
Viability of Glioblastoma Cells and Fibroblasts in the Presence of Imidazole-Containing Compounds


 , and
, and
Abstract
:1. Introduction
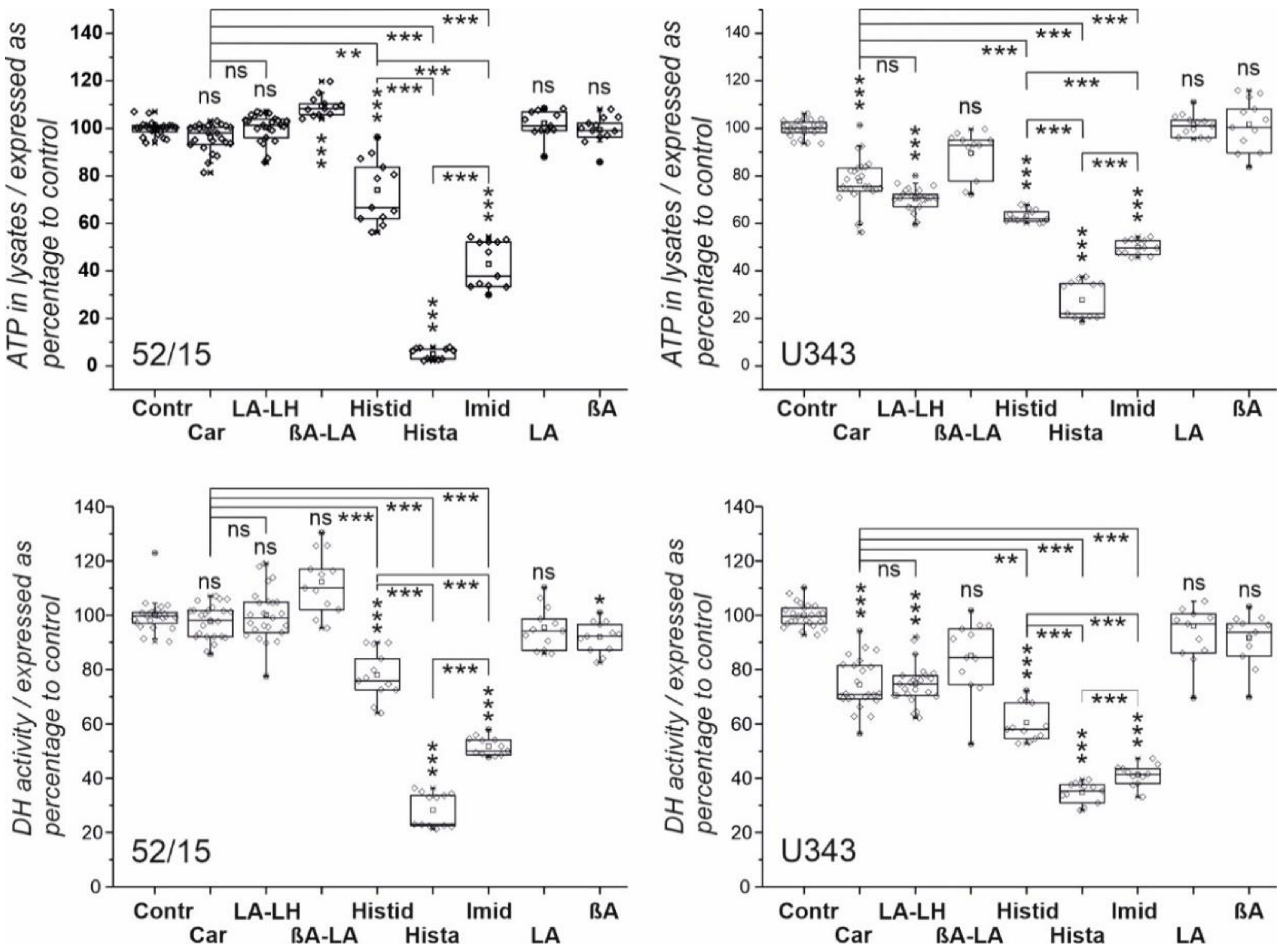
2. Results
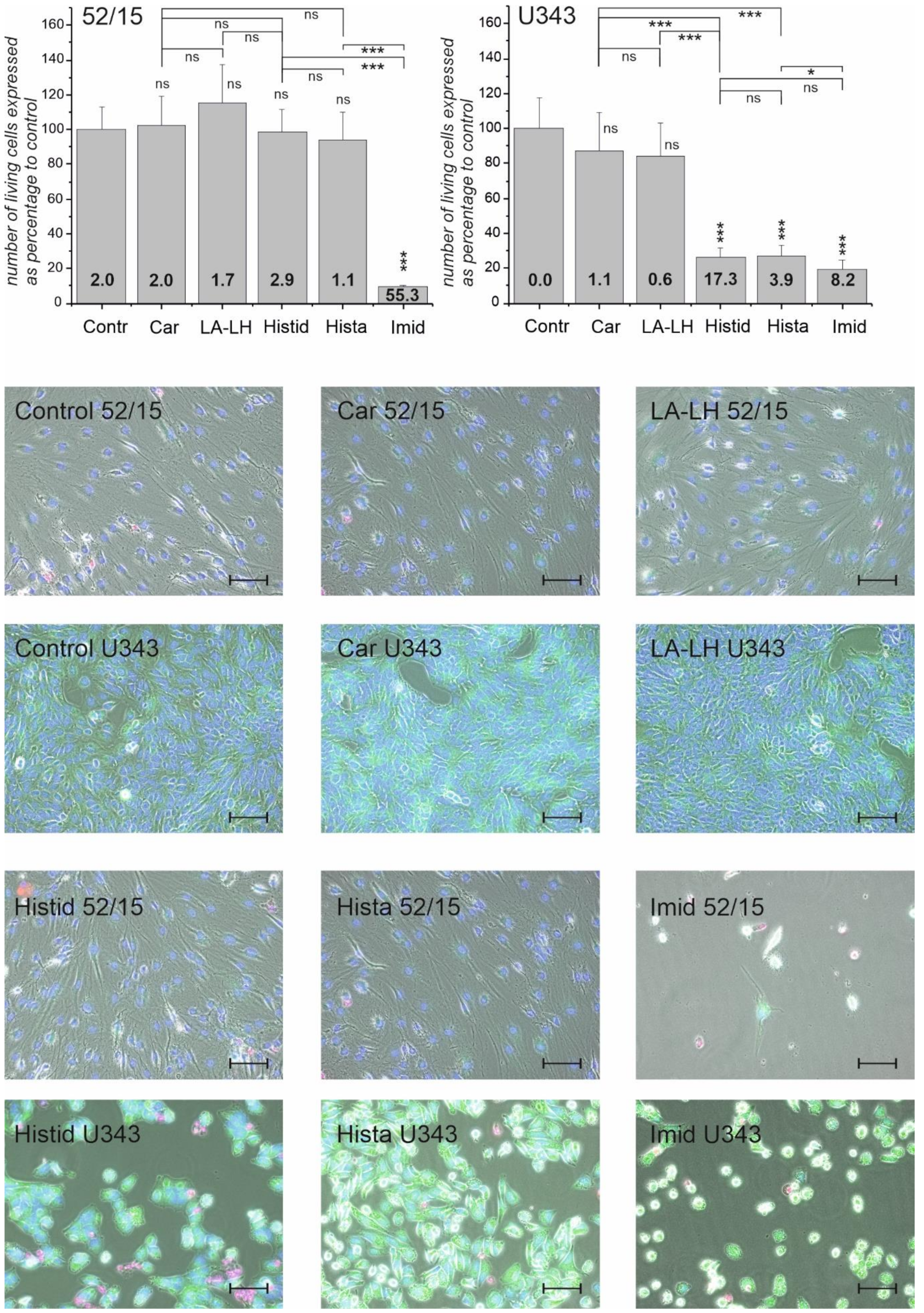
2.1. Viability of Glioblastoma Cells and Patient-Derived Fibroblasts in the Presence of Imidazole-Containing Compounds
2.2. Necrosis and Morphological Changes of GBM Cells and Patient-Derived Fibroblasts after Treatment with Different Imidazole-Containing Compounds
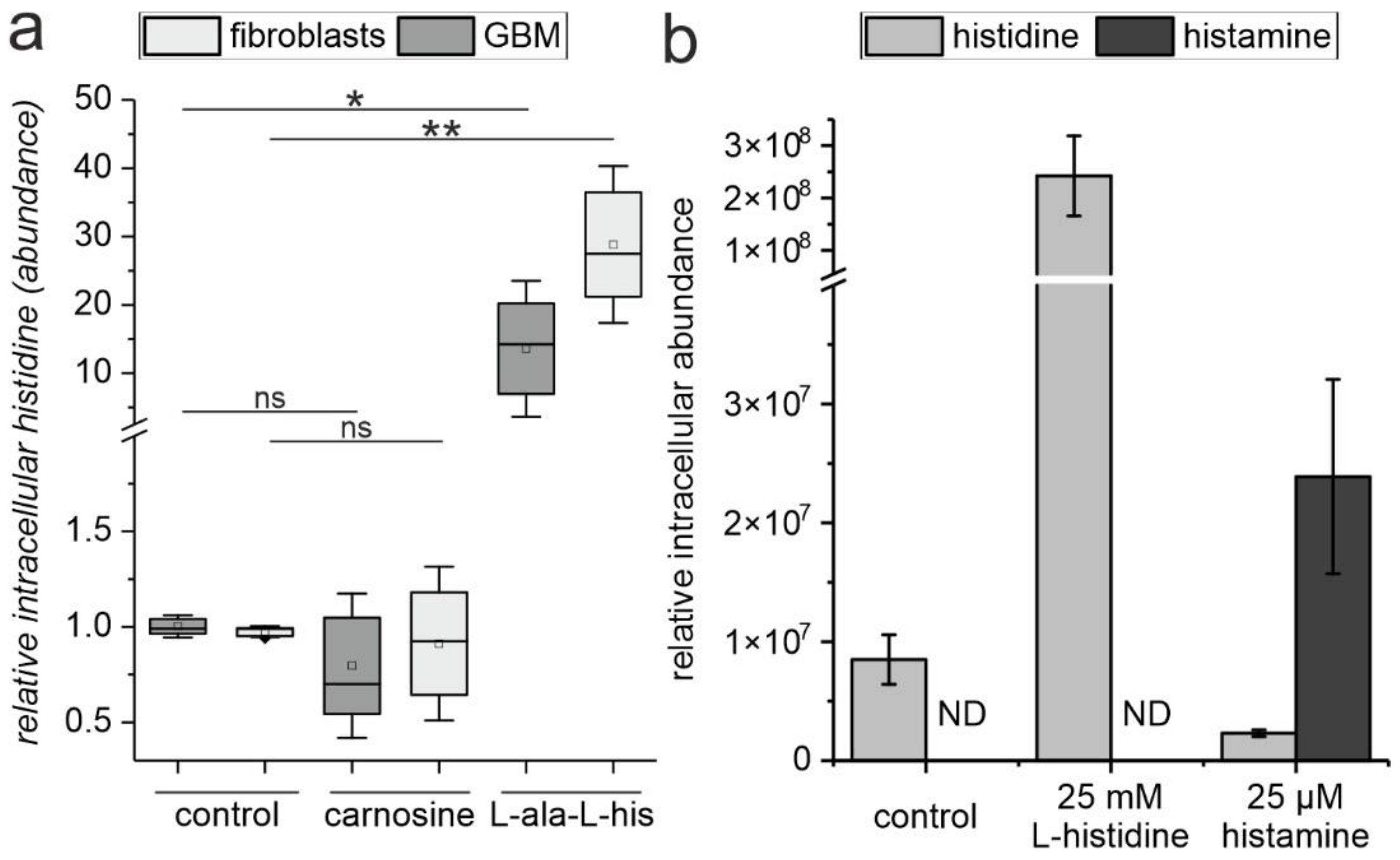
2.3. The Release of L-Histidine from L-Alanyl-L-Histidine Does Not Result in the Formation of Histamine
3. Discussion
4. Material and Methods
4.1. Reagents
4.2. Cell Lines and Fibroblast Cultures
4.3. Cell Viability Assays
4.4. Staining and Determination of Live and Dead Cells
4.5. Determination of Intracellular L-Histidine
4.6. HPLC–MS Set Up and Data Analysis
4.7. Statistical Analysis and Graphical Representation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ostrom, Q.T.; Patil, N.; Cioffi, G.; Waite, K.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary Brain and Other Central Nervous System Tumors Diagnosed in the United States in 2013–2017. Neuro-Oncology 2020, 22, iv1–iv96. [Google Scholar] [CrossRef] [PubMed]
- Hegi, M.E.; Diserens, A.-C.; Gorlia, T.; Hamou, M.-F.; de Tribolet, N.; Weller, M.; Kros, J.M.; Hainfellner, J.A.; Mason, W.; Mariani, L.; et al. MGMT gene silencing and benefit from temozolomide in glioblastoma. N. Engl. J. Med. 2005, 352, 997–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gulewitsch, W.; Amiradzibi, S. Ueber das Carnosin, eine neue organische Base des Fleischextraktes. Ber. Dtsch. Chem. Ges. 1900, 33, 1902–1903. [Google Scholar] [CrossRef] [Green Version]
- Turner, M.D.; Sale, C.; Garner, A.C.; Hipkiss, A.R. Anti-cancer actions of carnosine and the restoration of normal cellular homeostasis. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 119117. [Google Scholar] [CrossRef]
- Hipkiss, A.R.; Gaunitz, F. Inhibition of tumour cell growth by carnosine: Some possible mechanisms. Amino Acids 2014, 46, 327–337. [Google Scholar] [CrossRef]
- Oppermann, H.; Elsel, S.; Birkemeyer, C.; Meixensberger, J.; Gaunitz, F. Erythrocytes Prevent Degradation of Carnosine by Human Serum Carnosinase. Int. J. Mol. Sci. 2021, 22, 12802. [Google Scholar] [CrossRef]
- Letzien, U.; Oppermann, H.; Meixensberger, J.; Gaunitz, F. The antineoplastic effect of carnosine is accompanied by induction of PDK4 and can be mimicked by L-histidine. Amino Acids 2014, 46, 1009–1019. [Google Scholar] [CrossRef]
- Nagai, K.; Suda, T. Antineoplastic effects of carnosine and beta-alanine—Physiological considerations of its antineoplastic effects. J. Physiol. Soc. Jpn. 1986, 48, 741–747. [Google Scholar]
- Shen, Y.; Yang, J.; Li, J.; Shi, X.; Ouyang, L.; Tian, Y.; Lu, J. Carnosine inhibits the proliferation of human gastric cancer SGC-7901 cells through both of the mitochondrial respiration and glycolysis pathways. PLoS ONE 2014, 9, e104632. [Google Scholar] [CrossRef] [Green Version]
- Iovine, B.; Iannella, M.L.; Nocella, F.; Pricolo, M.R.; Baldi, M.R.; Bevilacqua, M.A. Carnosine inhibits KRas-mediated HCT-116 proliferation by affecting ATP and ROS production. Cancer Lett. 2012, 315, 122–128. [Google Scholar] [CrossRef]
- Ditte, Z.; Ditte, P.; Labudova, M.; Simko, V.; Iuliano, F.; Zatovicova, M.; Csaderova, L.; Pastorekova, S.; Pastorek, J. Carnosine inhibits carbonic anhydrase IX-mediated extracellular acidosis and suppresses growth of HeLa tumor xenografts. BMC Cancer 2014, 14, 358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renner, C.; Seyffarth, A.; de Arriba, S.; Meixensberger, J.; Gebhardt, R.; Gaunitz, F. Carnosine Inhibits Growth of Cells Isolated from Human Glioblastoma Multiforme. Int. J. Pept. Res. Ther. 2008, 14, 127–135. [Google Scholar] [CrossRef]
- Oppermann, H.; Purcz, K.; Birkemeyer, C.; Baran-Schmidt, R.; Meixensberger, J.; Gaunitz, F. Carnosine’s inhibitory effect on glioblastoma cell growth is independent of its cleavage. Amino Acids 2019, 51, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Holliday, R.; McFarland, G.A. Inhibition of the growth of transformed and neoplastic cells by the dipeptide carnosine. Br. J. Cancer 1996, 73, 966–971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oppermann, H.; Dietterle, J.; Purcz, K.; Morawski, M.; Eisenlöffel, C.; Müller, W.; Meixensberger, J.; Gaunitz, F. Carnosine selectively inhibits migration of IDH-wildtype glioblastoma cells in a co-culture model with fibroblasts. Cancer Cell Int. 2018, 18, 111. [Google Scholar] [CrossRef]
- Rauen, U.; Klempt, S.; de Groot, H. Histidine-induced injury to cultured liver cells, effects of histidine derivatives and of iron chelators. Cell. Mol. Life Sci. 2007, 64, 192–205. [Google Scholar] [CrossRef]
- Qiu, J.; Hauske, S.J.; Zhang, S.; Rodriguez-Niño, A.; Albrecht, T.; Pastene, D.O.; van den Born, J.; van Goor, H.; Ruf, S.; Kohlmann, M.; et al. Identification and characterisation of carnostatine (SAN9812), a potent and selective carnosinase (CN1) inhibitor with in vivo activity. Amino Acids 2019, 51, 7–16. [Google Scholar] [CrossRef] [Green Version]
- Gardner, M.L.G.; Illingworth, K.M.; Kelleher, J.; Wood, D. Intestinal-Absorption of the Intact Peptide Carnosine in Man, and Comparison with Intestinal Permeability to Lactulose. J. Physiol. 1991, 439, 411–422. [Google Scholar] [CrossRef]
- Baraniuk, J.N.; El-Amin, S.; Corey, R.; Rayhan, R.; Timbol, C. Carnosine Treatment for Gulf War Illness: A Randomized Controlled Trial. GJHS 2013, 5, 69–81. [Google Scholar] [CrossRef]
- Hajizadeh-Zaker, R.; Ghajar, A.; Mesgarpour, B.; Afarideh, M.; Mohammadi, M.-R.; Akhondzadeh, S. l-Carnosine as an Adjunctive Therapy to Risperidone in Children with Autistic Disorder: A Randomized, Double-Blind, Placebo-Controlled Trial. J. Child Adolesc. Psychopharmacol. 2018, 28, 74–81. [Google Scholar] [CrossRef]
- Mehrazad-Saber, Z.; Kheirouri, S.; Noorazar, S.-G. Effects of l-Carnosine Supplementation on Sleep Disorders and Disease Severity in Autistic Children: A Randomized, Controlled Clinical Trial. Basic Clin. Pharmacol. Toxicol. 2018, 123, 72–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akhtar, J.; Khan, A.A.; Ali, Z.; Haider, R.; Shahar Yar, M. Structure-activity relationship (SAR) study and design strategies of nitrogen-containing heterocyclic moieties for their anticancer activities. Eur. J. Med. Chem. 2017, 125, 143–189. [Google Scholar] [CrossRef] [PubMed]
- Siwach, A.; Verma, P.K. Synthesis and therapeutic potential of imidazole containing compounds. BMC Chem. 2021, 15, 12. [Google Scholar] [CrossRef] [PubMed]
- Saunders, B.; Elliott-Sale, K.; Artioli, G.G.; Swinton, P.A.; Dolan, E.; Roschel, H.; Sale, C.; Gualano, B. β-alanine supplementation to improve exercise capacity and performance: A systematic review and meta-analysis. Br. J. Sports Med. 2017, 51, 658–669. [Google Scholar] [CrossRef]
- Lopachev, A.V.; Abaimov, D.A.; Filimonov, I.S.; Kulichenkova, K.N.; Fedorova, T.N. An assessment of the transport mechanism and intraneuronal stability of L-carnosine. Amino Acids 2021, 1–8. [Google Scholar] [CrossRef]
- Long, Y.; Wang, D. Inhibition of Colon Cancer Cell Growth by Imidazole Through Activation of Apoptotic Pathway. Med. Sci. Monit. 2019, 25, 7597–7604. [Google Scholar] [CrossRef]
- Zhang, L.; Yao, K.; Fan, Y.; He, P.; Wang, X.; Hu, W.; Chen, Z. Carnosine protects brain microvascular endothelial cells against rotenone-induced oxidative stress injury through histamine H₁ and H₂ receptors in vitro. Clin. Exp. Pharmacol. Physiol. 2012, 39, 1019–1025. [Google Scholar] [CrossRef]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [Green Version]
- Bae, O.-N.; Majid, A. Role of histidine/histamine in carnosine-induced neuroprotection during ischemic brain damage. Brain Res. 2013, 1527, 246–254. [Google Scholar] [CrossRef]
- Shi, Y.; Lim, S.K.; Liang, Q.; Iyer, S.V.; Wang, H.-Y.; Wang, Z.; Xie, X.; Sun, D.; Chen, Y.-J.; Tabar, V.; et al. Gboxin is an oxidative phosphorylation inhibitor that targets glioblastoma. Nature 2019, 567, 341–346. [Google Scholar] [CrossRef]
- Morelli, M.B.; Amantini, C.; Nabissi, M.; Cardinali, C.; Santoni, M.; Bernardini, G.; Santoni, A.; Santoni, G. Axitinib induces senescence-associated cell death and necrosis in glioma cell lines: The proteasome inhibitor, bortezomib, potentiates axitinib-induced cytotoxicity in a p21(Waf/Cip1) dependent manner. Oncotarget 2017, 8, 3380–3395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uzlikova, M.; Nohynkova, E. The effect of metronidazole on the cell cycle and DNA in metronidazole-susceptible and -resistant Giardia cell lines. Mol. Biochem. Parasitol. 2014, 198, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Wenig, P.; Odermatt, J. OpenChrom: A cross-platform open source software for the mass spectrometric analysis of chromatographic data. BMC Bioinform. 2010, 11, 405. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Fibroblasts | Glioblastoma Cells | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Culture | 13/16 | 90/15 | 69/15 | 60/15 | 52/15 | U343 | T98G | U87 | G55T2 | |||||||||
| Assay | ATP | DH | ATP | DH | ATP | DH | ATP | DH | ATP | DH | ATP | DH | ATP | DH | ATP | DH | ATP | DH |
| Carnosine | ||||||||||||||||||
| LA-LH | ||||||||||||||||||
| Histidine | ||||||||||||||||||
| Imidazole | ||||||||||||||||||
| Histamine | ||||||||||||||||||
| bAla-L-Ala | ||||||||||||||||||
| L-Ala | ||||||||||||||||||
| b-Ala | ||||||||||||||||||
| Reduction to control (%): | 100–90 | 90–80 | 80–70 | 70–60 | 60–50 | 50–40 | <40 | |||||||||||
| Fibroblasts | Glioblastoma | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| culture | 90/15 | 69/15 | 60/15 | 52/15 | U343 | T98G | G55T2 | U87 | ||||||||
| % | ratio | % | ratio | % | ratio | % | ratio | % | ratio | % | ratio | % | ratio | % | ratio | |
| Car | ||||||||||||||||
| LALH | ||||||||||||||||
| Histid | ||||||||||||||||
| Imid | nd | |||||||||||||||
| Hista | ||||||||||||||||
| living cells to control (%) | ≥ 100 | <100–90 | <90–80 | <80–70 | <70–60 | <60–50 | <50–40 | <40–30 | <30–20 | <20–10 | <10 | |||||
| ratio dead/living | ≤0.5 | >0.5–1 | >1–2 | >2–4 | >4–8 | >8–16 | >16–32 | >32–64 | >65 | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seidel, E.C.; Birkemeyer, C.; Baran-Schmidt, R.; Meixensberger, J.; Oppermann, H.; Gaunitz, F. Viability of Glioblastoma Cells and Fibroblasts in the Presence of Imidazole-Containing Compounds. Int. J. Mol. Sci. 2022, 23, 5834. https://doi.org/10.3390/ijms23105834
Seidel EC, Birkemeyer C, Baran-Schmidt R, Meixensberger J, Oppermann H, Gaunitz F. Viability of Glioblastoma Cells and Fibroblasts in the Presence of Imidazole-Containing Compounds. International Journal of Molecular Sciences. 2022; 23(10):5834. https://doi.org/10.3390/ijms23105834
Chicago/Turabian StyleSeidel, Elisabeth Christiane, Claudia Birkemeyer, Rainer Baran-Schmidt, Jürgen Meixensberger, Henry Oppermann, and Frank Gaunitz. 2022. "Viability of Glioblastoma Cells and Fibroblasts in the Presence of Imidazole-Containing Compounds" International Journal of Molecular Sciences 23, no. 10: 5834. https://doi.org/10.3390/ijms23105834
APA StyleSeidel, E. C., Birkemeyer, C., Baran-Schmidt, R., Meixensberger, J., Oppermann, H., & Gaunitz, F. (2022). Viability of Glioblastoma Cells and Fibroblasts in the Presence of Imidazole-Containing Compounds. International Journal of Molecular Sciences, 23(10), 5834. https://doi.org/10.3390/ijms23105834