decarboxylase Gene from Aspergillus Parasiticus. FEMS Microbiol. Lett. 1994, 117, 207–210. [Google Scholar] [CrossRef]
- Eram, M.S.; Ma, K. Pyruvate Decarboxylase Activity of the Acetohydroxyacid Synthase of Thermotoga Maritima. Biochem. Biophys. Rep. 2016, 7, 394–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarado, O.; García-Meseguer, R.; Javier Ruiz-Pernía, J.; Tuñon, I.; Delgado, E.J. A Molecular Dynamics Study on the Role of the Protonation State in the Biosynthesis of R-PAC by AHAS. Chem. Phys. Lett. 2019, 716, 247–251. [Google Scholar] [CrossRef] [Green Version]
- Kheyrandish, M.; Doostmohammadi, M. Bioinformatics Analysis of Aceto Hydroxyl Acid Synthase as an Alternative Enzyme for Pyruvate Decarboxylase. J. Chem. Pharm. Res. 2018, 10, 71–74. [Google Scholar]
- Kunjapur, A.M.; Tarasova, Y.; Prather, K.L.J. Synthesis and Accumulation of Aromatic Aldehydes in an Engineered Strain of Escherichia Coli. J. Am. Chem. Soc. 2014, 136, 11644–11654. [Google Scholar] [CrossRef]
- Wu, R.-J.; Ren, T.; Gao, J.-Y.; Wang, L.; Yu, Q.; Yao, Z.; Song, G.-Q.; Ruan, W.-B.; Niu, C.-W.; Song, F.-H.; et al. Chemical Preparation, Biological Evaluation and 3D-QSAR of Ethoxysulfuron Derivatives as Novel Antifungal Agents Targeting Acetohydroxyacid Synthase. Eur. J. Med. Chem. 2019, 162, 348–363. [Google Scholar] [CrossRef]
- de la Plaza, M.; Fernández de Palencia, P.; Peláez, C.; Requena, T. Biochemical and Molecular Characterization of α-Ketoisovalerate Decarboxylase, an Enzyme Involved in the Formation of Aldehydes from Amino Acids by Lactococcus Lactis. FEMS Microbiol. Lett. 2004, 238, 367–374. [Google Scholar] [CrossRef] [Green Version]
- Miao, R.; Xie, H.; Ho, F.M.; Lindblad, P. Protein Engineering of α-Ketoisovalerate Decarboxylase for Improved Isobutanol Production in Synechocystis PCC 6803. Metab. Eng. 2018, 47, 42–48. [Google Scholar] [CrossRef]
- Sigrist, R.; da Costa, B.Z.; Marsaioli, A.J.; de Oliveira, L.G. Nature-Inspired Enzymatic Cascades to Build Valuable Compounds. Biotechnol. Adv. 2015, 33, 394–411. [Google Scholar] [CrossRef]
- France, S.P.; Hepworth, L.J.; Turner, N.J.; Flitsch, S.L. Constructing Biocatalytic Cascades: In Vitro and In Vivo Approaches to de Novo Multi-Enzyme Pathways. ACS Catal. 2017, 7, 710–724. [Google Scholar] [CrossRef]
- Wu, S.; Snajdrova, R.; Moore, J.C.; Baldenius, K.; Bornscheuer, U.T. Biocatalysis: Enzymatic Synthesis for Industrial Applications. Angewandte Chem. Int. Ed. 2020, 60, 88–119. [Google Scholar] [CrossRef]
- Noordermeer, M.A.; van der Goot, W.; van Kooij, A.J.; Veldsink, J.W.; Veldink, G.A.; Vliegenthart, J.F.G. Development of a Biocatalytic Process for the Production of C6-Aldehydes from Vegetable Oils by Soybean Lipoxygenase and Recombinant Hydroperoxide Lyase. J. Agric. Food Chem. 2002, 50, 4270–4274. [Google Scholar] [CrossRef] [Green Version]
- ul Hassan, M.N.; Zainal, Z.; Ismail, I. Green Leaf Volatiles: Biosynthesis, Biological Functions and Their Applications in Biotechnology. Plant Biotechnol. J. 2015, 13, 727–739. [Google Scholar] [CrossRef]
- Sánchez-Ortiz, A.; Bejaoui, M.A.; Quintero-Flores, A.; Jiménez, A.; Beltrán, G. Biosynthesis of Volatile Compounds by Hydroperoxide Lyase Enzymatic Activity during Virgin Olive Oil Extraction Process. Food Res. Int. 2018, 111, 220–228. [Google Scholar] [CrossRef]
- Robinson, D.S.; Wu, Z.; Domoney, C.; Casey, R. Lipoxygenases and the Quality of Foods. Food Chem. 1995, 54, 33–43. [Google Scholar] [CrossRef]
- Hornung, E.; Walther, M.; Kühn, H.; Feussner, I. Conversion of Cucumber Linoleate 13-Lipoxygenase to a 9-Lipoxygenating Species by Site-Directed Mutagenesis. Proc. Natl. Acad. Sci. USA 1999, 96, 4192–4197. [Google Scholar] [CrossRef] [Green Version]
- Brühlmann, F.; Bosijokovic, B.; Ullmann, C.; Auffray, P.; Fourage, L.; Wahler, D. Directed Evolution of a 13-Hydroperoxide Lyase (CYP74B) for Improved Process Performance. J. Biotechnol. 2013, 163, 339–345. [Google Scholar] [CrossRef]
- Matsui, K. Green Leaf Volatiles: Hydroperoxide Lyase Pathway of Oxylipin Metabolism. Curr. Opin. Plant Biol. 2006, 9, 274–280. [Google Scholar] [CrossRef]
- Shaaban, H.; Mahmoud, K.; Amin, A.; Banna, H. Application of Biotechnology to the Production of Natural Flavor and Fragrance Chemicals. Res. J. Pharm. Biol. Chem. Sci. 2016, 7, 2670–2717. [Google Scholar]
- Galliard, T.; Phillips, D.R.; Reynolds, J. The Formation of Cis-3-Nonenal, Trans-2-Nonenal and Hexanal from Linoleic Acid Hydroperoxide Isomers by a Hydroperoxide Cleavage Enzyme System in Cucumber (Cucumis Sativus) Fruits. Biochim. Biophys. Acta (BBA) Lipids Lipid Metab. 1976, 441, 181–192. [Google Scholar] [CrossRef]
- Liu, Q.; Hua, Y. Continuous Synthesis of Hexanal by Immobilized Hydroperoxide Lyase in Packed-Bed Reactor. Bioprocess Biosyst. Eng. 2015, 38, 2439–2449. [Google Scholar] [CrossRef]
- Vincenti, S.; Mariani, M.; Alberti, J.-C.; Jacopini, S.; Brunini-Bronzini de Caraffa, V.; Berti, L.; Maury, J. Biocatalytic Synthesis of Natural Green Leaf Volatiles Using the Lipoxygenase Metabolic Pathway. Catalysts 2019, 9, 873. [Google Scholar] [CrossRef] [Green Version]
- Bayer, T.; Milker, S.; Wiesinger, T.; Rudroff, F.; Mihovilovic, M.D. Designer Microorganisms for Optimized Redox Cascade Reactions—Challenges and Future Perspectives. Adv. Synth. Catal. 2015, 357, 1587–1618. [Google Scholar] [CrossRef]
- Wachtmeister, J.; Rother, D. Recent Advances in Whole Cell Biocatalysis Techniques Bridging from Investigative to Industrial Scale. Curr. Opin. Biotechnol. 2016, 42, 169–177. [Google Scholar] [CrossRef]
- Lin, B.; Tao, Y. Whole-Cell Biocatalysts by Design. Microb. Cell Fact. 2017, 16, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polakovič, M.; Švitel, J.; Bučko, M.; Filip, J.; Neděla, V.; Ansorge-Schumacher, M.B.; Gemeiner, P. Progress in Biocatalysis with Immobilized Viable Whole Cells: Systems Development, Reaction Engineering and Applications. Biotechnol. Lett. 2017, 39, 667–683. [Google Scholar] [CrossRef]
- Garzón-Posse, F.; Becerra-Figueroa, L.; Hernández-Arias, J.; Gamba-Sánchez, D. Whole Cells as Biocatalysts in Organic Transformations. Molecules 2018, 23, 1265. [Google Scholar] [CrossRef] [Green Version]
- Hauer, B. Embracing Nature’s Catalysts: A Viewpoint on the Future of Biocatalysis. ACS Catal. 2020, 10, 8418–8427. [Google Scholar] [CrossRef]
- Balagurunathan, B.; Tan, L.; Zhao, H. Metabolic Engineering of Escherichia Coli for Acetaldehyde Overproduction Using Pyruvate Decarboxylase from Zymomonas Mobilis. Enzyme Microb. Technol. 2018, 109, 58–65. [Google Scholar] [CrossRef]
- Welch, M.; Govindarajan, S.; Ness, J.E.; Villalobos, A.; Gurney, A.; Minshull, J.; Gustafsson, C. Design Parameters to Control Synthetic Gene Expression in Escherichia Coli. PLoS ONE 2009, 4, e7002. [Google Scholar] [CrossRef]
- Sudhamsu, J.; Kabir, M.; Airola, M.V.; Patel, B.A.; Yeh, S.-R.; Rousseau, D.L.; Crane, B.R. Co-Expression of Ferrochelatase Allows for Complete Heme Incorporation into Recombinant Proteins Produced in E. Coli. Protein Expr. Purif. 2010, 73, 78–82. [Google Scholar] [CrossRef] [Green Version]
- Tijet, N.; Wäspi, U.; Gaskin, D.J.; Hunziker, P.; Muller, B.L.; Vulfson, E.N.; Slusarenko, A.; Brash, A.R.; Whitehead, I.M. Purification, Molecular Cloning, and Expression of the Gene Encoding Fatty Acid 13-Hydroperoxide Lyase from Guava Fruit (Psidium Guajava). Lipids 2000, 35, 709–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horvat, M.; Winkler, M. In Vivo Reduction of Medium- to Long-Chain Fatty Acids by Carboxylic Acid Reductase (CAR) Enzymes: Limitations and Solutions. ChemCatChem 2020, 12, 5076–5090. [Google Scholar] [CrossRef]
- Han, L.; Liang, B. New Approaches to NAD(P)H Regeneration in the Biosynthesis Systems. World J. Microbiol. Biotechnol. 2018, 34, 141. [Google Scholar] [CrossRef] [PubMed]
- Schroer, K.; Peter Luef, K.; Stefan Hartner, F.; Glieder, A.; Pscheidt, B. Engineering the Pichia Pastoris Methanol Oxidation Pathway for Improved NADH Regeneration during Whole-Cell Biotransformation. Metab. Eng. 2010, 12, 8–17. [Google Scholar] [CrossRef]
- Wenk, S.; Schann, K.; He, H.; Rainaldi, V.; Kim, S.; Lindner, S.N.; Bar-Even, A. An “Energy-Auxotroph” Escherichia Coli Provides an in Vivo Platform for Assessing NADH Regeneration Systems. Biotechnol. Bioeng. 2020, 117, 3422–3434. [Google Scholar] [CrossRef]
- Lindner, S.N.; Ramirez, L.C.; Krüsemann, J.L.; Yishai, O.; Belkhelfa, S.; He, H.; Bouzon, M.; Döring, V.; Bar-Even, A. NADPH-Auxotrophic E. Coli: A Sensor Strain for Testing in Vivo Regeneration of NADPH. ACS Synth. Biol. 2018, 7, 2742–2749. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Huang, Z.; Du, C.; Li, Y.; Cao, Z. Introduction of an NADH Regeneration System into Klebsiella Oxytoca Leads to an Enhanced Oxidative and Reductive Metabolism of Glycerol. Metab. Eng. 2009, 11, 101–106. [Google Scholar] [CrossRef]
- Samuel, N.; Bao, T.; Zhang, X.; Yang, T.; Xu, M.; Li, X.; Komera, I.; Philibert, T.; Rao, Z. Optimized Whole Cell Biocatalyst from Acetoin to 2,3-Butanediol through Coexpression of Acetoin Reductase with NADH Regeneration Systems in Engineered Bacillus Subtilis. J. Chem. Technol. Biotechnol. 2017, 92, 2477–2487. [Google Scholar] [CrossRef]
- Strohmeier, G.A.; Eiteljörg, I.C.; Schwarz, A.; Winkler, M. Enzymatic One-Step Reduction of Carboxylates to Aldehydes with Cell-Free Regeneration of ATP and NADPH. Chem. Eur. J. 2019, 25, 6119–6123. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.-C.; Jiang, C.-M.; Ho, Y.-Y.; Shen, S.-C.; Chang, H.-M. Convenient Quantification of Methanol in Juices by Methanol Oxidase in Combination with Basic Fuchsin. Food Chem. 2007, 100, 412–418. [Google Scholar] [CrossRef]
- Schwendenwein, D.; Ressmann, A.K.; Doerr, M.; Höhne, M.; Bornscheuer, U.T.; Mihovilovic, M.D.; Rudroff, F.; Winkler, M. Random Mutagenesis-Driven Improvement of Carboxylate Reductase Activity Using an Amino Benzamidoxime-Mediated High-Throughput Assay. Adv. Synth. Catal. 2019, 361, 2544–2549. [Google Scholar] [CrossRef] [Green Version]
- Ressmann, A.K.; Schwendenwein, D.; Leonhartsberger, S.; Mihovilovic, M.D.; Bornscheuer, U.T.; Winkler, M.; Rudroff, F. Substrate-Independent High-Throughput Assay for the Quantification of Aldehydes. Adv. Synth. Catal. 2019, 361, 2538–2543. [Google Scholar] [CrossRef] [Green Version]
- Cai, H.; Liu, X.; Zou, J.; Xiao, J.; Yuan, B.; Li, F.; Cheng, Q. Multi-Wavelength Spectrophotometric Determination of Hydrogen Peroxide in Water with Peroxidase-Catalyzed Oxidation of ABTS. Chemosphere 2018, 193, 833–839. [Google Scholar] [CrossRef]
- Mu, W.; Xue, Q.; Jiang, B.; Hua, Y. Molecular Cloning, Expression, and Enzymatic Characterization of Solanum Tuberosum Hydroperoxide Lyase. Eur. Food Res. Technol. 2012, 234, 723–731. [Google Scholar] [CrossRef]
- Frazão, C.R.; Maton, V.; François, J.M.; Walther, T. Development of a Metabolite Sensor for High-Throughput Detection of Aldehydes in Escherichia Coli. Front. Bioeng. Biotechnol. 2018, 6. [Google Scholar] [CrossRef]
- Kunjapur, A.M.; Prather, K.L.J. Development of a Vanillate Biosensor for the Vanillin Biosynthesis Pathway in E. Coli. ACS Synth. Biol. 2019, 8, 1958–1967. [Google Scholar] [CrossRef]
- Santala, S.; Efimova, E.; Karp, M.; Santala, V. Real-Time Monitoring of Intracellular Wax Ester Metabolism. Microb. Cell Fact. 2011, 10, 75. [Google Scholar] [CrossRef] [Green Version]
- Lehtinen, T.; Efimova, E.; Santala, S.; Santala, V. Improved Fatty Aldehyde and Wax Ester Production by Overexpression of Fatty Acyl-CoA Reductases. Microb. Cell Fact. 2018, 17, 19. [Google Scholar] [CrossRef] [Green Version]
- Lehtinen, T.; Santala, V.; Santala, S. Twin-Layer Biosensor for Real-Time Monitoring of Alkane Metabolism. FEMS Microbiol. Lett. 2017, 364. [Google Scholar] [CrossRef]
- Vong, W.C.; Liu, S.-Q. Bioconversion of Green Volatiles in Okara (Soybean Residue) into Esters by Coupling Enzyme Catalysis and Yeast (Lindnera Saturnus) Fermentation. Appl. Microbiol. Biotechnol. 2018, 102, 10017–10026. [Google Scholar] [CrossRef]

















Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kazimírová, V.; Rebroš, M. Production of Aldehydes by Biocatalysis. Int. J. Mol. Sci. 2021, 22, 4949. https://doi.org/10.3390/ijms22094949
Kazimírová V, Rebroš M. Production of Aldehydes by Biocatalysis. International Journal of Molecular Sciences. 2021; 22(9):4949. https://doi.org/10.3390/ijms22094949
Chicago/Turabian StyleKazimírová, Veronika, and Martin Rebroš. 2021. "Production of Aldehydes by Biocatalysis" International Journal of Molecular Sciences 22, no. 9: 4949. https://doi.org/10.3390/ijms22094949
APA StyleKazimírová, V., & Rebroš, M. (2021). Production of Aldehydes by Biocatalysis. International Journal of Molecular Sciences, 22(9), 4949. https://doi.org/10.3390/ijms22094949