
Resveratrol-Encapsulated Mitochondria-Targeting Liposome Enhances Mitochondrial Respiratory Capacity in Myocardial Cells

 ,
,
Abstract
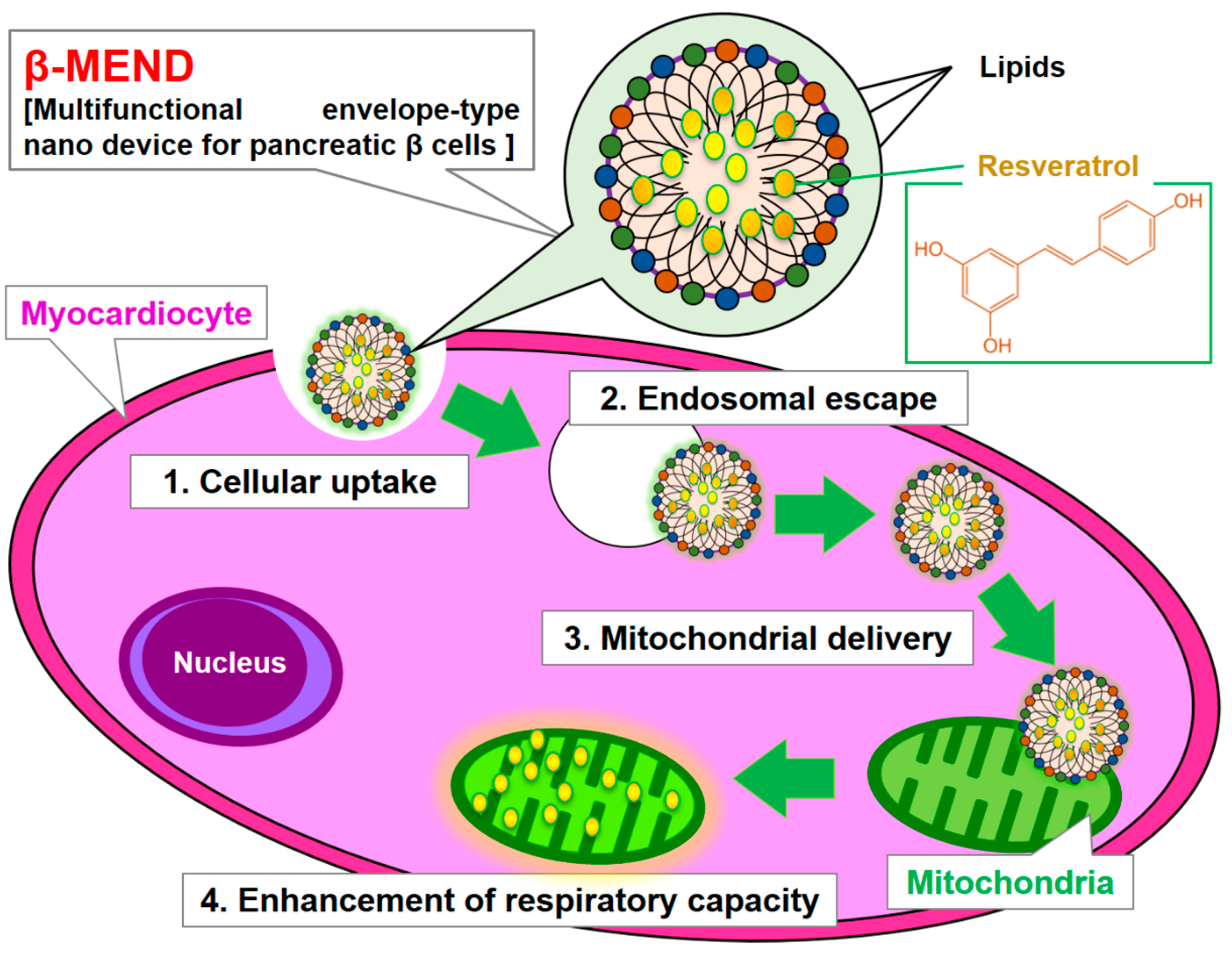
:1. Introduction
2. Results
2.1. Property of LPs
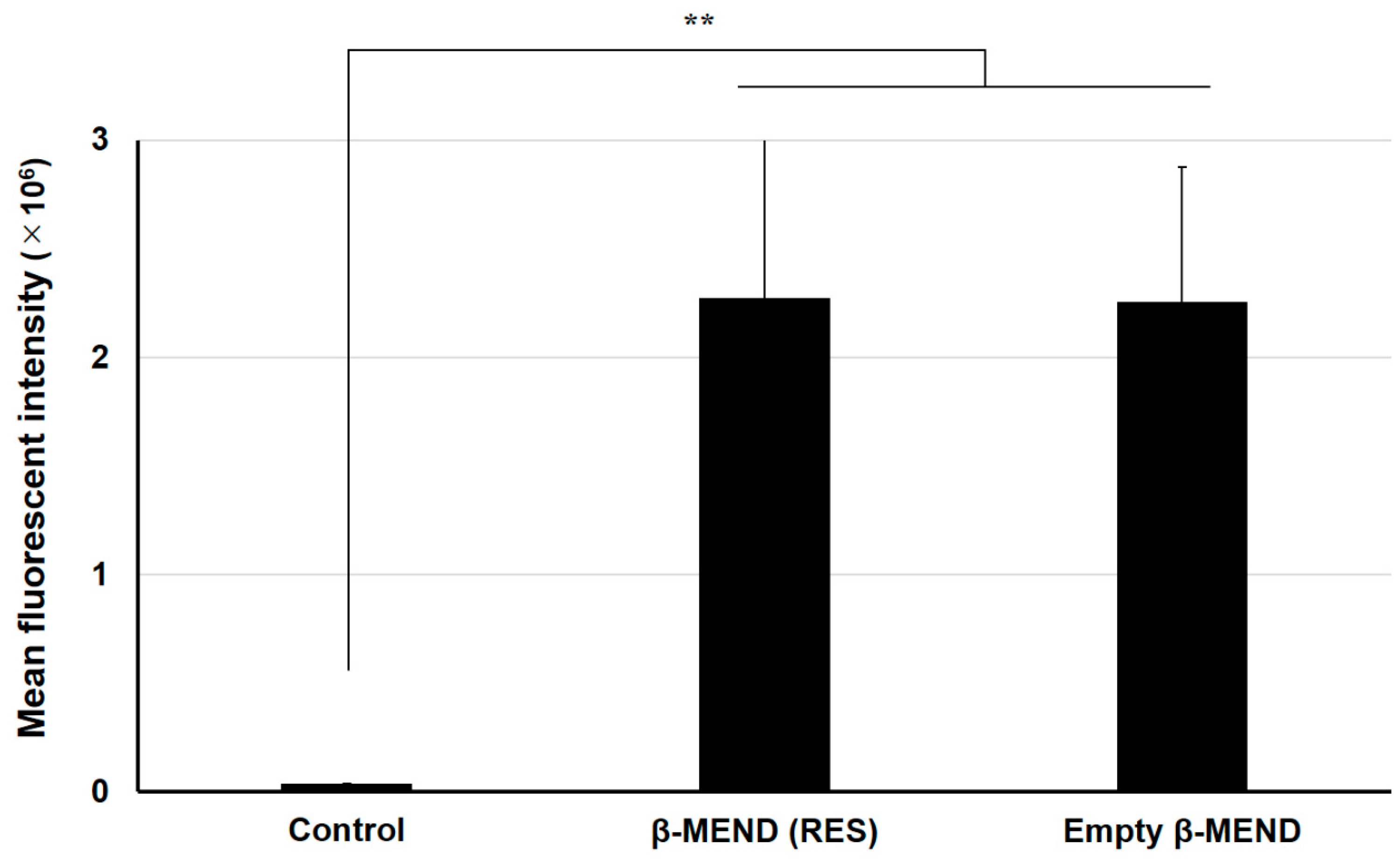
2.2. Evaluations of LP Uptake by Fluorescence-Activated Cell Sorting (FACS) Analyses
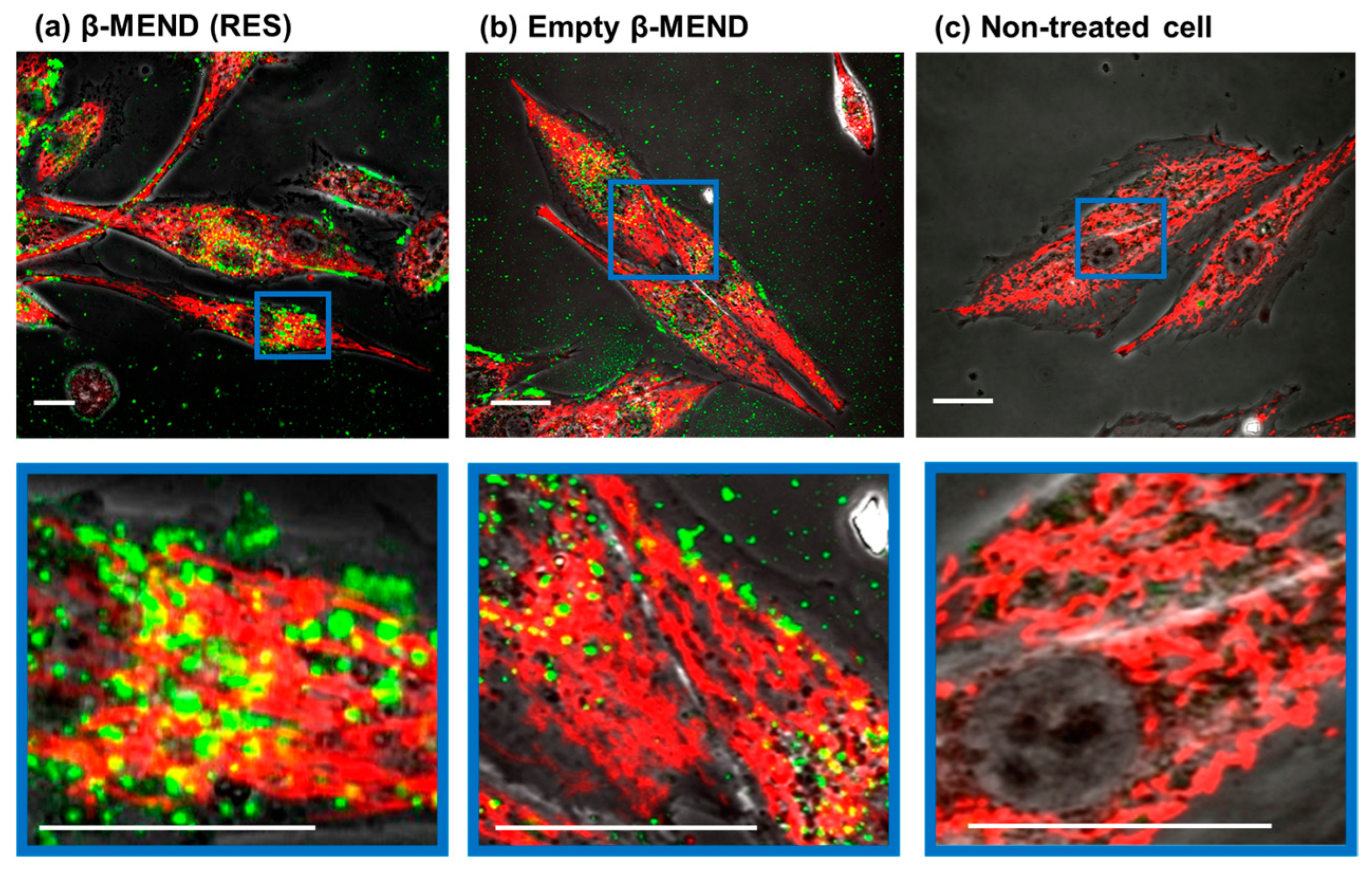
2.3. Evaluation of LP Uptake by Confocal Laser Scanning Microscopic (CLSM) Observations
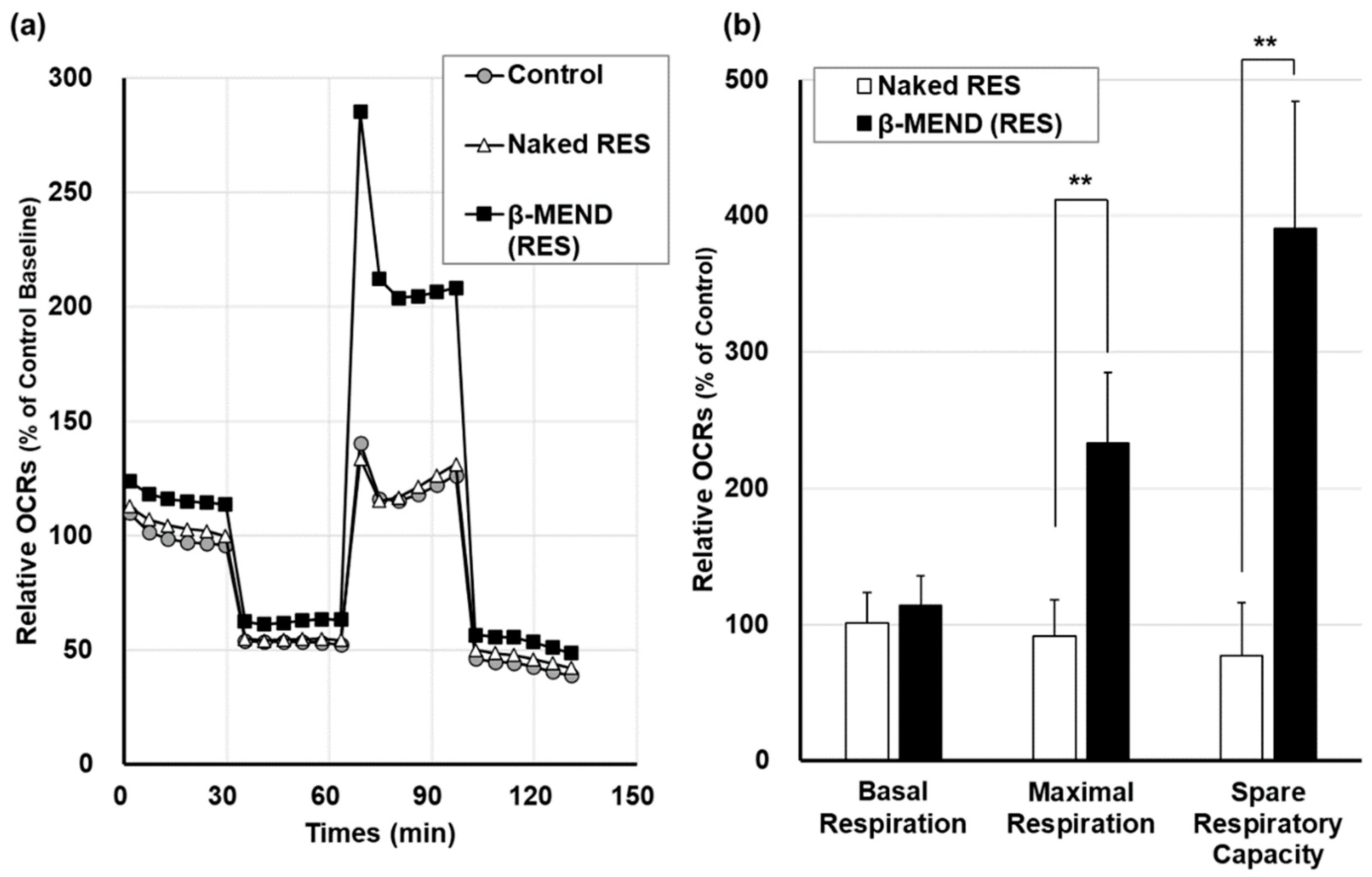
2.4. Measurement of Mitochondrial Respiratory Function
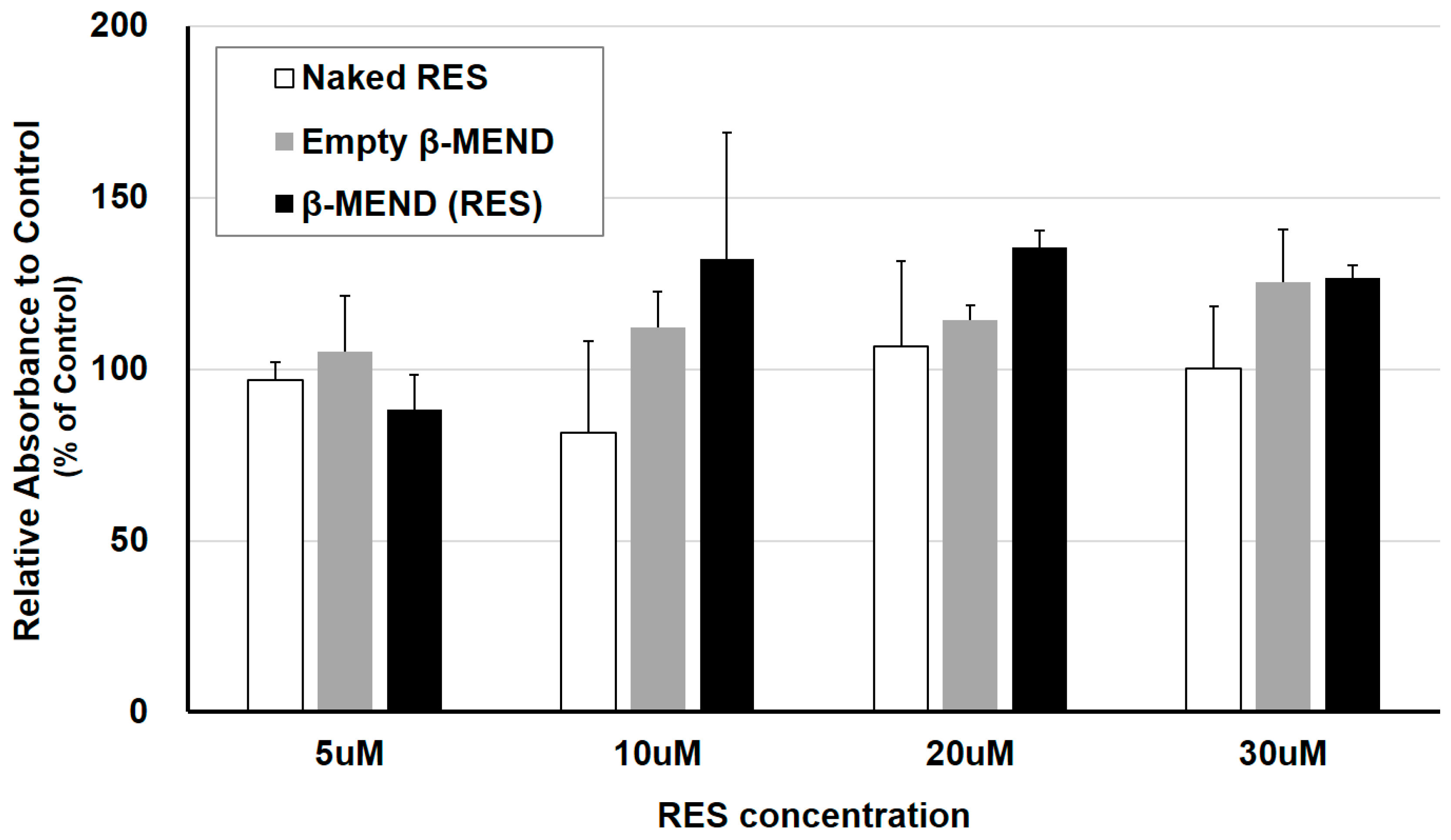
2.5. Cytotoxicity of the LPs
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Preparation of LPs
4.3. Cell Lines and Culturing
4.4. FACS Analyses of LP Uptake
4.5. CLSM Observation of LP Uptake
4.6. Analyses of Mitochondrial Respiratory Function
4.7. Evaluation of Cytotoxicity of LPs
4.8. Statistical Analyses
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pala, R.; Anju, V.T.; Dyavaiah, M.; Busi, S.; Nauli, S.M. Nanoparticle-Mediated Drug Delivery for the Treatment of Cardiovascular Diseases. Int. J. Nanomed. 2020, 15, 3741–3769. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Joshi, J.; Li, F.; Xu, B.; Khan, M.; Yang, J.; Zhu, W. Nanoparticle-Mediated Drug Delivery for Treatment of Ischemic Heart Disease. Front. Bioeng. Biotechnol. 2020, 8, 687. [Google Scholar] [CrossRef] [PubMed]
- Saludas, L.; Pascual-Gil, S.; Roli, F.; Garbayo, E.; Blanco-Prieto, M.J. Heart tissue repair and cardioprotection using drug delivery systems. Maturitas 2018, 110, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hausenloy, D.J.; Yellon, D.M. Myocardial ischemia-reperfusion injury: A neglected therapeutic target. J. Clin. Investig. 2013, 123, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.X.; Cheng, Y.; Liu, D.Z.; Liu, M.; Cui, H.; Zhang, B.L.; Mei, Q.B.; Zhou, S.Y. Mitochondria-targeted cyclosporin A delivery system to treat myocardial ischemia reperfusion injury of rats. J. Nanobiotechnol. 2019, 17, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.; Liu, D.Z.; Zhang, C.X.; Cui, H.; Liu, M.; Zhang, B.L.; Mei, Q.B.; Lu, Z.F.; Zhou, S.Y. Mitochondria-targeted antioxidant delivery for precise treatment of myocardial ischemia-reperfusion injury through a multistage continuous targeted strategy. Nanomedicine 2019, 16, 236–249. [Google Scholar] [CrossRef]
- Yamada, Y.; Akita, H.; Kamiya, H.; Kogure, K.; Yamamoto, T.; Shinohara, Y.; Yamashita, K.; Kobayashi, H.; Kikuchi, H.; Harashima, H. MITO-Porter: A liposome-based carrier system for delivery of macromolecules into mitochondria via membrane fusion. Biochim. Et Biophys. Acta (BBA) Biomembr. 2008, 1778, 423–432. [Google Scholar] [CrossRef]
- Yamada, Y.; Hibino, M.; Sasaki, D.; Abe, J.; Harashima, H. Power of mitochondrial drug delivery systems to produce innovative nanomedicines. Adv. Drug Deliv. Rev. 2020, 154–155, 187–209. [Google Scholar] [CrossRef]
- Yamada, Y.; Somiya, K.; Miyauchi, A.; Osaka, H.; Harashima, H. Validation of a mitochondrial RNA therapeutic strategy using fibroblasts from a Leigh syndrome patient with a mutation in the mitochondrial ND3 gene. Sci. Rep. 2020, 10, 7511. [Google Scholar] [CrossRef]
- Takano, Y.; Hirata, E.; Ushijima, N.; Harashima, H.; Yamada, Y. The effective In Vivo mitochondrial-targeting nanocarrier combined with a π-extended porphyrin-type photosensitizer. Nanoscale Adv. 2021, 3, 5919–5927. [Google Scholar] [CrossRef]
- Yamada, Y.; Tabata, M.; Yasuzaki, Y.; Nomura, M.; Shibata, A.; Ibayashi, Y.; Taniguchi, Y.; Sasaki, S.; Harashima, H. A nanocarrier system for the delivery of nucleic acids targeted to a pancreatic beta cell line. Biomaterials 2014, 35, 6430–6438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, Y.; Fujishita, N.; Harashima, H. A nanocarrier for the mitochondrial delivery of nucleic acids to cardiomyocytes. Nucleosides Nucleotides Nucleic Acids 2020, 39, 141–155. [Google Scholar] [CrossRef]
- Machado, N.D.; Fernandez, M.A.; Diaz, D.D. Recent Strategies in Resveratrol Delivery Systems. Chempluschem 2019, 84, 951–973. [Google Scholar] [CrossRef]
- Bresciani, L.; Calani, L.; Bocchi, L.; Delucchi, F.; Savi, M.; Ray, S.; Brighenti, F.; Stilli, D.; Del Rio, D. Bioaccumulation of resveratrol metabolites in myocardial tissue is dose-time dependent and related to cardiac hemodynamics in diabetic rats. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Sutanto, H.; Dobrev, D.; Heijman, J. Resveratrol: An effective pharmacological agent to prevent inflammation-induced atrial fibrillation? Naunyn. Schmiedebergs Arch. Pharm. 2018, 391, 1163–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, T.T.; Wang, X.Y.; Huang, J.; Deng, Y.Z.; Qiu, L.J.; Liu, H.Y.; Xu, X.W.; Ma, Z.X.; Tang, L.; Chen, H.P. Mitochondrial Translocation of DJ-1 Is Mediated by Grp75: Implication in Cardioprotection of Resveratrol Against Hypoxia/Reoxygenation-Induced Oxidative Stress. J. Cardiovasc. Pharmacol. 2020, 75, 305–313. [Google Scholar] [CrossRef]
- Schwartz, D.R.; Sack, M.N. Targeting the mitochondria to augment myocardial protection. Curr. Opin. Pharmacol. 2008, 8, 160–165. [Google Scholar] [CrossRef] [Green Version]
- Abe, J.; Yamada, Y.; Takeda, A.; Harashima, H. Cardiac progenitor cells activated by mitochondrial delivery of resveratrol enhance the survival of a doxorubicin-induced cardiomyopathy mouse model via the mitochondrial activation of a damaged myocardium. J. Control. Release 2018, 269, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Dolinsky, V.W.; Rogan, K.J.; Sung, M.M.; Zordoky, B.N.; Haykowsky, M.J.; Young, M.E.; Jones, L.W.; Dyck, J.R. Both aerobic exercise and resveratrol supplementation attenuate doxorubicin-induced cardiac injury in mice. Am. J. Physiol.-Endocrinol. Metab. 2013, 305, E243–E253. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Zhu, K.; Zeng, H.; Zhang, J.; Pu, Y.; Wang, Z.; Zhang, T.; Wang, B. Resveratrol solid lipid nanoparticles to trigger credible inhibition of doxorubicin cardiotoxicity. Int. J. Nanomed. 2019, 14, 6061–6071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alanazi, A.M.; Fadda, L.; Alhusaini, A.; Ahmad, R.; Hasan, I.H.; Mahmoud, A.M. Liposomal Resveratrol and/or Carvedilol Attenuate Doxorubicin-Induced Cardiotoxicity by Modulating Inflammation, Oxidative Stress and S100A1 in Rats. Antioxidants 2020, 9, 159. [Google Scholar] [CrossRef] [Green Version]
- Alanazi, A.; Fadda, L.; Alhusaini, A.; Ahmad, R. Antioxidant, antiapoptotic, and antifibrotic effects of the combination of liposomal resveratrol and carvedilol against doxorubicin-induced cardiomyopathy in rats. J. Biochem. Mol. Toxicol. 2020, 34, e22492. [Google Scholar] [CrossRef]
- Cheung, K.G.; Cole, L.K.; Xiang, B.; Chen, K.; Ma, X.; Myal, Y.; Hatch, G.M.; Tong, Q.; Dolinsky, V.W. Sirtuin-3 (SIRT3) Protein Attenuates Doxorubicin-induced Oxidative Stress and Improves Mitochondrial Respiration in H9c2 Cardiomyocytes. J. Biol. Chem. 2015, 290, 10981–10993. [Google Scholar] [CrossRef] [Green Version]
- Wen, J.; Wang, J.; Li, P.; Wang, R.; Wang, J.; Zhou, X.; Zhang, L.; Li, H.; Wei, S.; Cai, H.; et al. Protective effects of higenamine combined with [6]-gingerol against doxorubicin-induced mitochondrial dysfunction and toxicity in H9c2 cells and potential mechanisms. Biomed. Pharm. 2019, 115, 108881. [Google Scholar] [CrossRef]
- Hoes, M.F.; Grote Beverborg, N.; Kijlstra, J.D.; Kuipers, J.; Swinkels, D.W.; Giepmans, B.N.G.; Rodenburg, R.J.; van Veldhuisen, D.J.; de Boer, R.A.; van der Meer, P. Iron deficiency impairs contractility of human cardiomyocytes through decreased mitochondrial function. Eur. J. Heart Fail. 2018, 20, 910–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali Pour, P.; Kenney, M.C.; Kheradvar, A. Bioenergetics Consequences of Mitochondrial Transplantation in Cardiomyocytes. J. Am. Heart Assoc. 2020, 9, e014501. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.Y.; Chen, Y.F.; Chan, H.C.; Chung, C.H.; Peng, H.Y.; Ho, Y.C.; Chen, C.H.; Chang, K.C.; Tang, C.H.; Lee, A.S. Role of apolipoprotein E in electronegative low-density lipoprotein-induced mitochondrial dysfunction in cardiomyocytes. Metabolism 2020, 107, 154227. [Google Scholar] [CrossRef]
- Kokkinaki, D.; Hoffman, M.; Kalliora, C.; Kyriazis, I.D.; Maning, J.; Lucchese, A.M.; Shanmughapriya, S.; Tomar, D.; Park, J.Y.; Wang, H.; et al. Chemically synthesized Secoisolariciresinol diglucoside (LGM2605) improves mitochondrial function in cardiac myocytes and alleviates septic cardiomyopathy. J. Mol. Cell. Cardiol. 2019, 127, 232–245. [Google Scholar] [CrossRef] [PubMed]
- Nukaga, S.; Mori, T.; Miyagawa, Y.; Fujiwara-Tani, R.; Sasaki, T.; Fujii, K.; Mori, S.; Goto, K.; Kishi, S.; Nakashima, C.; et al. Combined administration of lauric acid and glucose improved cancer-derived cardiac atrophy in a mouse cachexia model. Cancer Sci. 2020, 111, 4605–4615. [Google Scholar] [CrossRef]
- Wanka, H.; Lutze, P.; Staar, D.; Grunow, B.; Peters, B.S.; Peters, J. An alternative renin isoform is cardioprotective by modulating mitochondrial metabolism. J. Cell. Mol. Med. 2018, 22, 5991–6001. [Google Scholar] [CrossRef] [Green Version]
- Gong, G.; Liu, J.; Liang, P.; Guo, T.; Hu, Q.; Ochiai, K.; Hou, M.; Ye, Y.; Wu, X.; Mansoor, A.; et al. Oxidative capacity in failing hearts. Am. J. Physiol.-Heart Circ. Physiol. 2003, 285, H541–H548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schipper, D.A.; Palsma, R.; Marsh, K.M.; O’Hare, C.; Dicken, D.S.; Lick, S.; Kazui, T.; Johnson, K.; Smolenski, R.T.; Duncker, D.J.; et al. Chronic Myocardial Ischemia Leads to Loss of Maximal Oxygen Consumption and Complex I Dysfunction. Ann. Thorac. Surg. 2017, 104, 1298–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dranka, B.P.; Hill, B.G.; Darley-Usmar, V.M. Mitochondrial reserve capacity in endothelial cells: The impact of nitric oxide and reactive oxygen species. Free Radic. Biol. Med. 2010, 48, 905–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Liposomes | Diameters (nm) | Polydispersity Index (PDI) | ζ-Potential (mV) |
|---|---|---|---|
| β-MEND (RES) | 79.7 ± 10.2 | 0.26 ± 0.03 | 37.7 ± 8.6 |
| Empty β-MEND | 78.8 ± 10.0 | 0.26 ± 0.02 | 28.4 ± 11.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsujioka, T.; Sasaki, D.; Takeda, A.; Harashima, H.; Yamada, Y. Resveratrol-Encapsulated Mitochondria-Targeting Liposome Enhances Mitochondrial Respiratory Capacity in Myocardial Cells. Int. J. Mol. Sci. 2022, 23, 112. https://doi.org/10.3390/ijms23010112
Tsujioka T, Sasaki D, Takeda A, Harashima H, Yamada Y. Resveratrol-Encapsulated Mitochondria-Targeting Liposome Enhances Mitochondrial Respiratory Capacity in Myocardial Cells. International Journal of Molecular Sciences. 2022; 23(1):112. https://doi.org/10.3390/ijms23010112
Chicago/Turabian StyleTsujioka, Takao, Daisuke Sasaki, Atsuhito Takeda, Hideyoshi Harashima, and Yuma Yamada. 2022. "Resveratrol-Encapsulated Mitochondria-Targeting Liposome Enhances Mitochondrial Respiratory Capacity in Myocardial Cells" International Journal of Molecular Sciences 23, no. 1: 112. https://doi.org/10.3390/ijms23010112
APA StyleTsujioka, T., Sasaki, D., Takeda, A., Harashima, H., & Yamada, Y. (2022). Resveratrol-Encapsulated Mitochondria-Targeting Liposome Enhances Mitochondrial Respiratory Capacity in Myocardial Cells. International Journal of Molecular Sciences, 23(1), 112. https://doi.org/10.3390/ijms23010112