
Telomerase Interaction Partners–Insight from Plants

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Candidate Telomerase-Binding Partners Interact with AtTERT, Pot1a or with Both Proteins

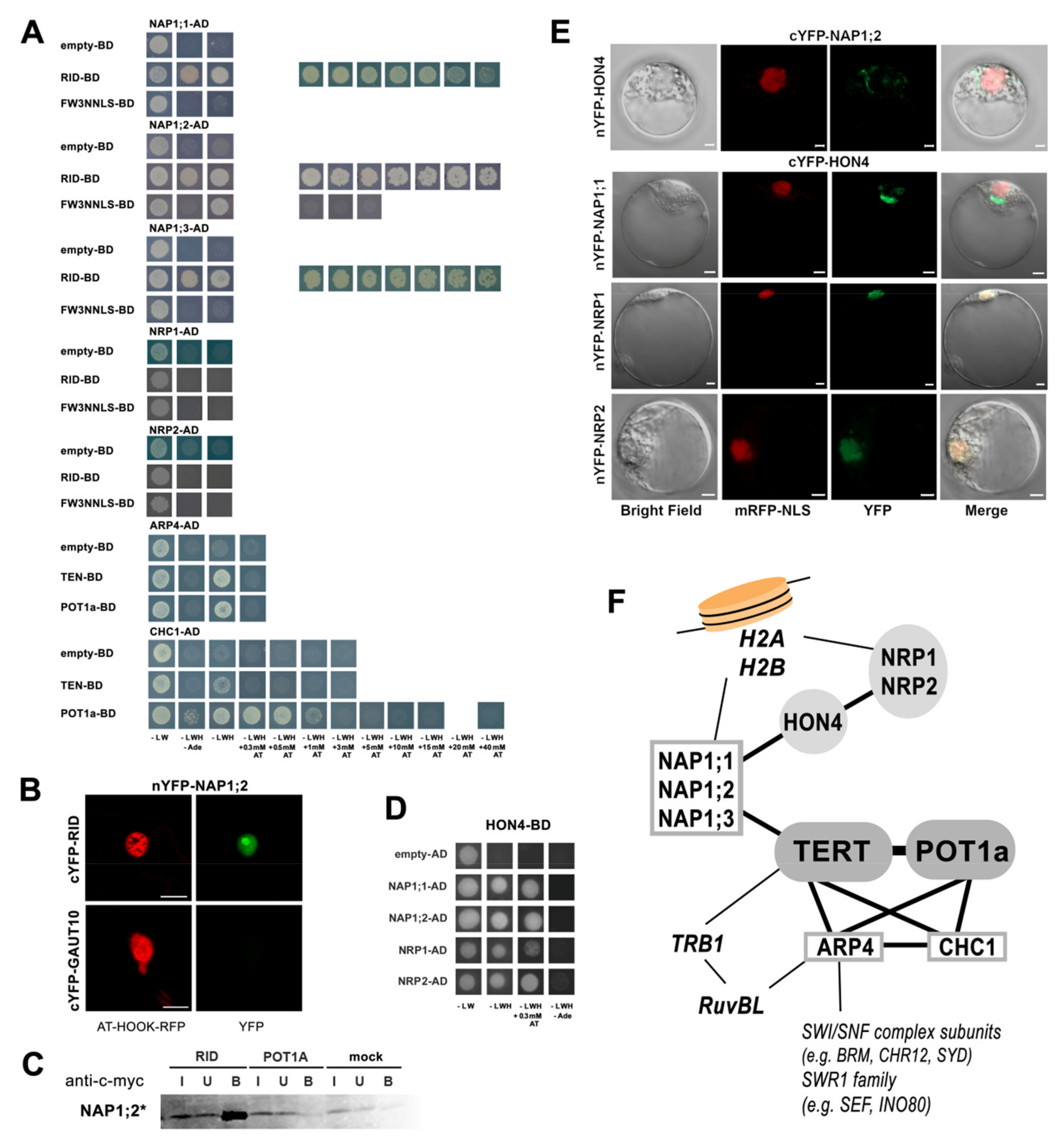
2.2. AtTERT Interacts with Chromatin Remodelling Proteins

2.3. Telomerase Interactions with DNA Replication Proteins
2.4. AtTERT N-Terminal Peptide Is Not Sufficient for Mitochondrial Entry
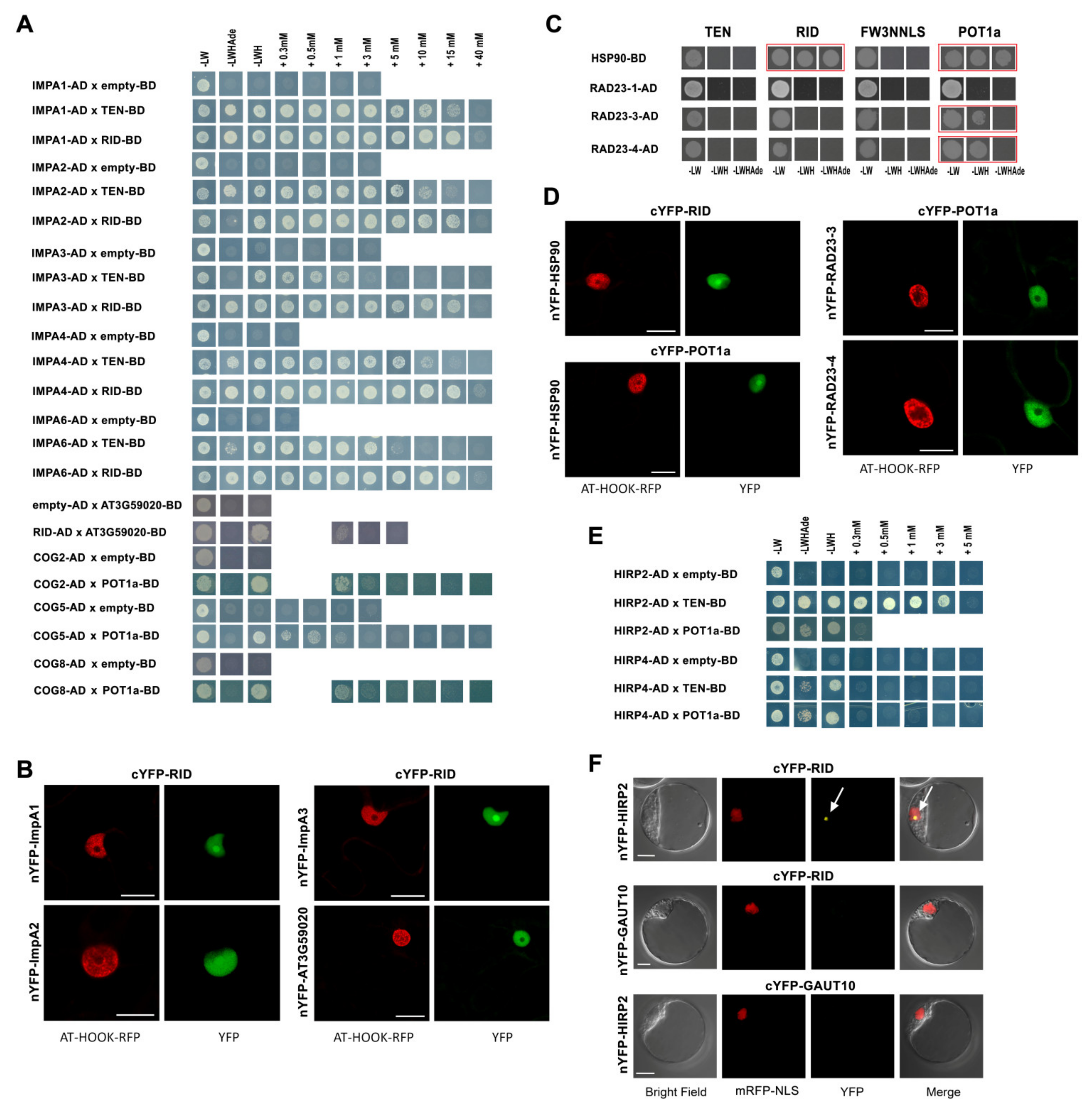
2.5. AtTERT and POT1a Interact with Importins, Chaperones and Proteins of Golgi Apparatus
2.6. AtTERT and POT1a Interact with Proteins Involved in Ribosome Maturation and RNA Processing
2.7. Connections between Telomere Length Maintenance System and Telomerase Interactors
2.8. The Plant Telomerase Interaction Network Is Comprised of Various Multiprotein Complexes
2.9. Limitations of the Study
3. Material and Methods
3.1. Preparation of Protein Constructs and Entry Clones
3.2. Yeast Two Hybrid (Y2H) Analysis
3.3. Bimolecular Fluorescence Complementation (BiFC) and Subcellular Localization of Protein Constructs
3.4. Co-Immunoprecipitation (co-IP) Analyses
3.5. In Vitro Mitochondrial Import Assay
3.6. Plant Material and Telomere Length Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chan, H.; Wang, Y.; Feigon, J. Progress in Human and Tetrahymena Telomerase Structure Determination. Annu. Rev. Biophys. 2017, 46, 199–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, J.C.; Cech, T.R. Human telomerase: Biogenesis, trafficking, recruitment, and activation. Genes Dev. 2015, 29, 1095–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wellinger, R.J.; Zakian, V.A. Everything you ever wanted to know about Saccharomyces cerevisiae telomeres: Beginning to end. Genetics 2012, 191, 1073–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, S.A.; Kim, K.; Lee, J.H.; Cha, J.S.; Khadka, P.; Cho, H.S.; Chung, I.K. Akt-mediated phosphorylation increases the binding affinity of hTERT for importin alpha to promote nuclear translocation. J. Cell Sci. 2015, 128, 2951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, Y.Y.; Her, J.; Oh, S.Y.; Chung, I.K. Hsp90-binding immunophilin FKBP52 modulates telomerase activity by promoting the cytoplasmic retrotransport of hTERT. Biochem. J. 2016, 473, 3517–3532. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, M.S.; Riha, K.; Gao, F.; Ren, S.; McKnight, T.D.; Shippen, D.E. Disruption of the telomerase catalytic subunit gene from Arabidopsis inactivates telomerase and leads to a slow loss of telomeric DNA. Proc. Natl. Acad. Sci. USA 1999, 96, 14813–14818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fajkus, P.; Peska, V.; Zavodnik, M.; Fojtova, M.; Fulneckova, J.; Dobias, S.; Kilar, A.; Dvorackova, M.; Zachova, D.; Necasova, I.; et al. Telomerase RNAs in land plants. Nucleic Acids Res. 2019, 47, 9842–9856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greider, C.W. Regulating telomere length from the inside out: The replication fork model. Genes Dev. 2016, 30, 1483–1491. [Google Scholar] [CrossRef] [Green Version]
- Pennock, E.; Buckley, K.; Lundblad, V. Cdc13 delivers separate complexes to the telomere for end protection and replication. Cell 2001, 104, 387–396. [Google Scholar] [CrossRef] [Green Version]
- Nandakumar, J.; Bell, C.F.; Weidenfeld, I.; Zaug, A.J.; Leinwand, L.A.; Cech, T.R. The TEL patch of telomere protein TPP1 mediates telomerase recruitment and processivity. Nature 2012, 492, 285–289. [Google Scholar] [CrossRef]
- De Lange, T. How telomeres solve the end-protection problem. Science 2009, 326, 948–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majerska, J.; Schrumpfova, P.P.; Dokladal, L.; Schorova, S.; Stejskal, K.; Oboril, M.; Honys, D.; Kozakova, L.; Polanska, P.S.; Sykorova, E. Tandem affinity purification of AtTERT reveals putative interaction partners of plant telomerase in vivo. Protoplasma 2017, 254, 1547–1562. [Google Scholar] [CrossRef]
- Rossignol, P.; Collier, S.; Bush, M.; Shaw, P.; Doonan, J.H. Arabidopsis POT1A interacts with TERT-V(I8), an N-terminal splicing variant of telomerase. J. Cell Sci. 2007, 120, 3678–3687. [Google Scholar] [CrossRef] [Green Version]
- Surovtseva, Y.V.; Shakirov, E.V.; Vespa, L.; Osbun, N.; Song, X.; Shippen, D.E. Arabidopsis POT1 associates with the telomerase RNP and is required for telomere maintenance. EMBO J. 2007, 26, 3653–3661. [Google Scholar] [CrossRef] [PubMed]
- Schrumpfova, P.P.; Vychodilova, I.; Dvorackova, M.; Majerska, J.; Dokladal, L.; Schorova, S.; Fajkus, J. Telomere repeat binding proteins are functional components of Arabidopsis telomeres and interact with telomerase. Plant J. 2014, 77, 770–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romaniuk, A.; Paszel-Jaworska, A.; Toton, E.; Lisiak, N.; Holysz, H.; Krolak, A.; Grodecka-Gazdecka, S.; Rubis, B. The non-canonical functions of telomerase: To turn off or not to turn off. Mol. Biol. Rep. 2019, 46, 1401–1411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackburn, E.H.; Epel, E.S.; Lin, J. Human telomere biology: A contributory and interactive factor in aging, disease risks, and protection. Science 2015, 350, 1193–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saretzki, G. Extra-telomeric functions of human telomerase: Cancer, mitochondria and oxidative stress. Curr. Pharm. Des. 2014, 20, 6386–6403. [Google Scholar] [CrossRef]
- Lin, K.W.; McDonald, K.R.; Guise, A.J.; Chan, A.; Cristea, I.M.; Zakian, V.A. Proteomics of yeast telomerase identified Cdc48-Npl4-Ufd1 and Ufd4 as regulators of Est1 and telomere length. Nat. Commun. 2015, 6, 8290. [Google Scholar] [CrossRef] [Green Version]
- Fu, D.; Collins, K. Purification of human telomerase complexes identifies factors involved in telomerase biogenesis and telomere length regulation. Mol. Cell 2007, 28, 773–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ungar, L.; Yosef, N.; Sela, Y.; Sharan, R.; Ruppin, E.; Kupiec, M. A genome-wide screen for essential yeast genes that affect telomere length maintenance. Nucleic Acids Res. 2009, 37, 3840–3849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zachova, D.; Fojtova, M.; Dvorackova, M.; Mozgova, I.; Lermontova, I.; Peska, V.; Schubert, I.; Fajkus, J.; Sykorova, E. Structure-function relationships during transgenic telomerase expression in Arabidopsis. Physiol. Plant. 2013, 149, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Dokladal, L.; Benkova, E.; Honys, D.; Duplakova, N.; Lee, L.Y.; Gelvin, S.B.; Sykorova, E. An armadillo-domain protein participates in a telomerase interaction network. Plant Mol. Biol. 2018, 97, 407–420. [Google Scholar] [CrossRef] [Green Version]
- Schrumpfova, P.P.; Fajkus, J. Composition and Function of Telomerase-A Polymerase Associated with the Origin of Eukaryotes. Biomolecules 2020, 10, 1425. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Zhu, Y.; Zhou, W.; Molinier, J.; Dong, A.; Shen, W.H. NAP1 family histone chaperones are required for somatic homologous recombination in Arabidopsis. Plant Cell 2012, 24, 1437–1447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolarova, K.; Nespor Dadejova, M.; Loja, T.; Lochmanova, G.; Sykorova, E.; Dvorackova, M. Disruption of NAP1 genes in Arabidopsis thaliana suppresses the fas1 mutant phenotype, enhances genome stability and changes chromatin compaction. Plant J. 2021, 106, 56–73. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Gao, J.; Ma, J.; Cao, L.; Zhang, C.; Zhu, Y.; Dong, A.; Shen, W.H. Distinct roles of the histone chaperones NAP1 and NRP and the chromatin-remodeling factor INO80 in somatic homologous recombination in Arabidopsis thaliana. Plant J. 2016, 88, 397–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charbonnel, C.; Rymarenko, O.; Da Ines, O.; Benyahya, F.; White, C.I.; Butter, F.; Amiard, S. The Linker Histone GH1-HMGA1 Is Involved in Telomere Stability and DNA Damage Repair. Plant Physiol. 2018, 177, 311–327. [Google Scholar] [CrossRef]
- Bieluszewski, T.; Galganski, L.; Sura, W.; Bieluszewska, A.; Abram, M.; Ludwikow, A.; Ziolkowski, P.A.; Sadowski, J. AtEAF1 is a potential platform protein for Arabidopsis NuA4 acetyltransferase complex. BMC Plant Biol. 2015, 15, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sacharowski, S.P.; Gratkowska, D.M.; Sarnowska, E.A.; Kondrak, P.; Jancewicz, I.; Porri, A.; Bucior, E.; Rolicka, A.T.; Franzen, R.; Kowalczyk, J.; et al. SWP73 Subunits of Arabidopsis SWI/SNF Chromatin Remodeling Complexes Play Distinct Roles in Leaf and Flower Development. Plant Cell 2015, 27, 1889–1906. [Google Scholar] [CrossRef]
- Liu, Z.; Zhu, Y.; Gao, J.; Yu, F.; Dong, A.; Shen, W.H. Molecular and reverse genetic characterization of NUCLEOSOME ASSEMBLY PROTEIN1 (NAP1) genes unravels their function in transcription and nucleotide excision repair in Arabidopsis thaliana. Plant J. 2009, 59, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.Q.; Gao, J.; Dong, A.W.; Shen, W.H. A truncated Arabidopsis NUCLEOSOME ASSEMBLY PROTEIN 1, AtNAP1;3T, alters plant growth responses to abscisic acid and salt in the Atnap1;3-2 mutant. Mol. Plant 2009, 2, 688–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schorova, S.; Fajkus, J.; Zaveska Drabkova, L.; Honys, D.; Schrumpfova, P.P. The plant Pontin and Reptin homologues, RuvBL1 and RuvBL2a, colocalize with TERT and TRB proteins in vivo, and participate in telomerase biogenesis. Plant J. 2019, 98, 195–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mozgova, I.; Mokros, P.; Fajkus, J. Dysfunction of chromatin assembly factor 1 induces shortening of telomeres and loss of 45S rDNA in Arabidopsis thaliana. Plant Cell 2010, 22, 2768–2780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaske, K.; Mokros, P.; Mozgova, I.; Fojtova, M.; Fajkus, J. A telomerase-independent component of telomere loss in chromatin assembly factor 1 mutants of Arabidopsis thaliana. Chromosoma 2013, 122, 285–293. [Google Scholar] [CrossRef]
- Van Leene, J.; Hollunder, J.; Eeckhout, D.; Persiau, G.; Van De Slijke, E.; Stals, H.; Van Isterdael, G.; Verkest, A.; Neirynck, S.; Buffel, Y.; et al. Targeted interactomics reveals a complex core cell cycle machinery in Arabidopsis thaliana. Mol. Syst. Biol. 2010, 6, 397. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Quimbaya, M.; Schubert, V.; Lammens, T.; Vandepoele, K.; Schubert, I.; Matsui, M.; Inze, D.; Berx, G.; De Veylder, L. The MCM-binding protein ETG1 aids sister chromatid cohesion required for postreplicative homologous recombination repair. PLoS Genet. 2010, 6, e1000817. [Google Scholar] [CrossRef] [Green Version]
- Tuteja, N.; Tran, N.Q.; Dang, H.Q.; Tuteja, R. Plant MCM proteins: Role in DNA replication and beyond. Plant Mol. Biol. 2011, 77, 537–545. [Google Scholar] [CrossRef]
- Froelich, C.A.; Kang, S.; Epling, L.B.; Bell, S.P.; Enemark, E.J. A conserved MCM single-stranded DNA binding element is essential for replication initiation. eLife 2014, 3, e01993. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Zhai, Y.; Zhang, Y.; Li, W.; Yang, M.; Lei, J.; Tye, B.K.; Gao, N. Structure of the eukaryotic MCM complex at 3.8 A. Nature 2015, 524, 186–191. [Google Scholar] [CrossRef]
- Lycka, M.; Peska, V.; Demko, M.; Spyroglou, I.; Kilar, A.; Fajkus, J.; Fojtova, M. WALTER: An easy way to online evaluate telomere lengths from terminal restriction fragment analysis. BMC Bioinform. 2021, 22, 145. [Google Scholar] [CrossRef] [PubMed]
- Forsburg, S.L. Eukaryotic MCM proteins: Beyond replication initiation. Microbiol. Mol. Biol. Rev. 2004, 68, 109–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drissi, R.; Dubois, M.L.; Douziech, M.; Boisvert, F.M. Quantitative Proteomics Reveals Dynamic Interactions of the Minichromosome Maintenance Complex (MCM) in the Cellular Response to Etoposide Induced DNA Damage. Mol. Cell. Proteom. 2015, 14, 2002–2013. [Google Scholar] [CrossRef] [Green Version]
- Dubois, M.L.; Bastin, C.; Levesque, D.; Boisvert, F.M. Comprehensive Characterization of Minichromosome Maintenance Complex (MCM) Protein Interactions Using Affinity and Proximity Purifications Coupled to Mass Spectrometry. J. Proteome Res. 2016, 15, 2924–2934. [Google Scholar] [CrossRef] [PubMed]
- Osman, K.; Yang, J.; Roitinger, E.; Lambing, C.; Heckmann, S.; Howell, E.; Cuacos, M.; Imre, R.; Durnberger, G.; Mechtler, K.; et al. Affinity proteomics reveals extensive phosphorylation of the Brassica chromosome axis protein ASY1 and a network of associated proteins at prophase I of meiosis. Plant J. 2018, 93, 17–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takashi, Y.; Kobayashi, Y.; Tanaka, K.; Tamura, K. Arabidopsis replication protein A 70a is required for DNA damage response and telomere length homeostasis. Plant Cell Physiol. 2009, 50, 1965–1976. [Google Scholar] [CrossRef]
- Aklilu, B.B.; Peurois, F.; Saintome, C.; Culligan, K.M.; Kobbe, D.; Leasure, C.; Chung, M.; Cattoor, M.; Lynch, R.; Sampson, L.; et al. Functional Diversification of Replication Protein A Paralogs and Telomere Length Maintenance in Arabidopsis. Genetics 2020, 215, 989–1002. [Google Scholar] [CrossRef]
- Huang, H.; Stromme, C.B.; Saredi, G.; Hodl, M.; Strandsby, A.; Gonzalez-Aguilera, C.; Chen, S.; Groth, A.; Patel, D.J. A unique binding mode enables MCM2 to chaperone histones H3-H4 at replication forks. Nat. Struct. Mol. Biol. 2015, 22, 618–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrie, C.; Whelan, J. Widespread dual targeting of proteins in land plants: When, where, how and why. Plant Signal. Behav. 2013, 8, 644–662. [Google Scholar] [CrossRef] [Green Version]
- Edmondson, A.C.; Song, D.; Alvarez, L.A.; Wall, M.K.; Almond, D.; McClellan, D.A.; Maxwell, A.; Nielsen, B.L. Characterization of a mitochondrially targeted single-stranded DNA-binding protein in Arabidopsis thaliana. Mol. Genet. Genom. 2005, 273, 115–122. [Google Scholar] [CrossRef]
- Nelson, B.K.; Cai, X.; Nebenfuhr, A. A multicolored set of in vivo organelle markers for co-localization studies in Arabidopsis and other plants. Plant J. 2007, 51, 1126–1136. [Google Scholar] [CrossRef]
- Yoo, H.H.; Kwon, C.; Lee, M.M.; Chung, I.K. Single-stranded DNA binding factor AtWHY1 modulates telomere length homeostasis in Arabidopsis. Plant J. 2007, 49, 442–451. [Google Scholar] [CrossRef] [PubMed]
- Palm, D.; Simm, S.; Darm, K.; Weis, B.L.; Ruprecht, M.; Schleiff, E.; Scharf, C. Proteome distribution between nucleoplasm and nucleolus and its relation to ribosome biogenesis in Arabidopsis thaliana. RNA Biol. 2016, 13, 441–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farmer, L.M.; Book, A.J.; Lee, K.H.; Lin, Y.L.; Fu, H.; Vierstra, R.D. The RAD23 family provides an essential connection between the 26S proteasome and ubiquitylated proteins in Arabidopsis. Plant Cell 2010, 22, 124–142. [Google Scholar] [CrossRef] [Green Version]
- Qi, Y.; Tsuda, K.; Nguyen le, V.; Wang, X.; Lin, J.; Murphy, A.S.; Glazebrook, J.; Thordal-Christensen, H.; Katagiri, F. Physical association of Arabidopsis hypersensitive induced reaction proteins (HIRs) with the immune receptor RPS2. J. Biol. Chem. 2011, 286, 31297–31307. [Google Scholar] [CrossRef] [Green Version]
- Koroleva, O.A.; Calder, G.; Pendle, A.F.; Kim, S.H.; Lewandowska, D.; Simpson, C.G.; Jones, I.M.; Brown, J.W.; Shaw, P.J. Dynamic behavior of Arabidopsis eIF4A-III, putative core protein of exon junction complex: Fast relocation to nucleolus and splicing speckles under hypoxia. Plant Cell 2009, 21, 1592–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, Z.; Dong, C.H.; Lee, H.; Zhu, J.; Xiong, L.; Gong, D.; Stevenson, B.; Zhu, J.K. A DEAD box RNA helicase is essential for mRNA export and important for development and stress responses in Arabidopsis. Plant Cell 2005, 17, 256–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palm, D.; Streit, D.; Shanmugam, T.; Weis, B.L.; Ruprecht, M.; Simm, S.; Schleiff, E. Plant-specific ribosome biogenesis factors in Arabidopsis thaliana with essential function in rRNA processing. Nucleic Acids Res. 2019, 47, 1880–1895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reichel, M.; Liao, Y.; Rettel, M.; Ragan, C.; Evers, M.; Alleaume, A.M.; Horos, R.; Hentze, M.W.; Preiss, T.; Millar, A.A. In Planta Determination of the mRNA-Binding Proteome of Arabidopsis Etiolated Seedlings. Plant Cell 2016, 28, 2435–2452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naprstkova, A.; Malinska, K.; Zaveska Drabkova, L.; Billey, E.; Naprstkova, D.; Sykorova, E.; Bousquet-Antonelli, C.; Honys, D. Characterization of ALBA Family Expression and Localization in Arabidopsis thaliana Generative Organs. Int. J. Mol. Sci. 2021, 22, 1652. [Google Scholar] [CrossRef]
- Lahmy, S.; Guilleminot, J.; Cheng, C.M.; Bechtold, N.; Albert, S.; Pelletier, G.; Delseny, M.; Devic, M. DOMINO1, a member of a small plant-specific gene family, encodes a protein essential for nuclear and nucleolar functions. Plant J. 2004, 39, 809–820. [Google Scholar] [CrossRef]
- Witkin, K.L.; Collins, K. Holoenzyme proteins required for the physiological assembly and activity of telomerase. Genes Dev. 2004, 18, 1107–1118. [Google Scholar] [CrossRef] [Green Version]
- Dona, M.; Mittelsten Scheid, O. DNA Damage Repair in the Context of Plant Chromatin. Plant Physiol. 2015, 168, 1206–1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleurdepine, S.; Deragon, J.M.; Devic, M.; Guilleminot, J.; Bousquet-Antonelli, C. A bona fide La protein is required for embryogenesis in Arabidopsis thaliana. Nucleic Acids Res. 2007, 35, 3306–3321. [Google Scholar] [CrossRef]
- Fojtova, M.; Peska, V.; Dobsakova, Z.; Mozgova, I.; Fajkus, J.; Sykorova, E. Molecular analysis of T-DNA insertion mutants identified putative regulatory elements in the AtTERT gene. J. Exp. Bot. 2011, 62, 5531–5545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perea-Resa, C.; Hernandez-Verdeja, T.; Lopez-Cobollo, R.; del Mar Castellano, M.; Salinas, J. LSM proteins provide accurate splicing and decay of selected transcripts to ensure normal Arabidopsis development. Plant Cell 2012, 24, 4930–4947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandasamy, M.K.; Deal, R.B.; McKinney, E.C.; Meagher, R.B. Silencing the nuclear actin-related protein AtARP4 in Arabidopsis has multiple effects on plant development, including early flowering and delayed floral senescence. Plant J. 2005, 41, 845–858. [Google Scholar] [CrossRef] [PubMed]
- Renfrew, K.B.; Song, X.; Lee, J.R.; Arora, A.; Shippen, D.E. POT1a and components of CST engage telomerase and regulate its activity in Arabidopsis. PLoS Genet. 2014, 10, e1004738. [Google Scholar] [CrossRef] [Green Version]
- Galati, A.; Micheli, E.; Cacchione, S. Chromatin structure in telomere dynamics. Front. Oncol. 2013, 3, 46. [Google Scholar] [CrossRef] [Green Version]
- Gentry, M.; Hennig, L. Remodelling chromatin to shape development of plants. Exp. Cell Res. 2014, 321, 40–46. [Google Scholar] [CrossRef]
- Ghanim, G.E.; Fountain, A.J.; van Roon, A.M.; Rangan, R.; Das, R.; Collins, K.; Nguyen, T.H.D. Structure of human telomerase holoenzyme with bound telomeric DNA. Nature 2021, 593, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Chuang, H.W.; Feng, J.H.; Feng, Y.L.; Wei, M.J. An Arabidopsis WDR protein coordinates cellular networks involved in light, stress response and hormone signals. Plant Sci. 2015, 241, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Trigg, S.A.; Garza, R.M.; MacWilliams, A.; Nery, J.R.; Bartlett, A.; Castanon, R.; Goubil, A.; Feeney, J.; O’Malley, R.; Huang, S.C.; et al. CrY2H-seq: A massively multiplexed assay for deep-coverage interactome mapping. Nat. Methods 2017, 14, 819–825. [Google Scholar] [CrossRef]
- Efroni, I.; Han, S.K.; Kim, H.J.; Wu, M.F.; Steiner, E.; Birnbaum, K.D.; Hong, J.C.; Eshed, Y.; Wagner, D. Regulation of leaf maturation by chromatin-mediated modulation of cytokinin responses. Dev. Cell 2013, 24, 438–445. [Google Scholar] [CrossRef] [Green Version]
- Kummari, D.; Palakolanu, S.R.; Kishor, P.B.K.; Bhatnagar-Mathur, P.; Singam, P.; Vadez, V.; Sharma, K.K. An update and perspectives on the use of promoters in plant genetic engineering. J. Biosci. 2020, 45, 119. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.Y.; Wu, F.H.; Hsu, C.T.; Shen, S.C.; Yeh, H.Y.; Liao, D.C.; Fang, M.J.; Liu, N.T.; Yen, Y.C.; Dokladal, L.; et al. Screening a cDNA library for protein-protein interactions directly in planta. Plant Cell 2012, 24, 1746–1759. [Google Scholar] [CrossRef] [Green Version]
- Citovsky, V.; Lee, L.Y.; Vyas, S.; Glick, E.; Chen, M.H.; Vainstein, A.; Gafni, Y.; Gelvin, S.B.; Tzfira, T. Subcellular localization of interacting proteins by bimolecular fluorescence complementation in planta. J. Mol. Biol. 2006, 362, 1120–1131. [Google Scholar] [CrossRef] [PubMed]
- Heinekamp, T.; Kuhlmann, M.; Lenk, A.; Strathmann, A.; Droge-Laser, W. The tobacco bZIP transcription factor BZI-1 binds to G-box elements in the promoters of phenylpropanoid pathway genes in vitro, but it is not involved in their regulation in vivo. Mol. Genet. Genom. 2002, 267, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Curtis, M.D.; Grossniklaus, U. A gateway cloning vector set for high-throughput functional analysis of genes in planta. Plant Physiol. 2003, 133, 462–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Alonso, J.M.; Stepanova, A.N.; Leisse, T.J.; Kim, C.J.; Chen, H.; Shinn, P.; Stevenson, D.K.; Zimmerman, J.; Barajas, P.; Cheuk, R.; et al. Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 2003, 301, 653–657. [Google Scholar] [CrossRef] [Green Version]
- Sessions, A.; Burke, E.; Presting, G.; Aux, G.; McElver, J.; Patton, D.; Dietrich, B.; Ho, P.; Bacwaden, J.; Ko, C.; et al. A high-throughput Arabidopsis reverse genetics system. Plant Cell 2002, 14, 2985–2994. [Google Scholar] [CrossRef]
- Rosso, M.G.; Li, Y.; Strizhov, N.; Reiss, B.; Dekker, K.; Weisshaar, B. An Arabidopsis thaliana T-DNA mutagenized population (GABI-Kat) for flanking sequence tag-based reverse genetics. Plant Mol. Biol. 2003, 53, 247–259. [Google Scholar] [CrossRef]
- Brunaud, V.; Balzergue, S.; Dubreucq, B.; Aubourg, S.; Samson, F.; Chauvin, S.; Bechtold, N.; Cruaud, C.; DeRose, R.; Pelletier, G.; et al. T-DNA integration into the Arabidopsis genome depends on sequences of pre-insertion sites. EMBO Rep. 2002, 3, 1152–1157. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Wang, J.; Miki, D.; Xia, R.; Yu, W.; He, J.; Zheng, Z.; Zhu, J.K.; Gong, Z. DNA replication factor C1 mediates genomic stability and transcriptional gene silencing in Arabidopsis. Plant Cell 2010, 22, 2336–2352. [Google Scholar] [CrossRef] [Green Version]
- Ni, D.A.; Sozzani, R.; Blanchet, S.; Domenichini, S.; Reuzeau, C.; Cella, R.; Bergounioux, C.; Raynaud, C. The Arabidopsis MCM2 gene is essential to embryo development and its over-expression alters root meristem function. New Phytol. 2009, 184, 311–322. [Google Scholar] [CrossRef]
- Herridge, R.P.; Day, R.C.; Macknight, R.C. The role of the MCM2-7 helicase complex during Arabidopsis seed development. Plant Mol. Biol. 2014, 86, 69–84. [Google Scholar] [CrossRef]
- Andreuzza, S.; Li, J.; Guitton, A.E.; Faure, J.E.; Casanova, S.; Park, J.S.; Choi, Y.; Chen, Z.; Berger, F. DNA LIGASE I exerts a maternal effect on seed development in Arabidopsis thaliana. Development 2010, 137, 73–81. [Google Scholar] [CrossRef] [Green Version]
- Griffith, M.E.; Mayer, U.; Capron, A.; Ngo, Q.A.; Surendrarao, A.; McClinton, R.; Jurgens, G.; Sundaresan, V. The TORMOZ gene encodes a nucleolar protein required for regulated division planes and embryo development in Arabidopsis. Plant Cell 2007, 19, 2246–2263. [Google Scholar] [CrossRef] [Green Version]
- Fojtova, M.; Fajkus, P.; Polanska, P.; Fajkus, J. Terminal Restriction Fragments (TRF) Method to Analyze Telomere Lengths. Bio-protocols 2015, 5, e1671. [Google Scholar] [CrossRef]
- Wan, X.; Wang, W.; Liu, J.; Tong, T. Estimating the sample mean and standard deviation from the sample size, median, range and/or interquartile range. BMC Med. Res. Methodol. 2014, 14, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higgins, J.P.T.; Thomas, J.; Chandler, J.; Cumpston, M.; Li, T.; Page, M.J.; Welch, W.A. (Eds.) Cochrane Handbook for Systematic Reviews of Interventions, 2nd ed.; John Wiley & Sons: Chichester, UK, 2019. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fulnečková, J.; Dokládal, L.; Kolářová, K.; Nešpor Dadejová, M.; Procházková, K.; Gomelská, S.; Sivčák, M.; Adamusová, K.; Lyčka, M.; Peska, V.; et al. Telomerase Interaction Partners–Insight from Plants. Int. J. Mol. Sci. 2022, 23, 368. https://doi.org/10.3390/ijms23010368
Fulnečková J, Dokládal L, Kolářová K, Nešpor Dadejová M, Procházková K, Gomelská S, Sivčák M, Adamusová K, Lyčka M, Peska V, et al. Telomerase Interaction Partners–Insight from Plants. International Journal of Molecular Sciences. 2022; 23(1):368. https://doi.org/10.3390/ijms23010368
Chicago/Turabian StyleFulnečková, Jana, Ladislav Dokládal, Karolína Kolářová, Martina Nešpor Dadejová, Klára Procházková, Sabina Gomelská, Martin Sivčák, Kateřina Adamusová, Martin Lyčka, Vratislav Peska, and et al. 2022. "Telomerase Interaction Partners–Insight from Plants" International Journal of Molecular Sciences 23, no. 1: 368. https://doi.org/10.3390/ijms23010368
APA StyleFulnečková, J., Dokládal, L., Kolářová, K., Nešpor Dadejová, M., Procházková, K., Gomelská, S., Sivčák, M., Adamusová, K., Lyčka, M., Peska, V., Dvořáčková, M., & Sýkorová, E. (2022). Telomerase Interaction Partners–Insight from Plants. International Journal of Molecular Sciences, 23(1), 368. https://doi.org/10.3390/ijms23010368