
Identification and Transcriptome Analysis of Genes Related to Membrane Lipid Regulation in Sweet Sorghum under Salt Stress

Abstract
:1. Introduction
2. Results
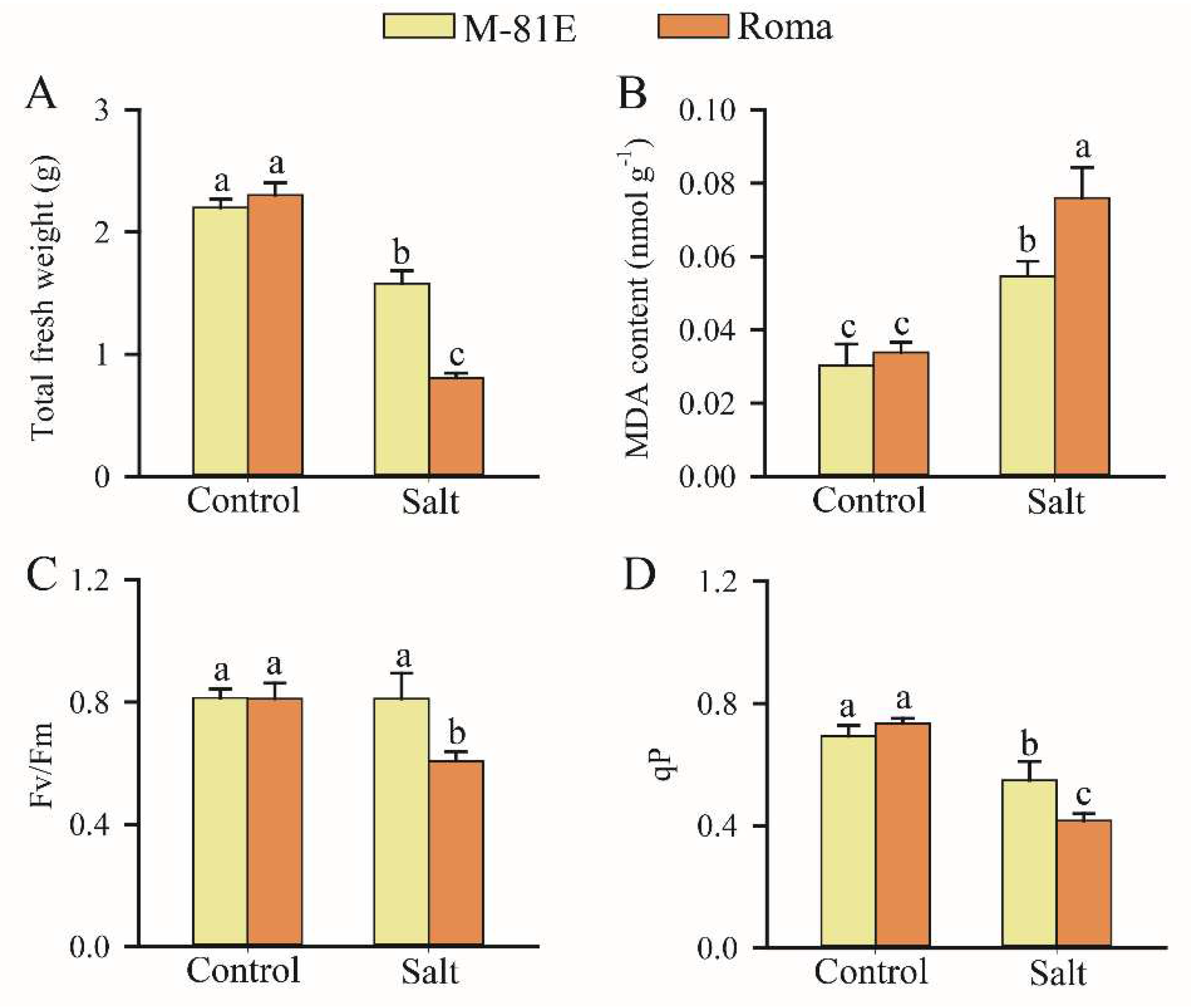
2.1. Effects of Salt Stress on Plants
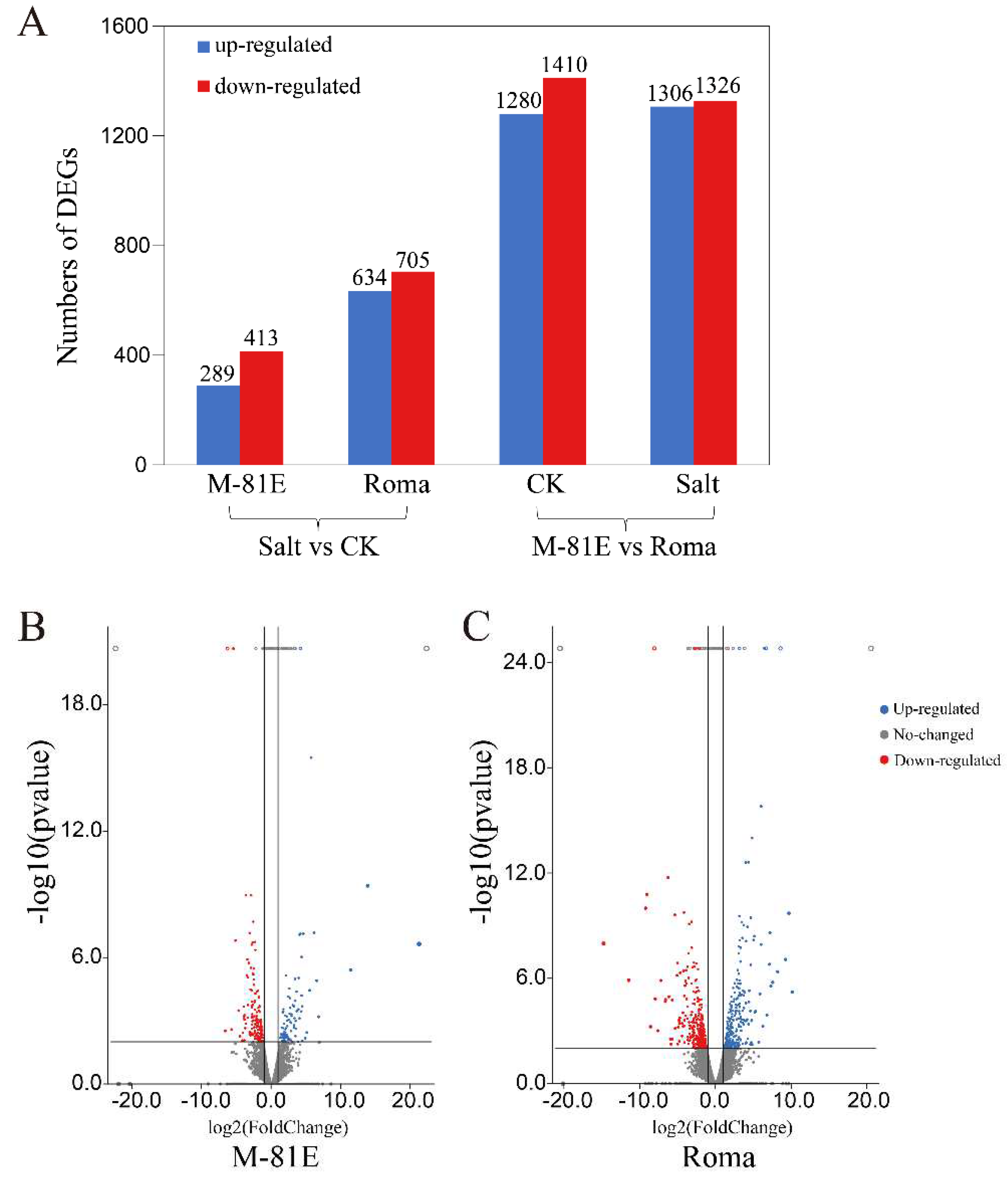
2.2. Identification of Differential Expression Genes (DEGs) Involved in Salt Stress
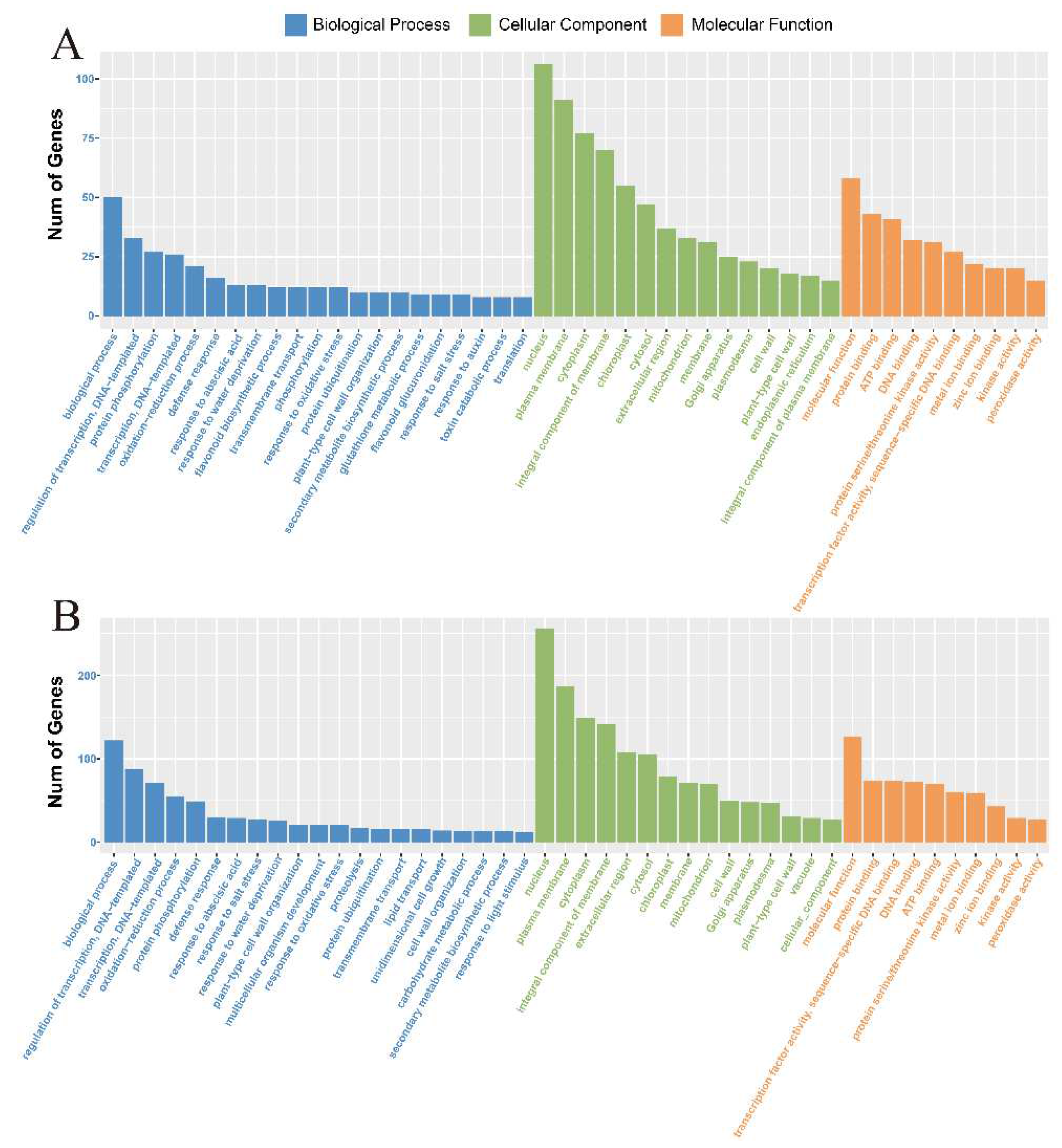
2.3. Functional Classification by Gene Ontology (GO)
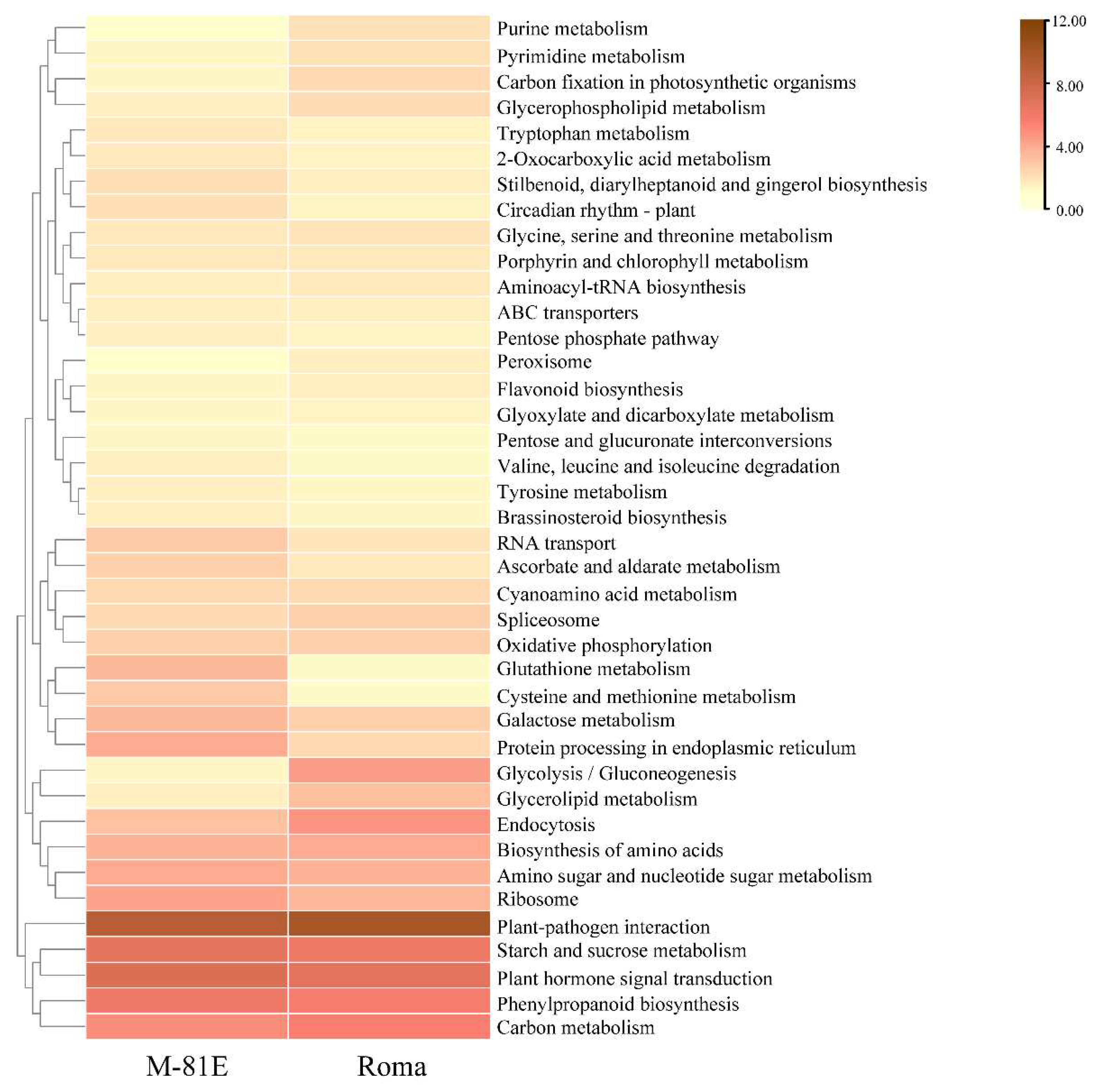
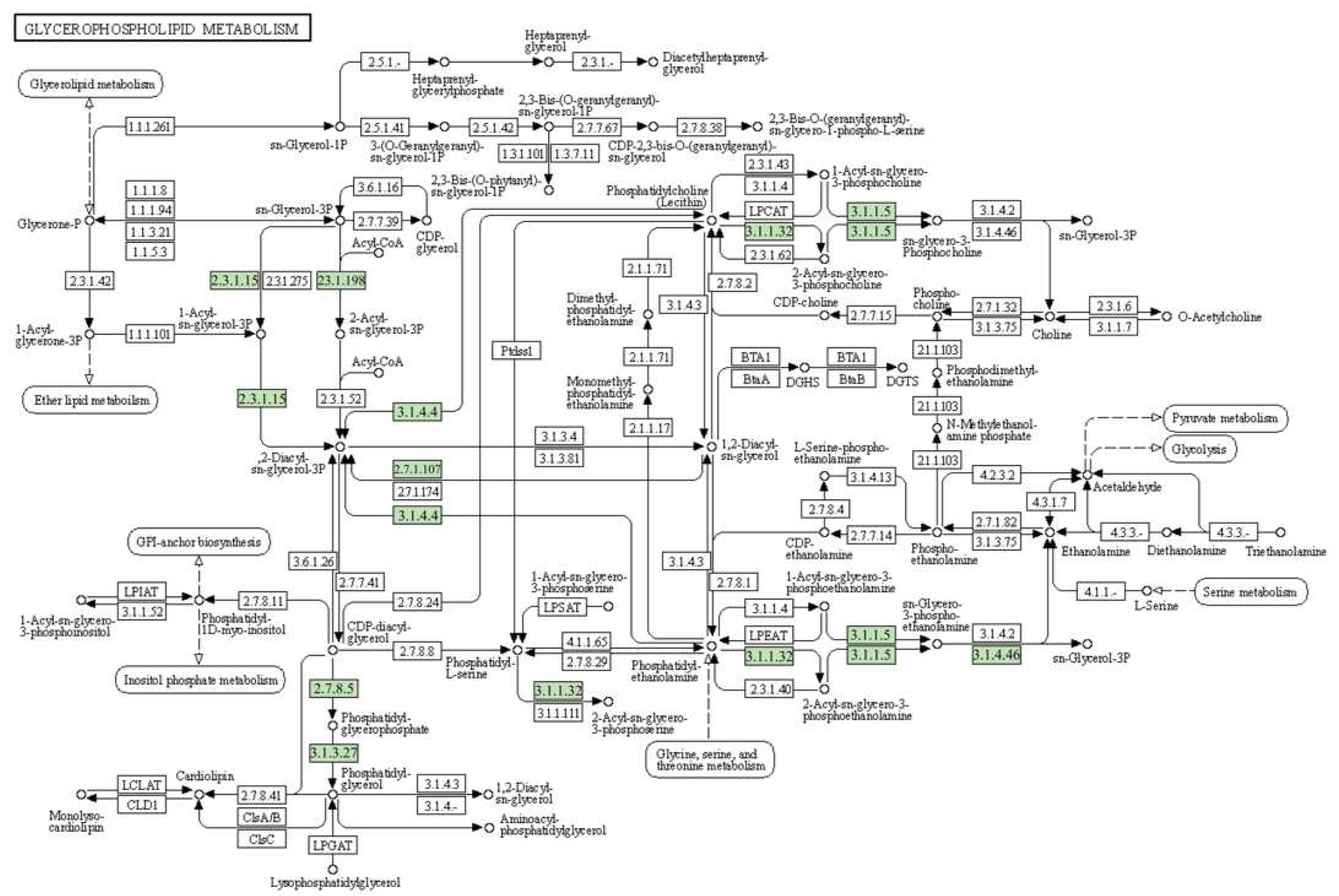
2.4. Functional Classification by KEGG
2.5. DEGs Related to Membrane Lipid Metabolism under Salt Stress
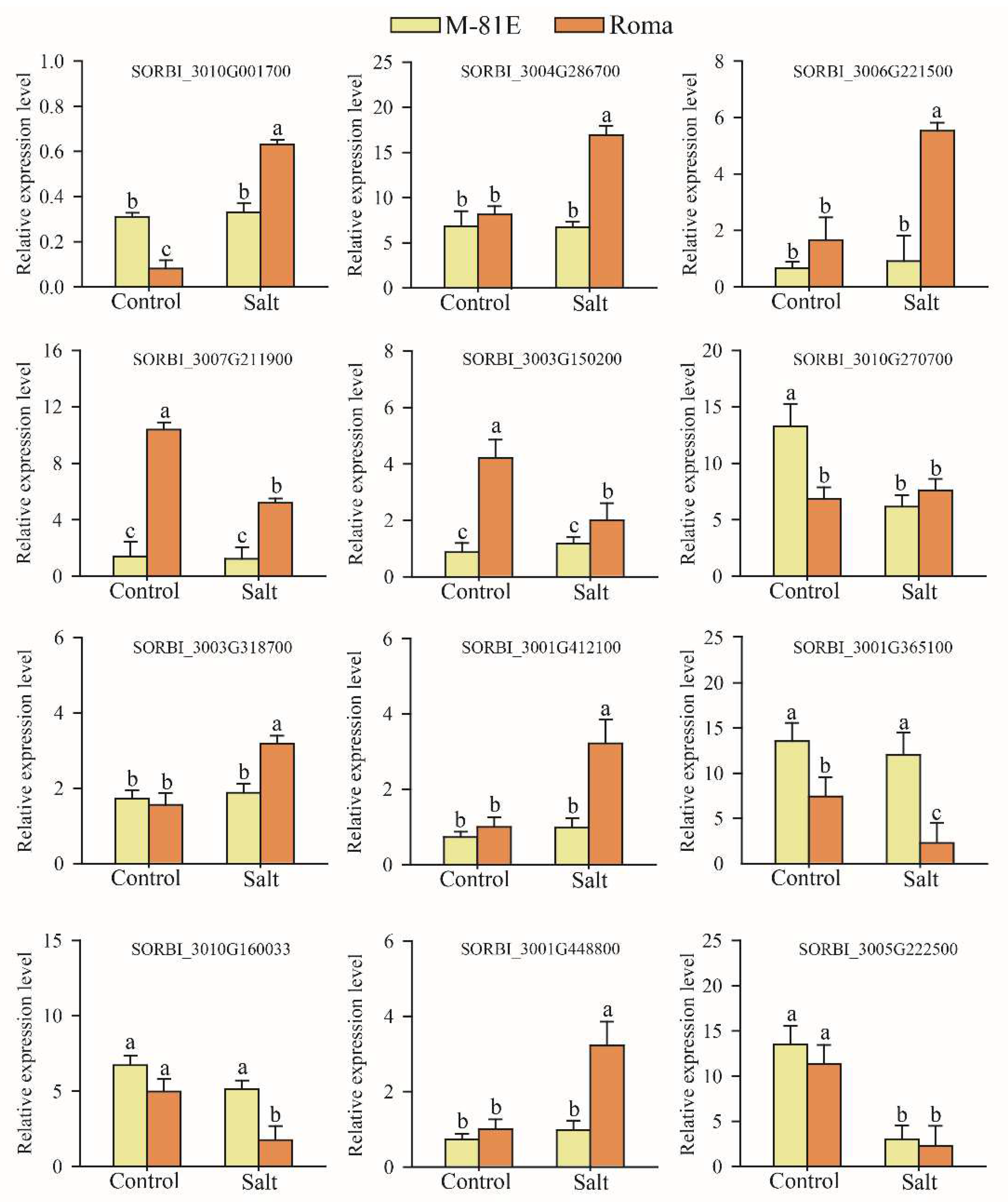
2.6. qRT-PCR Validation of DEGs
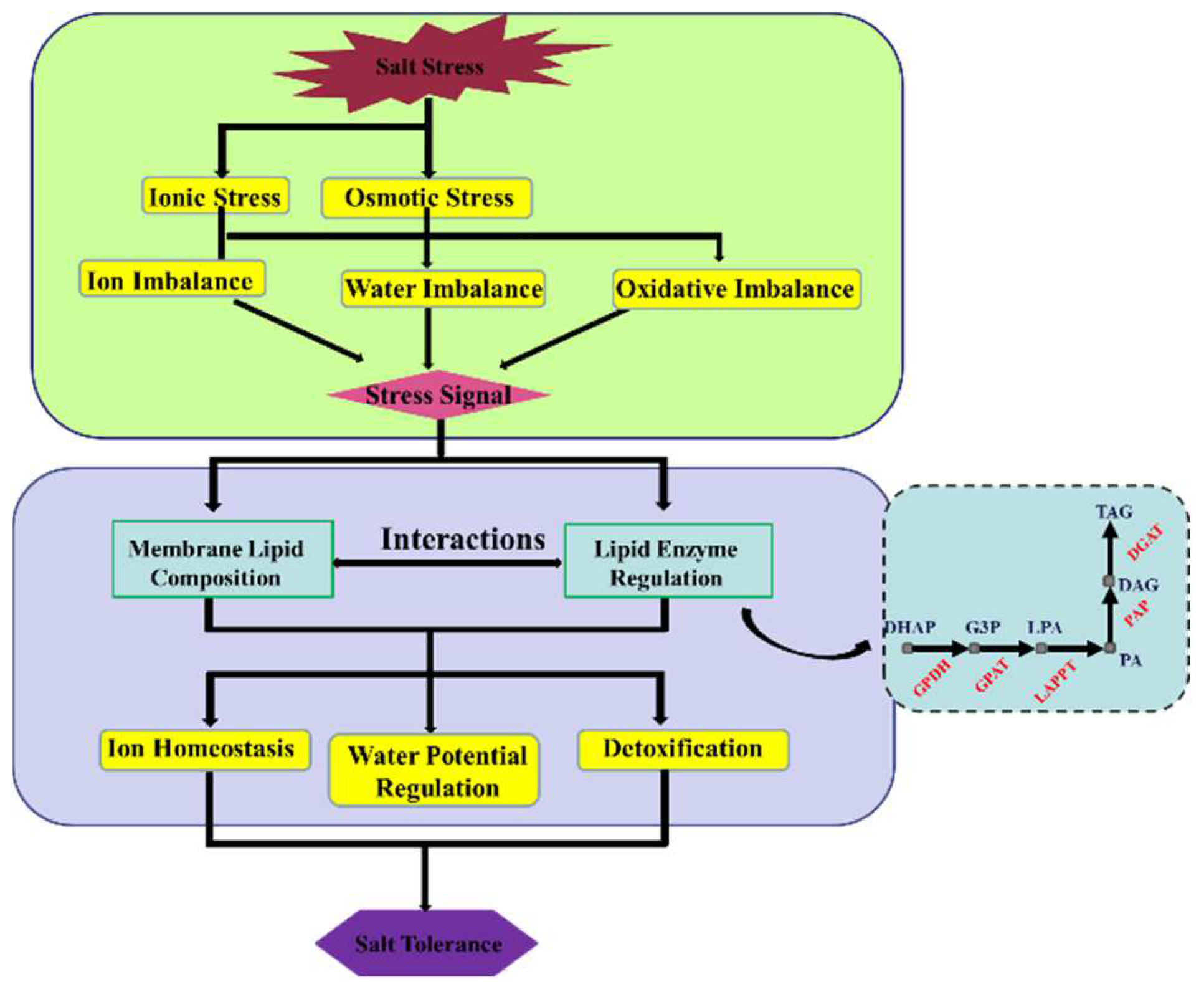
3. Discussion
4. Materials and Methods
4.1. Plant Material and Salt Treatment
4.2. Determination of MDA and Chlorophyll Fluorescence
4.3. Construction and High-Throughput Sequencing of Transcriptome Libraries
4.4. Transcriptome Assembly and Identification
4.5. Identification and Functional Annotation of DEGs
4.6. Quantitative Real-Time PCR Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Song, J.; Shi, G.; Gao, B.; Fan, H.; Wang, B.S. Waterlogging and salinity effects on two Suaeda salsa populations. Physiol. Plant. 2011, 141, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.J.; Sun, H.; Dong, Q.L.; Sun, T.Y.; Jin, Z.X.; Hao, Y.J.; Yao, Y.X. The enhancement of tolerance to salt and cold stresses by modifying the redox state and salicylic acid content via the cytosolic malate dehydrogenase gene in transgenic apple plants. Plant Biotechnol. J. 2016, 14, 1986–1997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, J.P.; Yao, J.F.; Xu, R.R.; You, C.X.; Wang, X.F.; Hao, Y.J. An apple NAC transcription factor enhances salt stress tolerance by modulating the ethylene response. Physiol. Plant. 2018, 164, 279–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Li, Y.; Lu, W.; Meng, F.; Wu, C.A.; Guo, X. Cotton GhMKK5 affects disease resistance, induces HR-like cell death, and reduces the tolerance to salt and drought stress in transgenic Nicotiana benthamiana. J. Exp. Bot. 2012, 63, 3935–3951. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Jiang, S.; Pan, J.; Kong, X.; Zhou, Y.; Liu, Y.; Li, D. The overexpression of a maize mitogen-activated protein kinase gene (ZmMPK5) confers salt stress tolerance and induces defence responses in tobacco. Plant Biol. 2014, 16, 558–570. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.G.; Ma, Q.J.; Sun, C.H.; Sun, M.H.; You, C.X.; Hao, Y.J. Overexpression of MdSOS2L1, a CIPK protein kinase, in-creases the antioxidant metabolites to enhance salt tolerance in apple and tomato. Physiol. Plant. 2016, 156, 201–214. [Google Scholar] [CrossRef]
- Tao, L.I.; Liu, R.; Xin, H.E.; Wang, B.S. Enhancement of Superoxide Dismutase and Catalase Activities and Salt Tolerance of Euhalophyte Suaeda salsa L. by Mycorrhizal Fungus Glomus mosseae. Pedosphere 2012, 22, 217–224. [Google Scholar]
- Shi, X.P.; Ren, J.J.; Yu, Q.; Zhou, S.M.; Ren, Q.P.; Kong, L.J.; Wang, X.L. Overexpression of SDH confers tolerance to salt and osmotic stress, but decreases ABA sensitivity in Arabidopsis. Plant Biol. 2018, 20, 327–337. [Google Scholar] [CrossRef]
- Zhao, B.; Wu, T.T.; Ma, S.S.; Jiang, D.J.; Bie, X.M.; Sui, N.; Zhang, X.S.; Wang, F. TaD27-B gene controls the tiller number in hexaploid wheat. Plant Biotechnol. J. 2020, 18, 513–525. [Google Scholar] [CrossRef] [Green Version]
- Munns, R.; Tester, M. Mechanism of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Elkahoui, S.; Smaoui, A.; Zarrouk, M.; Ghrir, R.; Limam, F. Salt-induced lipid changes in Catharanthus roseus cultured cell suspensions. Phytochemistry 2004, 65, 1911–1917. [Google Scholar] [CrossRef] [PubMed]
- Lopéz-Pérez, L.; Martinez-Ballesta, M.C.; Maurel, C.; Carvajal, M. Changes in plasma membrane lipids, aquaporins and proton pump of broccoli roots, as an adaptation mechanism to salinity. Phytochemistry 2009, 70, 492–500. [Google Scholar] [CrossRef] [PubMed]
- Kerkeb, L.; Donaire, J.P.; Venema, K.; Rodriguez-Rosales, M.P. Tolerance to NaCl induces changes in plasma membrane lipid composition, fluidity and H+-ATPase activity of tomato calli. Physiol. Plant. 2001, 113, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Mansour, M.M.F.; Salama, K.H.A. Cellular basis of salinity tolerance in plants. Environ. Exp. Bot. 2004, 52, 113–122. [Google Scholar] [CrossRef]
- Filek, M.; Walas, S.; Mrowiec, H.; Rudolphy-Skórska, E.; Sieprawska, A.; Biesaga-Kościelniak, J. Membrane permeability and micro- and macroelement accumulation in spring wheat cultivars during the short-term effect of salinity- and PEG-induced water stress. Acta Physiol. Plant. 2012, 34, 985–995. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.S.; Yu, C.; Tang, X.F.; Zhu, Z.J.; Ma, N.N.; Meng, Q.W. A tomato endoplasmic reticulum (ER)-type omega-3 fatty acid desaturase (LeFAD3) functions in early seedling tolerance to salinity stress. Plant Cell Rep. 2014, 33, 131–142. [Google Scholar] [CrossRef]
- Szymanski, J.; Brotman, Y.; Willmitzer, L.; Cuadros-Inostroza, A. Linking gene expression and membrane lipid composition of Arabidopsis. Plant Cell 2014, 26, 915–928. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.X.; Li, J.; Liu, Z.; Yin, J.; Chang, Z.Y.; Rong, C.; Wu, J.L.; Bi, F.C.; Yao, N. The Arabidopsis ceramidase AtACER functions in disease resistance and salt tolerance. Plant J. 2015, 81, 767–780. [Google Scholar] [CrossRef]
- Kejik, Z.; Briza, T.; Kralova, J.; Mikula, I.; Pouckova, P.; Martasek, P.; Kral, V. New method for recognition of sterol signalling molecules: Methinium salts as receptors for sulphated steroids. Steroids 2015, 94, 15–20. [Google Scholar] [CrossRef]
- Gidda, S.K.; Shockey, J.M.; Rothstein, S.J.; Dyer, J.M.; Mullen, R.T. Arabidopsis thaliana GPAT8 and GPAT9 are localized to the ER and possess distinct ER retrieval signals: Functional divergence of the dilysine ER retrieval motif in plant cells. Plant Physiol. Biochem. 2009, 47, 867–879. [Google Scholar] [CrossRef]
- Lin, Y.T.; Chen, L.J.; Herrfurth, C.; Feussner, I.; Li, H.M. Reduced Biosynthesis of Digalactosyldiacylglycerol, a Major Chlo-roplast Membrane Lipid, Leads to Oxylipin Overproduction and Phloem Cap Lignification in Arabidopsis. Plant Cell 2016, 28, 219–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moellering, E.R.; Muthan, B.; Benning, C.J. Freezing Tolerance in Plants Requires Lipid Remodeling at the Outer Chloroplast Membrane. Science 2010, 330, 226–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sui, N.; Li, M.; Li, K.; Song, J.; Wang, B.S. Increase in unsaturated fatty acids in membrane lipids of Suaeda salsa L. enhances protection of photosystem II under high salinity. Photosynthetica 2010, 48, 623–629. [Google Scholar] [CrossRef]
- Mansour, M.M.F. The plasma membrane transport systems and adaptation to salinity. J. Plant Physiol. 2014, 17, 1787–1800. [Google Scholar] [CrossRef] [PubMed]
- Yasar, F.; Uzal, O.; Ozpay, T. Changes of the lipid peroxidation and chlorophyll amount of green bean genotypes under drought stress. Afr. J. Agric. Res. 2010, 5, 2705–2709. [Google Scholar]
- Li, Q.Y.; Niu, H.B.; Yin, J.; Wang, M.B.; Shao, H.B.; Deng, D.Z.; Chen, X.X.; Ren, J.P.; Li, Y.C. Protective role of exogenous nitric oxide against oxidative-stress induced by salt stress in barley (Hordeum vulgare). Colloid Surf. B. 2008, 65, 220–225. [Google Scholar] [CrossRef]
- Li, M.; Guo, S.; Xu, Y.; Meng, Q.; Li, G.; Yang, X. Glycine betaine-mediated potentiation of HSP gene expression involves calcium signaling pathways in tobacco exposed to NaCl stress. Physiol. Plant. 2014, 150, 63–75. [Google Scholar] [CrossRef]
- Kumari, A.; Das, P.; Parida, A.K.; Agarwal, P.K. Proteomics, metabolomics, and ionomics perspectives of salinity tolerance in halophytes. Front. Plant Sci. 2015, 6, 537. [Google Scholar] [CrossRef] [Green Version]
- Salama, K.H.A.; Mansour, M.M.F.; Ali, F.Z.M.; Abou-Hadid, A.F. NaCl-induced changes in plasma membrane lipids and proteins of Zea mays L. cultivars differing in their response to salinity. Acta Physiol. Plant 2007, 29, 351–359. [Google Scholar] [CrossRef]
- Song, Y.; Li, J.; Sui, Y.; Han, G.; Zhang, Y.; Guo, S.; Sui, N. The sweet sorghum SbWRKY50 is negatively involved in salt re-sponse by regulating ion homeostasis. Plant Mol. Biol. 2020, 102, 603–614. [Google Scholar] [CrossRef]
- Sun, X.; Zheng, H.; Li, J.; Liu, L.; Zhang, X.; Sui, N. Comparative Transcriptome Analysis Reveals New lncRNAs Responding to Salt Stress in Sweet Sorghum. Front. Bioeng. Biotechnol. 2020, 8, 331–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Song, Y.; Zheng, H.; Yi, Z.; Sui, N. NADP-Malate Dehydrogenase of Sweet Sorghum Improves Salt Tolerance of Arabidopsis thaliana. J. Agric. Food Chem. 2018, 66, 5992–6002. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Li, J.L.; Liu, L.N.; Xie, Q.; Sui, N. Photosynthetic Regulation Under Salt Stress and Salt-Tolerance Mechanism of Sweet Sorghum. Front. Plant Sci. 2019, 10, 1722. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wang, Y.; Wei, X.; Zhao, X.; Wang, B.S.; Sui, N. Transcription Profiles of Genes Related to Hormonal Regulations Under Salt Stress in Sweet Sorghum. Plant Mol. Biol. Rep. 2017, 35, 586–599. [Google Scholar] [CrossRef]
- Guo, Y.Y.; Tian, S.S.; Liu, S.S.; Wang, W.Q.; Sui, N. Energy dissipation and antioxidant enzyme system protect photosystem II of sweet sorghum under drought stress. Photosynthetica 2018, 56, 861–872. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T. Gene Ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Minoru, K.; Susumu, G.; Shuichi, K.; Yasushi, O.; Masahiro, H.J. The KEGG resource for deciphering the genome. Nucleic Acids Res. 2004, 32, 277–280. [Google Scholar]
- Julkowska, M.M.; Testerink, C.J. Tuning plant signaling and growth to survive salt. Trends Plant Sci. 2015, 20, 586–594. [Google Scholar] [CrossRef] [Green Version]
- Zhai, S.; Gao, Q.; Liu, X.; Sui, Z.; Zhang, J. Overexpression of a Zea mays phospholipase C1 gene enhances drought tolerance in tobacco in part by maintaining stability in the membrane lipid composition. Plant Cell Tissue Organ Cult. 2013, 15, 253–262. [Google Scholar] [CrossRef]
- Guo, Y.; Liu, S.; Yang, Z.; Tian, S.; Sui, N. Responses of Unsaturated Fatty Acid in Membrane Lipid and Antioxidant Enzymes to Chilling Stress in Sweet Sorghum (Sorghum bicolor (L.) Moench) Seedling. J. Agric. Sci. 2016, 8, 71. [Google Scholar] [CrossRef]
- Blokhina, O.; Virolainen, E.; Fagerstedt, K.V. Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann. Bot. 2003, 91, 179–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chalbi, N.; Martínez-Ballesta, M.C.; Youssef, N.B.; Carvajal, M. Intrinsic stability of Brassicaceae plasma membrane in relation to changes in proteins and lipids as a response to salinity. J. Plant Physiol. 2015, 175, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Seliskar, D.M.; Gallagher, J.L. The response of plasma membrane lipid composition in callus of the halophyte Spartina patens (Poaceae) to salinity stress. Am. J. Bot. 2005, 92, 852–858. [Google Scholar] [CrossRef] [PubMed]
- Erdei, L.; Stuiver, B.E.P.; Kuiper, P.J.C. The effect of salinity on lipid composition and on activity of Ca2+ and Mg2+ stimulated ATPases in salt-sensitive and salt-tolerant Plantago species. Physiol. Plant. 1980, 49, 315–319. [Google Scholar] [CrossRef]
- Mansour, M.M.F.; Stadelmann, E.J.; Lee-Stadelmann, O.Y. Salt acclimation of Triticum aestivum by choline chloride: Plant growth, mineral content, and cell permeability. Plant Physiol. Biochem. 1993, 31, 341–348. [Google Scholar]
- Chen, W.L.; Zhang, Q.Q.; Tang, S.H.; Gong, W.; Hong, Y.Y. Glycerol-3-phosphate acyltransferase in lipid metabolism, growth and response to stresses in plants. Plant Physiol. J. 2018, 54, 725–735. [Google Scholar]
- Yang, W.; Pollard, M.; Li-Beisson, Y.; Beisson, F.; Feig, M.; Ohlrogge, J. A distinct type of glycerol-3-phosphate acyltransferase with sn-2 preference and phosphatase activity producing 2-monoacylglycerol. Proc. Natl. Acad. Sci. USA 2010, 107, 12040–12045. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Simpson, J.P.; Li-Beisson, Y.; Beisson, F.; Pollard, M.; Ohlrogge, J.B. A Land-Plant-Specific Glycerol-3-Phosphate Acyltransferase Family in Arabidopsis: Substrate Specificity, sn-2 Preference, and Evolution. Plant Physiol. 2012, 160, 638–652. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Shen, W.; Kazachkov, M.; Chen, G.; Chen, Q.; Carlsson, A.S.; Stymne, S.; Weselake, R.J.; Zou, J. Metabolic inter-actions between the Lands cycle and the Kennedy pathway of glycerolipid synthesis in Arabidopsis developing seeds. Plant Cell 2012, 24, 4652–4669. [Google Scholar] [CrossRef] [Green Version]
- Li-Beisson, Y.; Shorrosh, B.; Beisson, F.; Andersson, M.X.; Arondel, V.; Bates, P.D.; Baud, S.; Bird, D.; Debono, A.; Durrett, T. Acyl-Lipid Metabolism. Arab. Book 2010, 8, e0133. [Google Scholar] [CrossRef] [Green Version]
- Sui, N.; Tian, S.; Wang, W.; Wang, M.J.; Fan, H. Overexpression of Glycerol-3-Phosphate Acyltransferase from Suaeda salsa Improves Salt Tolerance in Arabidopsis. Front. Plant Sci. 2017, 8, 1337. [Google Scholar] [CrossRef] [PubMed]
- Kaup, M.T. A Role for Diacylglycerol Acyltransferase during Leaf Senescence. Plant Physiol. 2002, 129, 1616–1626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Kou, M.; Gao, Z.; Liu, Y.; Xuan, Y.; Liu, Y.; Tang, Z.; Cao, Q.; Li, Z.; Sun, J. Involvement of Phosphatidylserine and Triacylglycerol in the Response of Sweet Potato Leaves to Salt Stress. Front. Plant Sci. 2019, 10, 1086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cases, S.; Stone, S.J.; Zhou, P.; Yen, E.; Tow, B.; Lardizabal, K.D.; Voelker, T.; Farese, R.V.J. Cloning of DGAT2, a Second Mammalian Diacylglycerol Acyltransferase, and Related Family Members. J. Biol. Chem. 2001, 276, 38870. [Google Scholar] [CrossRef] [Green Version]
- Dahlqvist, A.; Stahl, U.; Lenman, M.; Banas, A.; Lee, M.; Sandager, L.; Ronne, H.; Stymne, S. Phospholipid:diacylglycerol acyltransferase: An enzyme that catalyzes the acyl-CoA-independent formation of triacylglycerol in yeast and plants. Proc. Natl. Acad. Sci. USA 2000, 97, 6487–6492. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, M.; Balazadeh, S.; Tohge, T.; Erban, A.; Giavalisco, P.; Kopka, J.; Mueller-Roeber, B.; Fernie, A.R.; Hoefgen, R. Comprehensive dissection of spatiotemporal metabolic shifts in primary, secondary, and lipid metabolism during developmental senescence in Arabidopsis. Plant Physiol. 2013, 162, 1290–1310. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.U.; Lee, K.R.; Jung, S.J.; Shin, H.A.; Go, Y.S.; Suh, M.C.; Kim, J.B. Senescence-inducible LEC2 enhances triacylglycerol accumulation in leaves without negatively affecting plant growth. Plant Biotechnol. J. 2015, 13, 1346–1359. [Google Scholar] [CrossRef] [Green Version]
- Trabelsi, H.; Renaud, J.; Mayer, P.; Boukhchina, S.J. Triacylglycerol and Glycerophospholipid Identification and Accumulation During Ripening of Pistacia lentiscus L. (Lentisc) Fruit. J. Am. Oil Chem. Soc. 2014, 91, 1189–1196. [Google Scholar] [CrossRef]
- Sui, N.; Yang, Z.; Liu, M.; Wang, B.S. Identification and transcriptomic profiling of genes involved in increasing sugar content during salt stress in sweet sorghum leaves. BMC Genom. 2015, 16, 534. [Google Scholar] [CrossRef] [Green Version]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants Without Soil. Circ. Calif. Agric. Exp. Stn. 1950, 347, 2–32. [Google Scholar]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplast I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved recon-struction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Frazee, A.C.; Pertea, G.; Jaffe, A.E.; Langmead, B.; Salzberg, S.L.; Leek, J.T. Ballgown bridges the gap between transcriptome assembly and expression analysis. Nat. Biotechnol. 2015, 33, 243–246. [Google Scholar] [CrossRef] [Green Version]
- Yangyang, D.; Jianqi, L.I.; Songfeng, W.U.; Yunping, Z.; Yaowen, C.; Fuchu, H.E. Integrated nr Database in Protein An-notation System and Its Localization. Comput. Eng. 2006, 32, 71–72. [Google Scholar]
- Apweiler, R.; Bairoch, A.; Wu, C.H.; Barker, W.C.; Boeckmann, B.; Ferro, S.; Gasteiger, E.; Huang, H.; Lopez, R.; Magrane, M.; et al. UniProt: The Universal Protein knowledgebase. Nucleic Acids Res. 2004, 32, 115–119. [Google Scholar] [CrossRef]
- Natale, D.A.; Galperin, M.Y.; Tatusov, R.L.; Koonin, E.V. Using the COG Database to Improve Gene Recognition in Complete Genomes. Genetica 2000, 108, 9–17. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Anotation | M-81E | Roma | ||||
|---|---|---|---|---|---|---|---|
| Log2FC | Regulated | p-Value | Log2FC | Regulated | p-Value | ||
| (A) Glycerolipid Metabolism | |||||||
| SORBI_3010G001700 | phospholipid:diacylglycerol acyltransferase | - | - | - | 2.26 | up | 0.02 |
| SORBI_3004G286700 | phospholipid:diacylglycerol acyltransferase | - | - | - | 1.09 | up | 0.04 |
| SORBI_3006G221500 | 1-acyl-sn-glycerol-3-phosphate acyltransferase(LAPPT) | - | - | - | 1.43 | up | 0.04 |
| SORBI_3007G211900 | 1,2-diacylglycerol 3-beta-galactosyltransferase | - | - | - | −1.65 | down | 0 |
| SORBI_3003G150200 | acylglycerol lipase | - | - | - | −1.41 | down | 0.02 |
| SORBI_3010G270700 | phospholipid:diacylglycerol acyltransferase (PDAT) | −1.18 | down | 0.02 | - | - | - |
| SORBI_3002G067000 | phospholipid:diacylglycerol acyltransferase (PDAT) | −1.05 | down | 0.02 | - | - | - |
| SORBI_3003G360700 | glycerol-3-phosphate O-acyltransferase | 1.65 | up | 0.02 | - | - | - |
| SORBI_3002G417800 | alpha-galactosidase | 1.25 | up | 0.01 | - | - | - |
| SORBI_3001G208100 | alpha-galactosidase | 1.71 | up | 0 | - | - | - |
| SORBI_3001G208200 | alpha-galactosidase | 2.52 | up | 0 | - | - | - |
| (B) Glycerophospholipid Metabolism | |||||||
| SORBI_3006G221500 | 1-acyl-sn-glycerol-3-phosphate acyltransferase | 1.43 | up | 0.04 | |||
| SORBI_3003G318700 | diacylglycerol kinase (ATP) | 1.14 | up | 0.04 | |||
| SORBI_3001G412100 | CDP-diacylglycerol-glycerol-3-phosphate 3-phosphatidyltransferase | 1.53 | up | 0.01 | |||
| SORBI_3001G365100 | phospholipase A1 | −2.56 | down | 0 | |||
| SORBI_3010G160033 | phospholipase A1 | −1.61 | down | 0 | |||
| SORBI_3001G448800 | lysophospholipase II | 1.61 | up | 0.03 | |||
| SORBI_3005G222500 | phospholipase D1/2 | −1.15 | down | 0.02 | −1.88 | down | 0 |
| SORBI_3008G183400 | phospholipase D1/2 | −1.73 | down | 0.01 | |||
| SORBI_3004G157300 | glycerophosphodiester phosphodiesterase | −1.04 | down | 0.01 | −1.68 | down | 0 |
| SORBI_3007G190700 | glycerophosphodiester phosphodiesterase | −1.09 | down | 0.03 | |||
| SORBI_3010G160000 | phospholipase A1 | −3.86 | down | 0.01 | |||
| SORBI_3009G014600 | phospholipase A1 | −1.35 | down | 0.01 | |||
| SORBI_3003G360700 | glycerol-3-phosphate O-acyltransferase (GPAT) | 1.77 | up | 0.03 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, F.; Chen, Z.; Zhang, F.; Zheng, H.; Li, S.; Gao, Y.; Yang, J.; Sui, N. Identification and Transcriptome Analysis of Genes Related to Membrane Lipid Regulation in Sweet Sorghum under Salt Stress. Int. J. Mol. Sci. 2022, 23, 5465. https://doi.org/10.3390/ijms23105465
Wu F, Chen Z, Zhang F, Zheng H, Li S, Gao Y, Yang J, Sui N. Identification and Transcriptome Analysis of Genes Related to Membrane Lipid Regulation in Sweet Sorghum under Salt Stress. International Journal of Molecular Sciences. 2022; 23(10):5465. https://doi.org/10.3390/ijms23105465
Chicago/Turabian StyleWu, Fenghui, Zengting Chen, Fangning Zhang, Hongxiang Zheng, Simin Li, Yinping Gao, Jie Yang, and Na Sui. 2022. "Identification and Transcriptome Analysis of Genes Related to Membrane Lipid Regulation in Sweet Sorghum under Salt Stress" International Journal of Molecular Sciences 23, no. 10: 5465. https://doi.org/10.3390/ijms23105465
APA StyleWu, F., Chen, Z., Zhang, F., Zheng, H., Li, S., Gao, Y., Yang, J., & Sui, N. (2022). Identification and Transcriptome Analysis of Genes Related to Membrane Lipid Regulation in Sweet Sorghum under Salt Stress. International Journal of Molecular Sciences, 23(10), 5465. https://doi.org/10.3390/ijms23105465