
Cysteinyl Leukotriene Pathway and Cancer

 ,
,  , , , , ,
, , , , ,
Abstract
:1. Unmet Need of Cancer Treatment
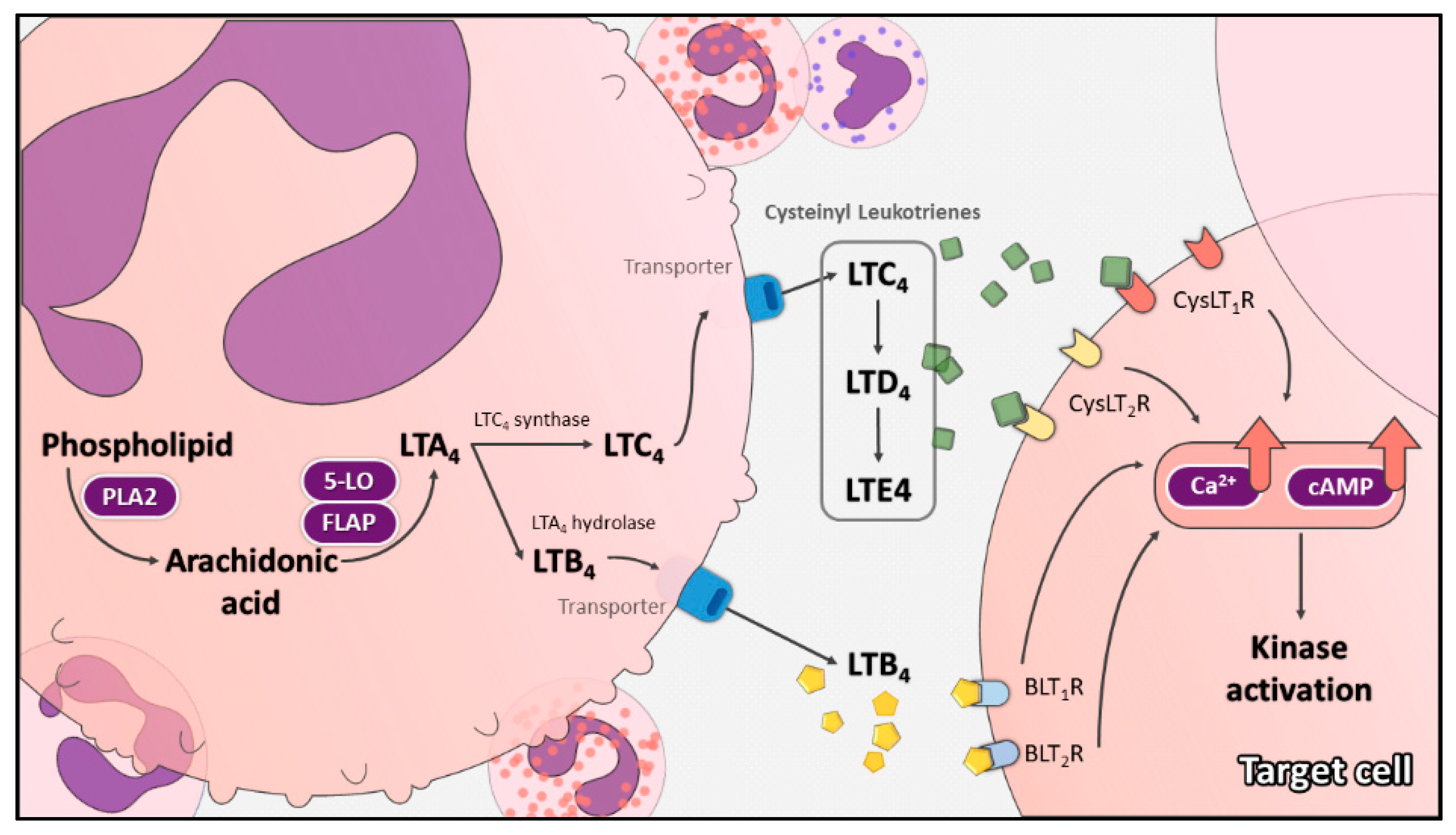
2. Cysteinyl Leukotriene Pathway
2.1. Leukotrienes
2.2. Cysteinyl Leukotrienes
- the affinity of CysLTs to CysLT1R: LTD4 > LTC4 >> LTE4;
- the affinity of CysLTs to CysLT2R: LTD4 = LTC4 >> LTE4;
- the affinity of CysLTs to CysLT3R: LTC4 > LTD4 [22].
2.3. CysLT1 Receptor (CysLT1R)
2.4. CysLT2 Receptor (CysLT2R)
2.5. CysLT3 Receptor (CysLT3R)
2.6. Cysteinyl Leukotriene Pathway Antagonists
3. In Vitro Studies about the Roles of Cysteinyl Leukotriene Pathway in Cancer
3.1. Hematologic Malignancies
3.2. Colorectal Cancer
3.3. Pancreatic Cancer and Hepatoma
3.4. Urological Malignancies
3.5. Breast Cancer
3.6. Lung Cancer
3.7. Neurological Malignancies
3.8. Other Malignancies
3.9. Drug Resistance and Cysteinyl Leukotriene Pathway
4. Animal Studies about the Roles of Cysteinyl Leukotriene Pathway in Cancer
4.1. The Role of Vascular Permeability Mediated by Cysteinyl Leukotrienes
4.2. Colorectal Cancer
4.3. Pancreatic Cancer
4.4. Lung Cancer
4.5. Neurological Malignancies
5. Clinical Studies about the Roles of Cysteinyl Leukotriene Pathway in Cancer
5.1. Hematological Malignancies
5.2. Colorectal Cancer
5.3. Esophageal Cancer and Gastric Cancer
5.4. Pancreatic Cancer and Hepatoma
5.5. Urological Malignancies
5.6. Breast Cancer
5.7. Other Malignancies
5.8. Chemopreventive Effects of Cysteinyl Leukotriene Inhibition
6. Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
| Author, Year | Cancer Cells | Major Findings |
|---|---|---|
| Hematologic malignancies | ||
| Stenke L, 1988 [37] | chronic myelogenous leukemia cell | Increased LTC4 synthase activity with increased LTC4 production in chronic myelogenous leukemia cells. |
| Stenke L, 1990 [38] | chronic myelogenous leukemia cell | Increased LTC4, rather than LTB4, producing capacity in patients of chronic myelogenous leukemia. |
| Chan CC, 1994 [16] | THP-1, a human monocytic leukemia cell line | LTD4 induced increased cytosolic calcium in THP-1 cells. |
| Hoshino M, 1998 [17] | THP-1, a human monocytic leukemia cell line | LTD4 activated the MAPK pathway. |
| Yektaei-Karin E, 2017 [39] | chronic myeloid leukemia (CML) cell lines (K562, KCL22, KU812), primary CD34 + blood cells from two CML patients, human colon carcinoma cell lung (HCT-116), lung fibroblast cell line (WI-38) | Montelukast and BWA4C (a 5-LO inhibitor) reduced cell proliferation of CML cells in a dose dependent manner. |
| Zovko A, 2018 [40] | chronic myeloid leukemia cell lines (K562 and JURL-MK1) | Montelukast, through a CysLT1R-dependent pathway, induced apoptosis of CML cells by inducing Bax overexpression, cytochrome c release, PARP-1 cleavage, and caspase-3 activation, as well as altered Wnt/β-catenin signaling. |
| Drost AC, 2012 [41] | chronic lymphocytic leukemia cell lines (EHEB, MEC-1); primary chronic lymphocytic leukemia cells from 54 patients; CD19 + (B cells) and CD19-cells from 8 healthy individuals | EHEB and MEC-1 cells expressed high levels of CysLT1R and low level of CysLT2R. LTD4 induced CysLT1R-mediated calcium fluxes, actin polymerization, chemotaxis and activation of MAPK pathway in chronic lymphocytic leukemia (CLL) cells. CysLT1R antagonists (MK571 and LY171883) reduced viability and increased apoptosis of CLL cells. |
| Schain F, 2008 [42] | human primary mediastinal B-cell lymphoma cell lines (Med-B1, Karpas-1106P) and a Hodgkin lymphoma cell line (L1236) | Med-B1, Karpas-1106P and L1236 cells expressed CysLT1R. LTC4 and LTD4 induced a calcium signal in Med-B1 cells, which could be blocked by zafirlukast. |
| Schain F, 2008 [43] | human Hodgkin lymphoma cell line cell lines (L1236, HDLM2, KMH2, L428, L591) | L1236 and KMH2 cells expressed functional CysLT1R, responding with a robust calcium signal upon LTD4 challenge, which could be blocked by zafirlukast. |
| Colorectal cancer | ||
| Ohd JF, 2003 [44] | Int 407 and two human colon cancer cell lines (Caco-2 and SW-480) | Int 407, Caco-2 and SW-480 expressed CysLT1R. The viability of Caco-2 cells increased with overexpression of CysLT1R. |
| Nielsen CK, 2003 [45] | Intestine 407 (Int 407), a human embryonic intestinal epithelial cell line | LTD4 prevented apoptosis of Int 407 cells induced by NS-398, a COX-2 inhibitor. |
| Wikström K, 2003 [46] | Intestine 407 (Int 407), a human embryonic intestinal epithelial cell line | LTD4 prevented apoptosis of Int 407 cells induced by NS-398, a COX-2 inhibitor. LTD4 also reduced the apoptotic potential by preventing caspase 8 activation and Bid cleavage. |
| Paruchuri S, 2002 [47] | Intestine 407 (Int 407), a human embryonic intestinal epithelial cell line | LTD4 enhance proliferation of Int 407 cells via two distinct signaling pathways (a Ras-independent and a Ras-dependent). |
| Wikström K, 2003 [48] | a human embryonic intestinal epithelial cell line (Int 407) and a human colorectal carcinoma cell line (Caco-2) | LTD4 upregulated COX-2 and Bcl-2 through a pertussis toxin sensitive G-protein and MAPK pathway in Int 407 and Caco-2 cells. |
| Parhamifar L, 2005 [49] | a human embryonic intestinal epithelial cell line (Int 407) and a human colorectal carcinoma cell line (Caco-2) | LTD4 stimulation induced cytosolic phospholipase A2α activation and nuclear translocation via a calcium-dependent mechanism involving activation of PKC and the MAPK pathway in both Int 407 and Caco-2 cells. |
| Mezhybovska M, 2006 [50] | Intestine 407 (Int 407), a human embryonic intestinal epithelial cell line | LTD4 induced β-catenin signaling in Int 407 cells, which activated TCF/LEF transcription factors in the nucleus and increased the association between β-catenin with Bcl-2 in the mitochondria. |
| Mezhybovska M, 2009 [51] | a human embryonic intestinal epithelial cell line (Int 407) and a human colorectal carcinoma cell line (Caco-2) | LTD4 increased mitochondrial metabolic activity and gene transcription and increased reactive oxygen species levels and subsequent activations of the p65 subunit of NF-κB, presumably through β-catenin accumulation in the mitochondria. |
| Paruchuri S, 2005 [52] | Intestine 407 (Int 407), a human embryonic intestinal epithelial cell line | LTD4 trigger a motile response of Int 407 cells via a PI3K/Rac signaling pathway. |
| Salim T, 2014 [53] | Human colorectal adenocarcinoma cell lines (HCT116, HT29) | LTD4 increased β-catenin level in colon cancer cells. In HCT116 cells but not in HT29 cells, LTD4 induced nuclear translocation of β-catenin, upregulated β-catenin target genes, and enhanced proliferation and migration; the effect could be prevented by pretreatment with ZM198,615, a CysLT1R antagonist. |
| Nielsen CK, 2005 [54] | a human embryonic intestinal epithelial cell line (Int 407) and a human colorectal carcinoma cell line (Caco-2) | CysLT1Rs were found in the outer nuclear membrane in Int 407 and Ca-co-2 cells. LTD4 induced nuclear translocation of the CysLT1Rs from the plasma membrane to the nucleus in Int 407 cells. |
| Yudina Y, 2008 [55] | Int 407 and 4 colon cancer cell lines (HT-29, SW-480, Caco-2, HCT-116) | The basal level of CysLT1R was higher in colon cancer cells compared to Int 407 cells. LTD4 significantly increased CysLT1R expression in Int 407 cells, but not in colon cancer cell lines; LTD4 induced upregulation of CysLT2R in colon cancer cell lines. |
| Paruchuri S, 2006 [56] | non-tumor intestinal epithelial cell lines (Int 407 and IEC-6) and tumor intestinal cell lines (Caco-2, SW480, HCT-116 and HT-29) | Constitutive CysLT1R signaling, which was maintained in an autocrine pattern, mediated both survival and proliferation of intestinal cells. A CysLT1R antagonist (MK571) induced apoptosis in non-tumor intestinal cells, but not in tumor-derived intestinal cell lines. |
| Bellamkonda K, 2018 [57] | human colon adenocarcinoma-derived cell lines (HT-29, SW-480) | Montelukast prevented LTD4-induced colony formation and disrupted colonospheres, as well as downregulation of cancer stem cell markers (ALDH1 and DCLK1). |
| Magnusson C, 2007 [59] | Int 407 and two human colon cancer cell lines (Caco-2 and SW-480) | The expression of CysLT2R, compared with CysLT1R, was higher in Int 407 but lower in two colon cancer cell lines (Caco-2 and SW480). CysLT2R was found to be located both at the plasma membrane and the nuclear membrane. CysLT2R signaling led to terminal differentiation of Caco-2 cells but had no effect on cell proliferation or apoptosis. |
| Magnusson C, 2011 [60] | Int 407 and 2 colon cancer cell lines (Caco-2, SW-480) | IFN-α could upregulate CysLT2R in Caco-2 cells. LTC4 induced expression mucin-2, and the effect could be blocked by AP 100984 (a specific CysLT2R antagonist) but not by montelukast (a specific CysLT1R antagonist). CysLT2R signaling was able to suppress cell migration induced by EGF signaling in Int 407 cells. |
| Bengtsson AM, 2013 [61] | colon cancer cell lines (Caco-2, SW-480, HCT-116) | All-trans retinoic acid (ATRA) treatment increased CysLT2R expression without affecting CysLT1R level, and upregulated LTC4 synthase in SW480 cells; the effect was not observed in HCT-116 cells. ATRA did not affect cell proliferation or induce apoptosis of SW480 cells. ATRA induced MUC-2 expression and alkaline phosphatase activity in SW480 cells, and a CysLT2R antagonist (AP 100984) reduced the effect. |
| Butler CT, 2019 [58] | HT29-Luc2 Bioware Ultra human colorectal cancer cell line | 1,4-dihydroxy quininib reduced clonal formation of HT29-Luc2. Gene silencing of CysLT1R in HT29-Luc2 cells significantly reduced expression of angiogenic marker calpain-2. |
| Pancreatic cancer and hepatoma | ||
| Kachi K, 2021 [62] | human pancreatic cancer cell lines (PA-TU-8988T, MIAPaCa-2, SUIT-2, PANC1) | CysLT1R was expressed in PA-TU-8988T, SUIT-2 and PANC1 cells, but not in MIAPaCa-2 cells. LTD4 promoted pancreatic cancer cell proliferation, whereas treatment with montelukast caused cell cycle arrest at G0/G1 phase without inducing apoptosis. |
| Arai J, 2021 [63] | human hepatoma cell lines (HepG2, PLC/PRF/5) | Pranlukast and montelukast inhibited ADAM9 activity and upregulated level of mem-brane-bound MHC class I-related chain A (mMICA). |
| Urological malignancies | ||
| Matsuyama M, 2010 [64] | a human RCC cell line (Caki-1), normal prostate stromal cells, a human bladder cancer cell line (T24), human prostate cancer cell lines (LNCaP, PC3, DU-145), a testicular cancer cell line (NEC-8), normal proximal tubular endothelial cells | All tested cancer cell lines showed CysLT1R expression, and treatment with montelukast downregulated CysLT1R expression, reduced cell viability, and induced early apoptosis in these cells. |
| Matsuyama M, 2009 [65] | T24, a human transitional cell carcinoma cell line | T24 cells expressed CysLT1R; montelukast induced apoptosis of T24 cells. |
| Matsuyama M, 2009 [66] | NEC-8, a human testicular cancer cell line | NEC-8 expressed CysLT1R. Montelukast reduced cell viability and induced apoptosis of NEC-8 cells. |
| Matsuyama M, 2007 [67] | human prostate cancer cell lines (LNCaP, PC3, DU-145) and normal stromal prostate cell lines | Montelukast significantly inhibited the proliferation and induced apoptosis of prostate cancer cell lines, but not normal stromal prostate cell lines. |
| Funao K, 2008 [68] | human renal cell carcinoma cell line (Caki-1) and normal proximal tubular endothelial cells | Montelukast significantly inhibited the proliferation and induced apoptosis of Caki-1, but not normal proximal tubular endothelial cells. |
| Tang C, 2018 [69] | human prostate cancer cell lines (LNCaP, PC-3, DU-145) and normal stromal prostate cell lines (NPCs) | Montelukast inhibited hypoxia-induced HIF-1α activation in prostate cancer cells, and inhibited their proliferation. Montelukast also induced downregulation of HIF-1α under hypoxic conditions, but the effect was not shown by pranlukast and zafirlukast. |
| Sarveswaran S, 2010 [70] | LNCaP, a human prostate cancer cell line | MK591, a 5-LO inhibitor developed to inhibit leukotriene biosynthesis, induced apoptosis in LNCaP cells. |
| Ghosh J, 1998 [71] | Hormone-responsive (LNCaP) and -nonresponsive (PC3) human prostate cancer cells | Inhibition of 5-LO by MK886 induced massive apoptosis in human prostate cancer cells, LNCaP and PC3. |
| Breast cancer | ||
| Przylipiak A, 1998 [72] | MCF-7, a human mammary cancer cell line | LTB4 and LTD4 inhibited growth of MCF-7 cells, and leukotriene antagonist LY171883 and 5-LO inhibitor MK886 further lifted the inhibitory effect. |
| Suknuntha K, 2018 [73] | MDAMB-231, a triple-negative breast cancer cell line | Montelukast and zafirlukast reduced cell viability of MDAMB-231 in a dose-dependent manner. Montelukast mainly induced apoptosis, while zafirlukast mainly exerted its action on cell cycle. |
| Magnusson C, 2011 [74] | breast cancer cell lines (MCF-7, MDA-MB-231) | Activating CysLT2R signaling with LTC4 did not affect cell proliferation or apoptosis of breast cancer cells but reduced cell migration. |
| Lung cancer | ||
| Avis IM, 1996 [75] | human small cell lung cancer cell lines (H209, H345, H82, N417) and human non-small cell lung cancer cell lines (H1155, H23, A549) | Inhibition of 5-LO resulted in enhanced levels of programmed cell death and significant growth reduction for a number of lung cancer cell lines. |
| Holm JB, 2013 [76] | A549, a human adenocarcinoma cell line | 5-LO inhibitors (AA861 or ETH 615-139) and zafirlukast (a CysLT1R antagonist) blocked the release of organic osmolytes (taurine, meAIB) and the concomitant cell volume restoration following hypoosmotic swelling of A549 cells; inhibition of 5-LO or CysLT1R did not affect caspase-3 activity during hypoxia. |
| Tsai MJ, 2017 [77] | human lung cancer cell lines (A549, H460, H1299, CL1-0, CL1-5) and mouse Lewis lung carcinoma cell line (LLC1) | Montelukast inhibited the viability/proliferation of lung cancer cells and induced cell death via nuclear translocation of apoptosis-inducing factor. |
| Neurological malignancies | ||
| Sveinbjörnsson B, 2008 [78] | human neuroblastoma cell lines (SH-SY5Y, SK-N-BE2, SK-N-SH, SK-N-AS, SK-N-FI, SK-N-DZ, IMR-32) and a human myelocytic cell line (U937) | All neuroblastoma cell lines expressed CysLT1R and CysLT2R. Neuroblastoma cells endogenously produced leukotrienes. LTD4 significantly increased cell viability and proliferation of neuroblastoma cells. Montelukast induced cell cycle arrest and apoptosis of neuroblastoma cells. |
| Piromkraipak P, 2018 [79] | glioblastoma cell lines (A172, U-87 MG) | Montelukast and zafirlukast inhibited proliferation and induced apoptosis in a concentration-dependent manner in glioblastoma cells; both medications decreased Bcl-2 expression without affecting Bax level. Montelukast induced more apoptosis than zafirlukast in A172 cells, but not in U-87 MG cells; zafirlukast caused cell cycle arrest at G0/G1 phase by upregulating the expression of p53 and p21 and showed a greater antiproliferative effect than montelukast. |
| Piromkraipak P, 2018 [80] | glioblastoma cell lines (A172, U373) and primary human astrocyte | Montelukast and zafirlukast, but not zileuton, significantly inhibited migration and invasion of glioblastoma cells, as well as inhibited the expression and activities of MMP-2 and MMP-9. |
| Other malignancies | ||
| Slater K, 2020 [81] | uveal melanoma cell lines derived from primary (Mel285, Mel270) and metastatic (OMM2.5) uveal melanoma | Uveal melanoma cell lines expressed CysLT1R and CysLT2R. Montelukast, quininib, and 1,4-dihydroxy quininib significantly inhibited uveal melanoma cells in a time- and dose- dependent manner, whereas a CysLT2-selective antagonist, HAMI 3379, did not show growth inhibition effect. Quininib significantly inhibited long-term proliferation, altered the cancer secretome of inflammatory and angiogenic factors, and inhibited oxidative phosphorylation. |
| Magi S, 2013 [82] | A431, an epidermoid carcinoma cell line | LTC4 mediated the second wave of Rac1 activation and cell migration; treatment with 5-LO inhibitors (AA861 and BU-4664L) or CysLT1R antagonists (MK571 and montelukast), as well as knockdown of CysLT1R, suppressed cell migration of A431 cells. |
| Association between cysteinyl leukotriene pathway and drug resistance | ||
| Gekeler V, 1995 [83] | multidrug resistant human leukemia cell line (HL60/AR) and human small cell lung cancer cell line (GLC4/ADR) | MK571 modulated MRP-associated multidrug resistance in HL60/AR and GLC4/ADR cells. |
| Brussel JP, 2004 [84] | PC3 and DU-145, human prostate cancer cell lines | Prostate cancer cell with multidrug resistant phenotype had increased expression of multidrug-resistance-associated protein 1 (MRP1) and adding MK571, zafirlukast or buthionine sulfoximine significantly increased the sensitivity to cytotoxic drugs. |
| Nagayama S, 1998 [85] | human epidermoid carcinoma cell line (KB-3-1) and its multidrug resistant subclone (CV 60) | ONO-1078, a LTD4 receptor antagonist, inhibited the transporting activity of MRP and increased the sensitivity to vincristin of CV 60 and KB-3-1 cells. |
| Nakano R, 1998 [86] | NCI-H520, a human lung cancer cell line | ONO-1078 enhanced the sensitivity of lung cancer NCI-H520 cells to vincristine, doxorubicin and etoposide through inhibiting the function of MRP. |
| Satapathy SR, 2020 [87] | human colon cancer cell lines (HCT116, HT-29) and their 5-FU-resistant (5-FU-R) cell lines | The 5-FU-R colon cancer cell lines expressed increased CysLT1R, which regulated 5-FU resistance via β-catenin activation and promoted 5-FU-R-derived stemness. Montelukast restricted the motility of 5-FU-R colon cancer cells, sensitized them to 5-FU, and decreased 5-FU-R-derived stemness. |
| Author, Year | Animal Model of Cancer | Major Findings |
|---|---|---|
| The role of vascular permeability mediated by cysteinyl leukotrienes | ||
| Chio CC, 1995 [88] | Rat C6 glioma model | LTE4 selectively opened the blood-tumor barrier and increased the tumor uptake of intravenously injected methotrexate. |
| Nozaki M, 2010 [89] | male Fisher rats, transplantable rat colon adenocarcinoma RCN9 cells implanted via the cisterna magna; Lewis lung carcinoma metastasis model in mice | Pranlukast, but not montelukast, inhibited tumor cell migration through brain capillary; both pranlukast and montelukast inhibited tumor cell migration through peripheral capillary. |
| Duah E, 2019 [90] | male C57BL/6 mice, including wild type, Cysltr1−/− and Cysltr2−/−, implanted with Matrigel plugs or subcutaneously injected with Lewis lung carcinoma (LLC) cells | CysLT2R regulated angiogenesis in isolated mouse endothelial cells and in Matrigel implants in mice. The growth and metastases of implanted LLC cells were significantly reduced in CysLT2R-null mice than in wild-type or CysLT1R-null mice. In wild-type mice, the expression of CysLT2R, but not CysLT1R, was increased in tumor vasculature, and BayCysLT2 (a selective CysLT2R antagonist), but not MK571 (a CysLT1R antagonist), reduced tumor growth, angiogenesis and lung metastasis of Lewis lung carcinoma cells in wild-type mice. |
| Colorectal cancer | ||
| Bellamkonda K, 2018 [57] | a xenograft model of nude mice subcutaneously injected with human colon adenocarcinoma-derived cell lines (HT-29, SW-480) | Montelukast significantly inhibited tumor growth and decreased amounts of cancer-stem cells. |
| Bellamkonda K, 2016 [91] | a nude mouse xenograft model with subcutaneous injection of human HCT-116 colon cancer cells | LTD4 promoted cancer-initiating cells in initiating tumor growth by allowing immuno-modulation of the tumor microenvironment. |
| Osman J, 2017 [92] | a colitis-associated colon cancer mice model induced by azoxymethane/dextran sulfate sodium | The mice with global disruption of CYSLTR1 gene expression had higher relative body weight, reduction in inflammation, and polyps with lower-grade dysplasia and decreased nuclear expression of β-catenin and COX-2. |
| Savari S, 2013 [93] | a xenograft model of nude mice inoculated with HCT-116 colon cancer cells | CysLT1R antagonists (montelukast and ZM198,615) inhibited proliferation and induced apoptosis of tumor cells, resulting in reduced size of the inoculated tumor. |
| Butler CT, 2019 [58] | HT29-Luc2 cells in immunocompromised mice (Balb/C nu/nu) | In the tumor xenograft model, 1,4-dihydroxy quininib significantly reduced tumor volume and the expression of angiogenic marker calpain-2. |
| Pancreatic cancer | ||
| Kachi K, 2021 [62] | a Syrian golden hamster model, using N-nitrosobis (2-oxopropyl) amine (BOP) to induce pancreatic ductal carcinomas | Montelukast suppressed pancreatic carcinogenesis by suppressing cell proliferation via the LTD4-CysLT1R axis. |
| Lung cancer | ||
| Tsai MJ, 2017 [77] | Mice with Lewis lung carcinoma injected subcutaneously | Montelukast significantly delayed the tumor growth, as well as decreased Ki-67 expression and increased terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL)-positive cells in the tumor tissue. |
| Gunning WT, 2002 [94] | vinyl carbamate injection to induce lung tumors in mice | Leukotriene pathway inhibitors (zafirlukast, MK886, and Zileuton) prevented lung tumor formation and slowed the growth and progression of adenomas to carcinoma. |
| Poczobutt JM, 2013 [95] | orthotopic immunocompetent mouse model of lung cancer with Lewis lung carcinoma cells injected into lungs of syngeneic mice | A later study, using an orthotopic immunocompetent mouse model of lung cancer with Lewis lung carcinoma cells injected into lungs of syngeneic mice, found increased production of leukotrienes (LTB4, LTC4, LTD4 and LTE4), in dependent on cytosolic phospholipase A2, in the tumor microenvironment. |
| Neurological malignancies | ||
| Black P, 1998 [96] | intracerebral C6 glioma in Sprague-Dawley rats | Pretreatment with LTC4 did significantly extend survival in rats treated with cisplatin |
| Author, Year | Patients/Specimens | Major Findings |
|---|---|---|
| Hematological malignancies | ||
| Schain F, 2008 [42] | lymph node biopsy specimens from 57 non-Hodgkin lymphoma patients | Primary mediastinal B-cell lymphoma was the only type showing CysLT1R expression in tumor cells, while other included lymphomas showed no CysLT1R expression. |
| Schain F, 2008 [43] | Specimens from 29 Hodgkin lymphoma patients | CysLT1R was expressed by primary Hodgkin Reed–Sternberg cells. |
| Sjölinder M, 2000 [97] | 17 patients in the chronic phase of Ph chromosome-positive (Ph+) chronic myeloid leukemia (CML) and 15 healthy medication-free volunteers | Mature CML CD16 (+) neutrophils had aberrantly increased expression of LTC4 synthase. |
| Roos C, 2008 [98] | 23 chronic myeloid leukemia (CML) patients and 18 healthy controls | Urinary excretion of LTE4 was significantly higher in CML patients than healthy controls. Neutrophilic LTC4 synthase expression and activity were markedly elevated and were normalized with imatinib mesylate treatment. |
| Dolinska M, 2017 [99] | bone marrow aspirates collected from 10 chronic myeloid leukemia (CML) patients and healthy adult volunteers | The majority of the single leukemic BCR-ABL+CD34+CD38− cells expressed CysLT1R and CysLT2R, but not ALOX5; treatment with zileuton or montelukast failed to suppress cell growth. |
| Colorectal cancer | ||
| Ohd JF, 2003 [44] | colorectal cancer samples from 84 patients | Using immunohistochemistry and in situ hybridization performed on tissue arrays showed that cysLT1R expression was significantly associated with poorer survival. |
| Nielsen CK, 2005 [54] | specimens from 44 patients with colorectal cancer | Colorectal cancer tissue showed higher CysLT1R and 5-LO immunohistochemical staining, particular in the nuclei, compared to normal colon tissue; the nuclear accumulation of CysLT1R was strongly correlated with stronger staining of proliferative marker Ki-67. |
| Magnusson C, 2007 [59] | tissues from 77 patients of colon cancer | Colon cancer tissue had a significantly lower CysLT2R expression than paired normal tissue; lower expression of CysLT2R in colon cancer was associated with later stage and poorer prognosis. |
| Magnusson C, 2011 [60] | specimens from 78 colon cancer patients | The tissue microarray showed that aggressive tumors generally expressed less IFN-α receptor and more EGFR, with a negative correlation between CysLT2R and EGFR expression. |
| Magnusson C, 2010 [100] | tissue microarray of 329 colorectal cancer patients | Higher nuclear expression of CysLT1R was associated with a poorer prognosis, whereas higher nuclear expression of CysLT2R was associated with a better prognosis. |
| Butler CT, 2019 [58] | human ex vivo colorectal tumor explants representing Dukes’ stage A, B and C colorectal cancer | In human ex vivo colorectal cancer tumor explants, 1,4-dihydroxy quininib significantly decreased the secretion of both angiogenic factor TIE-2 and adhesion molecule VCAM-1. |
| Esophageal cancer and gastric cancer | ||
| Shutt JD, 2012 [101] | esophageal biopsy specimens from 14 Barrett’s metaplasia, 2 high-grade dysplasia, 11 esophageal adenocarcinoma and 11 squamous control | Increased expression levels of LTB4 and CysLTs were found in esophageal adenocarcinoma tissue than in Barrett’s metaplasia and control tissues. |
| Venerito M, 2016 [102] | 19 patients with esophageal squamous cell cancer and 9 sex- and age-matched patients with functional dyspepsia | Significantly decreased expression levels of CysLT1R and CysLT2R were noted in esophageal cancer tissues compared with control squamous epithelium. |
| Venerito M, 2011 [103] | 35 gastric cancer tissue, 29 tumor-free tissue | Gastric cancer tissue showed significantly increased immunoreactive score of CysLT1R than tumor-free gastric mucosa; intestinal type had more CysLT1R and CysLT2R expression than the diffuse type. |
| Zhou Y, 2011 [104] | 92 hepatocellular carcinoma (HCC) patients and 20 health controls | Remarkably higher circulating LTD4 level was noted in HCC patients versus healthy subjects, especially in patients with chronic hepatitis B infection or metastasis. |
| Pancreatic cancer and hepatoma | ||
| Kachi K, 2021 [62] | 108 samples of pancreatic ductal adenocarcinoma tissues | Immunohistochemical analyses on pancreatic ductal adenocarcinoma tissues revealed that high CysLT1R expression was associated with worse overall survival. |
| Urological malignancies | ||
| Matsuyama M, 2010 [64] | tissue specimens from: 58 patients with renal cell carcinoma, paired with normal kidney tissue from 20 patients; 90 patients with bladder cancer, paired with normal bladder tissue from 30 patients; 151 patients with prostate cancer, 20 patients with prostatic intraepithelial neoplasia, 20 patients with benign prostatic hyperplasia, paired with normal prostate tissue from 20 patients; 30 patients with testicular cancer, paired with normal testis tissue from 10 patients | Immunohistochemistry revealed strong CysLT1R expression in all cancer samples, and more extensive and intense expression was noted in cancer with a higher grade or an advanced stage. Very weak CysLT1R expression was observed in relatively normal tissues. |
| Matsuyama M, 2009 [65] | tissue specimens from 90 transitional cell carcinoma (TCC) patients and 30 patients with normal bladder | CysLT1R expression was significantly more extensive and intense in TCC specimens, but not in normal bladder tissue; the expression level was greater in higher grade and advanced-stage cancer. |
| Matsuyama M, 2009 [66] | tissue specimens from 30 patients with testicular cancer and 10 patients with normal testes | Strong CysLT1R expression was observed in testicular cancer specimens, in contrast to weak expression in normal testes. |
| Matsuyama M, 2007 [67] | Tumor specimens were obtained from patients with prostate cancer (n = 151), prostatic intraepithelial neoplasia (n = 20), benign prostatic hyperplasia (n = 20), and normal prostate (n = 20) | Prostate cancer tissue had a significantly more extensive and intense CysLT1R expression, in nuclei and cytoplasm, than other groups, and the expression was stronger in those with higher Gleason score. |
| Funao K, 2008 [68] | Specimens from 58 patients with renal cell carcinoma and 20 patients with normal kidney tissues | Significantly more extensive and intense CysLT1R expression, in nuclei and cytoplasm, was observed in renal cell carcinoma tissues, compared with normal kidney tissue; the expression of CysLT1R was stronger in cancer tissues with higher grade. |
| Breast cancer | ||
| Magnusson C, 2011 [74] | specimens from 144 breast cancer patients | Breast cancers with higher CysLT1R and lower CysLT2R expression levels were associated with higher histological grade and worse overall survival. |
| Other malignancies | ||
| Sveinbjörnsson B, 2008 [78] | 27 neuroblastoma surgical specimens, 3 childhood ganglioneuromas, 3 samples of nonmalignant adrenals from children | CysLT1R staining was evident in all clinical tumor specimens and the adjacent vasculature. |
| Slater K, 2020 [81] | data of 80 primary uveal melanoma samples in The Cancer Genome Atlas (TCGA) and tissue from 52 patients of primary uveal melanoma in the Liverpool Ocular Oncology Centre | The data from TCGA showed that higher expression of CYSLTR1 and CYSLTR2 genes were significantly associated with poorer disease-free survival and overall survival. However, analysis with tissue microarray of 52 patients only showed poorer overall survival in those with higher CysLT1R expression but CysLT2R expression was not associated with survival. |
| Chemopreventive effects of cysteinyl leukotriene inhibition | ||
| Tsai MJ, 2016 [105] | 4185 CysLTR antagonist (LTRA) users and 20,925 LTRA non-users from Taiwan National Health Insurance Research Database | LTRA use decreased cancer risk in a dose-dependent manner in asthma patients. The chemopreventive effect of LTRA was markedly observed in terms of lung, colorectal, liver and breast cancer. |
| Sutton SS, 2021 [106] | 23,730 patients with leukotriene pathway inhibiting medication exposure and 534,736 patients from the data from the Department of Veteran Affairs | Patients with leukotriene pathway inhibiting medication (montelukast, zafirlukast, or zileuton) exposure had a significantly reduced risk of lung cancer than those without exposure. |
References
- Hsiao, A.J.; Chen, L.H.; Lu, T.H. Ten leading causes of death in Taiwan: A comparison of two grouping lists. J. Formos Med. Assoc. 2015, 114, 679–680. [Google Scholar] [CrossRef]
- World Health Organization. Cancer. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 15 November 2021).
- Robert, C. A decade of immune-checkpoint inhibitors in cancer therapy. Nat. Commun. 2020, 11, 3801. [Google Scholar] [CrossRef] [PubMed]
- Balkwill, F.R.; Capasso, M.; Hagemann, T. The tumor microenvironment at a glance. J. Cell Sci. 2012, 125, 5591–5596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurose, K.; Hoshaw-Woodard, S.; Adeyinka, A.; Lemeshow, S.; Watson, P.H.; Eng, C. Genetic model of multi-step breast carcinogenesis involving the epithelium and stroma: Clues to tumour-microenvironment interactions. Hum. Mol. Genet. 2001, 10, 1907–1913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, W.; Jiang, X.; Kim, D.; Guan, T.; Nicolls, M.R.; Rockson, S.G. Leukotrienes in Tumor-Associated Inflammation. Front. Pharmacol. 2020, 11, 1289. [Google Scholar] [CrossRef] [PubMed]
- Peters-Golden, M.; Gleason, M.M.; Togias, A. Cysteinyl leukotrienes: Multi-functional mediators in allergic rhinitis. Clin. Exp. Allergy 2006, 36, 689–703. [Google Scholar] [CrossRef] [Green Version]
- Ophir, J.; Brenner, S.; Kivity, S. Leukotrienes. Int. J. Dermatol. 1985, 24, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Peters-Golden, M.; Brock, T.G. 5-lipoxygenase and FLAP. Prostaglandins Leukot. Essent. Fat. Acids 2003, 69, 99–109. [Google Scholar] [CrossRef]
- Haeggstrom, J.Z.; Kull, F.; Rudberg, P.C.; Tholander, F.; Thunnissen, M.M. Leukotriene A4 hydrolase. Prostaglandins Other Lipid Mediat. 2002, 68-69, 495–510. [Google Scholar] [CrossRef]
- Peters-Golden, M.; Henderson, W.R., Jr. Leukotrienes. N. Engl. J. Med. 2007, 357, 1841–1854. [Google Scholar] [CrossRef]
- Al-Azzam, N.; Elsalem, L. Leukotriene D4 role in allergic asthma pathogenesis from cellular and therapeutic perspectives. Life Sci. 2020, 260, 118452. [Google Scholar] [CrossRef] [PubMed]
- Crooks, S.W.; Stockley, R.A. Leukotriene B4. Int. J. Biochem. Cell Biol. 1998, 30, 173–178. [Google Scholar] [CrossRef]
- Singh, R.K.; Gupta, S.; Dastidar, S.; Ray, A. Cysteinyl leukotrienes and their receptors: Molecular and functional characteristics. Pharmacology 2010, 85, 336–349. [Google Scholar] [CrossRef]
- Lam, B.K.; Austen, K.F. Leukotriene C4 synthase: A pivotal enzyme in cellular biosynthesis of the cysteinyl leukotrienes. Prostaglandins Other Lipid Mediat. 2002, 68, 511–520. [Google Scholar] [CrossRef]
- Chan, C.C.; Ecclestone, P.; Nicholson, D.W.; Metters, K.M.; Pon, D.J.; Rodger, I.W. Leukotriene D4-induced increases in cytosolic calcium in THP-1 cells: Dependence on extracellular calcium and inhibition with selective leukotriene D4 receptor antagonists. J. Pharmacol. Exp. Ther. 1994, 269, 891–896. [Google Scholar]
- Hoshino, M.; Izumi, T.; Shimizu, T. Leukotriene D4 activates mitogen-activated protein kinase through a protein kinase Calpha-Raf-1-dependent pathway in human monocytic leukemia THP-1 cells. J. Biol. Chem. 1998, 273, 4878–4882. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.; Ford-Hutchinson, A.W.; Nicholson, D.W. Activation of protein kinase C down-regulates leukotriene C4 synthase activity and attenuates cysteinyl leukotriene production in an eosinophilic substrain of HL-60 cells. J. Immunol. 1994, 153, 776–788. [Google Scholar]
- Murphy, A.G.; Casey, R.; Maguire, A.; Tosetto, M.; Butler, C.T.; Conroy, E.; Reynolds, A.L.; Sheahan, K.; O’Donoghue, D.; Gallagher, W.M.; et al. Preclinical validation of the small molecule drug quininib as a novel therapeutic for colorectal cancer. Sci. Rep. 2016, 6, 34523. [Google Scholar] [CrossRef] [Green Version]
- Burke, L.; Butler, C.T.; Murphy, A.; Moran, B.; Gallagher, W.M.; O’Sullivan, J.; Kennedy, B.N. Evaluation of Cysteinyl Leukotriene Signaling as a Therapeutic Target for Colorectal Cancer. Front. Cell Dev. Biol. 2016, 4, 103. [Google Scholar] [CrossRef] [Green Version]
- Tsopanoglou, N.E.; Pipili-Synetos, E.; Maragoudakis, M.E. Leukotrienes C4 and D4 promote angiogenesis via a receptor-mediated interaction. Eur. J. Pharmacol. 1994, 258, 151–154. [Google Scholar] [CrossRef]
- Rovati, G.E.; Capra, V. Cysteinyl-leukotriene receptors and cellular signals. Sci. World J. 2007, 7, 1375–1392. [Google Scholar] [CrossRef]
- Ravasi, S.; Citro, S.; Viviani, B.; Capra, V.; Rovati, G.E. CysLT1 receptor-induced human airway smooth muscle cells proliferation requires ROS generation, EGF receptor transactivation and ERK1/2 phosphorylation. Respir Res. 2006, 7, 42. [Google Scholar] [CrossRef] [Green Version]
- Profita, M.; Sala, A.; Bonanno, A.; Siena, L.; Ferraro, M.; Di Giorgi, R.; Montalbano, A.M.; Albano, G.D.; Gagliardo, R.; Gjomarkaj, M. Cysteinyl leukotriene-1 receptor activation in a human bronchial epithelial cell line leads to signal transducer and activator of transcription 1-mediated eosinophil adhesion. J. Pharmacol. Exp. Ther. 2008, 325, 1024–1030. [Google Scholar] [CrossRef] [Green Version]
- Winkler, J.D.; Sarau, H.M.; Foley, J.J.; Crooke, S.T. Leukotriene D4-induced homologous desensitization in basal and differentiated U-937 cells: Characterization with the partial agonist leukotriene E4 and assessment of receptor reserve. J. Pharmacol. Exp. Ther. 1988, 247, 54–62. [Google Scholar]
- Snetkov, V.A.; Hapgood, K.J.; McVicker, C.G.; Lee, T.H.; Ward, J.P. Mechanisms of leukotriene D4-induced constriction in human small bronchioles. Br. J. Pharmacol. 2001, 133, 243–252. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.Y.; Tang, S.S.; Hu, M.; Long, Y.; Li, Y.Q.; Liao, M.X.; Ji, H.; Hong, H. Leukotriene D4 induces amyloid-beta generation via CysLT(1)R-mediated NF-kappaB pathways in primary neurons. Neurochem. Int. 2013, 62, 340–347. [Google Scholar] [CrossRef]
- Heise, C.E.; O’Dowd, B.F.; Figueroa, D.J.; Sawyer, N.; Nguyen, T.; Im, D.S.; Stocco, R.; Bellefeuille, J.N.; Abramovitz, M.; Cheng, R.; et al. Characterization of the human cysteinyl leukotriene 2 receptor. J. Biol. Chem. 2000, 275, 30531–30536. [Google Scholar] [CrossRef] [Green Version]
- Sjostrom, M.; Johansson, A.S.; Schroder, O.; Qiu, H.; Palmblad, J.; Haeggstrom, J.Z. Dominant expression of the CysLT2 receptor accounts for calcium signaling by cysteinyl leukotrienes in human umbilical vein endothelial cells. Arterioscler. Thromb. Vasc. Biol. 2003, 23, e37–e41. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.; Ying, M.; Gu, S.; Yin, W.; Li, Y.; Yuan, H.; Fang, S.; Li, M. Cysteinyl Leukotriene Receptor 2 is Involved in Inflammation and Neuronal Damage by Mediating Microglia M1/M2 Polarization through NF-kappaB Pathway. Neuroscience 2019, 422, 99–118. [Google Scholar] [CrossRef]
- Liu, T.; Barrett, N.A.; Kanaoka, Y.; Buchheit, K.; Laidlaw, T.M.; Garofalo, D.; Lai, J.; Katz, H.R.; Feng, C.; Boyce, J.A. Cysteinyl leukotriene receptor 2 drives lung immunopathology through a platelet and high mobility box 1-dependent mechanism. Mucosal Immunol. 2019, 12, 679–690. [Google Scholar] [CrossRef]
- Kanaoka, Y.; Maekawa, A.; Austen, K.F. Identification of GPR99 protein as a potential third cysteinyl leukotriene receptor with a preference for leukotriene E4 ligand. J. Biol. Chem. 2013, 288, 10967–10972. [Google Scholar] [CrossRef] [Green Version]
- Bankova, L.G.; Lai, J.; Yoshimoto, E.; Boyce, J.A.; Austen, K.F.; Kanaoka, Y.; Barrett, N.A. Leukotriene E4 elicits respiratory epithelial cell mucin release through the G-protein-coupled receptor, GPR99. Proc. Natl. Acad. Sci. USA 2016, 113, 6242–6247. [Google Scholar] [CrossRef] [Green Version]
- Bankova, L.G.; Dwyer, D.F.; Yoshimoto, E.; Ualiyeva, S.; McGinty, J.W.; Raff, H.; von Moltke, J.; Kanaoka, Y.; Frank Austen, K.; Barrett, N.A. The cysteinyl leukotriene 3 receptor regulates expansion of IL-25-producing airway brush cells leading to type 2 inflammation. Sci. Immunol. 2018, 3, eaat9453. [Google Scholar] [CrossRef] [Green Version]
- Godson, C. Balancing the Effect of Leukotrienes in Asthma. N. Engl. J. Med. 2020, 382, 1472–1475. [Google Scholar] [CrossRef]
- Mong, S.; Wu, H.L.; Wong, A.; Sarau, H.M.; Crooke, S.T. Leukotriene D4 receptor-mediated phosphoinositol hydrolysis and calcium mobilization in rat basophilic leukemic cells. J. Pharmacol. Exp. Ther. 1988, 247, 803–813. [Google Scholar]
- Stenke, L.; Reizenstein, P.; Lindgren, J.A. Increased leukotriene C4 production in chronic myelogenous leukemia. Acta Oncol. 1988, 27, 803–805. [Google Scholar] [CrossRef] [Green Version]
- Stenke, L.; Samuelsson, J.; Palmblad, J.; Dabrowski, L.; Reizenstein, P.; Lindgren, J.A. Elevated white blood cell synthesis of leukotriene C4 in chronic myelogenous leukaemia but not in polycythaemia vera. Br. J. Haematol. 1990, 74, 257–263. [Google Scholar] [CrossRef]
- Yektaei-Karin, E.; Zovko, A.; Nilsson, A.; Nasman-Glaser, B.; Kanter, L.; Radmark, O.; Wallvik, J.; Ekblom, M.; Dolinska, M.; Qian, H.; et al. Modulation of leukotriene signaling inhibiting cell growth in chronic myeloid leukemia. Leuk. Lymphoma 2017, 58, 1903–1913. [Google Scholar] [CrossRef]
- Zovko, A.; Yektaei-Karin, E.; Salamon, D.; Nilsson, A.; Wallvik, J.; Stenke, L. Montelukast, a cysteinyl leukotriene receptor antagonist, inhibits the growth of chronic myeloid leukemia cells through apoptosis. Oncol. Rep. 2018, 40, 902–908. [Google Scholar] [CrossRef] [Green Version]
- Drost, A.C.; Seitz, G.; Boehmler, A.; Funk, M.; Norz, K.P.; Zipfel, A.; Xue, X.; Kanz, L.; Mohle, R. The G protein-coupled receptor CysLT1 mediates chemokine-like effects and prolongs survival in chronic lymphocytic leukemia. Leuk. Lymphoma 2012, 53, 665–673. [Google Scholar] [CrossRef]
- Schain, F.; Schain, D.; Mahshid, Y.; Liu, C.; Porwit, A.; Xu, D.; Claesson, H.E.; Sundstrom, C.; Bjorkholm, M.; Sjoberg, J. Differential expression of cysteinyl leukotriene receptor 1 and 15-lipoxygenase-1 in non-Hodgkin lymphomas. Clin. Lymphoma Myeloma 2008, 8, 340–347. [Google Scholar] [CrossRef]
- Schain, F.; Tryselius, Y.; Sjoberg, J.; Porwit, A.; Backman, L.; Malec, M.; Xu, D.; Vockerodt, M.; Baumforth, K.R.; Wei, W.; et al. Evidence for a pathophysiological role of cysteinyl leukotrienes in classical Hodgkin lymphoma. Int. J. Cancer 2008, 123, 2285–2293. [Google Scholar] [CrossRef]
- Ohd, J.F.; Nielsen, C.K.; Campbell, J.; Landberg, G.; Lofberg, H.; Sjolander, A. Expression of the leukotriene D4 receptor CysLT1, COX-2, and other cell survival factors in colorectal adenocarcinomas. Gastroenterology 2003, 124, 57–70. [Google Scholar] [CrossRef]
- Nielsen, C.K.; Ohd, J.F.; Wikstrom, K.; Massoumi, R.; Paruchuri, S.; Juhas, M.; Sjolander, A. The leukotriene receptor CysLT1 and 5-lipoxygenase are upregulated in colon cancer. Adv. Exp. Med. Biol. 2003, 525, 201–204. [Google Scholar]
- Wikstrom, K.; Juhas, M.; Sjolander, A. The anti-apoptotic effect of leukotriene D4 involves the prevention of caspase 8 activation and Bid cleavage. Biochem. J. 2003, 371, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Paruchuri, S.; Hallberg, B.; Juhas, M.; Larsson, C.; Sjolander, A. Leukotriene D(4) activates MAPK through a Ras-independent but PKCepsilon-dependent pathway in intestinal epithelial cells. J. Cell Sci. 2002, 115, 1883–1893. [Google Scholar] [CrossRef]
- Wikstrom, K.; Ohd, J.F.; Sjolander, A. Regulation of leukotriene-dependent induction of cyclooxygenase-2 and Bcl-2. Biochem. Biophys. Res. Commun. 2003, 302, 330–335. [Google Scholar] [CrossRef]
- Parhamifar, L.; Jeppsson, B.; Sjolander, A. Activation of cPLA2 is required for leukotriene D4-induced proliferation in colon cancer cells. Carcinogenesis 2005, 26, 1988–1998. [Google Scholar] [CrossRef] [Green Version]
- Mezhybovska, M.; Wikstrom, K.; Ohd, J.F.; Sjolander, A. The inflammatory mediator leukotriene D4 induces beta-catenin signaling and its association with antiapoptotic Bcl-2 in intestinal epithelial cells. J. Biol. Chem. 2006, 281, 6776–6784. [Google Scholar] [CrossRef] [Green Version]
- Mezhybovska, M.; Yudina, Y.; Abhyankar, A.; Sjolander, A. Beta-catenin is involved in alterations in mitochondrial activity in non-transformed intestinal epithelial and colon cancer cells. Br. J. Cancer 2009, 101, 1596–1605. [Google Scholar] [CrossRef] [Green Version]
- Paruchuri, S.; Broom, O.; Dib, K.; Sjolander, A. The pro-inflammatory mediator leukotriene D4 induces phosphatidylinositol 3-kinase and Rac-dependent migration of intestinal epithelial cells. J. Biol. Chem. 2005, 280, 13538–13544. [Google Scholar] [CrossRef] [Green Version]
- Salim, T.; Sand-Dejmek, J.; Sjolander, A. The inflammatory mediator leukotriene D4 induces subcellular beta-catenin translocation and migration of colon cancer cells. Exp. Cell Res. 2014, 321, 255–266. [Google Scholar] [CrossRef]
- Nielsen, C.K.; Campbell, J.I.; Ohd, J.F.; Morgelin, M.; Riesbeck, K.; Landberg, G.; Sjolander, A. A novel localization of the G-protein-coupled CysLT1 receptor in the nucleus of colorectal adenocarcinoma cells. Cancer Res. 2005, 65, 732–742. [Google Scholar]
- Yudina, Y.; Parhamifar, L.; Bengtsson, A.M.; Juhas, M.; Sjolander, A. Regulation of the eicosanoid pathway by tumour necrosis factor alpha and leukotriene D4 in intestinal epithelial cells. Prostaglandins Leukot. Essent. Fat. Acids 2008, 79, 223–231. [Google Scholar] [CrossRef]
- Paruchuri, S.; Mezhybovska, M.; Juhas, M.; Sjolander, A. Endogenous production of leukotriene D4 mediates autocrine survival and proliferation via CysLT1 receptor signalling in intestinal epithelial cells. Oncogene 2006, 25, 6660–6665. [Google Scholar] [CrossRef] [Green Version]
- Bellamkonda, K.; Satapathy, S.R.; Douglas, D.; Chandrashekar, N.; Selvanesan, B.C.; Liu, M.; Savari, S.; Jonsson, G.; Sjolander, A. Montelukast, a CysLT1 receptor antagonist, reduces colon cancer stemness and tumor burden in a mouse xenograft model of human colon cancer. Cancer Lett. 2018, 437, 13–24. [Google Scholar] [CrossRef]
- Butler, C.T.; Kennedy, S.A.; Buckley, A.; Doyle, R.; Conroy, E.; Gallagher, W.M.; O’Sullivan, J.; Kennedy, B.N. 1,4-dihydroxy quininib attenuates growth of colorectal cancer cells and xenografts and regulates the TIE-2 signaling pathway in patient tumours. Oncotarget 2019, 10, 3725–3744. [Google Scholar] [CrossRef] [Green Version]
- Magnusson, C.; Ehrnstrom, R.; Olsen, J.; Sjolander, A. An increased expression of cysteinyl leukotriene 2 receptor in colorectal adenocarcinomas correlates with high differentiation. Cancer Res. 2007, 67, 9190–9198. [Google Scholar] [CrossRef] [Green Version]
- Magnusson, C.; Bengtsson, A.M.; Liu, M.; Liu, J.; Ceder, Y.; Ehrnstrom, R.; Sjolander, A. Regulation of cysteinyl leukotriene receptor 2 expression--a potential anti-tumor mechanism. PLoS ONE 2011, 6, e29060. [Google Scholar] [CrossRef] [Green Version]
- Bengtsson, A.M.; Jonsson, G.; Magnusson, C.; Salim, T.; Axelsson, C.; Sjolander, A. The cysteinyl leukotriene 2 receptor contributes to all-trans retinoic acid-induced differentiation of colon cancer cells. BMC Cancer 2013, 13, 336. [Google Scholar] [CrossRef] [Green Version]
- Kachi, K.; Kato, H.; Naiki-Ito, A.; Komura, M.; Nagano-Matsuo, A.; Naitoh, I.; Hayashi, K.; Kataoka, H.; Inaguma, S.; Takahashi, S. Anti-Allergic Drug Suppressed Pancreatic Carcinogenesis via Down-Regulation of Cellular Proliferation. Int. J. Mol. Sci. 2021, 22, 7444. [Google Scholar] [CrossRef] [PubMed]
- Arai, J.; Goto, K.; Otoyama, Y.; Nakajima, Y.; Sugiura, I.; Kajiwara, A.; Tojo, M.; Ichikawa, Y.; Uozumi, S.; Shimozuma, Y.; et al. Leukotriene receptor antagonists enhance HCC treatment efficacy by inhibiting ADAMs and suppressing MICA shedding. Cancer Immunol. Immunother. 2021, 70, 203–213. [Google Scholar] [CrossRef]
- Matsuyama, M.; Yoshimura, R. Cysteinyl-leukotriene1 receptor is a potent target for the prevention and treatment of human urological cancer. Mol. Med. Rep. 2010, 3, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, M.; Funao, K.; Hayama, T.; Tanaka, T.; Kawahito, Y.; Sano, H.; Takemoto, Y.; Nakatani, T.; Yoshimura, R. Relationship between cysteinyl-leukotriene-1 receptor and human transitional cell carcinoma in bladder. Urology 2009, 73, 916–921. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, M.; Funao, K.; Kawahito, Y.; Sano, H.; Chargui, J.; Touraine, J.L.; Nakatani, T.; Yoshimura, R. Expression of cysteinylLT1 receptor in human testicular cancer and growth reduction by its antagonist through apoptosis. Mol. Med. Rep. 2009, 2, 163–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuyama, M.; Hayama, T.; Funao, K.; Kawahito, Y.; Sano, H.; Takemoto, Y.; Nakatani, T.; Yoshimura, R. Overexpression of cysteinyl LT1 receptor in prostate cancer and CysLT1R antagonist inhibits prostate cancer cell growth through apoptosis. Oncol. Rep. 2007, 18, 99–104. [Google Scholar] [CrossRef] [Green Version]
- Funao, K.; Matsuyama, M.; Naganuma, T.; Kawahito, Y.; Sano, H.; Nakatani, T.; Yoshimura, R. The cysteinylLT1 receptor in human renal cell carcinoma. Mol. Med. Rep. 2008, 1, 185–189. [Google Scholar]
- Tang, C.; Lei, H.; Zhang, J.; Liu, M.; Jin, J.; Luo, H.; Xu, H.; Wu, Y. Montelukast inhibits hypoxia inducible factor-1alpha translation in prostate cancer cells. Cancer Biol. Ther. 2018, 19, 715–721. [Google Scholar] [CrossRef] [Green Version]
- Sarveswaran, S.; Myers, C.E.; Ghosh, J. MK591, a leukotriene biosynthesis inhibitor, induces apoptosis in prostate cancer cells: Synergistic action with LY294002, an inhibitor of phosphatidylinositol 3′-kinase. Cancer Lett. 2010, 291, 167–176. [Google Scholar] [CrossRef]
- Ghosh, J.; Myers, C.E. Inhibition of arachidonate 5-lipoxygenase triggers massive apoptosis in human prostate cancer cells. Proc. Natl. Acad. Sci. USA 1998, 95, 13182–13187. [Google Scholar] [CrossRef] [Green Version]
- Przylipiak, A.; Hafner, J.; Przylipiak, J.; Runnebaum, B.; Rabe, T.; Kohn, F.M. Influence of leukotrienes on in vitro growth of human mammary carcinoma cell line MCF-7. Eur. J. Obstet. Gynecol. Reprod. Biol. 1998, 77, 61–65. [Google Scholar] [CrossRef]
- Suknuntha, K.; Yubolphan, R.; Krueaprasertkul, K.; Srihirun, S.; Sibmooh, N.; Vivithanaporn, P. Leukotriene Receptor Antagonists Inhibit Mitogenic Activity in Triple Negative Breast Cancer Cells. Asian Pac. J. Cancer Prev. 2018, 19, 833–837. [Google Scholar] [CrossRef] [PubMed]
- Magnusson, C.; Liu, J.; Ehrnstrom, R.; Manjer, J.; Jirstrom, K.; Andersson, T.; Sjolander, A. Cysteinyl leukotriene receptor expression pattern affects migration of breast cancer cells and survival of breast cancer patients. Int. J. Cancer 2011, 129, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Avis, I.M.; Jett, M.; Boyle, T.; Vos, M.D.; Moody, T.; Treston, A.M.; Martinez, A.; Mulshine, J.L. Growth control of lung cancer by interruption of 5-lipoxygenase-mediated growth factor signaling. J. Clin. Investig. 1996, 97, 806–813. [Google Scholar] [CrossRef] [Green Version]
- Holm, J.B.; Grygorczyk, R.; Lambert, I.H. Volume-sensitive release of organic osmolytes in the human lung epithelial cell line A549: Role of the 5-lipoxygenase. Am. J. Physiol. Cell Physiol. 2013, 305, C48–C60. [Google Scholar] [CrossRef] [Green Version]
- Tsai, M.J.; Chang, W.A.; Tsai, P.H.; Wu, C.Y.; Ho, Y.W.; Yen, M.C.; Lin, Y.S.; Kuo, P.L.; Hsu, Y.L. Montelukast Induces Apoptosis-Inducing Factor-Mediated Cell Death of Lung Cancer Cells. Int. J. Mol. Sci. 2017, 18, 1353. [Google Scholar] [CrossRef] [Green Version]
- Sveinbjornsson, B.; Rasmuson, A.; Baryawno, N.; Wan, M.; Pettersen, I.; Ponthan, F.; Orrego, A.; Haeggstrom, J.Z.; Johnsen, J.I.; Kogner, P. Expression of enzymes and receptors of the leukotriene pathway in human neuroblastoma promotes tumor survival and provides a target for therapy. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2008, 22, 3525–3536. [Google Scholar] [CrossRef] [Green Version]
- Piromkraipak, P.; Parakaw, T.; Phuagkhaopong, S.; Srihirun, S.; Chongthammakun, S.; Chaithirayanon, K.; Vivithanaporn, P. Cysteinyl leukotriene receptor antagonists induce apoptosis and inhibit proliferation of human glioblastoma cells by downregulating B-cell lymphoma 2 and inducing cell cycle arrest. Can. J. Physiol. Pharmacol. 2018, 96, 798–806. [Google Scholar] [CrossRef]
- Piromkraipak, P.; Sangpairoj, K.; Tirakotai, W.; Chaithirayanon, K.; Unchern, S.; Supavilai, P.; Power, C.; Vivithanaporn, P. Cysteinyl Leukotriene Receptor Antagonists Inhibit Migration, Invasion, and Expression of MMP-2/9 in Human Glioblastoma. Cell Mol. Neurobiol. 2018, 38, 559–573. [Google Scholar] [CrossRef]
- Slater, K.; Heeran, A.B.; Garcia-Mulero, S.; Kalirai, H.; Sanz-Pamplona, R.; Rahman, A.; Al-Attar, N.; Helmi, M.; O’Connell, F.; Bosch, R.; et al. High Cysteinyl Leukotriene Receptor 1 Expression Correlates with Poor Survival of Uveal Melanoma Patients and Cognate Antagonist Drugs Modulate the Growth, Cancer Secretome, and Metabolism of Uveal Melanoma Cells. Cancers 2020, 12, 2950. [Google Scholar] [CrossRef]
- Magi, S.; Takemoto, Y.; Kobayashi, H.; Kasamatsu, M.; Akita, T.; Tanaka, A.; Takano, K.; Tashiro, E.; Igarashi, Y.; Imoto, M. 5-Lipoxygenase and cysteinyl leukotriene receptor 1 regulate epidermal growth factor-induced cell migration through Tiam1 upregulation and Rac1 activation. Cancer Sci. 2013, 105, 290–296. [Google Scholar] [CrossRef]
- Gekeler, V.; Ise, W.; Sanders, K.H.; Ulrich, W.R.; Beck, J. The leukotriene LTD4 receptor antagonist MK571 specifically modulates MRP associated multidrug resistance. Biochem. Biophys. Res. Commun. 1995, 208, 345–352. [Google Scholar] [CrossRef]
- van Brussel, J.P.; Oomen, M.A.; Vossebeld, P.J.; Wiemer, E.A.; Sonneveld, P.; Mickisch, G.H. Identification of multidrug resistance-associated protein 1 and glutathione as multidrug resistance mechanisms in human prostate cancer cells: Chemosensitization with leukotriene D4 antagonists and buthionine sulfoximine. BJU Int. 2004, 93, 1333–1338. [Google Scholar] [CrossRef] [PubMed]
- Nagayama, S.; Chen, Z.S.; Kitazono, M.; Takebayashi, Y.; Niwa, K.; Yamada, K.; Tani, A.; Haraguchi, M.; Sumizawa, T.; Furukawa, T.; et al. Increased sensitivity to vincristine of MDR cells by the leukotriene D4 receptor antagonist, ONO-1078. Cancer Lett. 1998, 130, 175–182. [Google Scholar] [CrossRef]
- Nakano, R.; Oka, M.; Nakamura, T.; Fukuda, M.; Kawabata, S.; Terashi, K.; Tsukamoto, K.; Noguchi, Y.; Soda, H.; Kohno, S. A leukotriene receptor antagonist, ONO-1078, modulates drug sensitivity and leukotriene C4 efflux in lung cancer cells expressing multidrug resistance protein. Biochem. Biophys. Res. Commun. 1998, 251, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Satapathy, S.R.; Sjolander, A. Cysteinyl leukotriene receptor 1 promotes 5-fluorouracil resistance and resistance-derived stemness in colon cancer cells. Cancer Lett. 2020, 488, 50–62. [Google Scholar] [CrossRef]
- Chio, C.C.; Lin, S.J.; Lin, M.T. Leukotriene E4 selectively increase the delivery of methotrexate to the C6 gliomas in rats. J. Neuro-Oncol. 1995, 25, 89–95. [Google Scholar] [CrossRef]
- Nozaki, M.; Yoshikawa, M.; Ishitani, K.; Kobayashi, H.; Houkin, K.; Imai, K.; Ito, Y.; Muraki, T. Cysteinyl leukotriene receptor antagonists inhibit tumor metastasis by inhibiting capillary permeability. Keio J. Med. 2010, 59, 10–18. [Google Scholar] [CrossRef] [Green Version]
- Duah, E.; Teegala, L.R.; Kondeti, V.; Adapala, R.K.; Keshamouni, V.G.; Kanaoka, Y.; Austen, K.F.; Thodeti, C.K.; Paruchuri, S. Cysteinyl leukotriene 2 receptor promotes endothelial permeability, tumor angiogenesis, and metastasis. Proc. Natl. Acad. Sci. USA 2019, 116, 199–204. [Google Scholar] [CrossRef] [Green Version]
- Bellamkonda, K.; Chandrashekar, N.K.; Osman, J.; Selvanesan, B.C.; Savari, S.; Sjolander, A. The eicosanoids leukotriene D4 and prostaglandin E2 promote the tumorigenicity of colon cancer-initiating cells in a xenograft mouse model. BMC cancer 2016, 16, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Osman, J.; Savari, S.; Chandrashekar, N.K.; Bellamkonda, K.; Douglas, D.; Sjolander, A. Cysteinyl leukotriene receptor 1 facilitates tumorigenesis in a mouse model of colitis-associated colon cancer. Oncotarget 2017, 8, 34773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savari, S.; Liu, M.; Zhang, Y.; Sime, W.; Sjolander, A. CysLT(1)R antagonists inhibit tumor growth in a xenograft model of colon cancer. PLoS ONE 2013, 8, e73466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunning, W.T.; Kramer, P.M.; Steele, V.E.; Pereira, M.A. Chemoprevention by lipoxygenase and leukotriene pathway inhibitors of vinyl carbamate-induced lung tumors in mice. Cancer Res. 2002, 62, 4199–4201. [Google Scholar] [PubMed]
- Poczobutt, J.M.; Gijon, M.; Amin, J.; Hanson, D.; Li, H.; Walker, D.; Weiser-Evans, M.; Lu, X.; Murphy, R.C.; Nemenoff, R.A. Eicosanoid profiling in an orthotopic model of lung cancer progression by mass spectrometry demonstrates selective production of leukotrienes by inflammatory cells of the microenvironment. PLoS ONE 2013, 8, e79633. [Google Scholar] [CrossRef] [Green Version]
- Black, P.; Hand, C.M.; Vender, J.R.; Finkelstein, S.D. Chemotherapy in experimental brain tumor, part 2: Pretreatment with leukotriene C4 prolongs survival. J. Neuro-Oncol. 1998, 36, 7–19. [Google Scholar] [CrossRef]
- Sjolinder, M.; Stenke, L.; Nasman-Glaser, B.; Widell, S.; Doucet, J.; Jakobsson, P.J.; Lindgren, J.A. Aberrant expression of active leukotriene C(4) synthase in CD16(+) neutrophils from patients with chronic myeloid leukemia. Blood 2000, 95, 1456–1464. [Google Scholar] [CrossRef]
- Roos, C.; Stenke, L.; Ohm, L.; Widell, S.; Kumlin, M.; Lindgren, J.A.; Tornhamre, S. Clinical imatinib mesylate treatment induces early normalisation of aberrant neutrophil leukotriene C4 synthase expression and activity in chronic myeloid leukaemia. Br. J. Haematol. 2008, 142, 992–995. [Google Scholar] [CrossRef]
- Dolinska, M.; Piccini, A.; Wong, W.M.; Gelali, E.; Johansson, A.S.; Klang, J.; Xiao, P.; Yektaei-Karin, E.; Stromberg, U.O.; Mustjoki, S.; et al. Leukotriene signaling via ALOX5 and cysteinyl leukotriene receptor 1 is dispensable for in vitro growth of CD34(+)CD38(-) stem and progenitor cells in chronic myeloid leukemia. Biochem. Biophys. Res. Commun. 2017, 490, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Magnusson, C.; Mezhybovska, M.; Lorinc, E.; Fernebro, E.; Nilbert, M.; Sjolander, A. Low expression of CysLT1R and high expression of CysLT2R mediate good prognosis in colorectal cancer. Eur. J. Cancer 2010, 46, 826–835. [Google Scholar] [CrossRef]
- Shutt, J.D.; Boger, P.; Neale, J.R.; Patel, P.; Sampson, A.P. Activity of the leukotriene pathway in Barrett’s metaplasia and oesophageal adenocarcinoma. Inflamm. Res. 2012, 61, 1379–1384. [Google Scholar] [CrossRef]
- Venerito, M.; Helmke, C.; Jechorek, D.; Wex, T.; Rosania, R.; Antweiler, K.; Weigt, J.; Malfertheiner, P. Leukotriene receptor expression in esophageal squamous cell cancer and non-transformed esophageal epithelium: A matched case control study. BMC Gastroenterol. 2016, 16, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venerito, M.; Kuester, D.; Harms, C.; Schubert, D.; Wex, T.; Malfertheiner, P. Upregulation of leukotriene receptors in gastric cancer. Cancers 2011, 3, 3156–3168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Guo, D.; Li, H.; Jie, S. Circulating LTD4 in patients with hepatocellular carcinoma. Tumor Biol. 2011, 32, 139–144. [Google Scholar] [CrossRef]
- Tsai, M.J.; Wu, P.H.; Sheu, C.C.; Hsu, Y.L.; Chang, W.A.; Hung, J.Y.; Yang, C.J.; Yang, Y.H.; Kuo, P.L.; Huang, M.S. Cysteinyl Leukotriene Receptor Antagonists Decrease Cancer Risk in Asthma Patients. Sci. Rep. 2016, 6, 23979. [Google Scholar] [CrossRef]
- Sutton, S.S.; Magagnoli, J.; Cummings, T.H.; Hardin, J.W. Leukotriene inhibition and the risk of lung cancer among U.S. veterans with asthma. Pulm. Pharmacol. Ther. 2021, 71, 102084. [Google Scholar] [CrossRef] [PubMed]

| Medication | Pharmacologic Effect | Remark |
|---|---|---|
| Zileuton | 5-LO inhibitor | Clinically available * |
| AA861 | 5-LO inhibitor | |
| BU-4664L | 5-LO inhibitor | |
| BWA4C | 5-LO inhibitor | |
| ETH 615-139 | 5-LO inhibitor | |
| MK591 | 5-LO inhibitor | |
| MK886 | 5-LO inhibitor | |
| Montelukast | Selective CysLT1R antagonist | Clinically available * |
| Zafirlukast | Selective CysLT1R antagonist | Clinically available * |
| Pranlukast | Selective CysLT1R antagonist | Clinically available * |
| MK571 | Selective CysLT1R antagonist | |
| ZM198,615 | Selective CysLT1R antagonist | |
| AP 100984 | Selective CysLT2R antagonist | |
| BayCysLT2 | Selective CysLT2R antagonist | |
| HAMI 3379 | Selective CysLT2R antagonist | |
| Quininib 1,4-dihydroxy quininib | Selective CysLT1R (and CysLT2R) antagonist |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, M.-J.; Chang, W.-A.; Chuang, C.-H.; Wu, K.-L.; Cheng, C.-H.; Sheu, C.-C.; Hsu, Y.-L.; Hung, J.-Y. Cysteinyl Leukotriene Pathway and Cancer. Int. J. Mol. Sci. 2022, 23, 120. https://doi.org/10.3390/ijms23010120
Tsai M-J, Chang W-A, Chuang C-H, Wu K-L, Cheng C-H, Sheu C-C, Hsu Y-L, Hung J-Y. Cysteinyl Leukotriene Pathway and Cancer. International Journal of Molecular Sciences. 2022; 23(1):120. https://doi.org/10.3390/ijms23010120
Chicago/Turabian StyleTsai, Ming-Ju, Wei-An Chang, Cheng-Hao Chuang, Kuan-Li Wu, Chih-Hung Cheng, Chau-Chyun Sheu, Ya-Ling Hsu, and Jen-Yu Hung. 2022. "Cysteinyl Leukotriene Pathway and Cancer" International Journal of Molecular Sciences 23, no. 1: 120. https://doi.org/10.3390/ijms23010120
APA StyleTsai, M. -J., Chang, W. -A., Chuang, C. -H., Wu, K. -L., Cheng, C. -H., Sheu, C. -C., Hsu, Y. -L., & Hung, J. -Y. (2022). Cysteinyl Leukotriene Pathway and Cancer. International Journal of Molecular Sciences, 23(1), 120. https://doi.org/10.3390/ijms23010120