
Insecticidal Triterpenes in Meliaceae: Plant Species, Molecules, and Activities: Part II (Cipadessa, Melia)


Abstract
:1. Introduction
2. Plant Species and Their Insecticidal Chemicals
2.1. Cipadessa
2.1.1. Mexicanolide Limonoids
2.1.2. Rings B,D-Seco Limonoids
2.1.3. 10,11-Linkage Limonoids
2.2. Entandrophragma
2.2.1. Ring-Seco Limonoids
2.2.2. Rearranged Limonoids
2.2.3. Ring-Intact Limonoid
2.3. Guarea
2.4. Khaya
2.4.1. Rearranged Limonoids: 2,30-Linkage Type Chemicals
2.4.2. Ring-Seco Limonoids
2.4.3. Ring-Intact Limonoids: Anthothecol and Azadirone
2.5. Melia

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
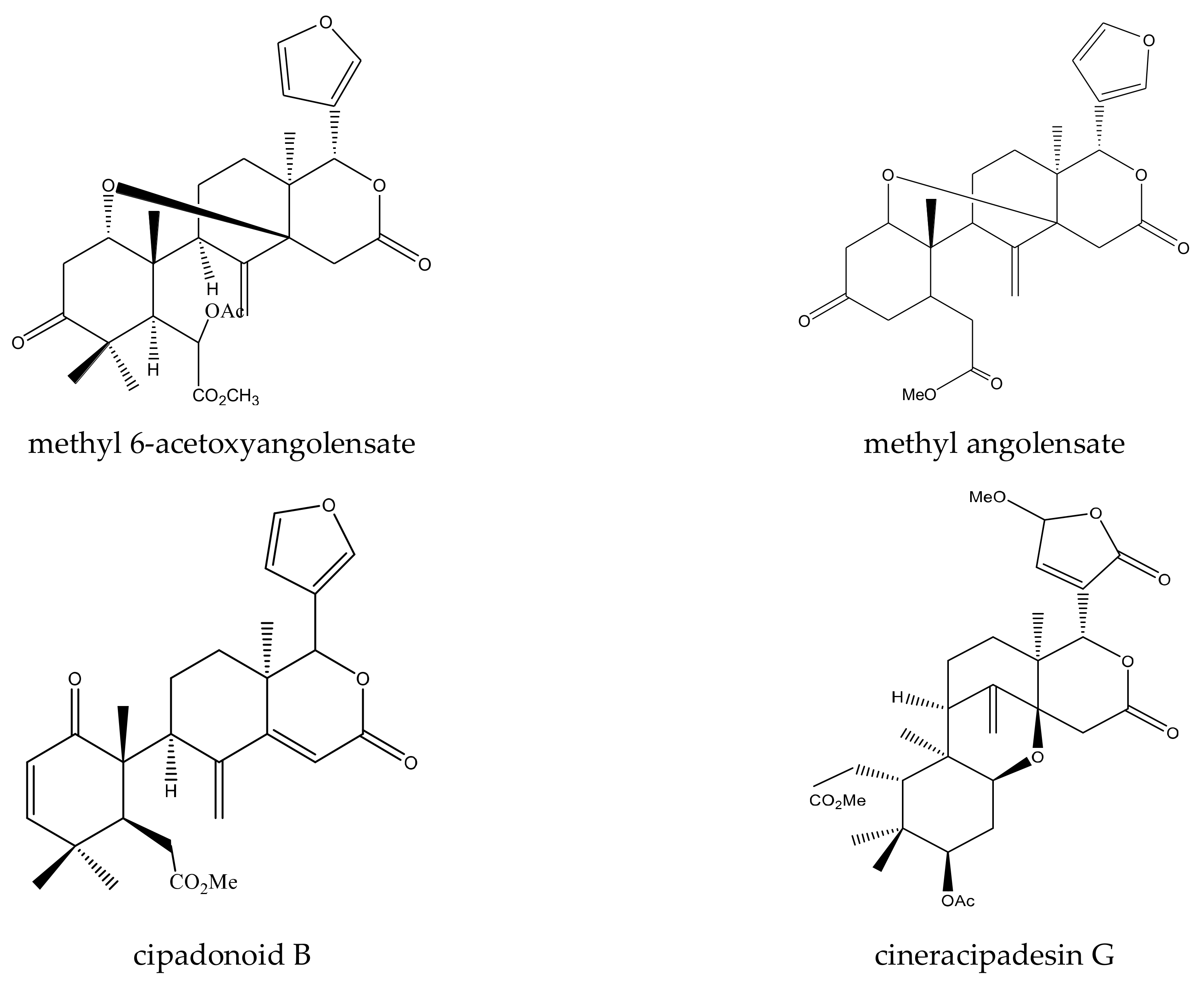{kind=link}
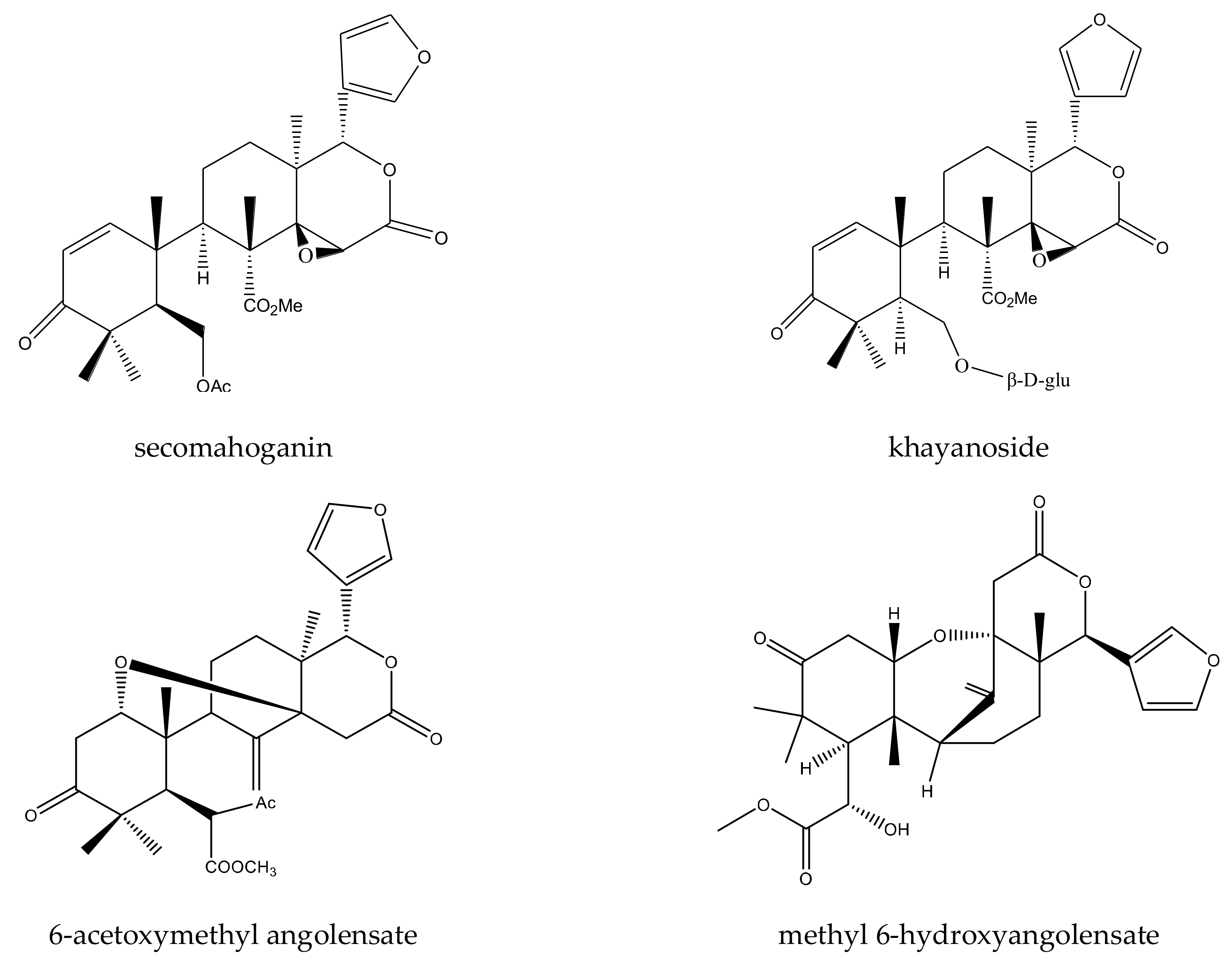{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
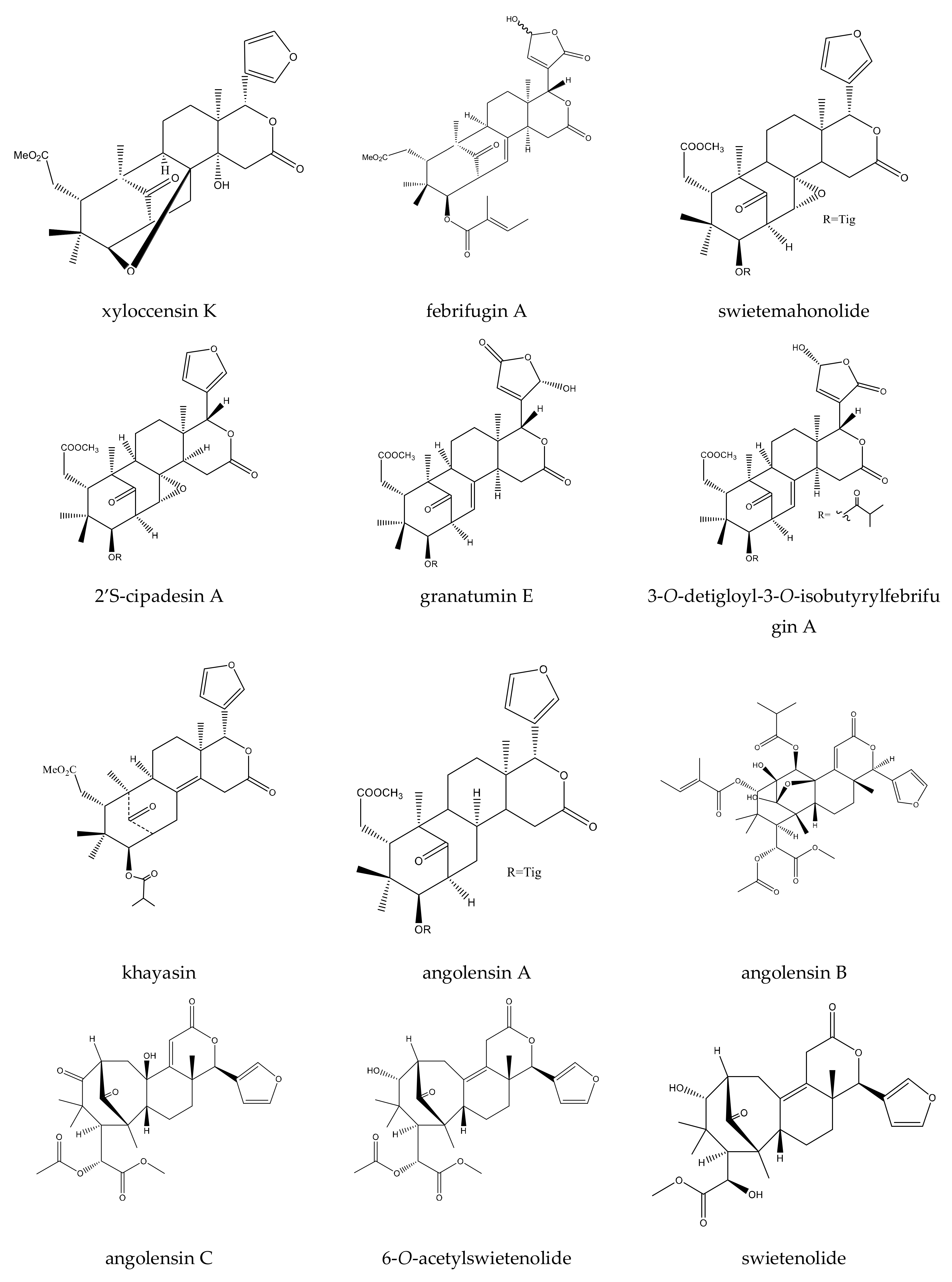{kind=link}
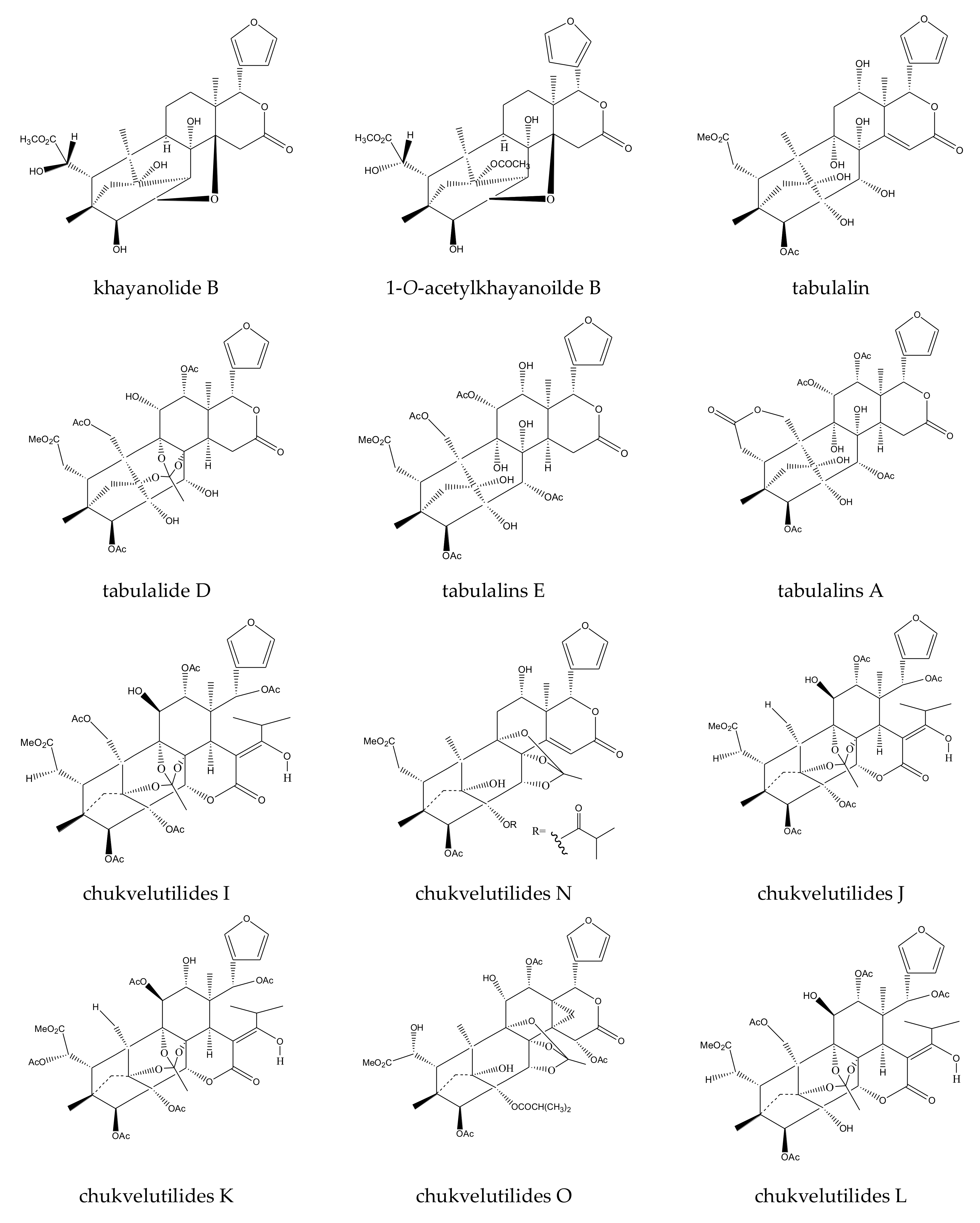{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Plant Source | Insect | Activity | Ref. |
|---|---|---|---|---|
| cineracipadesin G | Cipadessa cinerascens | Drosophila melanogaster | AI = 32.8% (1 mM) | [42] |
| febrifugin | Cipadessa fruticosa Cipadessa baccifera Cipadessa cinerascens | Spodoptera frugiperda | AFD at 100 mg/kg | [43] |
| 3-deacetyl-cipadonoid D | Cipadessa cinerascens | Drosophila melanogaster | AI = 39.1% (1 mM) (17 h) | [42] |
| angolensin A | Entandrophragma angolense | Spodoptera littoralis | AFD at 500 μg/mL | [31] |
| angolensin B | Entandrophragma angolense | Spodoptera littoralis | AFD at 500 μg/mL | [31] |
| angolensin C | Entandrophragma angolense | Spodoptera littoralis | AFD at 1000 μg/mL | [31] |
| 3β-hydroxy-3-deoxy- carapin | Entandrophragma angolense | Spodoptera littoralis | AFD at 1000 μg/mL | [31] |
| xyloccensin K | Entandrophragma angolense | Spodoptera littoralis | AFD at 1000 μg/mL | [31] |
| entandrophragmin | Entandrophragma cylindricum Entandrophragma bussei Entandrophragma spicatum Entandrophragma caudatum | Ostrinia nubilalis | FI at 500 μg/mL (48 h) | [11,46,131,132] |
| bussein | Entandrophragma bussei Entandrophragma caudatum | Ostrinia nubilalis | FI at 500 μg/mL (48 h) | [11,46] |
| 6-acetoxymethyl angolensate | Entandrophragma angolense | Spodoptera littoralis | MAC = 500 μg/mL | [11] |
| methyl angolensate | Entandrophragma angolense Entandrophragma macrophyllum Guarea thompsonii Khaya anthotheca Khaya senegalensis Khaya grandifoliola Khaya ivoremis | Spodoptera litura | PFI = 65.3 (24 h) | [40] |
| secomahoganin | Entandrophragma angolense | Spodoptera littoralis | AFD at 1000 μg/mL | [31] |
| azadirone | Entandrophragma delevoyi Khaya anthotheca | Leptinotarsa decemlineata | AI = 11.6–26.9 at 100–500 μg/mL (20 h) | [5] |
| gedunin | Entandrophragma angolense Entandrophragma delevoyi Entandrophragma macrophyllum Guarea grandiflora Khaya grandifoliola | Reticulitermes speratus | PC95 = 218.4 μg/disc (30 days) | [5,77,78] |
| melianone | Guarea grandiflora | Reticulitermes speratus | antifeeding activity at 100 μg/disc (30 days) | [41] |
| khayanolide D | Khaya senegalensis | Spodoptera littoralis | AI = 57.1 at 100 μg/mL (6 h) | [34] |
| khayanolide E | Khaya senegalensis | Spodoptera littoralis | MIC = 100 μg/mL | [33] |
| khayanolide A | Khaya senegalensis | Spodoptera littoralis | AI = 21.7 at 300 μg/mL (6 h) | [34] |
| khayanolide B | Khaya senegalensis | Spodoptera littoralis | AI = 24.8 at 1000 μg/mL (6 h) | [34] |
| 2-hydroxysenega- nolide | Khaya senegalensis | Spodoptera littoralis | AI = 38.4 at 200 μg/mL (6 h) | [34] |
| 1-O-acetylkhayanolide A | Khaya senegalensis | Spodoptera littoralis | AI = 31.4 at 100 μg/mL (6 h) | [34] |
| 1-O-acetylkhayanoilde B | Khaya senegalensis | Spodoptera littoralis | AI = 17.3 at 300 μg/mL (6 h) | [34] |
| khayanolide C | Khaya senegalensis | Spodoptera littoralis | AI = 31.5 at 500 μg/mL (6 h) | [34] |
| khayalactol | Khaya senegalensis | Spodoptera littoralis | AI = 83.8 at 1000 μg/mL (6 h) | [34] |
| khayanone | Khaya senegalensis | Spodoptera littoralis | AI = 47.4 at 1000 μg/mL (6 h) | [34] |
| 6-O-acetylswietenolide | Khaya grandifoliola | Spodoptera littoralis | AI = 72.2 at 1000 μg/mL | [5,35] |
| swietenolide | Khaya grandifoliola | Spodoptera littoralis | AI = 94.1 at 1000 μg/mL | [5,35] |
| seneganolide | Khaya senegalensis | Spodoptera littoralis | AI = 48.0 at 1000 μg/mL (6 h) | [34] |
| azadirone | Entandrophragma delevoyi Khaya anthotheca | Leptinotarsa decemlineata | AI = 11.6–26.9 at 100–500 μg/mL (20 h) | [5] |
| khayanoside | Khaya senegalensis | Spodoptera littoralis | AI = 15.1 at 1000 μg/mL (6 h) | [34] |
| methyl 6-hydroxyangolensate | Khaya senegalensis | Spodoptera littoralis | AI = 23.6 at 1000 μg/mL (6 h) | [34] |
| methyl 6-acetoxyangolensate | Khaya senegalensis | Spodoptera littoralis | AI = 18.0 at 1000 μg/mL (6 h) | [47] |
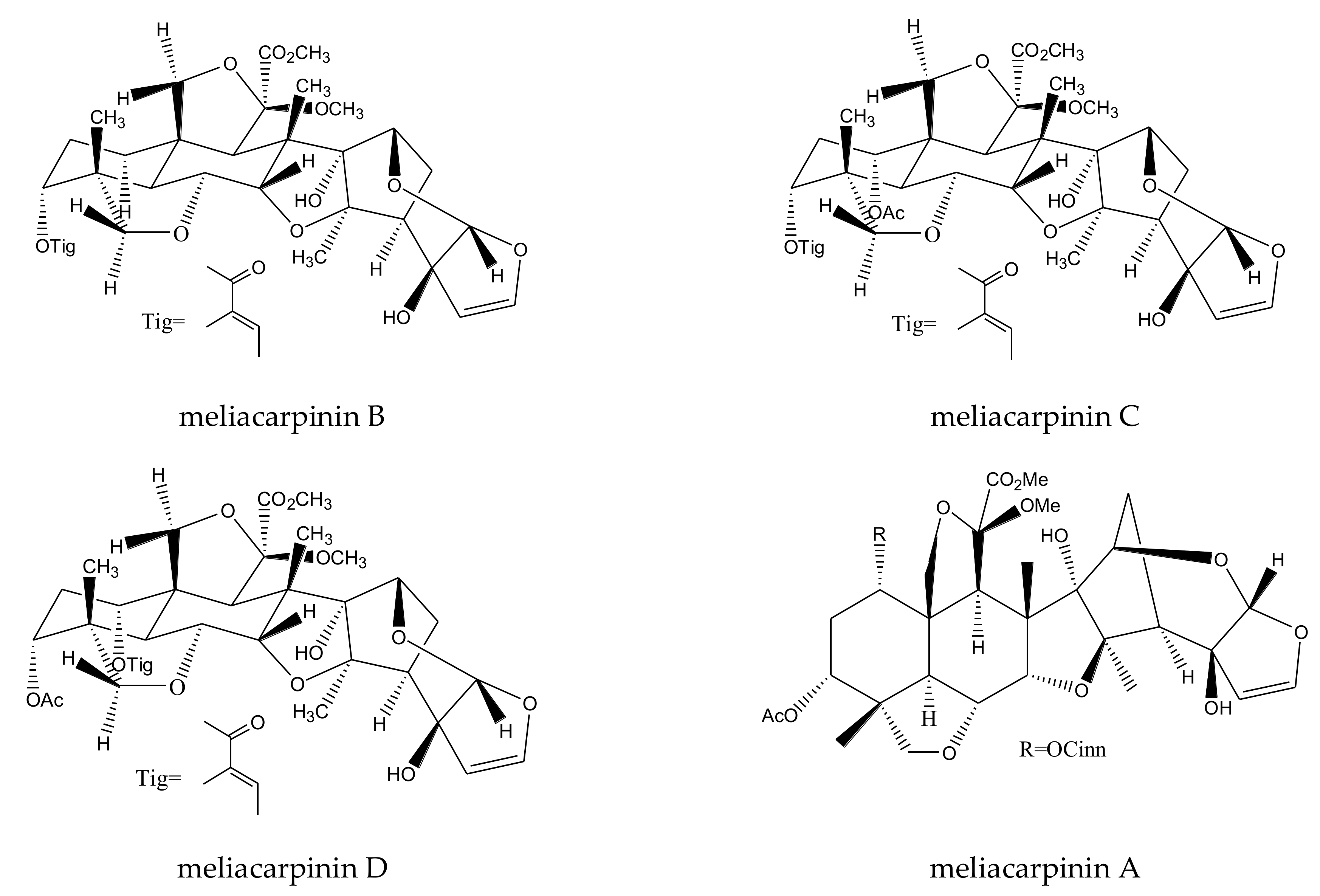
| meliacarpinin B | Melia azedarach | Spodoptera exigua | MIC = 50 μg/mL (2–24 h) | [36] |
| meliacarpinin C | Melia azedarach | Spodoptera exigua | MIC = 50 μg/mL (2–24 h) | [36] |
| meliacarpinin D | Melia azedarach | Spodoptera exigua | MIC = 50 μg/mL (2–24 h) | [36] |
| meliacarpinin A | Melia azedarach | Spodoptera exigua | MIC = 50 μg/mL (2–24 h) | [36] |
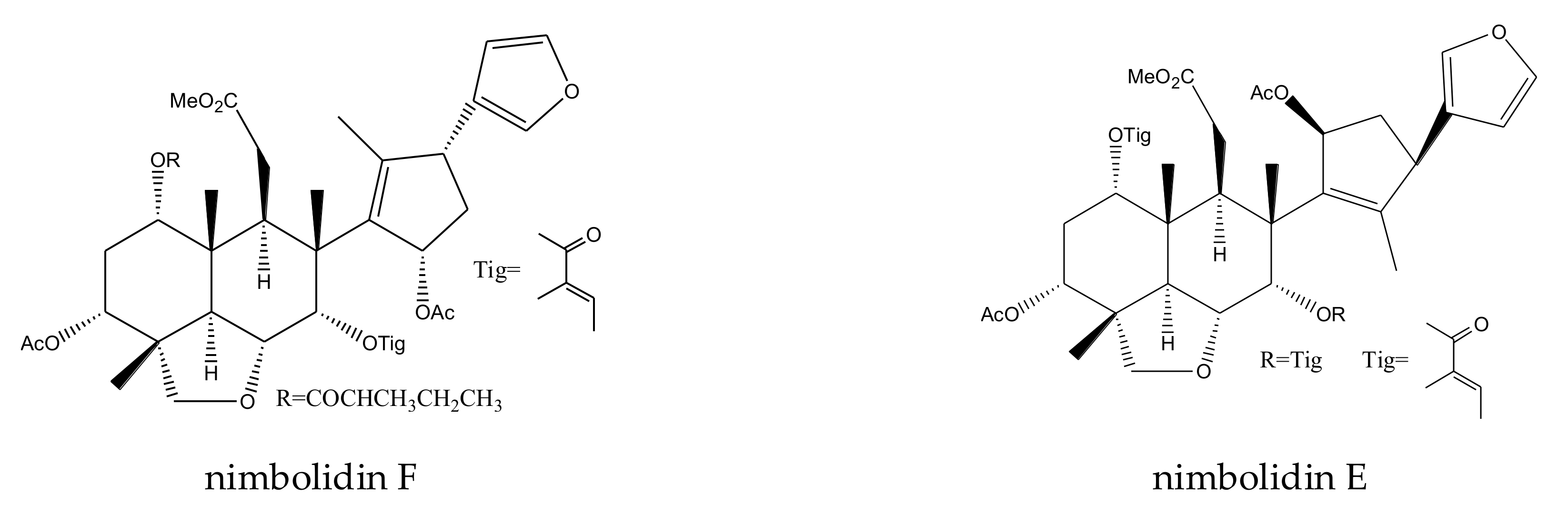
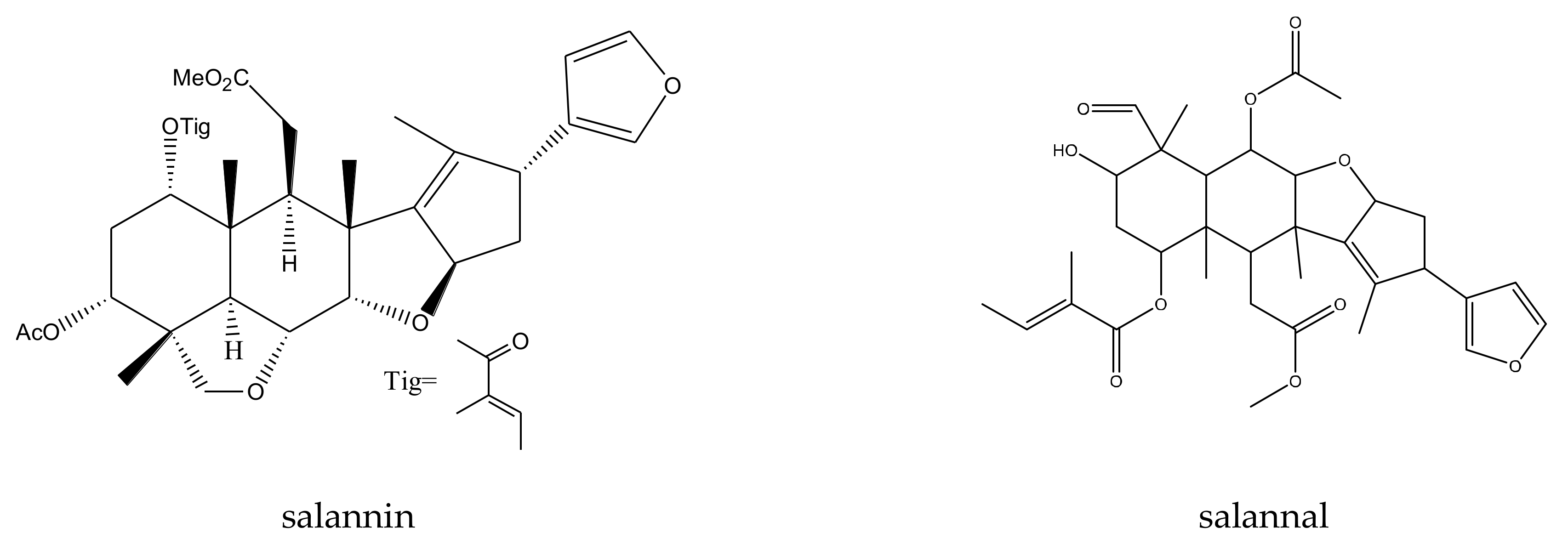
| salannal | Melia toosendan | Pieris rapae | AFC50 = 1.26 mM | [44] |
| 3-O-acetylohchinolal | Melia toosendan | Pieris rapae | AFC50 = 0.89 mM | [44] |
| salannin | Melia toosendan Melia azedarach | Pieris rapae | AFC50 = 1.35 mM | [44] |
| Spodoptera eridania | MIC = 1000 μg/mL (2–24 h) | [37] | ||
| ohchinal | Melia toosendan | Pieris rapae | AFC50 = 1.79 mM | [44] |
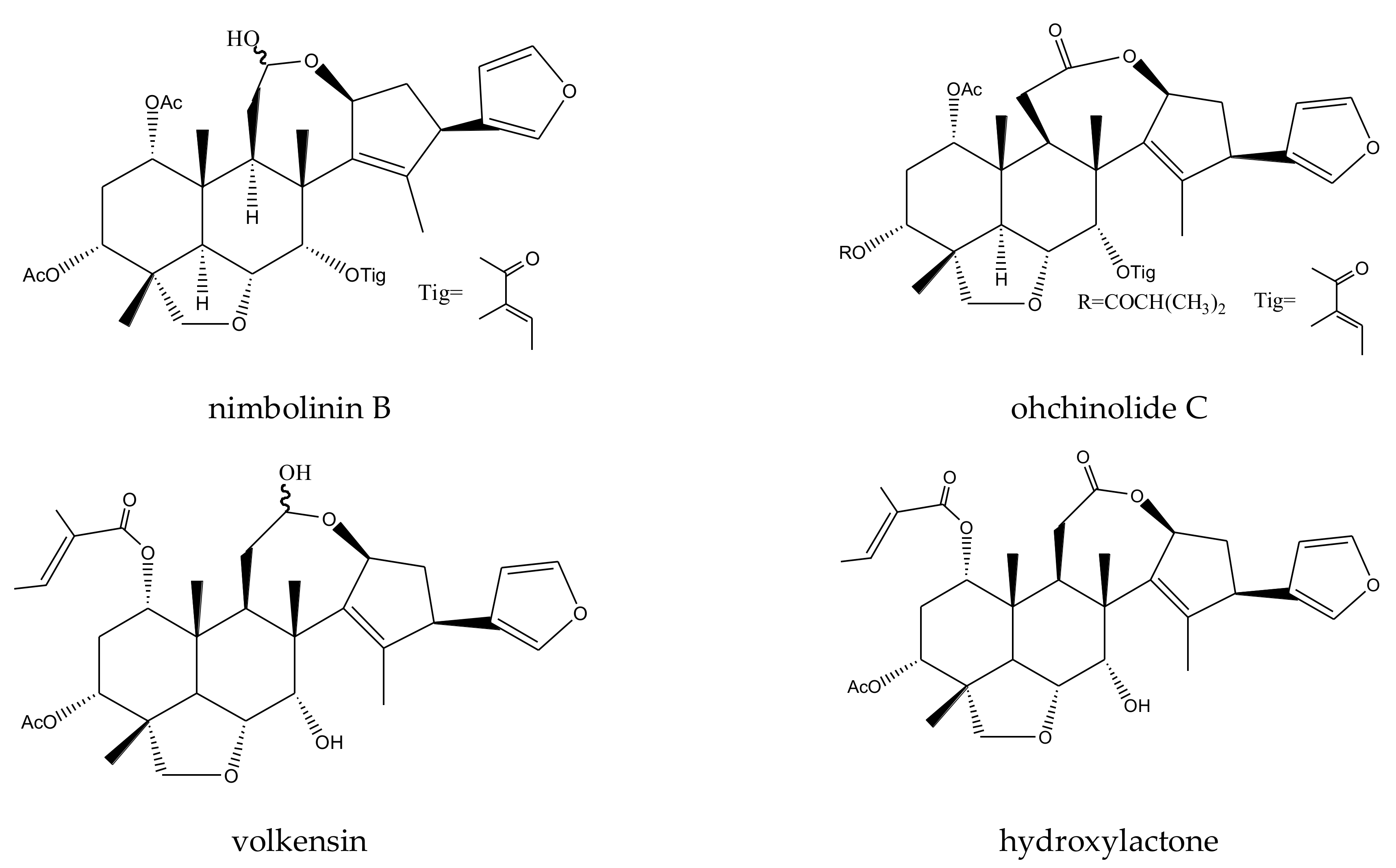
| nimbolinin B | Melia toosendan | Spodoptera eridania | MIC = 1000 μg/mL (2–24 h) | [37] |
| toosendanin | Melia azedarach Melia toosendan | Spodoptera eridania | MIC = 300 μg/mL (2–24 h) | [5,32,37,39,44,57,58] |
| Pieris rapae | AFC50 = 0.21 mM | |||
| Peridroma saucia | DC50 = 8.04 μg/cm2 | |||
| Helicoverpa armigera | FI50 = 56.6 μg/mL (6 h) | |||
| Epilachna paenulata | ED50 = 3.69 μg/cm2 (24 h) | |||
| Spodoptera littoralis | AFC50 = 200 μg/mL | |||
| Spodoptera litura | 100% antifeedant rate at 0.01% toosendanin | |||
| Spodoptera abyssinia | 76.5% antifeedant rate at 0.1% toosendanin | |||
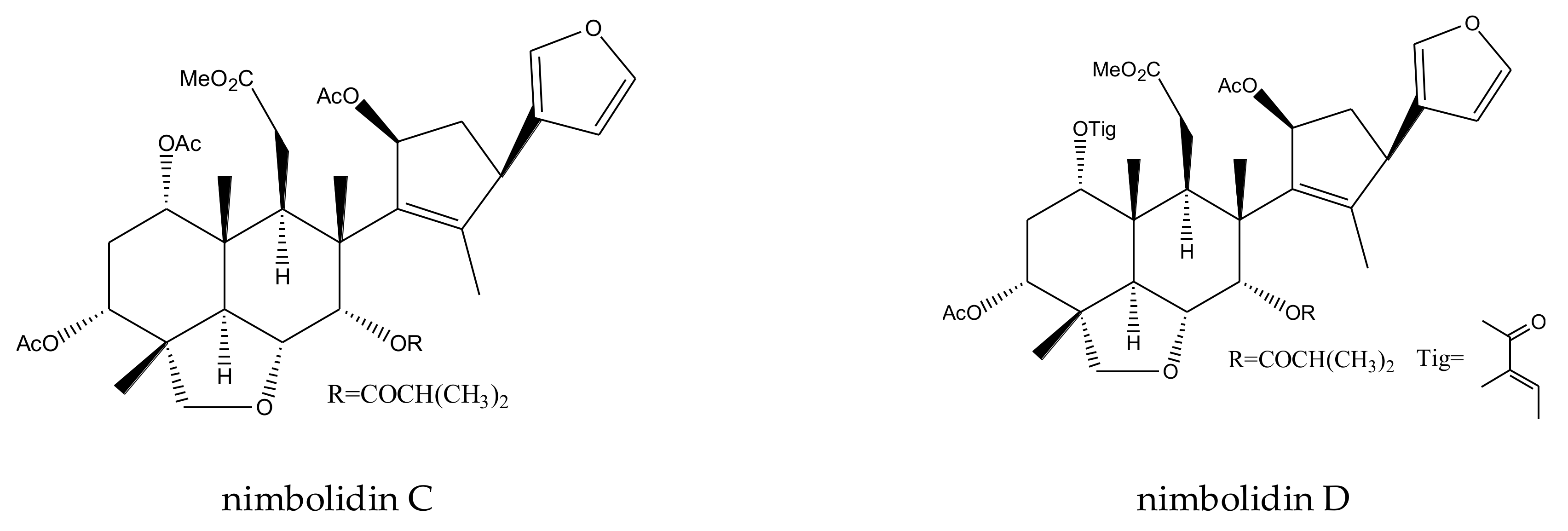
| nimbolidin C | Melia toosendan | Spodoptera eridania | MIC = 500 μg/mL (2–24 h) | [37] |
| nimbolidin D | Melia toosendan | Spodoptera eridania | MIC = 500 μg/mL (2–24 h) | [37] |
| nimbolidin E | Melia toosendan | Spodoptera eridania | MIC = 500μg/mL (2–24 h) | [37] |
| nimbolidin F | Melia toosendan | Spodoptera eridania | MIC = 500 μg/mL (2–24 h) | [37] |
| 3-O-acetylohchinolal | Melia toosendan | Spodoptera eridania | MIC = 1000 μg/mL (2–24 h) | [37] |
| ohchinolide C | Melia toosendan | Spodoptera eridania | MIC = 1000 μg/mL (2–24 h) | [37] |
| volkensin | Melia volkensii | Spodoptera frugiperda | ED50 = 3.5 μg/cm2 (15 h) | [29] |
| hydroxylactone | Melia volkensii | Spodoptera frugiperda | ED50 = 6 μg/cm2 (15 h) | [29] |
| 6-acetylsendanal | Melia toosendan | Pieris rapae | AFC50 = 1.32 mM | [44] |
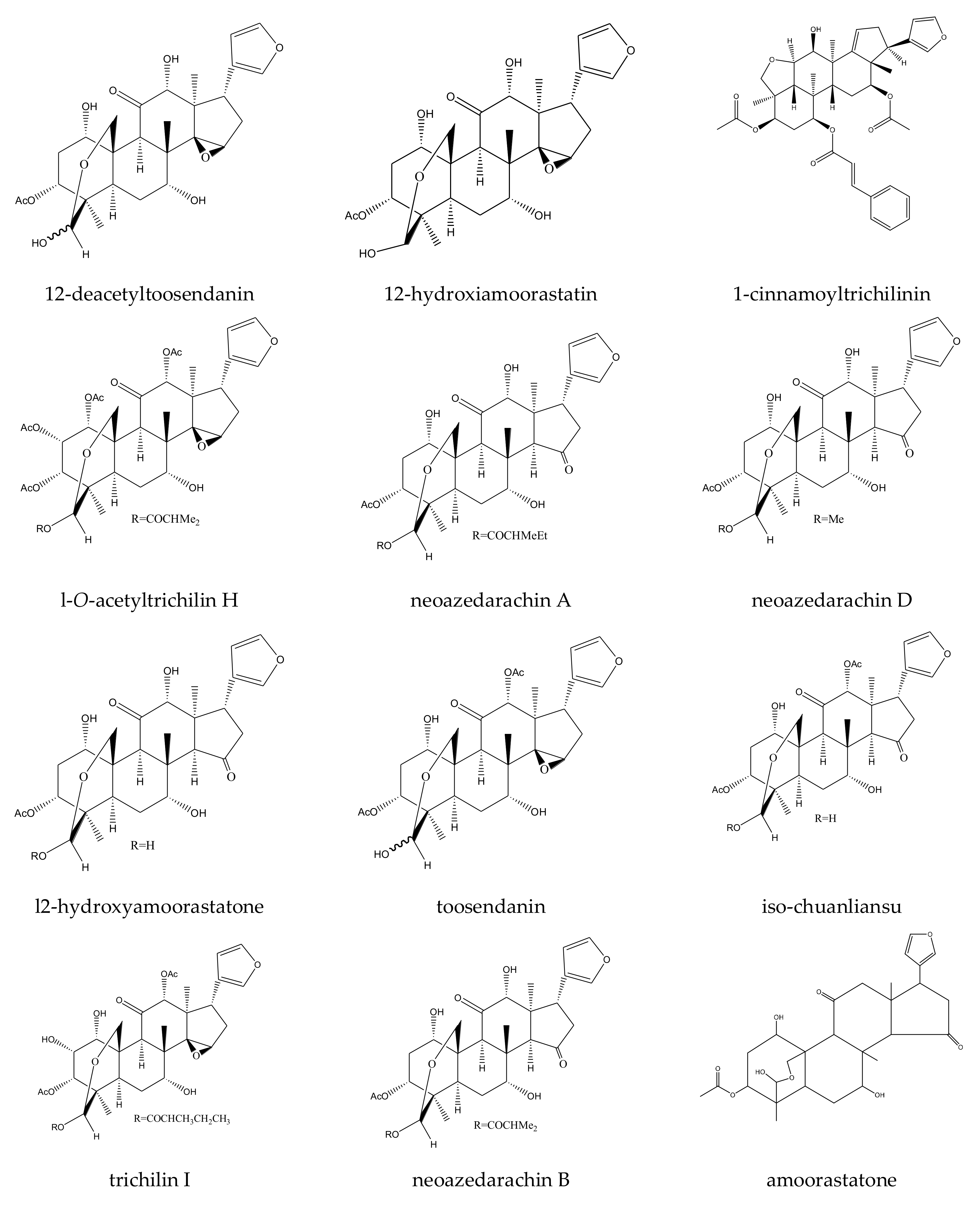
| iso-chuanliansu | Melia toosendan Melia toosendan | Pieris rapae | AFC50 = 0.46 mM | [44] |
| Spodoptera littoralis | MIC = 300 μg/mL (2–24 h) | [29] | ||
| amoorastatone | Melia toosendan | Pieris rapae | AFC50 = 0.63 mM | [44] |
| 12-hydroxyamoorastatone | Melia toosendan | Pieris rapae | AFC50 = 0.64 mM | [44] |
| mesendanin H | Melia toosendan | Pieris rapae | AFC50 = 0.11 mM | [44] |
| meliatoosenin E | Melia toosendan | Pieris rapae | AFC50 = 1.03 mM | [44] |
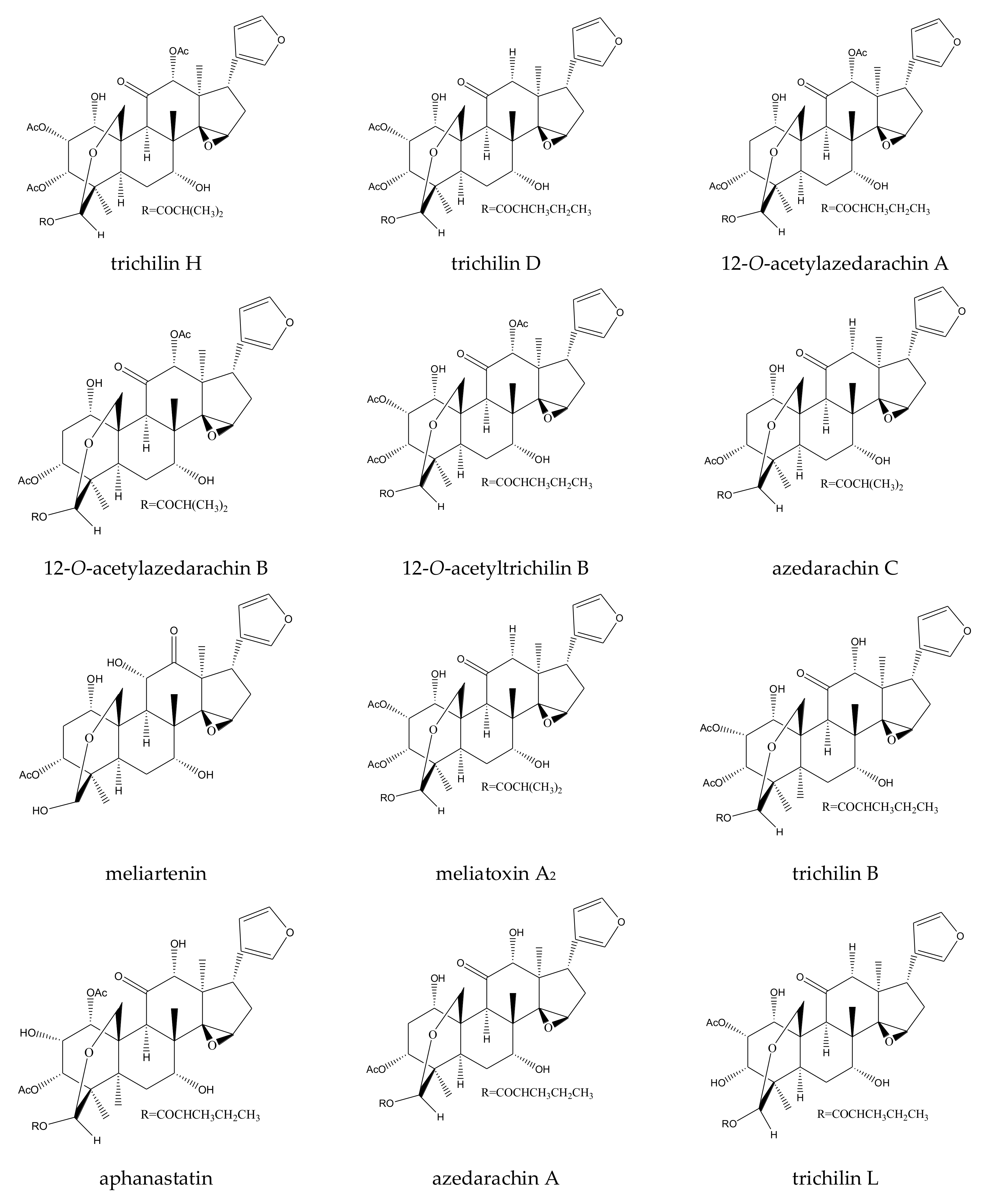
| trichilin B | Melia azedarach | Spodoptera exigua | MIC = 200 μg/mL (6–24 h) | [36] |
| aphanastatin | Melia azedarach | Spodoptera exigua | MIC = 200 μg/mL (6–24 h) | [36] |
| azedarachin A | Melia azedarach | Spodoptera exigua | MIC = 200 μg/mL (6–24 h) | [36] |
| 12-O-acetyltrichilin B | Melia azedarach | Spodoptera exigua | MIC = 400 μg/mL (6–24 h) | [36] |
| Spodoptera eridania | MIC = 400 μg/mL (6–24 h) | [37] | ||
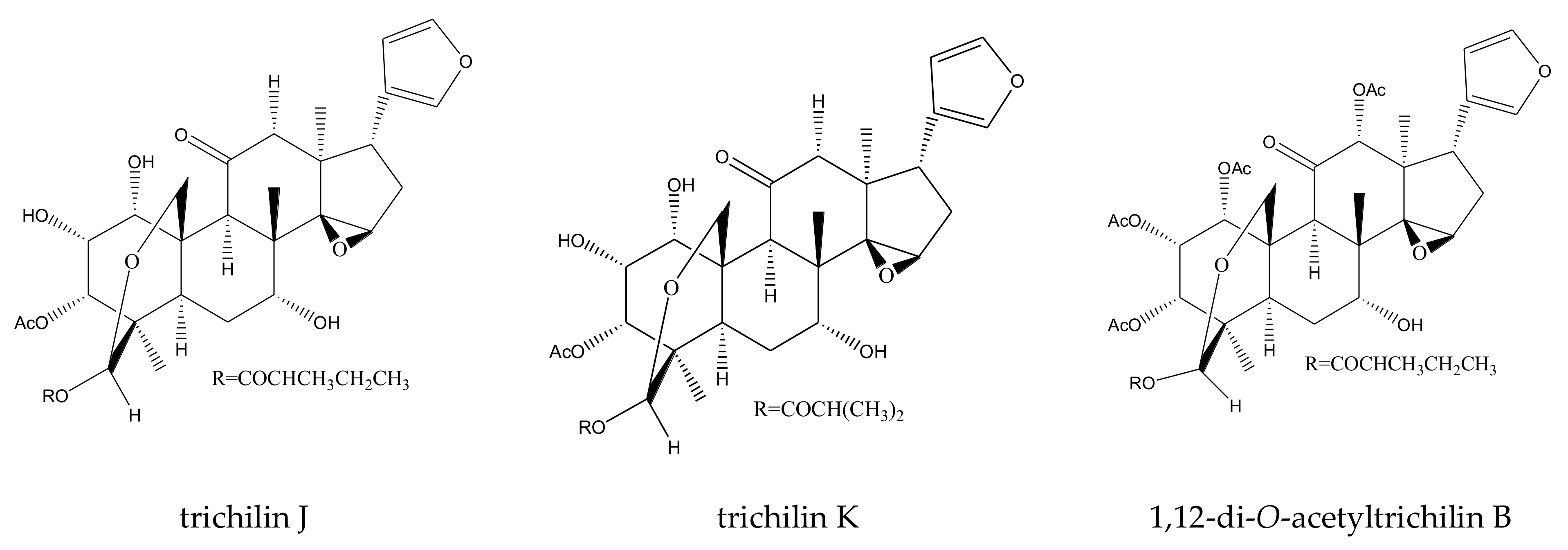
| 1,12-di-O-acetyltrichilin B | Melia azedarach | Spodoptera exigua | MIC = 400 μg/mL (6–24 h) | [36] |
| trichilin H | Melia azedarach | Spodoptera exigua | MIC = 400 μg/mL (6–24 h) | [36] |
| trichilin D | Melia azedarach | Spodoptera exigua | MIC = 400 μg/mL (6–24 h) | [36] |
| meliatoxin A2 | Melia azedarach | Spodoptera exigua | MIC = 400 μg/mL (6–24 h) | [36] |
| 12-O-acetylazedarachin A | Melia azedarach | Spodoptera exigua | MIC = 400 μg/mL (6–24 h) | [36] |
| 12-O-acetylazedarachin B | Melia azedarach | Spodoptera exigua | MIC = 400 μg/mL (6–24 h) | [36] |
| azedarachin C | Melia azedarach | Spodoptera exigua | MIC = 400 μg/mL (6–24 h) | [36] |
| trichilin I | Melia toosendan | Spodoptera eridania | MIC = 400 μg/mL (2–24 h) | [37] |
| trichilin J | Melia toosendan | Spodoptera eridania | MIC = 400 μg/mL (2–24 h) | [37] |
| trichilin K | Melia toosendan | Spodoptera eridania | MIC = 400 μg/mL (2–24 h) | [37] |
| trichilin L | Melia toosendan | Spodoptera eridania | MIC = 400 μg/mL (2–24 h) | [37] |
| 12-deacetyltoosendanin | Melia toosendan | Spodoptera eridania | MIC = 150 μg/mL (2–24 h) | [37] |
| Spodoptera littoralis | MIC = 250 μg/mL (2–24 h) | [29] | ||
| l-O-acetyltrichilin H | Melia toosendan | Spodoptera littoralis | MIC = 300 μg/mL (2–24 h) | [29] |
| neoazedarachin A | Melia toosendan | Spodoptera littoralis | MIC = 400 μg/mL (2–24 h) | [29] |
| neoazedarachin B | Melia toosendan | Spodoptera littoralis | MIC = 400 μg/mL (2–24 h) | [29] |
| neoazedarachin D | Melia toosendan | Spodoptera littoralis | MIC = 400 μg/mL (2–24 h) | [29] |
| meliartenin | Melia azedarach | Epilachna paenulata | ED50 = 0.80 µg/cm2 (24 h) | [39] |
| 12-hydroxia-moorastatin | Melia azedarach | Epilachna paenulata | ED50 = 0.80 µg/cm2 (24 h) | [39] |
| 1-cinnamoyltrichilinin | Melia volkensii | Spodoptera littoralis | MAC = 1000 μg/mL | [45] |
2.5.1. Ring-Intact Limonoids
| Compound | Plant Source | Insect | Activity | Ref. |
|---|---|---|---|---|
| cipadesin | Cipadessa fruticosa Cipadessa baccifera | Spodoptera frugiperda | S50 = 7 days at 100 μg/mL | [51] |
| febrifugin | Cipadessa fruticosa Cipadessa baccifera Cipadessa cinerascens | Spodoptera frugiperda | S50 = 6 days at 100 μg/mL | [51] |
| ruageanin A | Cipadessa fruticosa | Atta sexdens rubropilosa | S50 = 6 days at 100 μg/mL | [51] |
| swietemahonolide | Cipadessa fruticosa | Atta sexdens rubropilosa | S50 = 8 days at 100 μg/mL | [51] |
| mexicanolide | Cipadessa fruticosa | Atta sexdens rubropilosa | S50 = 6 days at 100 μg/mL | [51] |
| cipadesin A | Cipadessa fruticosa Cipadessa baccifera Cipadessa cinerascens | Spodoptera frugiperda | MR: less than 40% at 50 mg/kg | [47] |
| febrifugin A | Cipadessa fruticosa | Spodoptera frugiperda | MR: 73.3% at 50 mg/kg | [47] |
| khayasin T | Cipadessa fruticosa Cipadessa baccifera | Spodoptera frugiperda | MR: 50% at 50 mg/kg | [47] |
| melianodiol | Guarea grandiflora Guarea kunthiana | Aedes aegypti | LC50 = 14.44 μg/mL LC90 = 17.54 μg/mL (24 h) | [20] |
| melianone | Guarea grandiflora | Reticulitermes speratus | MR: 95% at 100 μg/disc (30 days) | [41] |
| gedunin | Entandrophragma angolense Entandrophragma delevoyi Entandrophragma macrophyllum Guarea grandiflora Khaya grandifoliola | Spodoptera frugiperda | LM: LC50 = 39 μg/mL (30 days) | [5,77,78] |
| methyl angolensate | Entandrophragma angolense Entandrophragma macrophyllum Guarea thompsonii Khaya anthotheca Khaya senegalensis Khaya grandifoliola Khaya ivoremis | Spodoptera frugiperda | MR: 40% at 50 mg/kg (7 days) | [50] |
| 7-deacetoxy-7-oxo -gedunin | Guarea grandiflora Guarea guidonia Carapa guianensis | Atta sexdens rubropilosa | S50 = 9 days at 100 μg/mL | [25] |
| anthothecol | Khaya anthotheca | Plutella xylostella | MR: 30–80% at 0.25–1 mg/mL (48 h) | [48] |
| Myzus persicae | MR: 30%, 60% at 0.5, 1 mg/mL (48 h) | |||
| toosendanin | Melia azedarach Melia toosendan | Sitophilus oryzae | LC50 = 675 μg/mL (6 weeks) | [5,49,195] |
| Cryptolestes ferrugineus | LC50 = 1875 μg/mL (6 weeks) | |||
| Ostrinia furnacalis | Mortality: 58.33% at 0.4 μg (average 3.1 days) | |||
| Spodoptera frugiperda | LC50 = 7.0 μg/mL | |||
| 1,3-dicinnamoyl-11- hydroxymeliacarpin | Melia azedarach | Spodoptera littoralis | LC50 = 2.36 μg/mL (12 days) | [52] |
| 1-cinnamoyl-3- methacryl-11-hydroxy-meliacarpin | Melia azedarach | Spodoptera littoralis | LC50 = 1.19 μg/mL (12 days) | [52] |
| 1-cinnamoyl-3-acetyl-11-hydroxymeliacarpin | Melia azedarach | Spodoptera littoralis | LC50 = 0.48 μg/mL (12 days) | [52] |
| 2’S-cipadesin A | Cipadessa baccifera | AChE | inhibitory activity (AChE) at 50 mM | [47] |
| granatumin E | Cipadessa baccifera | AChE | inhibitory activity (AChE) at 50 mM | [47] |
| 3-O-detigloyl-3-O-isobutyrylfebrifugin A | Cipadessa baccifera | AChE | inhibitory activity (AChE) at 50 mM | [47] |
| cipadonoid B | Cipadessa baccifera Cipadessa cinerascens | insect nAChR | pI50 = 4.2 | [47] |
| khayasin | Cipadessa baccifera | Brontispa longissima | LC50 = 7.28 μg/mL (24 h) | [53] |
| Compound | Plant Source | Insect | Activity | Ref. |
|---|---|---|---|---|
| cipadesin | Cipadessafruticosa Cipadessa baccifera | Spodoptera frugiperda | LPE: 0.8 days | [43] |
| febrifugin | Cipadessa fruticosa Cipadessa baccifera Cipadessa cinerascens | Spodoptera frugiperda | LPE: 1.8 days | [43] |
| khayasin T | Cipadessa fruticosa Cipadessa baccifera | Spodoptera frugiperda | LPE: 1.2 days | [43] |
| cipadesin A | Cipadessa fruticosa Cipadessa baccifera Cipadessa cinerascens | Spodoptera frugiperda | LPE: 2.1 days | [43] |
| prieurianin | Entandrophragma candolei Guarea guidonia | Helicoverpa armigera | GIL, EC50 = 18.8 μg/mL (7 days) | [32] |
| epoxyprieurianin | Entandrophragma candolei | Helicoverpa armigera | GIL, EC50 = 3.2 μg/mL (7 days) | [32] |
| prieurianin acetate | Entandrophragma candolei | Helicoverpa armigera | GIL, EC50 = 11.5 μg/mL (7 days) | [32] |
| epoxyprieurianin acetate | Entandrophragma candolei | Helicoverpa armigera | GIL, EC50 = 2.6 μg/mL (7 days) | [32] |
| 6α-acetoxy-gedunin | Guarea kunthiana Guarea grandiflora | Ostrinia nubilalis | Reduced growth at 50 μg/mL | [18,56] |
| 3β-O-tigloylmelianol | Guarea kunthiana | Rhipicephalus microplus | GSI reduced 50% (48 h) | [21] |
| 7-deacetoxy-7-oxogedunin | Guarea grandiflora Guarea guidonia Carapa guianensis | Spodoptera frugiperda | LPE: 1.2 days | [50] |
| 1-O-acetylkhayanoilde B | Khaya senegalensis | Spodoptera littoralis | GI, EC50 = 16.75 mg/kg (7 days) | [54] |
| khyanolide A | Khaya senegalensis | Spodoptera littoralis | GI, EC50 = 14.65 mg/kg (7 days) | [54] |
| khyanolide B | Khaya senegalensis | Spodoptera littoralis | GI, EC50 = 6.96 mg/kg (7 days) | [54] |
| khayalactol | Khaya senegalensis | Spodoptera littoralis | GI, EC50 = 11.48 mg/kg (7 days) | [54] |
| toosendanin | Melia azedarach Melia toosendan | Peridroma saucia | EC50 = 42.3 μg/mL after 7 days | [55,175,194] |
| Ostrinia furnacalis | Inhibition of body weight and pupation | |||
| Spodoptera frugiperda | Inhibition of body weight |
2.5.2. Ring C-Seco Limonoids
3. Structures and Structure–Activity Relationship (SAR) of the Insecticidal Chemicals
3.1. Structures of the Insecticidal Chemicals
3.2. Structure–Activity Relationship (SAR) of the Insecticidal Chemicals
4. Insecticidal Mechanism of Action
5. Environmental Toxicity
| Chemicals or Plant Extracts | Mice | Aquatic Organisms | Natural Enemies | Ref. |
|---|---|---|---|---|
| methyl angolensate | inhibition of smooth muscle, decrease of propulsive action of the gastrointestinal tract | - | - | [225] |
| toosendanin | serious hepatotoxicity, pregnancy-toxic | cytoplasmic vacuolation and nuclear shrinkage in liver of zebrafish | - | [226,227,228] |
| neoazedarachin B | - | low toxicity to brine shrimp, LC50 = 0.0059 μg/mL (48 h) | - | [215] |
| methanol extracts of K. ivorensis | relatively toxic, LD50 = 549 mg/kg | - | - | [220] |
| ethanolic extract of K. senegalensis | adverse effect on liver and kidney | - | - | [221,222,223] |
| chloroform fraction of M. azedarach | potent pregnancy interceptive property | - | - | [224] |
| andiroba oil (C. guianensis oil) | not toxic at 2000 mg/kg (14 d) | - | - | [214] |
| water extract of K. grandifoliola | adverse effect on bone minerals (at 500 mg/kg) | - | - | [219] |
| aqueous leaf extracts of M. azedarach | - | - | no direct negative effects on the survival and foraging of Cotesia plutellae, Diadromus collaris, Trichogramma evanescens, Aphidius ervi, Aphidius colemani, Eretmocerus eremicus and Encarsia formosa | [138,139] |
| M. azaderach unripe fruit extracts | - | - | no detrimental effects on Eriopis connexa | [216] |
6. Future Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Sun, Z.Q.; Lv, M.; Huang, W.W.; Li, T.; Xu, H. Development of botanical pesticides: Exploration on the phenotype of vestigial wings of insect pests induced by plant natural products or their derivatives by blocking tyrosine phosphorylation of insulin receptor 1. J. Agric. Food Chem. 2022, 70, 2117–2126. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, R.A.; Camara, M.C.; Oliveira, J.L.; Carvalho, L.B.; Proença, P.L.F.; Casagrande, M.G.; Lima, R.; Nascimento, J.; Gonçalves, K.C.; Gonçalves, R.A.; et al. Zein based-nanoparticles loaded botanical pesticides in pest control: An enzyme stimuli-responsive approach aiming sustainable agriculture. J. Hazard. Mater. 2021, 417, 126004. [Google Scholar] [CrossRef] [PubMed]
- Stephen, O.D.; Antonio, E.; Maurizio, V. Natural products in pest management: Innovative approaches for increasing their use. Pest Manag. Sci. 2019, 75, 2299–2300. [Google Scholar]
- Lorsbach, B.A.; Sparks, T.C.; Cicchillo, R.M.; Garizi, N.V.; Hahn, D.R.; Meyer, K.G. Natural products: A strategic lead generation approach in crop protection discovery. Pest Manag. Sci. 2019, 75, 2301–2309. [Google Scholar] [CrossRef] [PubMed]
- Tan, Q.G.; Luo, X.D. Meliaceous limonoids: Chemistry and biological activities. Chem. Rev. 2011, 111, 7437–7522. [Google Scholar] [CrossRef]
- Fan, W.X.; Fan, L.H.; Wang, Z.T.; Yang, L. Limonoids from the genus Melia (Meliaceae): Phytochemistry, synthesis, bioactivities, pharmacokinetics, and toxicology. Front. Pharmacol. 2021, 12, 795565. [Google Scholar] [CrossRef]
- Lin, M.H.; Yang, S.F.; Huang, J.G.; Zhou, L.J. Insecticidal triterpenes in meliaceae: Plant species, molecules and activities: Part I (Aphanamixis-Chukrasia). Int. J. Mol. Sci. 2021, 22, 13262. [Google Scholar] [CrossRef]
- Yu, J.H.; Zhang, H.; Zhou, B.; Zimbres, F.M.; Dalal, S.; Liu, Q.F.; Cassera, M.B.; Yue, J.M. Limonoids from Cipadessa baccifera. J. Nat. Prod. 2020, 83, 1751–1765. [Google Scholar] [CrossRef]
- Yu, J.H.; Zhou, B.; Dalal, S.; Liu, Q.F.; Cassera, M.B.; Yue, J.M. Cipaferoids A–C, three limonoids represent two different scaffolds from Cipadessa baccifera. Chin. J. Chem. 2018, 36, 124–128. [Google Scholar] [CrossRef]
- Yakusu, E.K.; Monthe, F.S.; Bourland, N.; Hardy, O.J.; Louppe, D.; Lokanda, F.B.M.; Hubau, W.; Muhongya, J.M.K.; Bulcke, J.V.D.; Acker, J.V.; et al. Entandrophragma: Taxonomy and ecology of a genus of African tree species with economic importance. A review. Biotechnol. Agron. Soc. Environ. 2018, 22, 113–127. [Google Scholar] [CrossRef]
- Arnason, J.T.; Philogène, B.J.R.; Donskov, N.; Kubo, I. Limonoids from the meliaceae and rutaceae reduce feeding, growth and development of Ostrinia nubilalis. Entomol. Exp. Appl. 1987, 43, 221–226. [Google Scholar] [CrossRef]
- Eckebil, P.P.T.; Verheggen, F.; Doucet, J.L.; Malaisse, F.; Daïnou, K.; Cerutti, P.O.; Vermeulen, C. Entandrophragma cylindricum (Sprague) Sprague (Meliaceae), a competitive tree species in central Africa. A review. Biotechnol. Agron. Soc. Environ. 2017, 21, 80–97. [Google Scholar] [CrossRef]
- Mulholland, D.A.; Schwikkard, S.L.; Sandor, P.; Nuzillard, J.M. Delevoyin C, a tetranortriterpenoid from Entandrophragma delevoyi. Phytochemistry 2000, 53, 465–468. [Google Scholar] [CrossRef]
- Zhang, W.Y.; An, F.L.; Zhou, M.M.; Chen, M.H.; Jian, K.L.; Quasie, O.; Yang, M.H.; Luo, J.; Kong, L.Y. Limonoids with diverse frameworks from the stem bark of Entandrophragma angolense and their bioactivities. RSC Adv. 2016, 6, 97160–97171. [Google Scholar] [CrossRef]
- Mulholland, D.A.; Osborne, R.; Roberts, S.L.; Taylor, D.A.H. Limonoids and triterpenoid acids from the bark of Entandrophragma delevoyi. Phytochemistry 1994, 37, 1417–1420. [Google Scholar] [CrossRef]
- Connolly, J.D.; Phillips, W.R.; Mulholland, D.A.; Taylor, D.A.H. Spicatin, a protolimonoid from Entandrophragma spicatum. Phytochemistry 1981, 20, 2596–2597. [Google Scholar] [CrossRef]
- Lukacova, V.; Polonsky, J.; Moretti, C.; Pettit, G.R.; Schmidt, J.M. Isolation and structure of 14,15β-epoxyprieurianin from the south american tree Guarea guidona. J. Nat. Prod. 1982, 45, 288–294. [Google Scholar] [CrossRef]
- Jimenez, A.; Villarreal, C.; Toscano, R.A.; Cook, M.; Arnason, J.T.; Bye, R.; Mata, R. Limonoids from Swietenia humilis and Guarea grandiflora (Meliaceae). Phytochemistry 1998, 49, 1981–1988. [Google Scholar] [CrossRef]
- Housley, J.R.; King, F.E.; King, T.J.; Taylor, P.R. The chemistry of hardwood extractives. Part XXXIV. Constituents of Guarea species. J. Chem. Soc. 1962, 5095–5104. [Google Scholar] [CrossRef]
- Sarmento, U.C.; Miguita, C.H.; Almeida, L.M.G.E.; Souza, A.S.; Garcez, W.S.; Garcez, F.R. Larvicidal efficacies of plants from midwestern Brazil: Melianodiol from Guarea kunthiana as a potential biopesticide against Aedes aegypti. Mem. Inst. Oswaldo Cruz 2016, 111, 469–474. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, C.D.S.; Borges, L.M.F.; Louly, C.C.B.; Rocha, T.L.; Morais, S.M.T.S.; Miguita, C.H.; Garcez, W.S.; Garcez, F.R. In vitro activity of 3β-O-tigloylmelianol from Guarea kunthiana A. Juss (Meliaceae) on oogenesis and ecdysis of the cattle tick Rhipicephalus (Boophilus) microplus (Canestrini) (Acari: Ixodidae). Exp. Parasitol. 2016, 164, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Csdiz-Gurrea, M.D.; Sinan, K.I.; Zengin, G.; Bene, K.; Etienne, O.K.; Leyva-Jiménez, F.J.; Fernandez-Ochoa, A.; Villegas-Aguilar, M.C.; Mahomoodally, M.F.; Lobine, D.; et al. Bioactivity assays, chemical characterization, ADMET predictions and network analysis of Khaya senegalensis A. Juss (Meliaceae) extracts. Food Res. Int. 2021, 139, 109970. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Tan, J.; Vanderveer, D.; Wang, X.; Wargovich, M.J.; Chen, F. Khayanolides from African mahogany Khaya senegalensis (Meliaceae): A revision. Phytochemistry 2009, 70, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Mukaila, Y.O.; Ajao, A.N.; Moteetee, A.N. Khaya grandifoliola CDC. (Meliaceae: Sapindales): Ethnobotany, phytochemistry, pharmacological properties, and toxicology. J. Ethnopharmacol. 2021, 278, 114253. [Google Scholar] [CrossRef]
- Ambrozin, A.R.; Leite, A.C.; Bueno, F.C. Limonoids from andiroba oil and Cedrela fissilis and their insecticidal activity. J. Braz. Chem. Soc. 2006, 17, 542–547. [Google Scholar] [CrossRef] [Green Version]
- Yuan, T.; Zhang, C.R.; Yang, S.P.; Yue, J.M. Limonoids and triterpenoids from Khaya senegalensis. J. Nat. Prod. 2010, 73, 669–674. [Google Scholar] [CrossRef]
- Chiffelle, T.; Huerta, A.; Bobadilla, V.; Macuada, G.; Ceballos, R. Antifeedant and insecticidal effects of extracts from Melia azedarach fruits and Peumus boldus leaves on Xanthogaleruca luteola larvae. Chil. J. Agric. Res. 2019, 79, 609–615. [Google Scholar] [CrossRef] [Green Version]
- Nakatani, M.; Zhou, J.B.; Minami, Y.; Yagi, F.; Tadera, K. Antifeeding limonoids from Melia toosendan. Heterocycles 1997, 45, 1781–1786. [Google Scholar] [CrossRef]
- Rajab, M.S.; Bentley, M.D.; Alford, A.R.; Mendel, M.J. A new limonoid insect antifeedant from the fruit of Melia volkensii. J. Nat. Prod. 1988, 51, 168–171. [Google Scholar] [CrossRef]
- Koul, O.; Multani, J.S.; Singh, G.; Wahab, S. Bioefficacy of toosendanin from Melia dubia (syn. M. Azedarach) against gram pod-borer, Helicoverpa armigera (Hübner). Curr. Sci. India 2002, 83, 1387–1391. [Google Scholar]
- Kipassa, N.T.; Iwagawa, T.; Okamura, H.; Doe, M.; Morimoto, Y.; Nakatani, M. Limonoids from the stem bark of Cedrela odorata. Phytochemistry 2008, 69, 1782–1787. [Google Scholar] [CrossRef] [PubMed]
- Koul, O.; Daniewski, W.M.; Multani, J.S.; Maria, G.; Gurmeet, S. Antifeedant effects of the limonoids from Entandrophragma candolei (Meliaceae) on the gram pod borer, Helicoverpa armigera (Lepidoptera: Noctuidae). J. Agric. Food Chem. 2003, 51, 7271–7275. [Google Scholar] [CrossRef] [PubMed]
- Nakatani, M.; Abdelgaleil, S.A.; Kassem, S.M.; Takezaki, K.; Okamura, H.; Iwagawa, T.; Doe, M. Three new modified limonoids from Khaya senegalensis. J. Nat. Prod. 2002, 65, 1219–1221. [Google Scholar] [CrossRef] [PubMed]
- Abdelgaleil, S.A.M.; Nakatani, M. Antifeeding activity of limonoids from Khaya senegalensis (Meliaceae). J. Appl. Entomol. 2003, 127, 236–239. [Google Scholar] [CrossRef]
- Mootoo, B.S.; Ali, A.; Motilal, R.; Pingal, R.; Mclean, S. Limonoids from Swietenia macrophylla and S. aubrevilleana. J. Nat. Prod. 1999, 62, 1514–1517. [Google Scholar] [CrossRef]
- Munehiro, N.; Huang, R.C.; Hiroaki, O.; Iwagawa, T.; Tadera, K.; Naokic, H. Three new antifeeding meliacarpinins from Chinese Melia azedarach Linn. Tetrahedron 1995, 51, 11731–11736. [Google Scholar]
- Huang, C.R.; Zhou, J.B.; Suenaga, H.; Takezaki, K.; Nakatani, M. Insect antifeeding property of limonoids from Okinawan and Chinese Melia azedarach L., and from Chinese Melia toosendan (Meliaceae). Biosci. Biotechnol. Biochem. 1995, 59, 1755–1757. [Google Scholar] [CrossRef]
- Zhou, J.B.; Tadera, K.; Minami, Y.; Wang, Z.T.; Bo, Y.Y. New limonoids from Melia toosendan. Biosci. Biotechnol. Biochem. 1998, 62, 496–500. [Google Scholar] [CrossRef] [Green Version]
- Carpinel, C.; Defagó, M.T.; Valladares, G.; Palacios, S.M. Antifeedant and insecticide properties of a limonoid from Melia azedarach (Meliaceae) with potential use for pest management. J. Agric. Food Chem. 2003, 51, 369–374. [Google Scholar] [CrossRef]
- Suresh, G.; Gopalakrishnan, G.; Wesley, D.; Singh, N.D.P.; Malathi, R.; Rajan, S.S. Insect antifeedant activity of the tetranortriterpenoids from the rutales. A perusal of structural relations. J. Agric. Food Chem. 2002, 50, 4484–4490. [Google Scholar] [CrossRef]
- Hui, S.R.; Mujo, K.; Takehiko, Y.; Shozo, T. Antifeeding constituents of Phellodendron chinense fruit against Reticulitermes speratus. J. Pestic. Sci. 1990, 15, 567–572. [Google Scholar]
- Fu, L.R.; Ma, Q.Y.; Huang, S.Z.; Dai, H.F.; Guo, Z.K.; Yu, Z.F.; Zhao, Y.X. Terpenoids and their anti-feedant activity from Cipadessa cinerascens. J. Asian Nat. Prod. Res. 2015, 16, 1054–1059. [Google Scholar] [CrossRef] [PubMed]
- Matos, A.P.; Leite, A.C.; Batista-Pereira, L.G.; Vieira, P.C.; Fernandes, J.B.; Silva, M.F. Effects of limonoids from Cipadessa fruticosa on fall armyworm. J. Biosci. 2009, 64, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Dong, H.Y.; He, Q.M.; Liang, J.L.; Zhou, L. Characterization of limonoids isolated from the fruits of Melia toosendan and their antifeedant activity against Pieris rapae. Chem. Biodivers. 2020, 17. [Google Scholar] [CrossRef]
- Jaoko, V.; Taning, C.N.; Backx, S.; Mulatya, J.; Abeele, J.V.; Magomere, T.; Olubayo, F.; Mangelinckx, S.; Werbrouck, S.P.O.; Smagghe, G. The phytochemical composition of Melia volkensii and its potential for insect pest management. Plants 2020, 9, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gervais, M.H.; Bonaventure, T.N.; Robert, G.I.; Simeon, F.K. Phytochemistry and pharmacology of the genus Entandrophragma over the 50 years from 1967 to 2018: A ‘golden’ overview. J. Pharm. Pharmacol. 2018, 70, 1431–1460. [Google Scholar]
- Bandi, A.K.R.; Lee, D. Secondary metabolites of plants from the genus Cipadessa: Chemistry and biological activity. Chem. Biodivers. 2012, 9, 1403–1421. [Google Scholar] [CrossRef] [PubMed]
- Park, E.S.; Bae, I.K.; Jeon, H.J.; Lee, S.E. Limonoid derivatives and its pesticidal activities. Entomol. Res. 2014, 44, 158–162. [Google Scholar] [CrossRef]
- Xie, Y.S.; Fields, P.G.; Isman, M.B.; Chen, W.K.; Zhang, X. Insecticidal activity of Melia toosendan extracts and toosendanin against three stored-product insects. J. Stored Prod. Res. 1995, 31, 259–265. [Google Scholar] [CrossRef]
- Sarria, A.L.F.; Soares, M.S.; Matos, A.P.; Fernandes, J.B.; Vieira, P.C.; Silva, M.F.G.F. Effect of triterpenoids and limonoids isolated from Cabralea canjerana and Carapa guianensis (Meliaceae) against Spodoptera frugiperda (JE Smith). Z. Naturforsch. C 2011, 66, 245–250. [Google Scholar] [CrossRef]
- Leite, A.C.; Bueno, F.C.; Oliveira, C.G.; Fernandes, J.B.; Vieira, P.C.; Silva, M.F.G.F.; Bueno, O.C.; Pagnocca, F.C.; Hebling, M.J.A.; Bacci, M., Jr. Limonoids from Cipadessa fruticosa and Cedrela fissilis and their insecticidal activity. J. Braz. Chem. Soc. 2005, 16, 1391–1395. [Google Scholar] [CrossRef] [Green Version]
- Bohnenstengel, F.I.; Wray, V.; Witte, L.; Srivastava, R.P.; Proksch, P. Insecticidal meliacarpins (C-seco limonoids) from Melia azedarach. Phytochemistry 1999, 50, 977–982. [Google Scholar] [CrossRef]
- Li, M.Y.; Zhang, J.; Feng, G.; Satyanandamurty, T.; Wu, J. Khayasin and 2′S-methylbutanoylproceranolide: Promising candidate insecticides for the control of the coconut leaf beetle, Brontispa longissima. Balk. J. Geom. Appl. 2011, 36, 22–26. [Google Scholar] [CrossRef] [Green Version]
- El-Aswad, A.F.; Abdelgaleil, S.A.; Nakatani, M. Feeding deterrent and growth inhibitory properties of limonoids from Khaya senegalensis against the cotton leafworm, Spodoptera littoralis. Pest Manag. Sci. 2004, 60, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Isman, M.B.; Chiu, S.F. Antifeedant and growth inhibitory effects of the limonoid toosendanin and extracts on the variegated cutworm, (Lep., Noctuidae). J. Appl. Eentomol. 2010, 119, 367–370. [Google Scholar] [CrossRef]
- Hofer, M.; Greger, H.; Mereiter, K. 6α-Acetoxygedunin. Acta Crystallogr. E 2009, 65, 1942–1943. [Google Scholar] [CrossRef] [PubMed]
- Chiu, S.F.; Zhang, X. A critical review of toosendanin a novel insecticide isolated from Melia toosendan. J. South China Agric. Univ. 1987, 8, 57–67. [Google Scholar]
- Zhang, X.; Wang, X.L.; Feng, J.T.; Chiu, S.F. The development of botanical insecticides-toosendanin. Acta Univ. Agric. Boreali-Occident. 1993, 21, 1–5. [Google Scholar]
- Ramkumar, G.; Karthi, S.; Muthusamy, R.; Natarajan, D.; Shivakumar, M.S. Adulticidal and smoke toxicity of Cipadessa baccifera (Roth) plant extracts against Anopheles stephensi, Aedes aegypti, and Culex quinquefasciatus. Parasitol. Res. 2014, 114, 167–173. [Google Scholar] [CrossRef]
- Ramkumar, G.; Karthi, S.; Shivakumar, M.S.; Kweka, E.J. Culex quinquefasciatus egg membrane alteration and ovicidal activity of Cipadessa baccifera (Roth) plant extracts compared to synthetic insect growth regulators. Res. Rep. Trop. Med. 2019, 10, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Leite, A.C.; Oliveira, C.G.; Bacci, M. Toxicity of Cipadessa fruticosa to the leaf-cutting ants Atta sexdens rubropilosa (Hymenoptera: Formicidae) and their symbiotic fungus. Sociobiology 2005, 46, 17–26. [Google Scholar]
- Malarvannan, S.; Sekar, S.; Subashini, H.D. Efficacy of Cipadessa baccifera leaf extracts against gram pod borer, Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). J. Entomol. Res. 2008, 32, 19–25. [Google Scholar]
- Gan, L.S.; Wang, X.N.; Wu, Y.; Yue, J.M. Tetranortriterpenoids from Cipadessa baccifera. J. Nat. Prod. 2007, 70, 1344–1347. [Google Scholar] [CrossRef]
- Faber, J.M.; Eger, W.A.; Williams, C.M. Enantioselective total synthesis of the mexicanolides: Khayasin, proceranolide, and mexicanolide. J. Org. Chem. 2012, 77, 8913–8921. [Google Scholar] [CrossRef]
- Fang, X.; Di, Y.T.; Li, C.S.; Geng, Z.L.; Zhang, Z.; Zhang, Y.; Lu, Y.; Zheng, Q.T. Tetranortriterpenoids from the leaves of Cipadessa cinerascens. J. Nat. Prod. 2009, 72, 714–718. [Google Scholar] [CrossRef]
- Yuan, Y.H.; Li, B.G.; Zhou, M.; Qi, H.Y.; Zhang, G.L. Cipadesins A–C: Novel tetranortriterpenoids from Cipadessa cinerascens. Org. Lett. 2005, 7, 5051–5053. [Google Scholar] [CrossRef]
- Fang, X.; Zhang, Q.; Tan, C.J.; Zhen, M.S.; Lu, Y.; Lu, Y.B.; Zheng, Q.T.; Di, Y.T.; Hao, X.J. Cipadonoids B–G, six new limonoids from Cipadessa cinerascens. Tetrahedron 2009, 65, 7408–7414. [Google Scholar] [CrossRef]
- Lin, L.G.; Tang, C.P.; Ke, C.Q.; Zhang, Y.; Ye, Y. Terpenoids from the stems of Cipadessa baccifera. J. Nat. Prod. 2008, 71, 628–632. [Google Scholar] [CrossRef]
- Luo, X.D.; Wu, S.H.; Ma, Y.B.; Wu, D.G. Tirucallane triterpenoids from Dysoxylum hainanense. Phytochemistry 2000, 54, 801–805. [Google Scholar] [CrossRef]
- Marpaung, L.; Nakamura, N.; Kakuda, H.; Hattori, M. Absolute configuration of cipadesin and febrifugin, from the seeds of Cipadessa baccifera. Nat. Med. 2001, 55, 220. [Google Scholar]
- Leite, A.C.; Neto, A.P.; Ambrozin, A.R.P.; Fernandes, J.B.; Vieira, P.C.; Silva, M.F.G.F.; Albuquerque, S. Trypanocidal activity of flavonoids and limonoids isolated from Myrsinaceae and Meliaceae active plant extracts. Rev. Brar. Farmacogn. 2010, 20, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Faber, J.M.; Williams, C.M. A concise total synthesis of (±)-cipadonoid B from synthetic azedaralide. Chem. Commun. 2011, 47, 2258–2260. [Google Scholar] [CrossRef] [PubMed]
- Akisanya, A.; Bevan, C.W.L.; Hirst, J.; Halsall, T.G.; Taylor, D.A.H. West African timbers. Part III. Petroleum extracts from the genus Entandrophragma. J. Chem. Soc. 1960, 758, 3827–3829. [Google Scholar] [CrossRef]
- Daniewski, W.M.; Gumulka, M.; Anczewski, W.; Truszewska, D.; Bloszyk, E.; Drozdz, B. Constituents of some Meliaceae plants and their antifeedant activity. Limonoids and sesquiterpenoids. Pol. J. Chem. 1996, 70, 1265–1269. [Google Scholar]
- Adesida, G.A.; Taylor, D.A.H. The chemistry of the genus Entandrophragma. Phytochemistry 1967, 6, 1429–1433. [Google Scholar] [CrossRef]
- Pettit, G.; Barton, G.H.R.; Herald, C.I.; Polonsky, J.; Schmidt, J.M.; Connolly, J.D. Evaluation of limonoids against the murine P388 lymphocytic leukemia cell line. J. Nat. Prod. 1983, 46, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Ishida, M.; Serit, M.; Nakata, K. Several antifeedants from neem oil, Azadirachta indica A. Juss. against Reticulitermes speratus Kolbe (Isoptera: Rhinotermitidae). Biosci. Biotech. Bioch. 2014, 56, 1835–1838. [Google Scholar] [CrossRef] [Green Version]
- Koul, O.; Multani, J.S.; Singh, G. 6β-hydroxygedunin from Azadirachta indica. Its potentiation effects with some non-azadirachtin limonoids in neem against lepidopteran larvae. J. Agric. Food Chem. 2003, 51, 2937–2942. [Google Scholar] [CrossRef] [PubMed]
- Lago, J.H.G.; Claudia, B.B.; Roque, N.F. Terpenoids from Guarea guidonia. Phytochemistry 2002, 60, 333–338. [Google Scholar] [CrossRef]
- Jessica, A.P.; Fabiana, G.; Mayara, C.S.; Luis, F.A.A.; Claudecir, C.M. Antimicrobial, insecticidal, and antioxidant activity of essential oil and extracts of Guarea kunthiana A. Juss. J. Med. Plant Res. 2015, 9, 48–55. [Google Scholar] [CrossRef] [Green Version]
- Achnine, L.; Mata, R.; Lotina-Hennsen, B. Interference of the natural product 7-oxo-7-deacetoxygedunin with CF0 of H+-ATPase of Spinach Chloroplasts. Pestic. Biochem. Phys. 1999, 63, 139–149. [Google Scholar] [CrossRef]
- Bueno, F.C.; Godoy, M.P.; Leite, A.C.; Bueno, O.C.; Pagnocca, F.C.; Fernandes, J.B.; Hebling, M.J.A.; Bacci, J.M.; Vieira, P.C.; Silva, M.F.G.F. Toxicity of Cedrela fissilis to Atta sexdens rubropilosa (Hymenoptera: Formicidae) and its symbiotic fungus. Sociobiology 2005, 45, 389–399. [Google Scholar]
- Okieimen, F.E.; Eromosele, C.O. Fatty acid composition of the seed oil of Khaya senegalensis. Bioresour. Technol. 1999, 69, 279–280. [Google Scholar] [CrossRef]
- Adedeji, G.A.; Ogunsanwo, O.Y.; Eguakun, F.S.; Elufioye, O.T. Chemical composition and termiticidal activity of Khaya ivorensis stem bark extracts on woods. Maderas-Cienc. Tecnol. 2018, 20, 315–324. [Google Scholar] [CrossRef] [Green Version]
- Babarinde, S.A.; Ewete, F.K. Comparative bioactivity of three Khaya species (Meliaceae) against Callosobruchus maculatus Fabricius (Coleoptera: Bruchidae). J. Entomol. Res. Soc. 2008, 10, 27–35. [Google Scholar]
- Bamaiyi, L.J.; Ndams, I.S.; Toro, W.A.; Odekina, S. Effect of Mahogany Khaya senegalensis seed oil in the control of Callosobruchus maculatus on stored cowpea. Plant Protec. Sci. 2006, 42, 130–134. [Google Scholar] [CrossRef] [Green Version]
- Shaalan, E.A.; Canyon, D.V.; Younes, M.W.F.; Hoda, A.W.; Abdel-Hamid, M. Efficacy of eight larvicidal botanical extracts from Khaya senegalensis and Daucus carota against Culex annulirostris. J. Am. Mosquito Cont. 2006, 22, 433–436. [Google Scholar] [CrossRef]
- Kuevi, D.N.O.; Kuntworbe, N.; Ayertey, E. Effects of pH and electrolytes on castor oil emulsions with various stabilisers using Khaya senegalensis gum as an emulsifier. Adv. Pharm. Pharm. Sci. 2021, 2021, 7049332. [Google Scholar] [CrossRef]
- Verma, R.K.; Yu, W.; Singh, S.P.; Shankar, S.; Srivastava, R.K. Anthothecol-encapsulated PLGA nanoparticles inhibit pancreatic cancer stem cell growth by modulating sonic hedgehog pathway. Nanomed. Nanotechnol. Biol. Med. 2015, 11, 2061–2070. [Google Scholar] [CrossRef]
- Halsall, T.G.; Troke, J.A. The structures of three new meliacins isolated from Khaya anthotheca heartwood. J. Chem. Soc. Perkin Trans. 1 1975, 6, 1758–1764. [Google Scholar]
- Adesida, G.A.; Adesogan, E.K.; Okorie, D.A. The limonoid chemistry of the genus Khaya (Meliaceae). Phytochemistry 1971, 10, 1845–1853. [Google Scholar] [CrossRef]
- Adesogan, E.K.; Okorie, D.A.; Taylor, D.A.H. Limonoids from Khaya anthotheca (Welw.) C.DC. J. Chem. Soc. 1970, 205, 205–211. [Google Scholar] [CrossRef]
- Ekong, D.E.U.; Joseph, I.; Sondengam, B.L. The meliacins (limonoids): Minor constituents of Khaya anthotheca: Reduction of the meliacins with zinc–copper couple. J. Chem. Soc. Perkin Trans. 1 1975, 21, 2118–2122. [Google Scholar] [CrossRef]
- Bevan, C.W.L.; Rees, A.H.; Taylor, D.A.H. West African timbers. part VII. anthothecol, an extractive from Khaya anthotheca. J. Chem. Soc. 1963, 983–989. [Google Scholar] [CrossRef]
- Bickii, J.; Njifutie, N.; Foyere, J.A.; Basco, L.K.; Ringwald, P. In vitro antimalarial activity of limonoids from Khaya grandifoliola C.D.C. (Meliaceae). J. Ethnopharmacol. 2000, 69, 27–33. [Google Scholar] [CrossRef]
- Adesogan, E.K.; Taylor, D.A.H. Limonoid extractives from Khaya ivorensis. J. Chem. Soc. 1970, 12, 1710–1714. [Google Scholar] [CrossRef]
- Luco, J.M.; Sosa, M.C.; Cesco, J.C. Molecular connectivity and hydrophobicity in the study of antifeedant activity of clerodane diterpenoids. Pestic. Sci. 1994, 41, 1–6. [Google Scholar] [CrossRef]
- Koul, O.; Isman, M.B. Toxicity of the limonoid allelochemical cedrelone to noctuid larvae. Entomol. Exp. Appl. 2011, 64, 281–287. [Google Scholar] [CrossRef]
- Madhusudana, R.M.; Meshulam, H.; Zelnik, R. Structure and stereochemistry of limonoids of Cabralea eichleriana. Phytochemistry 1975, 14, 1071–1075. [Google Scholar] [CrossRef]
- Govindachari, T.R.; Narsimhan, N.S.; Suresh, G.; Partho, P.D.; Gopalakrishnan, G.; Kumari, G.N.K. Structure related insect antifeedant and growth regulating activities of some limonoids. J. Chem. Ecol. 1995, 21, 1585–1600. [Google Scholar] [CrossRef]
- Braga, P.; Soares, M.S.M.; Fátima, G.F.; Vieira, P.C.; Fernandes, J.B.; Pinheiro, A.L. Dammarane triterpenes from Cabralea canjerana (Vell.) Mart. (Meliaceae): Their chemosystematic significance. Biochem. Syst. Ecol. 2006, 34, 282–290. [Google Scholar] [CrossRef]
- Adesogan, E.K.; Taylor, D.A.H. Extractives from Khaya senegalensis (Desr.) A. Juss. Chem. Comm. 1968, 16, 790–791. [Google Scholar] [CrossRef]
- Magrini, F.E.; Specht, A.; Gaio, J.; Girelli, C.P.; Migues, I.; Heinzen, C.; Saldana, J.; Sartori, V.C.; Cesio, V. Antifeedant activity and effects of fruits and seeds extracts of Cabralea canjerana canjerana (Vell.) Mart. (Meliaceae) on the immature stages of the fall armyworm Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae). Ind. Crops Prod. 2015, 65, 150–158. [Google Scholar] [CrossRef]
- Abdel-Ghany, H.S.M.; Fahmy, M.M.; Abuowarda, M.M.; Abdel-Shafy, S.; Hoballah, E.M. In vitro acaricidal effect of Melia azedarach and Artemisia herba-alba extracts on Hyalomma dromedarii (Acari: Ixodidae): Embryonated eggs and engorged nymphs. J. Parasit. Dis. 2019, 43, 696–710. [Google Scholar] [CrossRef] [PubMed]
- Bhakyaraj, K.; Kumaraguru, S.; Gopinath, K.; Sabitha, V.; Arumugam, A. Eco-friendly synthesis of palladium nanoparticles using Melia azedarach leaf extract and their evaluation for antimicrobial and larvicidal activities. J. Clust. Sci. 2017, 28, 463–476. [Google Scholar] [CrossRef]
- Ramanibai, R.; Velayutham, K. Bioactive compound synthesis of Ag nanoparticles from leaves of Melia azedarach and its control for mosquito larvae. Res. Vet. Sci. 2015, 98, 82–88. [Google Scholar] [CrossRef]
- Ntalli, N.; Kopiczko, A.; Radtke, K.; Marciniak, P.; Rosinski, G.; Adamski, Z. Biological activity of Melia azedarach extracts against Spodoptera exigua. Biologia 2014, 69, 1606–1614. [Google Scholar] [CrossRef]
- Ashrafju, M.; Ahmadi, K.; Hamidi, H. Different concentrations of Melia azedarach L. (Meliaceae) ethanolic extract on the developmental time and egg laying of Tetranychus urticae Koch. Arch. Phytopathol. Plant Prot. 2014, 47, 2242–2247. [Google Scholar] [CrossRef]
- Maurya, R.P.; Kushwaha, M.L.; Yadav, L.B. Insecticidal activity of ethanolic extracts from seed and leaf of bakain (Melia azedarach) and its effect on oviposition and egg hatching of Pieris brassicae in mid hills of Uttarakhand. Indian J. Agric. Sci. 2013, 83, 866–871. [Google Scholar]
- Jung, P.H.; Silveira, A.C.D.; Nieri, E.M.; Potrich, M.; Refatti, M. Insecticidal activity of Eugenia uniflora L. and Melia azedarach L. on Atta laevigata Smith. Floresta Ambiente 2013, 20, 191–196. [Google Scholar] [CrossRef] [Green Version]
- Mckenna, M.M.; Hammad, A.F.; Farran, M.T. Effect of Melia azedarach (Sapindales: Meliaceae) fruit extracts on citrus leafminer Phyllocnistis citrella (Lepidoptera: Gracillariidae). Springerplus 2013, 2, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inacio, M.D.F.; Carvalho, M.G.D. Insecticidal activity of dichloromethane and methanolic extracts of Azadirachta indica (A. Juss), Melia azedarach (L.) and Carapa guianenses (Aubl.) (Meliaceae) on the subterranean termite Coptotermes gestroi (Wasmann) (Isoptera, Rhinotermitidae). Biosci. J. 2012, 28, 676–683. [Google Scholar]
- Bullangpoti, V.; Wajnberg, E.; Audant, P.; Feyereisen, R. Antifeedant activity of Jatropha gossypifolia and Melia azedarach senescent leaf extracts on Spodoptera frugiperda (Lepidoptera: Noctuidae) and their potential use as synergists. Pest Manag. Sci. 2012, 68, 1255–1264. [Google Scholar] [CrossRef]
- Orhan, I.E.; Guner, E.; Ozcelik, B.; Senol, F.S.; Caglar, S.S.; Emecen, G.; Kocak, O.; Sener, B. Assessment of antimicrobial, insecticidal and genotoxic effects of Melia azedarach L. (Chinaberry) naturalized in Anatolia. Int. J. Food Sci. Nutr. 2012, 63, 560–565. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.K.; Sharma, I.D.; Sharma, K.C. Effect of Melia extracts on oviposition, gonad development and field infestation of melon fruit fly (Bactrocera tau) in cucumber (Cucumis sativus). Indian J. Agric. Sci. 2012, 82, 523–527. [Google Scholar]
- Roy, S.; Mukhopadhyay, A. Bioefficacy assessment of Melia azedarach (L.) seed extract on tea red spider mite, Oligonychus coffeae (Nietner) (Acari: Tetranychidae). Int. J. Acarol. 2012, 38, 79–86. [Google Scholar] [CrossRef]
- Chiffelle, I.; Huerta, A.; Azua, F.; Puga, K.; Araya, J.E. Antifeeding and insecticide properties of aqueous and ethanolic fruit extracts from Melia azedarach L. on the elm leaf beetle Xanthogaleruca luteola Müller. Chil. J. Agric. Res. 2011, 71, 218–225. [Google Scholar] [CrossRef]
- Maroneze, D.M.; Gallegos, D.M.N. Effect of Melia azedarach aqueous extract on the development of immature and reproductive stages of Spodoptera frugiperda (J. E. Smith, 1797) (Lepidoptera: Noctuidae). Semin. Cienc. Agrar. 2009, 30, 537–550. [Google Scholar]
- Defagó, M.T.; Dumón, A.; Avalos, D.S.; Palacios, S.M.; Valladares, G. Effects of Melia azedarach extract on Cotesia ayerza, parasitoid of the alfalfa defoliator Colias lesbia. Biol. Control 2011, 57, 75–78. [Google Scholar] [CrossRef]
- Brunherotto, R.; Vendramim, J.D.; Oriani, M.A.D.G. Effects of tomato genotypes and aqueous extracts of Melia azedarach leaves and Azadirachta indica seeds on Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Neotrop. Entomol. 2010, 39, 784–791. [Google Scholar] [CrossRef] [Green Version]
- Cabral, M.M.O.; Crescente, E.R.F.; Mendonça, P.M.; Gomes, C.M.S.; Oliveira, V.C.; Kelecom, A. Extratos de Melia azedarach L. extracts and their activity on Musca domestica L. (Diptera: Muscidae). Rev. Bras. Farmacogn. 2008, 18, 699–702. [Google Scholar] [CrossRef] [Green Version]
- Trudel, R.E.; Bomblies, A. Larvicidal effects of Chinaberry (Melia azederach) powder on Anopheles arabiensis in Ethiopia. Parasite Vector. 2011, 4, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prophiro, J.S.; Rossi, J.C.N.; Pedroso, M.F.; Kanis, L.A.; Silva, O.S. Leaf extracts of Melia azedarach Linnaeus (Sapindales: Meliaceae) act as larvicide against Aedes aegypti (Linnaeus, 1762) (Diptera: Culicidae). Rev. Soc. Bras. Med. Trop. 2008, 41, 560–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.I.; Kim, S.G.; Kang, B.R.; Ko, S.J.; Kim, J.S.; Kim, S.S. Management of two spotted spider mite, Tetranychus urticae, on organic strawberry field in jeonnam area and toxicity of natural enemies against crude extract of Chrysanthimum cinerariefolium and Melia azedarach. Korean J. Org. Agric. 2009, 17, 211–226. [Google Scholar]
- Farag, M.; Ahmed, M.H.M.; Yousef, H.; Abdel-Rahman, A.A.H. Repellent and insecticidal activities of Melia azedarach L. against cotton leafworm, Spodoptera littoralis (Boisd.). Z. Naturforsch. C 2011, 66, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Nathala, E.; Dhingra, S.; Walia, S.; Shankar, G.K. Antifeedant and toxic activity of Melia azedarach seed extracts against Helicoverpa armigera (Lepipoptera. Noctuiuae). Pestic. Res. J. 2008, 20, 16–20. [Google Scholar]
- Rossetti, M.R.; Defagó, M.T.; Carpinella, M.C.; Palacios, S.M.; Valladares, G. Biological activity of extracts of Melia azedarach on larvae of Spodoptera eridania (Lepidoptera: Noctuidae). Rev. Soc. Entomol. Argent. 2008, 67, 115–125. [Google Scholar]
- Ibrahem, M.; Seedo, M.; Abdollah, J. Biological effectiveness of Melia azedarach L., fruits extract on potato tuber moth, Phthorimaea operculella (Zeller), (Lepidoptera: Gelechiidae) under lab conditions. Dirasat Agric. Sci. 2010, 35, 11–12. [Google Scholar]
- Sharma, A.; Sharma, K.K. Effect of Melia azedarach extract on the feeding of Diorhabda lusca (Maulik) on Celtis australis Linn. J. Entomol. Res. 2008, 32, 117–118. [Google Scholar]
- Sharma, A.; Sehgal, R.N.K.C.; Kumar, R.; Gupta, R. Field efficacy of various extracts of Melia azedarach (Linn) seeds against Ascotis imparta Hubn. (Lepidoptera: Geometridae) larvae. J. Entomol. Res. 2007, 31, 47–49. [Google Scholar]
- Rachokarn, S.; Piyasaengthong, N.; Bullangpoti, V. Impact of botanical extracts derived from leaf extracts Melia azedarach L. (Meliaceae) and Amaranthus viridis L. (Amaranthaceae) on populations of Spodoptera exigua (Hübner) (Lepidoptera: Noctuidae) and detoxification enzyme activities. Commun. Agric. Appl. Biol. Sci. 2008, 73, 451–457. [Google Scholar]
- Defagó, M.; Valladares, G.; Banchio, E.; Carpinella, C.; Palacios, S. Insecticide and antifeedant activity of different plant parts of Melia azedarach on Xanthogaleruca luteola. Fitoterapia 2006, 77, 500–505. [Google Scholar] [CrossRef] [PubMed]
- Rossi, J.C.N.; Prophiro, J.S.; Mendes, A.M.; Kanis, L.A.; Silva, O.S. Larvicidal effect of ethanolic extracts from Melia azedarach dried leaves and ripe fruits in Aedes albopictus larvae. Lat. Am. J. Pharm. 2007, 26, 737–740. [Google Scholar]
- Coria, C.; Almiron, W.; Valladares, G.; Carpinella, C.; Luduena, F.; Defago, M.; Palacios, S. Larvicide and oviposition deterrent effects of fruit and leaf extracts from Melia azedarach L. on Aedes aegypti (L.) (Diptera: Culicidae). Bioresour. Technol. 2008, 99, 3066–3070. [Google Scholar] [CrossRef] [PubMed]
- El-Sawi, S.A. Toxicity and bioactivity of Melia azedarach L. on the two spotted spider mite Tetranychus urticae Koch (Acari: Tetranychidae), and its predacious mite, Euseius scutalis (Athais-Henriot) (Acari: Phytoseiidae). Egypt. J. Biol. Pest Control 2008, 18, 289–295. [Google Scholar]
- Mitchell, P.L.; Thielen, J.B.; Stell, F.M.; Fescemyer, H.W. Activity of Melia volkensii (Meliaceae) extract against southern green stink bug (Hemiptera: Heteroptera: Pentatomidae). J. Agric. Urban. Entomol. 2004, 21, 131–141. [Google Scholar]
- Nathan, S.S.; Sehoon, K. Effects of Melia azedarach L. extract on the teak defoliator Hyblaea puera Cramer (Lepidoptera: Hyblaeidae). Crop Prot. 2006, 25, 287–291. [Google Scholar] [CrossRef]
- Charleston, D.S.; Kfir, R.; Dicke, M.; Vet, L.E.M. Impact of botanical extracts derived from Melia azedarach and Azadirachta indica on populations of Plutella xylostella and its natural enemies: A field test of laboratory findings. Biol. Control 2006, 39, 105–114. [Google Scholar] [CrossRef]
- Charleston, D.S.; Gols, R.; Hordijk, K.A.; Kfir, R.; Vet, L.E.M.; Dicke, M. Impact of botanical pesticides derived from Melia azedarach and Azadirachta indica plants on the emission of volatiles that attract parasitoids of the diamondback moth to cabbage plants. J. Chem. Ecol. 2006, 32, 325–349. [Google Scholar] [CrossRef]
- Bounechada, M.; Doumandji, S.E.; Laouer, H. Laboratory evaluation of Melia azedarach L. and Eucalyptus globulus Labill. extracts in order to control Ocneridia volxemi Bolivar (Orthoptera, Pamphaginae) hoppers. Commun. Agric. Appl. Biol. Sci. 2004, 69, 235–244. [Google Scholar]
- Wandscheer, C.B.; Duque, J.E.; Silva, M.A.N.D.; Fukuyama, Y.; Wohlke, J.L.; Adelmann, J.; Fontana, J.D. Larvicidal action of ethanolic extracts from fruit endocarps of Melia azedarach and Azadirachta indica against the dengue mosquito Aedes aegypti. Toxicon 2004, 44, 829–835. [Google Scholar] [CrossRef]
- Gursharan, S.; Vishaldeep, K.; Darshan, S. Lethal and sublethal effects of different ecotypes of Melia azedarach against Plutella xylostella (Lepidoptera: Plutellidae). Int. J. Trop. Insect Sci. 2006, 2, 92–100. [Google Scholar]
- Nathala, E.; Dhingra, S. Relative effect of various extracts of Melia azedarach (Linn) leaf against Helicoverpa armigera (Hübner) and their comparison with seed extracts. J. Entomol. Res. 2005, 29, 201–206. [Google Scholar]
- Haryadi, Y.; Yuniarti, S. Study on the insecticidal effects of custard apple (Annona reticulata L.) and mindi (Melia azedarach L.) leaves against Sitophilus zeamais Motschulsky (Coleoptera: Curculionidae). In Proceedings of the 8th International Working Conference on Stored Product Protection, York, UK, 22–26 July 2002; pp. 600–602. [Google Scholar]
- Jazzar, C.; Hammad, A.F. The efficacy of enhanced aqueous extracts of Melia azedarach leaves and fruits integrated with the Camptotylus reuteri releases against the sweetpotato whitefly nymphs. Bull. Insectology 2003, 56, 169–275. [Google Scholar]
- Juan, A.; Sans, A.; Riba, M. Antifeedant activity of fruit and seed extracts of Melia azedarach and Azadirachta indica on larvae of Sesamia nonagrioides. Phytoparasitica 2000, 28, 311–319. [Google Scholar] [CrossRef]
- Hoste, B.; De, L.A. Effect of an insecticidal Melia azedarach extract on food transport in the gut of insects. In Proceedings of the 52nd International Symposium on Crop Protection, PTS I and II, Ghent, Belgium, 9 May 2000; Volume 65, pp. 423–430. [Google Scholar]
- Breuer, M.; Schmidt, G.H. Effect of Melia azedarach extract incorporated into an artificial diet on growth, development and fecundity of Spodoptera frugiperda (J. E. Smith) (Lep., Noctuidae). Acta Neurochir. 1996, 103, 171–194. [Google Scholar]
- Schmidt, G.H.; Ahmed, A.A.I.; Breuer, M. Effect of Melia azedarach extract on larval development and reproduction parameters of Spodoptera littoralis (Boisd.) and Agrotis ipsilon (Hufn.) (Lep. Noctuidae). Anzger Schdlingskunde Pflanzenschutz Umweltschutz 1997, 70, 4–12. [Google Scholar] [CrossRef]
- Koul, O.; Jain, M.P.; Sharma, V.K. Growth inhibitory and antifeedant activity of extracts from Melia dubia to Spodoptera litura and Helicoverpa armigera larvae. Indian J. Exp. Biol. 2000, 38, 63–68. [Google Scholar] [PubMed]
- Breuer, M.; Schmidt, G.H. Influence of a short period treatment with Melia aedarach extract on food intake and growth of the larvae of Spodoptera frugiperda (J.E.Smith) (Lep., Noctuidae). Z. Pflanzenk. Pflanzen. 1995, 102, 633–654. [Google Scholar]
- Tillman, P.G.; Cottrell, T.E.; Buntin, G.D. Potential of Melia azedarach L. (Meliaceae) as a host plant for Halyomorpha halys (Stl) (Hemiptera: Pentatomidae). Fla. Entomol. 2019, 102, 222–226. [Google Scholar]
- Nardo, E.A.B.D.; Costa, A.S.D.; Loureno, A.L. Melia azedarach extract as an antifeedant to Bemisia tabaci (Homoptera: Aleyrodidae). Fla. Entomol. 1997, 80, 92–94. [Google Scholar] [CrossRef]
- Irungu, L.W.; Mwangi, R.W. Effects of a biologically active fraction from Melia volkensii on Culex quinquefasciatus. Int. J. Trop. Insect Sci. 1995, 16, 159–162. [Google Scholar] [CrossRef]
- Dilawari, V.K.; Singh, K.; Dhaliwal, G.S. Effects of Melia azedarach L. on oviposition and feeding of Plutella xylostella L. Int. J. Trop. Insect Sci. 1994, 15, 203–205. [Google Scholar] [CrossRef]
- Yousef, H.; El-Lakwah, S.F. Effects of Melia azedarach ripe fruit extract on some enzyme activities of the cotton leafworm Spodoptera littoralis (Boisd.). Egypt. J. Pest Control 2014, 24, 315–320. [Google Scholar]
- Scapinello, J.; Oliveira, J.V.; Ribeiros, M.L. Effects of supercritical CO2 extracts of Melia azedarach L. on the control of fall armyworm (Spodoptera frugiperda). J. Supercrit. Fluids 2014, 93, 20–26. [Google Scholar] [CrossRef]
- Mwangi, R.W.; Mukiama, T.K. Evaluation of Melia volkensii extract fractions as mosquito larvicides. J. Am. Mosquito Control 1988, 4, 442–447. [Google Scholar]
- Raja, N. Field evaluation of aqueous extract of Melia azedarach Linn. seeds against cabbage aphid, Brevicoryne brassicae Linn. (Homoptera: Aphididae), and its predator Coccinella septempunctata Linn. (Coleoptera: Coccinellidae). Arch. Phytopathol. Plant Prot. 2012, 45, 1273–1279. [Google Scholar]
- Souza, B.H.S.; Costa, E.N.; Forim, M.R. Formulations of Melia azedarach to control Diabrotica speciosa (Germar) (Coleoptera: Chrysomelidae) larvae in corn and plant enhancement. Neotrop. Entomol. 2015, 44, 173–179. [Google Scholar] [CrossRef]
- Kebede, Y.; Gebre-Michael, T.; Balkew, M. Laboratory and field evaluation of neem (Azadirachta indica A. Juss) and Chinaberry (Melia azedarach L.) oils as repellents against Phlebotomus orientalis and P. bergeroti (Diptera: Psychodidae) in Ethiopia. Acta Trop. 2010, 113, 145–150. [Google Scholar] [CrossRef]
- Al-Mehmadi, R.M.; Al-Khalaf, A.A. Larvicidal and histological effects of Melia azedarach extract on Culex quinquefasciatus Say larvae (Diptera: Culicidae). J. King Saud Univ. Sci. 2010, 22, 77–85. [Google Scholar] [CrossRef] [Green Version]
- Kamaraj, C.; Rahuman, A.A.; Bagavan, A.; Mohamed, M.J.; Elango, G.; Rajakumar, G.; Zahir, A.A.; Santhoshkumar, T.; Marimuthu, S. Ovicidal and larvicidal activity of crude extracts of Melia azedarach against Haemonchus contortus (Strongylida). Parasitol. Res. 2010, 106, 1071–1077. [Google Scholar] [CrossRef]
- Shrestha, S.S.; Ferrarese, I.; Sut, S.; Zengin, G.; Grana, S.; Ak, G.; Pant, D.R.; Dall’Acqua, S.; Rajbhandary, S. Phytochemical investigations and in vitro bioactivity screening on Melia azedarach L. leaves extract from Nepal. Chem. Biodivers. 2021, 18, e2001070. [Google Scholar] [CrossRef] [PubMed]
- Abiy, E.; Gebre-Michael, T.; Balkew, M.; Medhin, G. Repellent efficacy of DEET, MyggA, neem (Azedirachta indica) oil and chinaberry (Melia azedarach) oil against Anopheles arabiensis, the principal malaria vector in Ethiopia. Malaria J. 2015, 14, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.B.; Minami, Y.; Yagi, F.; Tadera, K.; Nakatani, M. Ring C-seco limonoids from Melia toosendan. Phytochemistry 1997, 46, 911–914. [Google Scholar] [CrossRef]
- Munehiro, N. Limonoids from Melia toosendan (Meliaceae) and their antifeedant activity. Heterocycles 1999, 118, 313–318. [Google Scholar]
- Huang, R.C.; Okamura, H.; Iwagawa, T.; Nakatani, M. The structures of azedarachins, limonoid antifeedants from Chinese Melia azedarach Linn. B Chem. Soc. Jpn. 1994, 67, 2468–2472. [Google Scholar] [CrossRef]
- Nakatani, M.; Huang, R.C.; Okamura, H.; Naokia, H.; Iwagawa, T. Limonoid antifeedants from Chinese Melia azedarach. Phytochemistry 1994, 36, 39–41. [Google Scholar] [CrossRef]
- Oelrichs, P.B.; Hill, M.W.; Vallely, P.J.; Macleod, J.K.; Molinski, T.F. Toxic tetranortriterpenes of the fruit of Melia azedarach. Phytochemistry 1983, 22, 531–534. [Google Scholar] [CrossRef]
- Takeya, K.; Qiao, Z.S.; Hirobe, C.; Itokawa, H. Cytotoxic azadirachtin-type limonoids from Melia azedarach. Phytochemistry 1996, 42, 709–712. [Google Scholar] [CrossRef]
- Valladares, G.R.; Ferreyra, D.; Defago, M.T.; Carpinella, M.C.; Palacios, S. Effects of Melia azedarach on Triatoma infestans. Fitoterapia 1999, 70, 421–424. [Google Scholar] [CrossRef]
- Valladares, G.; Defago, M.T.; Palacios, S. Laboratory evaluation of Melia azedarach (Meliaceae) extracts against the elm leaf beetle (Coleoptera: Chrysomelidae). J. Econ. Entomol. 1997, 90, 747–750. [Google Scholar] [CrossRef]
- Itokawa, H.; Qiao, Z.S.; Hirobe, C.; Takeya, K. Cytotoxic limonoids and tetranortriterpenoids from Melia azedarach. Chem. Pharm. Bull. 1995, 43, 1171–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Céspedes, C.L.; Calderón, J.S.; Lina, L.; Aranda, E. Growth inhibitory effects on fall armyworm Spodoptera frugiperda of some limonoids isolated from Cedrela spp. (Meliaceae). J. Agric. Food Chem. 2000, 48, 1903–1908. [Google Scholar] [CrossRef] [PubMed]
- Isman, M.B.; Koul, O.; Luczynski, A.; Kaminski, J. Insecticidal and antifeedant bioactivities of neem oils and their relationship to azadirachtin content. J. Agric. Food Chem. 1990, 38, 1406–1411. [Google Scholar] [CrossRef]
- Carpinella, M.C.; Defagó, M.T.; Valladares, G.; Palacios, S.M. Chapter 5: Role of Melia azedarach L. (Meliaceae) for the control of insects and acari: Present status and future prospects. Adv. Phytomed. 2006, 3, 81–123. [Google Scholar]
- Wiart, C. A note on Melia toosendan Siebold & Zucc. Toxicology 2012, 295, 68. [Google Scholar]
- Araújo, S.A.C.D.; Teixeira, M.; Dantas, T.; Melo, V.S.P.; Lima, F.E.S.; Ricarte, A.R.F.; Costa, E.C.; Miranda, A.M. Usos potenciais de Melia azedarach L. (Meliaceae): Um levantamento. Arq. Inst. Biol. 2009, 76, 141–148. [Google Scholar] [CrossRef]
- Carpinella, C.; Ferrayoli, C.; Valladares, G.; Defago, M.; Palacios, S. Potent limonoid insect antifeedant from Melia azedarach (organic chemistry). Biosci. Biotech. Bioch. 2002, 66, 1731–1736. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.B.; Okamura, H.; Iwagawa, T.; Nakatani, M. Limonoid antifeedants from Melia toosendan. Phytochemistry 1996, 41, 117–120. [Google Scholar] [CrossRef]
- Huang, R.C.; Okamura, H.; Iwagawa, T.; Tadera, K.; Nakatani, M. Azedarachin C, a limonoid antifeedant from Melia azedarach. Phytochemistry 1995, 38, 593–594. [Google Scholar] [CrossRef]
- Sombatsiri, K.; Ermel, K.; Schmutterer, H.; Ascher, K.R.S.; Zebitz, C.P.W.; Naqvi, S.N.H. Other meliaceous plants containing ingredients for integrated pest management and further purposes: Sections 8.2.1.3–8.2.1.10. In The Neem Tree; Wiley-VCH Verlag GmbH Co. KGaA: Weinheim, Germany, 2005; pp. 606–612. [Google Scholar] [CrossRef]
- Nakatani, M.; Zhou, J.B.; Iwagawa, T.; Okamura, H. Isolation of two limonoid antifeedants from Melia toosendan. Heterocycles 1996, 3, 739–743. [Google Scholar] [CrossRef]
- Nakatani, M.; Fukuman, Y.; Sakumoto, T.; Yamashita, N.; Okamura, H.; Iwagawa, T. Nimbolinins, C-Seco limonoids from the fruits of Melia toosendan. Hetercocycles 2000, 53, 689–695. [Google Scholar] [CrossRef]
- Huang, R.C.; Tadera, K.; Yagi, F.; Minami, Y.; Okamura, H.; Iwagawa, T.; Nakatani, M. Limonoids from Melia azedarach. Phytochemistry 1996, 43, 581–583. [Google Scholar] [CrossRef]
- Nakatani, M.; Zhou, J.B.; Nakayama, N.; Okamura, H.; Iwagawa, T. Nimbolidins C-E, limonoid antifeedants from Melia toosendan. Phytochemistry 1996, 41, 739–743. [Google Scholar] [CrossRef]
- Kraus, W.; Bokel, M.; Klenk, A.; Pöhnl, H. The structure of azadirachtin and 22,23-dihydro-23β-methoxyazadirachtin. Tetrahedron Lett. 1985, 26, 6435–6438. [Google Scholar] [CrossRef]
- Kraus, W.; Bokel, M.; Bruhn, A.; Cramer, R.; Klaiber, I.; Klenk, A.; Nagl, G.; Pöhnl, H.; Sadlo, H.; Vogler, B. Structure determination by NMR of azadirachtin and related compounds from Azadirachta indica A. Juss. Tetrahedron 1987, 43, 2817–2830. [Google Scholar] [CrossRef]
- Zhou, Y.W.; Li, H.; Feng, J.T. Preliminary study on the relationship between bioactivity and formulation of toosendanin. J. Hebei Agric. Univ. 2018, 41, 6–11. [Google Scholar]
- Zhang, J.; Feng, G.; Luo, L.; Yu, X.Y.; Ma, Z.Q.; Feng, J.T.; Liu, X.J.; Zhang, X. Development of an enzyme-linked immunosorbent assay for toosendanin. Anal. Chim. Acta. 2008, 622, 182–188. [Google Scholar] [CrossRef]
- Busato, M.A.; Lutinski, J.A.; Hermes, D.; Magro, J.D.; Bordignon, C.M.; Bohn, A.; Simoes, D.A. Larvicidal effects of supercritical extract of Melia azedarach and Bacillus thuringiensis var. israelensis product on late instar of Aedes aegypti (Diptera: Culicidae). Rev. Colomb. Entomol. 2018, 44, 39–43. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.L.; Wang, Y.I.; Shinfoon, C. The toxic chemical factors in the fruits ofmelia azedarach and their bio-activitiestoward Pieris rapae. Acta Entomol. Sinica. 1994, 37, 20–24. [Google Scholar]
- Chiu, S.F.; Huang, D.P.; Zhang, X. The antifeedant and growth disruption effects of extracts from some meliaceous plants to the Asiatic corn borer. Acta Entomol. Sin. 1985, 28, 450–453. [Google Scholar]
- Chiu, S.F.; Zhang, X. Experiments on the antifeedant and systemic properties of some botanical insecticides against the rice yellow stem borer. J. Integr. Agric. 1982, 2, 55–62. [Google Scholar]
- Zhang, X.; Chiu, S.F. The antifeeding and repellent effects of meliaceus plants to some insect pests. J. South China Agric. Univ. 1983, 4, 1–7. [Google Scholar]
- Wei, T.; Wen, X.; Niu, C.; An, S.J.; Wang, D.W.; Xi, Z.; Wang, N.N. Design of acetohydroxyacid synthase herbicide-resistant germplasm through MB-QSAR and CRISPR/Cas9-mediated base-editing approaches. J. Agric. Food Chem. 2022, 70, 2817–2824. [Google Scholar] [CrossRef]
- Lin, X.D.; Li, Y.; Zhong, W.Q.; Hong, T.; Li, L.H.; Song, S.Y.; He, D.H. Synthesis, bioactivity, and QSAR Study of 3, 4-dichlorophenyl isoxazole-substituted stilbene derivatives against the phytopathogenic fungus Botrytis cinerea. J. Agric. Food Chem. 2021, 69, 9520–9528. [Google Scholar] [CrossRef]
- Duran, A.G.; Chinchilla, N.; Molinillo, J.M.G.; Macias, F.A. Structure-activity relationship studies on naphthoquinone analogs. The search for new herbicides based on natural products. Pest Manag. Sci. 2019, 75, 2517–2529. [Google Scholar] [CrossRef]
- Xu, H.; Zhang, J.L. Natural products-based insecticidal agents 9. Design, semisynthesis and insecticidal activity of 28-acyloxy derivatives of toosendanin against Mythimna separata Walker in vivo. Bioorg. Med. Chem. Lett. 2011, 21, 1974–1977. [Google Scholar] [CrossRef]
- Zhang, J.L.; Qu, H.; Yu, X.; Zhi, X.Y.; Chen, H.; Xu, H. Combinatorial synthesis of a series of alkyl/alkenylacyloxy derivatives at the C-28 position of toosendanin as insecticidal agents. Comb. Chem. High Throughput Screen. 2013, 16, 394–399. [Google Scholar] [CrossRef]
- Kawasaki, A.; Hashimoto, H.; Shimotoyodome, Y.; Ito, S.; Ishikawa, J.; Sugai, Y.; Fujimori, T. Limonoids and unsaturated fatty acids present in Melia toosendan increase ceramide production in keratinocytes. Fitoterapia 2021, 155, 105058. [Google Scholar] [CrossRef]
- Jabeen, B.; Riaz, N.; Saleem, M.; Naveed, M.A.; Ahmed, M.; Tahir, M.N.; Pescitelli, G.; Ashraf, M.; Ejaz, S.A.; Ahmed, I.; et al. Isolation and characterization of limonoids from Kigelia africana. Z. Naturforsch. B 2013, 68, 1041–1048. [Google Scholar] [CrossRef]
- Sarker, S.D.; Savchenko, T.; Whiting, P.; Sik, V.; Dinan, L. Two limonoids from Turraea obtusifolia (Meliaceae), prieurianin and rohitukin, antagonise 20-hydroxyecdysone action in a drosophila cell line. Arch. Insect Biochem. Physiol. 1997, 35, 211–217. [Google Scholar] [CrossRef]
- Schoonhoven, L.M.; Luo, L.E. Multiple mode of action of the feeding deterrent, toosendanin, on the sense of taste in Pieris brassicae larvae. J. Comp. Physiol. A 1994, 175, 519–524. [Google Scholar] [CrossRef]
- Zhang, X.; Chiu, S.F. Effects of toosendanin on several enzyme systems of the cabbage worm Pieris rapae L. Acta Entomol. Sin. 1992, 35, 171–177. [Google Scholar]
- Zhang, X.; Chiu, S.F. Study on the histopathology of the midgut of cabbage worm Pieris rapae L. caused by toosendanin. Acta Entomol. Sin. 1991, 34, 501–502. [Google Scholar]
- Zhang, X.; Chiu, S.F. Effects of toosendanin on the respiration and other physiological parameters of the imported cabbage worm (Pieris rapae L.). J. South China Agric. Univ. 1992, 13, 5–11. [Google Scholar]
- Li, H.; Zhang, J.; Ma, T.; Li, C.; Ma, Z.Q.; Zhang, X. Acting target of toosendanin locates in the midgut epithelium cells of Mythimna separate Walker larvae (Lepidoptera: Noctuidae). Ecotox. Environ. Safe 2020, 201, 110828. [Google Scholar] [CrossRef]
- Ma, Z.Q.; Monika, G.N.; Zhang, X.; Brown, M.R. Effects of the botanical insecticide, toosendanin, on blood digestion and egg production by female Aedes aegypti (Diptera: Culicidae): Topical application and ingestion. J. Med. Entomol. 2013, 50, 112–122. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.L. Toosendanin modifies k+ and Ca2+ channel activity and intracellular Ca2+ concentration. Prog. Biochem. Biophys. 2007, 34, 132–137. [Google Scholar]
- Olurishe, T.; Kwanashie, H.; Anuka, J. Effect of Khaya senegalensis on uterine contractility in rats. J. Bioresour. Bioprod. 2006, 3, 30–35. [Google Scholar] [CrossRef]
- Dong, Y.; Zhu, G.; Wang, S.F.; Keon, K.A.; Rubinstein, J.L.; Zeng, S.X.; Zhang, S.; Chen, Q.L.; Fu, J.; Li, M.; et al. Toosendanin, a novel potent vacuolar-type H+-translocating ATPase inhibitor, sensitizes cancer cells to chemotherapy by blocking protective autophagy. Int. J. Biol. Sci. 2022, 18, 2684–2702. [Google Scholar] [CrossRef]
- Miranda, R.N.C.; Dolabela, M.F.; da Silva, M.N.; Povoa, M.M.; Maia, J.G.S. Antiplasmodial activity of the andiroba (Carapa guianensis Aubl., Meliaceae) oil and its limonoid-rich fraction. J. Ethnopharmacol. 2012, 142, 679–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuyama, Y.; Nakaoka, M.; Yamamoto, T.; Takahashi, H.; Minami, H. Degraded and oxetane-bearing limonoids from the roots of Melia azedarach. Chem. Pharm. Bull. 2006, 54, 1219–1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haramboure, M.; Mirande, L.; Smagghe, G.; Pineda, S.; Schneider, M.I. Compatibility of a Melia azedarach extract with Eriopis connexa (Coleoptera: Coccinellidae). Commun. Agric. Appl. Biol. Sci. 2010, 75, 373–378. [Google Scholar] [PubMed]
- Caboni, P.; Sarais, G.; Angioni, A.; Garcia, A.J.; Lai, F.; Dedola, F.; Cabras, P. Residues and persistence of neem formulations on strawberry after field treatment. J. Agric. Food Chem. 2006, 54, 10026–10032. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; He, S.; Liu, G.; Li, C.; Ma, Z.Q.; Zhang, X. Residue and dissipation kinetics of toosendanin in cabbage, tobacco and soil using IC-ELISA detection. Food Chem. 2021, 335, 127600. [Google Scholar] [CrossRef]
- Bumah, V.V.; Essien, E.U.; Agbedahunsi, J.M.; Eka, O.U. Effects of Khaya grandifoliola on red blood cells and bone mineral content in rats. Phytother. Res. 2010, 19, 928–931. [Google Scholar] [CrossRef]
- Nwozo, S.O.; Ajaiyeoba, E.O.; Oladeinde, F.O.; Olanrewaju, I. In vivo animal model toxicological studies of Nigerian medicinal plants II: Eniantha chlorantha and Khaya ivorensis. Eur. J. Sci. Res. 2006, 14, 133–138. [Google Scholar]
- Adebayo, J.O.; Yakubu, M.T.; Egwim, E.C.; Owoyele, V.B.; Enaibe, B.U. Effect of ethanolic extract of Khaya senegalensis on some biochemical parameters of rat kidney. J. Ethnopharmacol. 2003, 88, 69–72. [Google Scholar] [CrossRef]
- Loica, Z.; Loetitia, M.; Tiburce, H.; Roland, K.; Joachim, G.; Alphonse, S. Chronophysiology of rat wistar under extracts from Khaya Senegalensis. Int. J. Sci. Res. 2018, 7, 65–71. [Google Scholar]
- Yakubu, M.T.; Adebayo, O.J.; Egwim, E.C.; Owoyele, V.B. Increased liver alkaline phosphatase and aminotransferase activities following administration of ethanolic extract of Khaya senegalensis stem bark to rats. Biokemistri 2005, 17, 27–32. [Google Scholar] [CrossRef] [Green Version]
- Keshri, G.; Bajpai, M.; Lakshmi, V.; Setty, B.S.; Gupta, G. Role of energy metabolism in the pregnancy interceptive action of Ferula assafoetida and Melia azedarach extracts in rat. Contraception 2004, 70, 429–432. [Google Scholar] [CrossRef] [PubMed]
- Orisadipe, A.; Amos, S.; Adesomoju, A.; Binda, L.; Emeje, M.; Okogun, J.; Wambebe, C.; Gamaniel, K. Spasmolytic activity of methyl angolensate: A triterpenoid isolated from Entandrophragma angolense. Biol. Pharm. Bull. 2001, 24, 364–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.T.; Chang, C.H.; Wang, Z.T. Determination of toosendanin in rat plasma by ultra-performance liquid chromatography-electrospray ionization-mass spectrometry and its application in a pharmacokinetic study. Biomed. Chromatogr. 2013, 27, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Liu, Q.; Liang, Q.X.; Gao, S.; Zhuang, K.Y.; Zhang, Y.; Zhang, H.Z.; Liu, K.C.; She, G.M.; Xia, Q. Toosendanin triggered hepatotoxicity in zebrafish via inflammation, autophagy, and apoptosis pathways. Comp. Biochem. Phys. C 2021, 250, 109171. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.L.; Shi, W.Y.; Zhong, W.; Ma, A.T.; Wang, X.D.; Zhao, Y.T.; Wang, M.; Zhong, X.H. Effects of toosendanin on pregnancy and uterine immunity alterations in mice. Am. J. Chin. Med. 2010, 38, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Hodgson, H.; De, L.P.R.; Stephenson, M.J.; Thimmappa, R.; Vincent, J.L.; Sattely, E.S.; Osbourn, A. Identification of key enzymes responsible for protolimonoid biosynthesis in plants: Opening the door to azadirachtin production. Proc. Natl. Acad. Sci. USA 2019, 116, 17096–17104. [Google Scholar] [CrossRef] [Green Version]
- Lian, X.; Zhang, X.; Wang, F.; Wang, X.N.; Xue, Z.Y.; Qi, X.Q. Characterization of a 2,3-oxidosqualene cyclase in the toosendanin biosynthetic pathway of Melia toosendan. Physiol. Plant. 2020, 170, 528–536. [Google Scholar] [CrossRef]




























| Family | Genus | Species |
|---|---|---|
| Meliaceae | Cipadessa | Cipadessa baccifera (Roth) Miq. |
| Cipadessa cinerascens (Pell.) Hand-Mazz | ||
| Entandrophragma | Entandrophragma angolense C. DC | |
| Entandrophragma bussei Harms ex Engl. | ||
| Entandrophragma caudatum Sprague | ||
| Entandrophragma candolei (Harms) | ||
| Entandrophragma delevoyi (de Wild) | ||
| Entandrophragma cylindricum (Sprague) Sprague | ||
| Entandrophragma spicatum (C.DC.) Sprague | ||
| Entandrophragma macrophyllum A. Chev. | ||
| Guarea | Guarea guidonia (L.) Sleumer | |
| Guarea grandiflora ADC | ||
| Guarea thompsonii Sprague et Hutch. | ||
| Guarea kunthiana A. Juss | ||
| Khaya | Khaya anthotheca (Welv.) C. DC | |
| Khaya senegalensis (Desr.) A. Juss. | ||
| Khaya grandifoliola C. DC | ||
| Khaya ivorensis A. Chev. | ||
| Melia | Melia azedarach L. | |
| Melia toosendan Sieb. Et Zucc. | ||
| Melia dubia Cav. | ||
| Melia volkensii Gurke |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, M.; Bi, X.; Zhou, L.; Huang, J. Insecticidal Triterpenes in Meliaceae: Plant Species, Molecules, and Activities: Part II (Cipadessa, Melia). Int. J. Mol. Sci. 2022, 23, 5329. https://doi.org/10.3390/ijms23105329
Lin M, Bi X, Zhou L, Huang J. Insecticidal Triterpenes in Meliaceae: Plant Species, Molecules, and Activities: Part II (Cipadessa, Melia). International Journal of Molecular Sciences. 2022; 23(10):5329. https://doi.org/10.3390/ijms23105329
Chicago/Turabian StyleLin, Meihong, Xiaoyang Bi, Lijuan Zhou, and Jiguang Huang. 2022. "Insecticidal Triterpenes in Meliaceae: Plant Species, Molecules, and Activities: Part II (Cipadessa, Melia)" International Journal of Molecular Sciences 23, no. 10: 5329. https://doi.org/10.3390/ijms23105329
APA StyleLin, M., Bi, X., Zhou, L., & Huang, J. (2022). Insecticidal Triterpenes in Meliaceae: Plant Species, Molecules, and Activities: Part II (Cipadessa, Melia). International Journal of Molecular Sciences, 23(10), 5329. https://doi.org/10.3390/ijms23105329