
Persistent Infection of a Canine Histiocytic Sarcoma Cell Line with Attenuated Canine Distemper Virus Expressing Vasostatin or Granulocyte-Macrophage Colony-Stimulating Factor


 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. DH82 Cell Growth Rate and Virus Replication Ability Are Independent of the Virus Strain
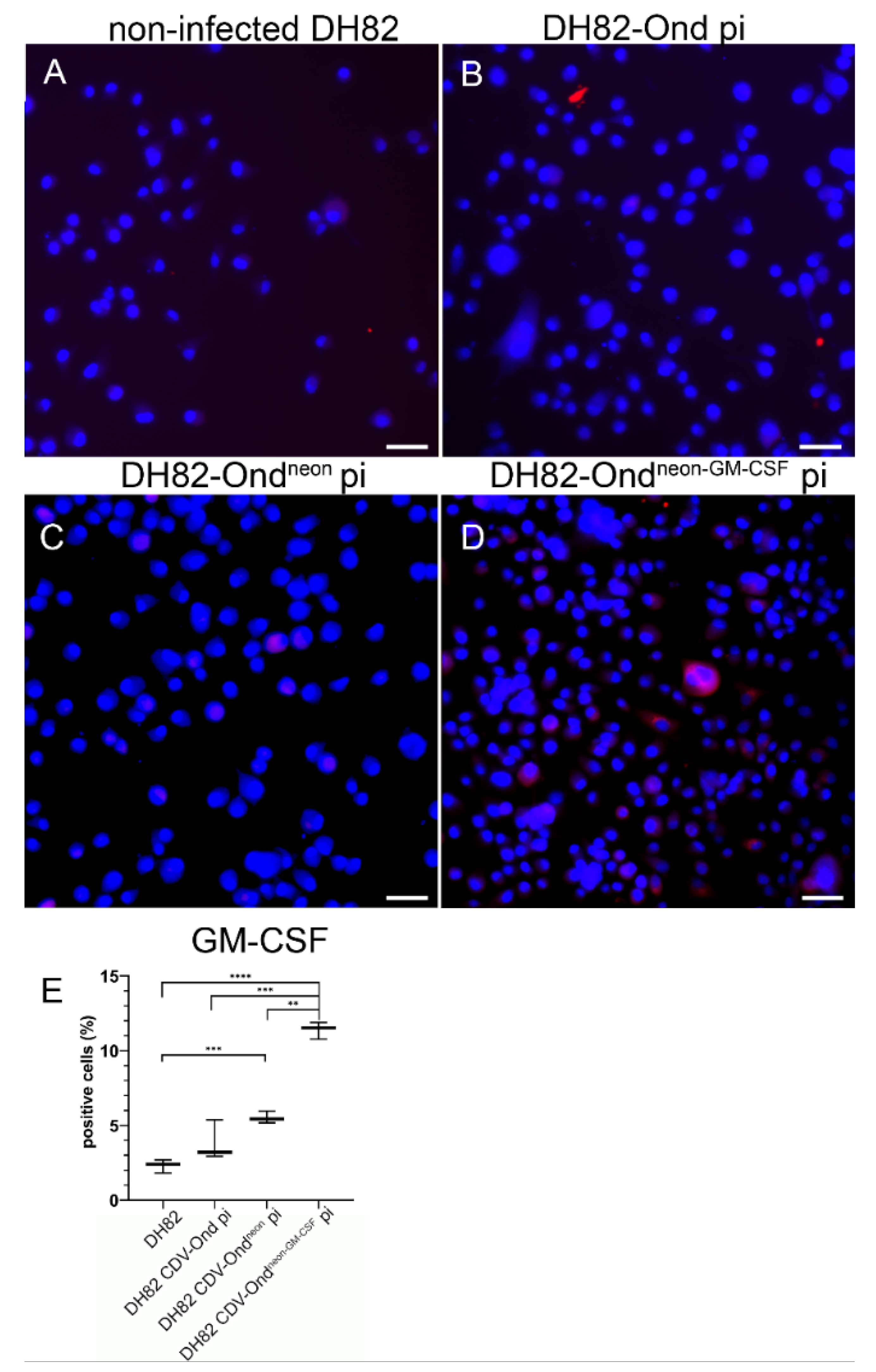
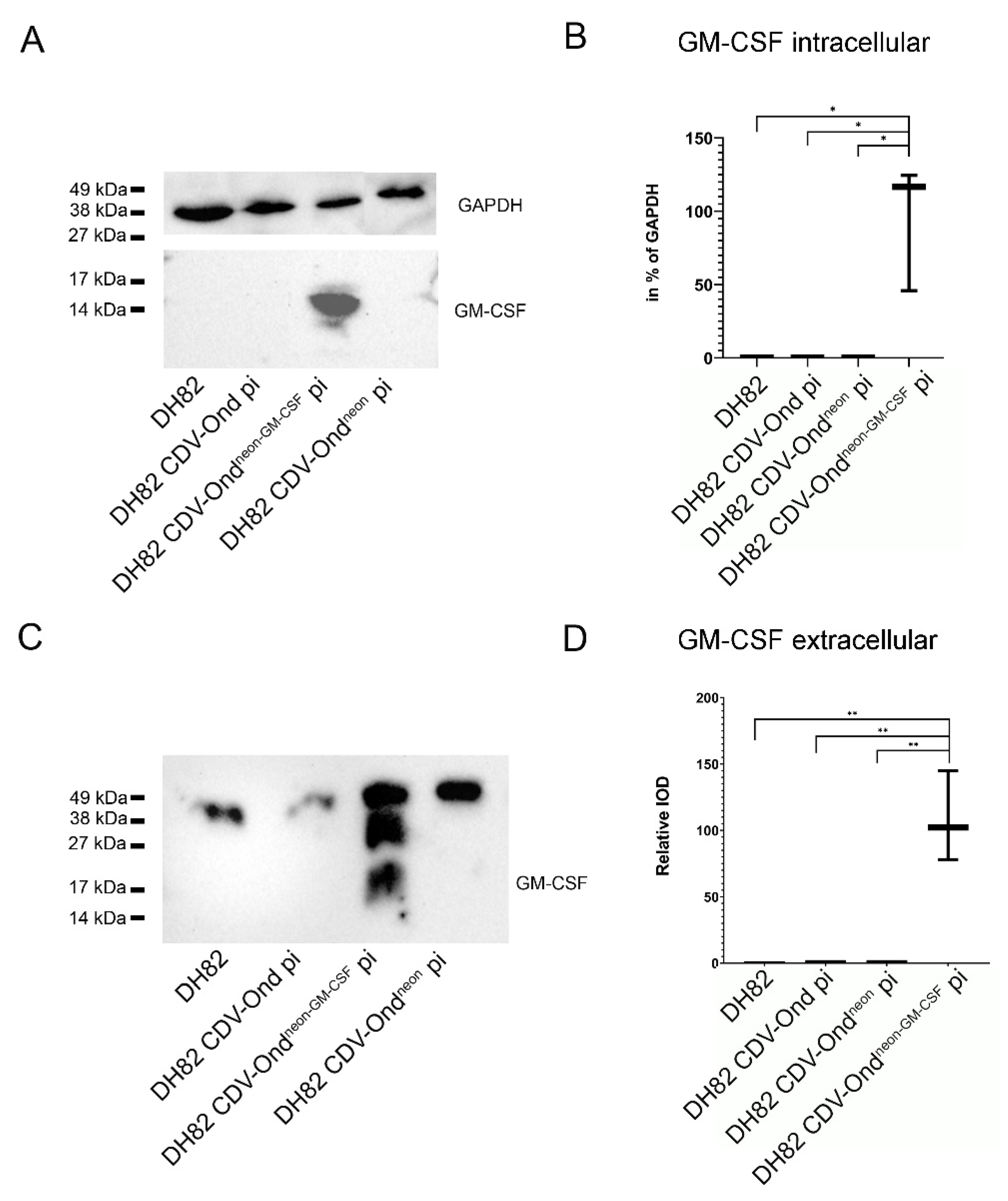
2.2. GM-CSF Is Increased in DH82 Cells Persistently Infected with CDV-Ondneon-GM-CSF on the Transcriptional and Protein Level
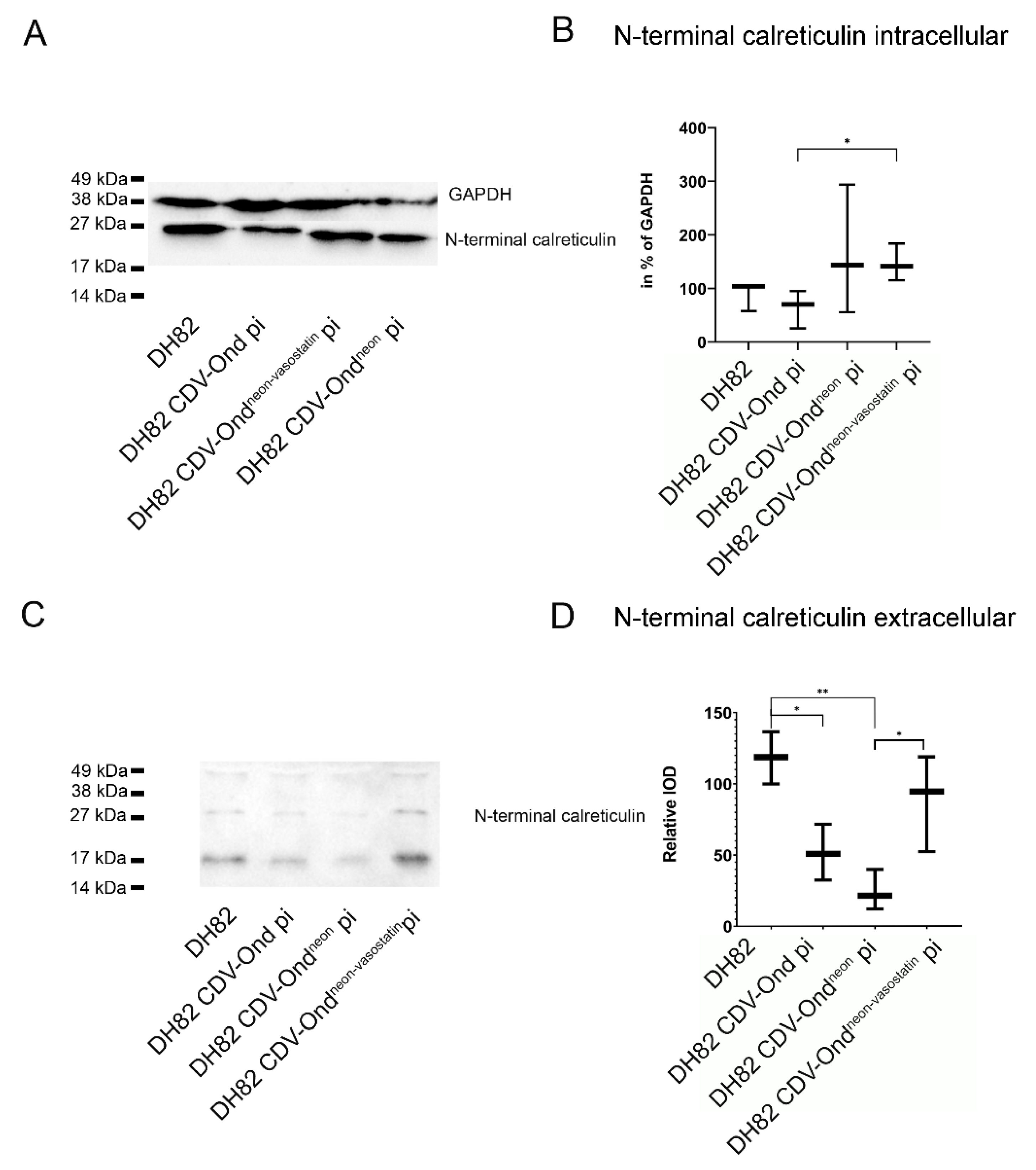
2.3. DH82 Cells Persistently Infected with CDV-Ondneon-vasostatin Show an Increased Number of Vasostatin mRNA Transcripts
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Cumulative Population Doubling Assay
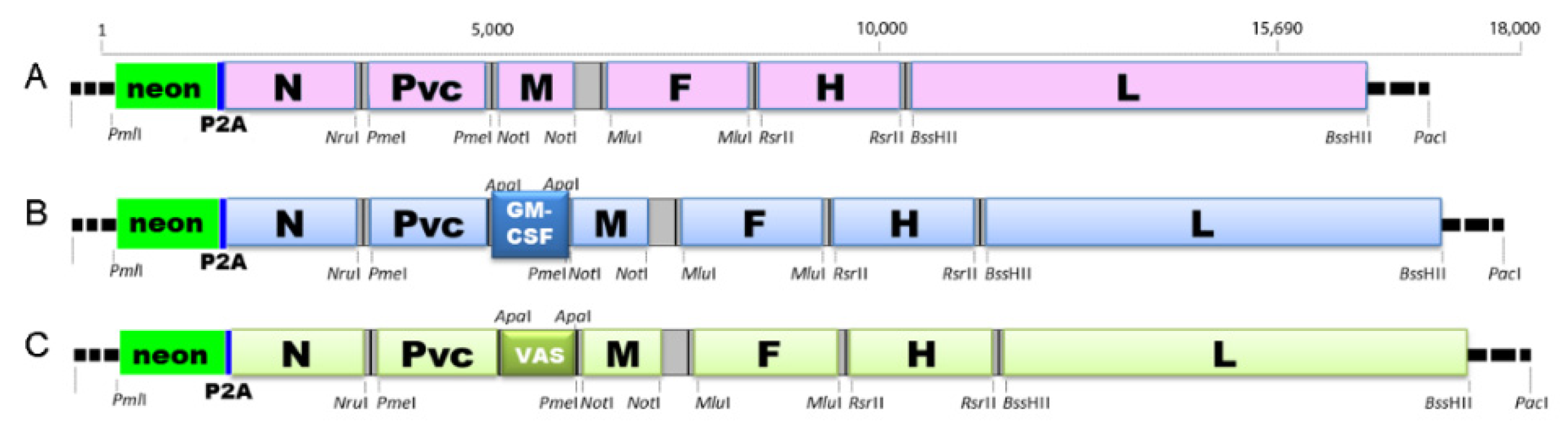
4.3. Genetically Engineered Virus Strains
4.4. Virus Titration
4.5. RNA Isolation and cDNA Synthesis
4.6. Primer Design
4.7. Qualitative PCR
4.8. Reverse Transcription Quantitative RT-qPCR
4.9. Immunoblotting
4.10. Immunohistochemistry
4.11. Immunofluorescence
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lewis, T.W.; Wiles, B.M.; Llewellyn-Zaidi, A.M.; Evans, K.M.; O’Neill, D.G. Longevity and mortality in Kennel Club registered dog breeds in the UK in 2014. Canine Genet. Epidemiol. 2018, 5, 10. [Google Scholar] [CrossRef] [PubMed]
- Thongtharb, A.; Uchida, K.; Chambers, J.K.; Nakayama, H. Variations in histiocytic differentiation of cell lines from canine cerebral and articular histiocytic sarcomas. Vet Pathol. 2017, 54, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Skorupski, K.A.; Rodriguez, C.O.; Krick, E.L.; Clifford, C.A.; Ward, R.; Kent, M.S. Long-term survival in dogs with localized histiocytic sarcoma treated with CCNU as an adjuvant to local therapy. Vet. Comp. Oncol. 2009, 7, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Berg, D.R.; Offord, C.P.; Kemler, I.; Ennis, M.K.; Chang, L.; Paulik, G.; Bajzer, Z.; Neuhauser, C.; Dingli, D. In vitro and in silico multidimensional modeling of oncolytic tumor virotherapy dynamics. PLoS Comput. Biol. 2019, 15, e1006773. [Google Scholar] [CrossRef] [Green Version]
- Lapp, S.; Pfankuche, V.M.; Baumgärtner, W.; Puff, C. Viral oncolysis—Can insights from measles be transferred to canine distemper virus? Viruses 2014, 6, 2340–2375. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharjee, S.; Yadava, P.K. Measles virus: Background and oncolytic virotherapy. Biochem. Biophys. Rep. 2018, 13, 58–62. [Google Scholar] [CrossRef]
- Suter, S.E.; Chein, M.B.; von Messling, V.; Yip, B.; Cattaneo, R.; Vernau, W.; Madewell, B.R.; London, C.A. In vitro canine distemper virus infection of canine lymphoid cells: A prelude to oncolytic therapy for lymphoma. Clin. Cancer Res. 2005, 11, 1579–1587. [Google Scholar] [CrossRef] [Green Version]
- Del Puerto, H.L.; Martins, A.S.; Milsted, A.; Souza-Fagundes, E.M.; Braz, G.F.; Hissa, B.; Andrade, L.O.; Alves, F.; Rajao, D.S.; Leite, R.C.; et al. Canine distemper virus induces apoptosis in cervical tumor derived cell lines. Virol. J. 2011, 8, 334. [Google Scholar] [CrossRef] [Green Version]
- Puff, C.; Krudewig, C.; Imbschweiler, I.; Baumgärtner, W.; Alldinger, S. Influence of persistent canine distemper virus infection on expression of RECK, matrix-metalloproteinases and their inhibitors in a canine macrophage/monocytic tumour cell line (DH82). Vet. J. 2009, 182, 100–107. [Google Scholar] [CrossRef]
- Armando, F.; Fayyad, A.; Arms, S.; Barthel, Y.; Schaudien, D.; Rohn, K.; Gambini, M.; Lombardo, M.S.; Beineke, A.; Baumgärtner, W.; et al. Intratumoral canine distemper virus infection inhibits tumor growth by modulation of the tumor microenvironment in a murine xenograft model of canine histiocytic sarcoma. Int. J. Mol. Sci. 2021, 22, 3578. [Google Scholar] [CrossRef]
- Armando, F.; Gambini, M.; Corradi, A.; Becker, K.; Marek, K.; Pfankuche, V.M.; Mergani, A.E.; Brogden, G.; de Buhr, N.; von Köckritz-Blickwede, M.; et al. Mesenchymal to epithelial transition driven by canine distemper virus infection of canine histiocytic sarcoma cells contributes to a reduced cell motility in vitro. J. Cell Mol. Med. 2020, 24, 9332–9348. [Google Scholar] [CrossRef] [PubMed]
- Armando, F.; Gambini, M.; Corradi, A.; Giudice, C.; Pfankuche, V.M.; Brogden, G.; Attig, F.; von Köckritz-Blickwede, M.; Baumgärtner, W.; Puff, C. Oxidative stress in canine histiocytic sarcoma cells induced by an infection with canine distemper virus led to a dysregulation of HIF-1alpha downstream pathway resulting in a reduced expression of VEGF-B in vitro. Viruses 2020, 12, 200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfankuche, V.M.; Sayed-Ahmed, M.; Contioso, V.B.; Spitzbarth, I.; Rohn, K.; Ulrich, R.; Deschl, U.; Kalkuhl, A.; Baumgärtner, W.; Puff, C. Persistent morbillivirus infection leads to altered cortactin distribution in histiocytic sarcoma cells with decreased cellular migration capacity. PLoS ONE 2016, 11, e0167517. [Google Scholar] [CrossRef]
- Pfankuche, V.M.; Spitzbarth, I.; Lapp, S.; Ulrich, R.; Deschl, U.; Kalkuhl, A.; Baumgärtner, W.; Puff, C. Reduced angiogenic gene expression in morbillivirus-triggered oncolysis in a translational model for histiocytic sarcoma. J. Cell Mol. Med. 2017, 21, 816–830. [Google Scholar] [CrossRef]
- Amatya, B.; Lee, H.; Asico, L.D.; Konkalmatt, P.; Armando, I.; Felder, R.A.; Jose, P.A. SNX-PXA-RGS-PXC subfamily of SNXs in the regulation of receptor-mediated signaling and membrane trafficking. Int. J. Mol. Sci. 2021, 22, 2319. [Google Scholar] [CrossRef]
- Nishida, N.; Yano, H.; Nishida, T.; Kamura, T.; Kojiro, M. Angiogenesis in cancer. Vasc. Health Risk Manag. 2006, 2, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Pike, S.E.; Yao, L.; Jones, K.D.; Cherney, B.; Appella, E.; Sakaguchi, K.; Nakhasi, H.; Teruya-Feldstein, J.; Wirth, P.; Gupta, G.; et al. Vasostatin, a calreticulin fragment, inhibits angiogenesis and suppresses tumor growth. J. Exp. Med. 1998, 188, 2349–2356. [Google Scholar] [CrossRef]
- Cai, K.X.; Tse, L.Y.; Leung, C.; Tam, P.K.; Xu, R.; Sham, M.H. Suppression of lung tumor growth and metastasis in mice by adeno-associated virus-mediated expression of vasostatin. Clin. Cancer Res. 2008, 14, 939–949. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Yuan, Y.Z.; Lu, J.; Xia, L.; Zhu, Y.; Zhang, Y.P.; Qiao, M.M. Treatment of pancreatic carcinoma by adenoviral mediated gene transfer of vasostatin in mice. Gut 2006, 55, 259–265. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, F.; Tsagozis, P.; Lundberg, K.; Parsa, R.; Mangsbo, S.M.; Persson, M.A.; Harris, R.A.; Pisa, P. Tumor-specific bacteriophages induce tumor destruction through activation of tumor-associated macrophages. J. Immunol. 2009, 182, 3105–3111. [Google Scholar] [CrossRef] [Green Version]
- Boutilier, A.J.; Elsawa, S.F. Macrophage polarization states in the tumor microenvironment. Int. J. Mol. Sci. 2021, 22, 6995. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Marchesi, F.; Malesci, A.; Laghi, L.; Allavena, P. Tumour-associated macrophages as treatment targets in oncology. Nat. Rev. Clin. Oncol. 2017, 14, 399–416. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Wang, X.; Cheng, P. Remodeling of tumor immune microenvironment by oncolytic viruses. Front. Oncol. 2020, 10, 561372. [Google Scholar] [CrossRef]
- Sica, A.; Schioppa, T.; Mantovani, A.; Allavena, P. Tumour-associated macrophages are a distinct M2 polarised population promoting tumour progression: Potential targets of anti-cancer therapy. Eur. J. Cancer 2006, 42, 717–727. [Google Scholar] [CrossRef] [PubMed]
- Schmid, M.C.; Varner, J.A. Myeloid cells in the tumor microenvironment: Modulation of tumor angiogenesis and tumor inflammation. J. Oncol. 2010, 2010, 201026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howells, A.; Marelli, G.; Lemoine, N.R.; Wang, Y. Oncolytic viruses—Interaction of virus and tumor cells in the battle to eliminate cancer. Front. Oncol. 2017, 7, 195. [Google Scholar] [CrossRef] [Green Version]
- Grote, D.; Russell, S.J.; Cornu, T.I.; Cattaneo, R.; Vile, R.; Poland, G.A.; Fielding, A.K. Live attenuated measles virus induces regression of human lymphoma xenografts in immunodeficient mice. Blood 2001, 97, 3746–3754. [Google Scholar] [CrossRef] [Green Version]
- Grossardt, C.; Engeland, C.E.; Bossow, S.; Halama, N.; Zaoui, K.; Leber, M.F.; Springfeld, C.; Jaeger, D.; von Kalle, C.; Ungerechts, G. Granulocyte-macrophage colony-stimulating factor-armed oncolytic measles virus is an effective therapeutic cancer vaccine. Hum. Gene Ther. 2013, 24, 644–654. [Google Scholar] [CrossRef]
- Russell, L.; Peng, K.W. The emerging role of oncolytic virus therapy against cancer. Chin. Clin. Oncol. 2018, 7, 16. [Google Scholar] [CrossRef]
- De Silva, N.; Atkins, H.; Kirn, D.H.; Bell, J.C.; Breitbach, C.J. Double trouble for tumours: Exploiting the tumour microenvironment to enhance anticancer effect of oncolytic viruses. Cytokine Growth Factor Rev. 2010, 21, 135–141. [Google Scholar] [CrossRef]
- Hutzen, B.; Bid, H.K.; Houghton, P.J.; Pierson, C.R.; Powell, K.; Bratasz, A.; Raffel, C.; Studebaker, A.W. Treatment of medulloblastoma with oncolytic measles viruses expressing the angiogenesis inhibitors endostatin and angiostatin. BMC Cancer 2014, 14, 206. [Google Scholar] [CrossRef] [PubMed]
- Grote, D.; Cattaneo, R.; Fielding, A.K. Neutrophils contribute to the measles virus-induced antitumor effect: Enhancement by granulocyte macrophage colony-stimulating factor expression. Cancer Res. 2003, 63, 6463–6468. [Google Scholar] [PubMed]
- Heinrich, F.; Lehmbecker, A.; Raddatz, B.B.; Kegler, K.; Tipold, A.; Stein, V.M.; Kalkuhl, A.; Deschl, U.; Baumgärtner, W.; Ulrich, R.; et al. Morphologic, phenotypic, and transcriptomic characterization of classically and alternatively activated canine blood-derived macrophages in vitro. PLoS ONE 2017, 12, e0183572. [Google Scholar] [CrossRef] [PubMed]
- Padilla, L.; Martinez-Hernandez, J.; Barranco, I.; Lucas, X.; Pastor, L.M.; Rodriguez-Martinez, H.; Roca, J.; Parrilla, I. Granulocyte-macrophage colony stimulating factor (GM-CSF) is fully expressed in the genital tract, seminal plasma and spermatozoa of male pigs. Sci. Rep. 2020, 10, 13360. [Google Scholar] [CrossRef]
- Willemsen, A.; Zwart, M.P. On the stability of sequences inserted into viral genomes. Virus Evol. 2019, 5, vez045. [Google Scholar] [CrossRef]
- Janke, M.; Peeters, B.; de Leeuw, O.; Moorman, R.; Arnold, A.; Fournier, P.; Schirrmacher, V. Recombinant Newcastle disease virus (NDV) with inserted gene coding for GM-CSF as a new vector for cancer immunogene therapy. Gene Ther. 2007, 14, 1639–1649. [Google Scholar] [CrossRef]
- Koch, M.S.; Lawler, S.E.; Chiocca, E.A. HSV-1 Oncolytic viruses from bench to bedside: An overview of current clinical trials. Cancers 2020, 12, 3514. [Google Scholar] [CrossRef]
- Pol, J.; Kroemer, G.; Galluzzi, L. First oncolytic virus approved for melanoma immunotherapy. Oncoimmunology 2016, 5, e1115641. [Google Scholar] [CrossRef] [Green Version]
- Eubank, T.D.; Roberts, R.; Galloway, M.; Wang, Y.; Cohn, D.E.; Marsh, C.B. GM-CSF induces expression of soluble VEGF receptor-1 from human monocytes and inhibits angiogenesis in mice. Immunity 2004, 21, 831–842. [Google Scholar] [CrossRef] [Green Version]
- Pyonteck, S.M.; Akkari, L.; Schuhmacher, A.J.; Bowman, R.L.; Sevenich, L.; Quail, D.F.; Olson, O.C.; Quick, M.L.; Huse, J.T.; Teijeiro, V.; et al. CSF-1R inhibition alters macrophage polarization and blocks glioma progression. Nat. Med. 2013, 19, 1264–1272. [Google Scholar] [CrossRef] [Green Version]
- Marcelo, A.; Koppenol, R.; de Almeida, L.P.; Matos, C.A.; Nobrega, C. Stress granules, RNA-binding proteins and polyglutamine diseases: Too much aggregation? Cell Death Dis. 2021, 12, 592. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Na, Z.; Slavoff, S.A. P-Bodies: Composition, Properties, and Functions. Biochemistry 2018, 57, 2424–2431. [Google Scholar] [CrossRef] [PubMed]
- Bibi, A.; Agarwal, N.K.; Dihazi, G.H.; Eltoweissy, M.; Van Nguyen, P.; Mueller, G.A.; Dihazi, H. Calreticulin is crucial for calcium homeostasis mediated adaptation and survival of thick ascending limb of Henle’s loop cells under osmotic stress. Int. J. Biochem. Cell Biol. 2011, 43, 1187–1197. [Google Scholar] [CrossRef] [PubMed]
- Waisman, D.M.; Salimath, B.P.; Anderson, M.J. Isolation and characterization of CAB-63, a novel calcium-binding protein. J. Biol. Chem. 1985, 260, 1652–1660. [Google Scholar] [CrossRef]
- Greenwood, S.K.; Hill, R.B.; Sun, J.T.; Armstrong, M.J.; Johnson, T.E.; Gara, J.P.; Galloway, S.M. Population doubling: A simple and more accurate estimation of cell growth suppression in the in vitro assay for chromosomal aberrations that reduces irrelevant positive results. Environ. Mol. Mutagen. 2004, 43, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, N.; Gall, F.M.; Mathieu, C.; Hierweger, M.M.; Brugger, M.; Alves, M.P.; Vesin, J.; Banfi, D.; Kalbermatter, D.; Horvat, B.; et al. Highly potent host-specific small-molecule inhibitor of paramyxovirus and pneumovirus replication with high resistance barrier. mBio 2021, 12, e0262121. [Google Scholar] [CrossRef]
- Shrestha, N.; Gall, F.M.; Vesin, J.; Chambon, M.; Turcatti, G.; Fotiadis, D.; Riedl, R.; Plattet, P. Antiviral screen against canine distemper virus-induced membrane fusion activity. Viruses 2021, 13, 128. [Google Scholar] [CrossRef]
- Techangamsuwan, S.; Haas, L.; Rohn, K.; Baumgärtner, W.; Wewetzer, K. Distinct cell tropism of canine distemper virus strains to adult olfactory ensheathing cells and Schwann cells in vitro. Virus Res. 2009, 144, 195–201. [Google Scholar] [CrossRef]
- von Smolinski, D.; Leverkoehne, I.; von Samson-Himmelstjerna, G.; Gruber, A.D. Impact of formalin-fixation and paraffin-embedding on the ratio between mRNA copy numbers of differently expressed genes. Histochem. Cell Biol. 2005, 124, 177–188. [Google Scholar] [CrossRef]
- Krüger, N.; Rocha, C.; Runft, S.; Krüger, J.; Färber, I.; Armando, F.; Leitzen, E.; Brogden, G.; Gerold, G.; Pöhlmann, S.; et al. The upper respiratory tract of felids is highly susceptible to SARS-CoV-2 infection. Int. J. Mol. Sci. 2021, 22, 10636. [Google Scholar] [CrossRef]
- Armando, F.; Ferrari, L.; Arcari, M.L.; Azzali, G.; Dallatana, D.; Ferrari, M.; Lombardi, G.; Zanfabro, M.; Di Lecce, R.; Lunghi, P.; et al. Endocanalicular transendothelial crossing (ETC): A novel intravasation mode used by HEK-EBNA293-VEGF-D cells during the metastatic process in a xenograft model. PLoS ONE 2020, 15, e0239932. [Google Scholar] [CrossRef] [PubMed]
- Armando, F.; Pigoli, C.; Gambini, M.; Ghidelli, A.; Ghisleni, G.; Corradi, A.; Passeri, B.; Caniatti, M.; Grieco, V.; Baumgärtner, W.; et al. Peripheral nerve sheath tumors resembling human atypical neurofibroma in goldfish (Carassius auratus, Linnaeus, 1758). Animals 2021, 11, 2621. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Percentage of CDV-Infected Cells | Median (in %) | Range (in %) |
|---|---|---|
| Non-infected DH82 | 0 | 0 |
| DH82 CDV-Ond pi | 98.26 | 97.67–99.12 |
| DH82 CDV-Ondneon pi | 96.76 | 96.08–97.13 |
| DH82 CDV-Ondneon-vasostatin pi | 95.56 | 95.44–97.91 |
| DH82 CDV-Ondneon-GM-CSF pi | 97.88 | 97.30–98.00 |
| Percentage of mNeonGreen-Expressing Cells | Median (in %) | Range (in %) |
|---|---|---|
| Non-infected DH82 | 0 | 0 |
| DH82 CDV-Ond pi | 0 | 0 |
| DH82 CDV-Ondneon pi | 92.19 | 91.89–92.53 |
| DH82 CDV-Ondneon-vasostatin pi | 88.44 | 83.60–93.34 |
| DH82 CDV-Ondneon-GM-CSF pi | 90.63 | 89.32–94.90 |
| Number of GM-CSF mRNA Transcripts | Median (in % of GAPDH) | Range (in % of GAPDH) |
|---|---|---|
| Non-infected DH82 | 0.09 | 0.07–0.14 |
| DH82 CDV-Ond pi | 0.08 | 0.00–0.29 |
| DH82 CDV-Ondneon pi | 0.11 | 0.02–0.26 |
| DH82 CDV-Ondneon-GM-CSF pi | 3183.90 | 2422.74–5158.06 |
| Percentage of GM-CSF Expressing Cells | Median (in %) | Range (in %) |
|---|---|---|
| Non-infected DH82 | 2.41 | 1.82–2.70 |
| DH82 CDV-Ond pi | 3.20 | 2.93–5.38 |
| DH82 CDV-Ondneon pi | 5.43 | 5.18–5.43 |
| DH82 CDV-Ondneon-GM-CSF pi | 11.53 | 10.77–11.89 |
| Vasostatin mRNA Transcripts (in % of GAPDH) | Median (in %) | Range (in %) |
|---|---|---|
| Non-infected DH82 | 87.36 | 56.28–160.97 |
| DH82 CDV-Ond pi | 96.33 | 11.92–472 |
| DH82 CDV-Ondneon pi | 36.22 | 17.55–137.98 |
| DH82 CDV-Ondneon-vasostatin pi | 1688.40 | 923.58–11,081.08 |
| Genes | Primer Sequences (5′-3′) | Length of Amplicon (bp) | Position | GenBank Accession Number | |
|---|---|---|---|---|---|
| GAPDH | Forward * | AAGGTCGGAGTCAACGGATT | 365 | 7–26 | AB038240 [9] |
| Reverse * | GCAGAAGAAGCAGAGATGATG | 371–351 | |||
| Forward | GTCATCAACGGGAAGTCCATCTC | 84 | 96–218 | ||
| Reverse | AACATACTCAGCACCAGCATCAC | 279–257 | |||
| CDV | Forward * | ACAGGATTGCTGAGGACCTAT | 287 | 769–789 | AF378705 [9,49] |
| Reverse * | CAAGATAACCATGTACGGTGC | 1055–1035 | |||
| Forward | GCTCTTGGGTTGCATGAGTT | 83 | 954–973 | ||
| Reverse | GCTGTTTCACCCATCTGTTG | 1036–1017 | |||
| Vasostatin | Forward * | TGTTCTCAGTTCCGGCAAGT | 235 | 83–103 | XM038428450.1 |
| Reverse * | ACATGCACGGAGACTCTGAG | 317–298 | |||
| Forward | TGACCAGGAGAAGGATAAAGGG | 100 | 110–132 | ||
| Reverse | GCAACAAAGGCCAGACACTG | 209–189 | |||
| GM-CSF | Forward * | CTCACCCACCCTTGTCACTC | 218 | 63–83 | NM001003245.1 |
| Reverse * | CACCAGCCTCAAGAATCCCT | 280–260 | |||
| Forward | TTGTCACTCGGCCCTCTCA | 93 | 74–93 | ||
| Reverse | TCATCACAGCAGTCACGTCA | 160–140 | |||
| Primary Antibody | Host Species, Clonality | Antigen Retrieval | Blocking Serum | Dilution | Secondary Antibody (1:200) |
|---|---|---|---|---|---|
| CDV-NP (University of Bern, Prof. Zurbriggen) | Mouse, monoclonal, clone D110 | na | Goat serum | 1:100 (IF) | GaM-Cy3 |
| GM-CSF (R&D Systems, Minneapolis, MN, USA) | Goat, polyclonal | na | Horse serum | 1:80 (IF) | DaG-Cy3 |
| N-terminal calreticulin (Sigma-Aldrich, Taufkirchen, Germany) | Rabbit, polyclonal | Citrate buffer, MW (800 W, 20′) | Goat serum | 1:100 (IHC) | GaR-b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marek, K.; Armando, F.; Nippold, V.M.; Rohn, K.; Plattet, P.; Brogden, G.; Gerold, G.; Baumgärtner, W.; Puff, C. Persistent Infection of a Canine Histiocytic Sarcoma Cell Line with Attenuated Canine Distemper Virus Expressing Vasostatin or Granulocyte-Macrophage Colony-Stimulating Factor. Int. J. Mol. Sci. 2022, 23, 6156. https://doi.org/10.3390/ijms23116156
Marek K, Armando F, Nippold VM, Rohn K, Plattet P, Brogden G, Gerold G, Baumgärtner W, Puff C. Persistent Infection of a Canine Histiocytic Sarcoma Cell Line with Attenuated Canine Distemper Virus Expressing Vasostatin or Granulocyte-Macrophage Colony-Stimulating Factor. International Journal of Molecular Sciences. 2022; 23(11):6156. https://doi.org/10.3390/ijms23116156
Chicago/Turabian StyleMarek, Katarzyna, Federico Armando, Vanessa Maria Nippold, Karl Rohn, Philippe Plattet, Graham Brogden, Gisa Gerold, Wolfgang Baumgärtner, and Christina Puff. 2022. "Persistent Infection of a Canine Histiocytic Sarcoma Cell Line with Attenuated Canine Distemper Virus Expressing Vasostatin or Granulocyte-Macrophage Colony-Stimulating Factor" International Journal of Molecular Sciences 23, no. 11: 6156. https://doi.org/10.3390/ijms23116156
APA StyleMarek, K., Armando, F., Nippold, V. M., Rohn, K., Plattet, P., Brogden, G., Gerold, G., Baumgärtner, W., & Puff, C. (2022). Persistent Infection of a Canine Histiocytic Sarcoma Cell Line with Attenuated Canine Distemper Virus Expressing Vasostatin or Granulocyte-Macrophage Colony-Stimulating Factor. International Journal of Molecular Sciences, 23(11), 6156. https://doi.org/10.3390/ijms23116156