
Transcriptome Analysis Reveals Crosstalk between the Abscisic Acid and Jasmonic Acid Signaling Pathways in Rice-Mediated Defense against Nilaparvata lugens


Abstract
:1. Introduction
2. Results
2.1. RNA-seq Assembly and Mapping
2.2. Functional Classification, Annotation and Quality Assessment of DEGs
2.3. Analysis of KEGG Metabolic Pathways
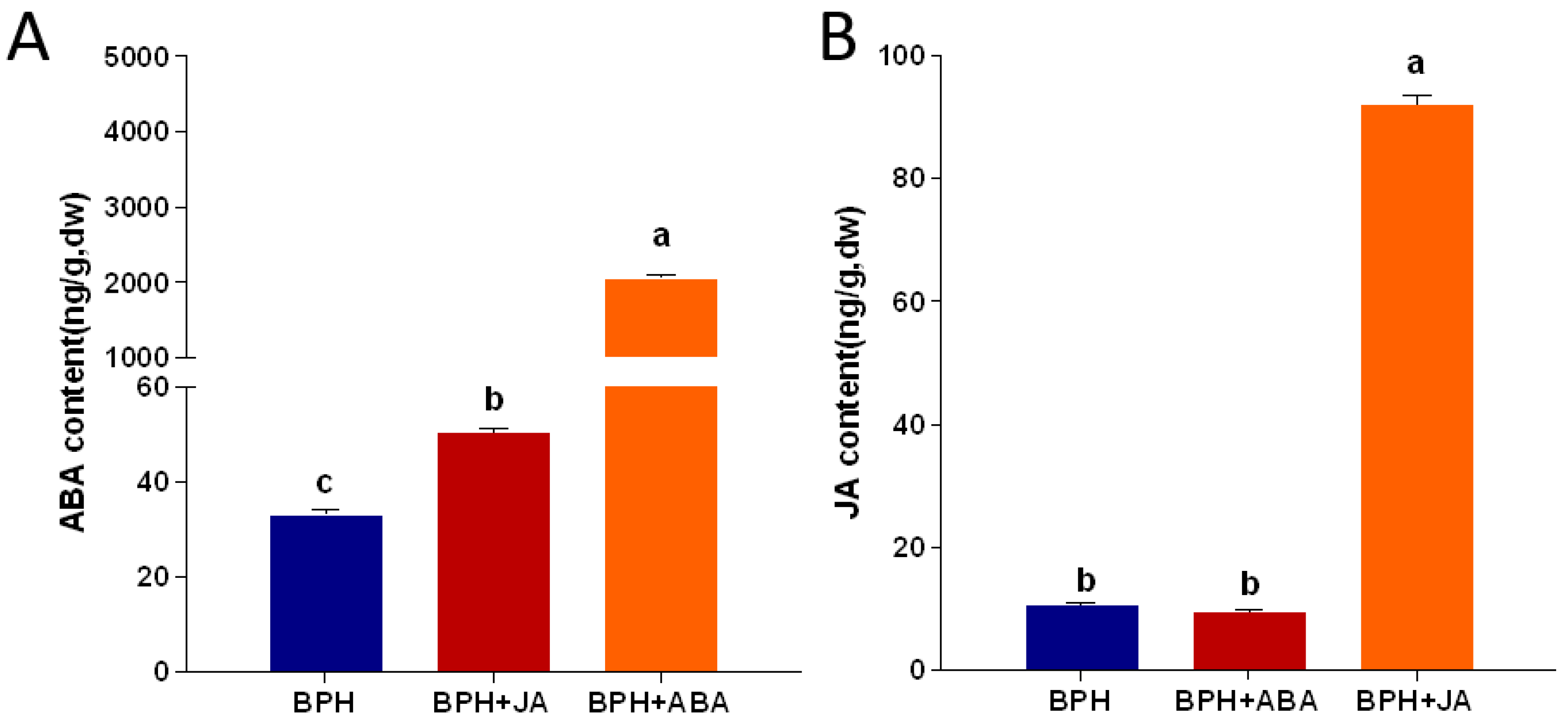
2.4. Changes in JA and ABA Hormone Content and Gene Expression after BPH Infestation
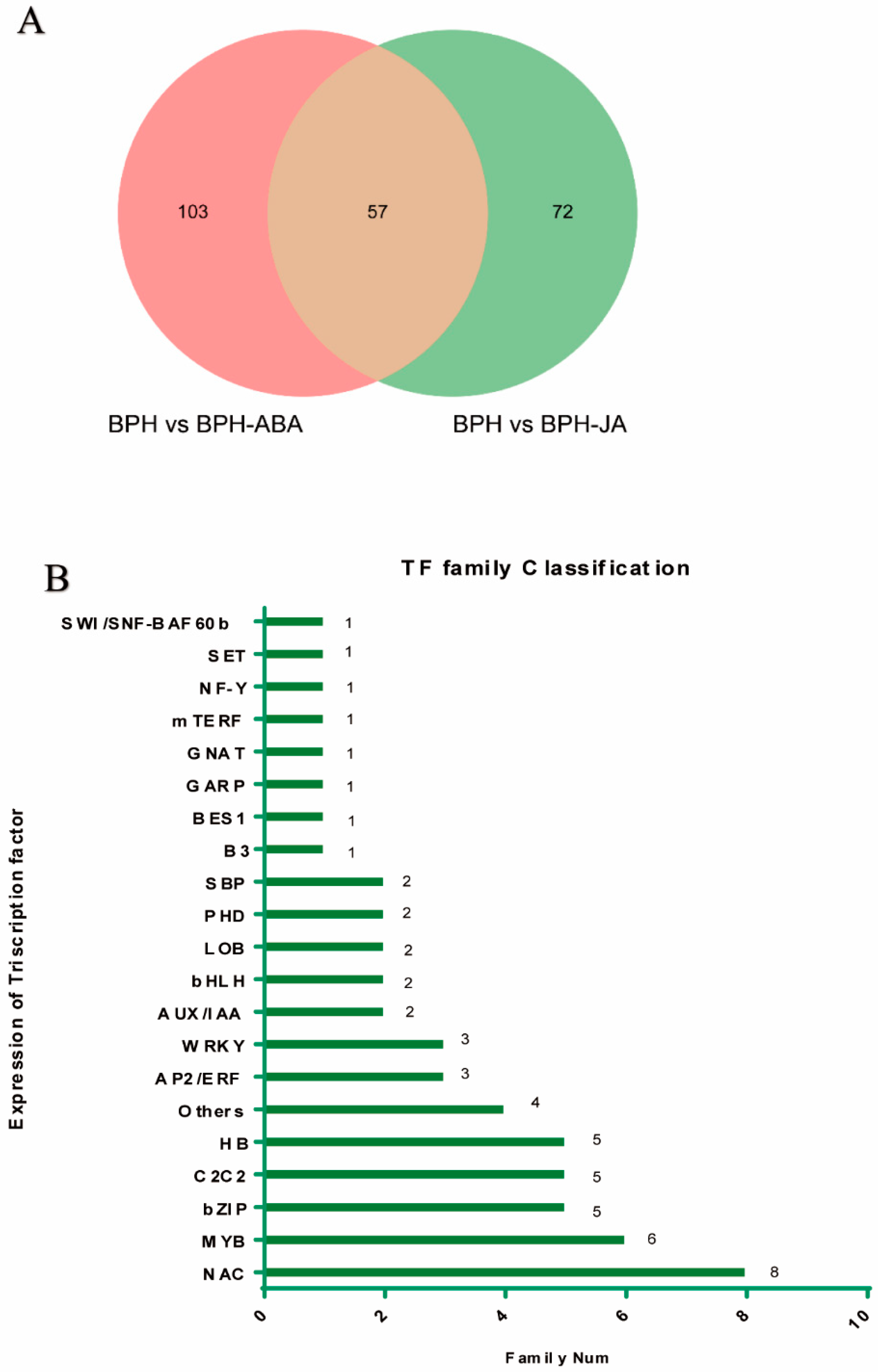
2.5. Expression of Rice Transcription Factors in Response to Hormone Treatments
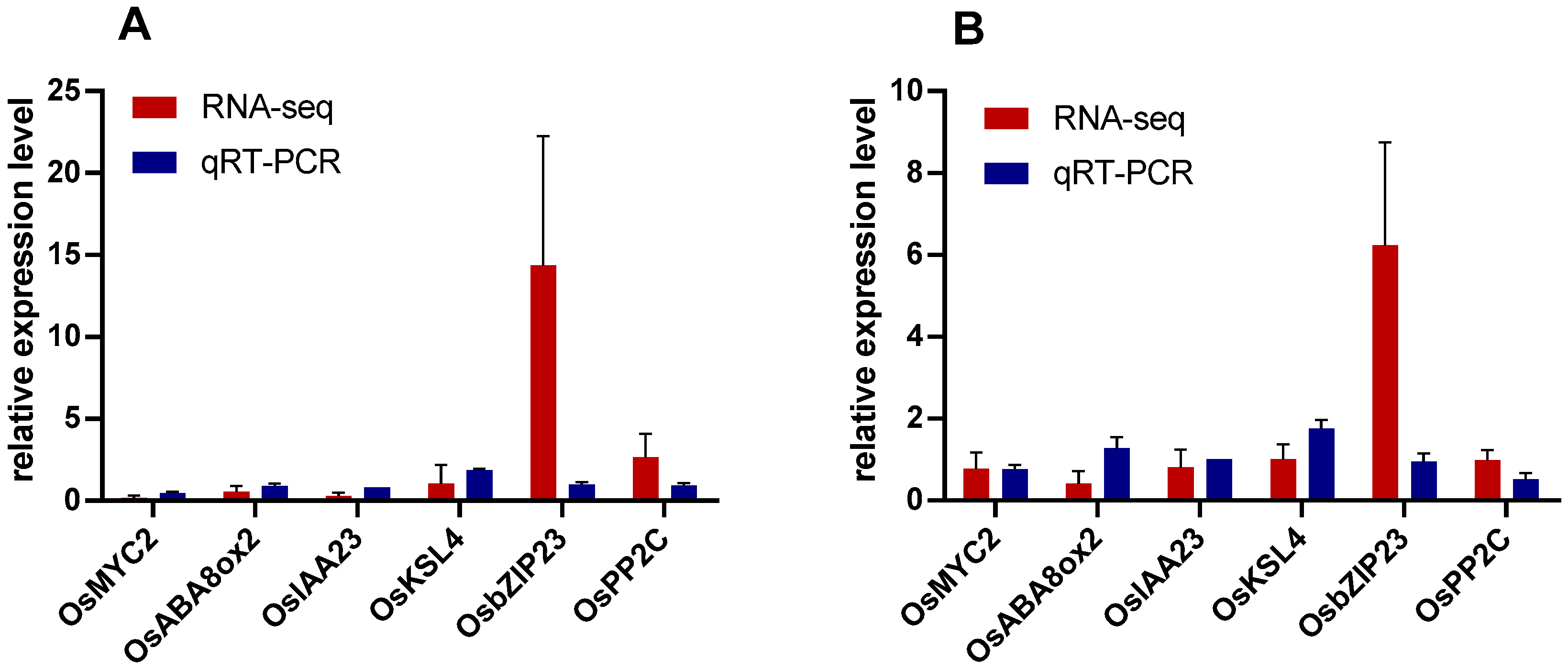
2.6. Quantitative RT-PCR
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. ABA, JA and BPH Treatments
4.3. RNA Library Construction, Sequencing and Quality Control
4.4. RNA Sequencing
4.5. Quantification of Phytohormones
4.6. Identification of DEGs and Verification by qRT-PCR
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Skamnioti, P.; Gurr, S.J. Against the grain: Safeguarding rice from rice blast disease. Trends Biotechnol. 2009, 27, 141–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, M.P.; Bari, M.N.; Haque, S.S.; Kabir, M.M.M.; Afrin, S.; Nowrin, F.; Islam, M.S.; Landis, D.A. Establishing next-generation pest control services in rice fields: Eco-agriculture. Sci. Rep. 2019, 9, 10180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heong, K.L.; Wong, L.; Reyes, J.H.D. Addressing Planthopper Threats to Asian Rice Farming and Food Security: Fixing Insecticide Misuse; Asian Development Bank: Mandaluyong, Philippines, 2013; pp. 67–76. [Google Scholar]
- Almeida, D.M.; Almadanim, M.C.; Lourenço, T.; Abreu, I.A.; Saibo, N.J.; Oliveira, M.M. Screening for abiotic stress tolerance in rice: Salt, cold, and drought. Methods Mol. Biol. 2016, 1398, 155–182. [Google Scholar]
- Wang, Y.J.; Huang, J.K.; Wang, J.X.; Christopher, F. Mitigating rice production risks from drought through improving irrigation infrastructure and management in China. Agric. Resour. Econ. 2018, 62, 161–176. [Google Scholar] [CrossRef]
- Cheng, J.A. Rice planthopper problems and relevant causes in China. In Planthoppers: New Threats to the Sustainability of Intensive Rice Production Systems in Asia; International Rice Research Institute: Los Baños, Philippines, 2009; pp. 157–178. [Google Scholar]
- Hogenhout, S.A.; Ammar, D.; Whitfield, A.E.; Redinbaugh, M.G. Insect vector interactions with persistently transmitted viruses. Annu. Rev. Phytopathol. 2008, 46, 327–359. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.H.; Liu, X.G.; Zhu, Y.C.; Inzé, D.; Van Camp, W. Inheritance mode and realized heritability of resistance to imidacloprid in the brown planthopper, Nilaparvata lugens (Stål) (Homoptera: Delphacidae). Pest Manag. Sci. 2009, 65, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.D.; Verdeprado, H.; Zita, D.; Sanada-Morimura, S. The development and characterization of near-isogenic and pyramided lines carrying resistance genes to brown planthopper with the genetic background of Japonica rice (Oryza sativa L.). Plants 2019, 8, 498. [Google Scholar] [CrossRef] [Green Version]
- Chanclud, E.; Lacombe, B. Plant hormones: Key players in gut microbiota and human diseases? Trends Plant Sci. 2017, 22, 754–758. [Google Scholar] [CrossRef]
- Nakashima, K.; Yamaguchi-Shinozaki, K. ABA signaling in stress-response and seed development. Plant Cell Rep. 2013, 32, 959–970. [Google Scholar] [CrossRef]
- Hazman, M.; Sühnel, M.; Schäfer, S.; Zumsteg, J. Characterization of jasmonoyl-isoleucine (JA-Ile) hormonal catabolic pathways in rice upon wounding and salt stress. Rice 2019, 12, 45. [Google Scholar] [CrossRef]
- Liu, J.L.; Chen, X.; Zhang, H.M.; Yang, X.; Wong, A. Effects of exogenous plant growth regulator abscisic acid-induced resistance in rice on the expression of vitellogenin mRNA in Nilaparvata lugens (Hemiptera: Delphacidae) adult females. J. Insect Sci. 2014, 14, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.L.; Du, H.T.; Ding, X.; Zhou, Y.; Xie, P.; Wu, J. Mechanisms of callose deposition in rice regulated by exogenous abscisic acid and its involvement in rice resistance to Nilaparvata lugens Stål (Hemiptera: Delphacidae). Pest Manag. Sci. 2017, 73, 2559–2568. [Google Scholar] [CrossRef]
- Adie, B.A.T.; Pérez-Pérez, J.; Pérez-Pérez, M.M.; Godoy, M.; Sánchez-Serrano, J.J.; Schmelz, E.A.; Solano, R. ABA is an essential signal for plant resistance to pathogens affecting JA biosynthesis and the activation of defenses in Arabidopsis. Plant Cell 2007, 19, 1665–1681. [Google Scholar] [CrossRef] [Green Version]
- Dinh, S.T.; Baldwin, I.T.; Galis, I. The HERBIVORE ELICITOR-REGULATED1 gene enhances abscisic acid levels and defenses against herbivores in Nicotiana attenuata plants. Plant Physiol. 2013, 162, 2106–2124. [Google Scholar] [CrossRef] [Green Version]
- Vos, I.A.; Adriaan, V.; Schuurink, R.C.; Watt, L.G.; Pieterse, C.M.; Van Wees, S.C. Onset of herbivore-induced resistance in systemic tissue primed for jasmonate-dependent defenses is activated by abscisic acid. Front. Plant Sci. 2013, 4, 539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aleman, F.; Yazaki, J.; Lee, M.; Takahashi, Y.; Kim, A.Y.; Li, Z.; Kinoshita, T.; Ecker, J.R.; Schroeder, J.I. An ABA-increased interaction of the PYL6 ABA receptor with MYC2 transcription factor: A putative link of ABA and JA signaling. Sci. Rep. 2016, 6, 28941. [Google Scholar] [CrossRef]
- Nahar, K.; Kyndt, T.; Nzogela, Y.B.; Gheysen, G. Abscisic acid interacts antagonistically with classical defense pathways in rice-migratory nematode interaction. New Phytol. 2012, 196, 901–913. [Google Scholar] [CrossRef] [PubMed]
- Kyndt, T.; Nahar, K.; Haeck, A.; Verbeek, R.; Demeestere, K.; Gheysen, G. Interplay between carotenoids, abscisic acid and jasmonate guides the compatible rice-meloidogyne graminicola interaction. Front. Plant Sci. 2017, 8, 951. [Google Scholar] [CrossRef]
- Howe, G.A.; Jander, G. Plant immunity to insect herbivores. Annu. Rev. Plant Biol. 2008, 59, 41–66. [Google Scholar] [CrossRef] [Green Version]
- Erb, M.; Meldau, S.; Howe, G.A. Role of phytohormones in insect-specific plant reactions. Trends Plant Sci. 2012, 17, 250–259. [Google Scholar] [CrossRef] [Green Version]
- Bodenhausen, N.; Reymond, P. Signaling pathways controlling induced resistance to insect herbivores in Arabidopsis. Mol. Plant-Microbe Interact. 2007, 20, 1406–1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, B.; Zhang, W.; Liu, B.; Hu, J.; Wei, Z.; Shi, Z.; He, R.; Zhu, L.; Chen, R.; Han, B.; et al. Identification and characterization of BPH14, a gene conferring resistance to brown planthopper in rice. Proc. Natl. Acad. Sci. USA 2009, 106, 22163–22168. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Guo, H.; Yuan, L.; Wei, J.; Zhang, W.; Ge, F. Plant stomatal closure improves aphid feeding under elevated CO2. Glob. Change Biol. 2015, 21, 2739–2748. [Google Scholar] [CrossRef] [PubMed]
- Hillwig, M.S.; Chiozza, M.; Casteel, C.L.; Lau, S.T.; Hohenstein, J.; Hernández, E.; Jander, G.; MacIntosh, G.C. Abscisic acid deficiency increases defense responses against Myzus persicae in Arabidopsis. Mol. Plant Pathol. 2016, 17, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Zong, W.; Tang, N.; Yang, J.; Peng, L.; Ma, S.; Xu, Y.; Li, G.; Xiong, L. Feedback regulation of ABA signaling and biosynthesis by a bZIP transcription factor targets drought-resistance-related genes. Plant Physiol. 2016, 171, 2810–2825. [Google Scholar] [CrossRef] [Green Version]
- Vranová, E.; Langebartels, C.; Van, M.M.; Inzé, D.; Van Camp, W. Oxidative stress, heat shock and drought differentially affect expression of a tobacco protein phosphatase 2C. J. Exp. Bot. 2000, 51, 1763–1764. [Google Scholar] [CrossRef] [Green Version]
- Schweighofer, A.; Hirt, H.; Meskiene, I. Plant PP2C phosphatases: Emerging functions in stress signaling. Trends Plant Sci. 2004, 9, 236–243. [Google Scholar] [CrossRef]
- Gonzalez, A.; Zhao, M.; Leavitt, J.M.; Lloyd, A.M. Regulation of the anthocyanin biosynthetic pathway by the TTG1/bHLH/Myb transcriptional complex in Arabidopsis seedlings. Plant J. 2008, 53, 14. [Google Scholar] [CrossRef]
- Purva, B.; Chetna, S.; Anshu, A.; Mehrotra, S.; Mehrotra, R. PP2C-like promoter and its deletion variants are induced by ABA but not by MeJA and SA in Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 547. [Google Scholar]
- Millar, A.A.; Gubler, F. The Arabidopsis GAMYB-like genes, MYB33 and MYB65, are microRNA-regulated genes that redundantly facilitate anther development. Plant Cell 2005, 17, 705–721. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, A.; Mendenhall, J.; Huo, Y.J.; Lloyd, A. TTG1 complex MYBs, MYB5 and TT2, control outer seed coat differentiation. Dev. Biol. 2009, 325, 412–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Gao, L.; Wang, H.; Chen, X.; Wang, Y.; Yang, H.; Wei, C.; Wan, X.; Xia, T. The R2R3-MYB, bHLH, WD40, and related transcription factors in flavonoid biosynthesis. Funct. Integr. Genom. 2013, 13, 75–98. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhang, J.; Li, J.; Zhou, G.; Zhou, G.; Wang, Q.; Bian, W.; Erb, M.; Lou, Y. Prioritizing plant defence overgrowth through WRKY regulation facilitates infestation by non-target herbivores. eLife 2015, 4, e04805. [Google Scholar] [CrossRef] [Green Version]
- Huangfu, J.; Li, J.; Li, R.; Ye, M.; Kuai, P.; Zhang, T.; Lou, Y. The transcription factor OsWRKY45 negatively modulates the resistance of rice to the brown planthopper Nilaparvata lugens. Int. J. Mol. Sci. 2016, 17, 697. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.; Wang, L.; Zhao, J.; Li, C.; Ge, F.; Kang, L. Ecological trade-offs between jasmonic acid-dependent direct and indirect plant defences in tritrophic interaction. New Phytol. 2011, 189, 557–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shangguan, X.; Zhang, J.; Liu, B.; Zhao, Y.; Wang, H.; Wang, Z.; Guo, J.; Rao, W.; Jing, S.; Guan, W. A mucin-like protein of planthopper is required for feeding and induces immunity response in plants. Plant Physiol. 2018, 176, 552–565. [Google Scholar] [CrossRef] [Green Version]
- Qi, J.; Li, J.; Han, X.; Li, R.; Wu, J.; Yu, H.; Hu, L.; Xiao, Y.; Lu, J.; Lou, Y. Jasmonic acid carboxyl methyltransferase regulates development and herbivory-induced defense response in rice. J. Integr. Plant Biol. 2016, 58, 564–576. [Google Scholar] [CrossRef]
- Lou, Y.G.; Du, M.H.; Turlings, T.C.; Cheng, J.A.; Shan, W.F. Exogenous application of jasmonic acid induces volatile emissions in rice and enhances parasitism of Nilaparvata lugens eggs by the parasitoid Anagrus nilaparvatae. J. Chem. Ecol. 2005, 31, 1985–2002. [Google Scholar] [CrossRef]
- Shi, X.; Pan, H.; Xie, W.; Wang, S.; Wu, Q.; Chen, G.; Tian, L.; Zhou, X.; Liu, Y.; Zhang, Y. Different effects of exogenous jasmonic acid on preference and performance of viruliferous Bemisia tabaci B and Q. Entomol. Exp. Appl. 2017, 165, 148–158. [Google Scholar] [CrossRef]
- Ma, F.; Yang, X.; Shi, Z.; Miao, X. Novel crosstalk between ethylene- and jasmonic acid-pathway responses to a piercing-sucking insect in rice. New Phytol. 2020, 225, 474–487. [Google Scholar] [CrossRef] [Green Version]
- Lackman, P.; Goossens, A. Jasmonate signaling involves the abscisic acid receptor PYL4 to regulate metabolic reprogramming in Arabidopsis and tobacco. Proc. Natl. Acad. Sci. USA 2011, 108, 5891–5896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenzo, O.; Chico, J.M.; Sánchez-Serrano, J.J.; Solano, R. JASMONATE-INSENSITIVE1 encodes a MYC transcription factor essential to discriminate between different jasmonate-regulated defense responses in Arabidopsis. Plant Cell 2004, 16, 1938–1950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abe, H.; Urao, T.T.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as transcriptional activators in abscisic acid signaling. Plant Cell 2003, 15, 63–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerchev, P.I.; Foyer, C.H. The transcription factor ABI4 is required for the ascorbic acid-dependent regulation of growth and regulation of jasmonate-dependent defense signaling pathways in Arabidopsis. Plant Cell 2011, 23, 3319–3334. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Sun, Y.; Peng, X.; Wang, Q.; Harris, M.; Ge, F. Up-regulation of abscisic acid signaling pathway facilitates aphid xylem absorption and osmoregulation under drought stress. J. Exp. Bot. 2016, 67, 681–693. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, D.; D’Agostino, N.; Tytgat, T.O.; Sun, P.; Lortzing, T.; Visser, E.J.; Cristescu, S.M.; Steppuhn, A.; Mariani, C.; van Dam, N.M.; et al. Drought and flooding have distinct effects on herbivore-induced responses and resistance in Solanum dulcamara. Plant Cell Environ. 2016, 39, 1485–1499. [Google Scholar] [CrossRef] [Green Version]
- Gene Ontology Consortiom; Blake, J.A.; Dolan, M.; Drabkin, H.; Hill, D.P.; Li, N.; Sitnikov, D.; Bridges, S.; Burgess, S.; Buza, T.; et al. Gene Ontology annotations and resources. Nucleic Acids Res. 2013, 41, D530–D535. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Read Number | Base Number | % ≥Q30 | Reads Aligned | Exonic |
|---|---|---|---|---|---|
| BPH1 | 29004617 | 8701385100 | 92.74 | 51063904 (88.03%) | 35,168,954 (82.64%) |
| BPH2 | 23242201 | 6972660300 | 92.24 | 37905421 (81.54%) | 22,739,057 (79.34%) |
| BPH3 | 26550424 | 7965127200 | 92.54 | 45041229 (84.82%) | 33,151,024 (84.38%) |
| BPH + ABA1 | 26551481 | 7965444300 | 92.38 | 48669904 (91.65%) | 36,079,719 (84.7%) |
| BPH + ABA2 | 29957564 | 8987269200 | 94.73 | 53778358 (89.76%) | 36,055,653 (83.67%) |
| BPH + ABA3 | 36308246 | 10892473800 | 95.25 | 60138179 (82.82%) | 37,540,070 (81.7%) |
| BPH + JA1 | 27950890 | 8385267000 | 95.37 | 51810937 (92.68%) | 38,656,560 (86.07%) |
| BPH + JA2 | 28574794 | 8572438200 | 93.63 | 54632427 (95.60%) | 42,975,066 (86.27%) |
| BPH + JA3 | 32635686 | 9790705800 | 95.31 | 61075150 (93.57%) | 44,054,381 (85.55%) |
| Treatment | All DEGs | Up-Regulated DEGs | Down-Regulated DEGs |
|---|---|---|---|
| BPH vs. BPH + ABA | 3491 | 1795 | 1696 |
| BPH vs. BPH + JA | 2727 | 1674 | 1053 |
| Pathway ID | Gene Name | Functional Annotation | p-Value | log2FC | |
|---|---|---|---|---|---|
| LOC_Os02g10120 | K00454 | OsLOX1 | Lipoxygenase | 8.38 × 10−12 | −3.58132 |
| LOC_Os03g12500 | K01723 | OsAOS2 | Oxidoreductase activity | 2.94 × 10−9 | 3.099447 |
| LOC_Os08g35740 | K05894 | OsOPR | 12-oxophytodienoic acid reductase | 7.45 × 10−5 | 2.655998 |
| LOC_Os05g07090 | K00232 | OsACOX1 | Acyl-CoA oxidase | 6.23 × 10−5 | 2.050977 |
| LOC_Os06g24704 | K00232 | OsACOX3 | Acyl-CoA oxidase | 7.72 × 10−9 | 1.708204 |
| LOC_Os02g57260 | K07513 | OsACAA1 | Acetyl-CoA acyltransferase 1 | 7.89 × 10−10 | 1.422267 |
| LOC_Os03g08320 | K13464 | OsJAZ11 | Jasmonate ZIM domain-containing protein | 5.65 × 10−3 | 1.686453 |
| LOC_Os03g28940 | K13464 | OsJAZ6 | Jasmonate ZIM domain-containing protein | 3.51 × 10−4 | 1.574457 |
| LOC_Os07g42370 | K13464 | OsJAZ7 | Jasmonate ZIM domain-containing protein | 3.37 × 10−5 | 1.789341 |
| LOC_Os09g26780 | K13464 | OsJAZ8 | Jasmonate ZIM domain-containing protein | 2.33 × 10−3 | 2.0051 |
| LOC_Os10g25290 | K13464 | OsJAZ12 | Jasmonate ZIM domain-containing protein | 2.49 × 10−7 | 2.292083 |
| FC, fold change |
| Pathway ID | Gene Name | Functional Annotation | p-Value | log2FC | |
|---|---|---|---|---|---|
| LOC_Os06g36670 | K14496 | OsPYL9; OsPYL/RCAR9 | Abscisic acid receptor, PYR/PYL family | 7.97 × 10−4 | −1.63928 |
| LOC_Os02g47510 | K09840 | OsNCED1 | 9-cis-epoxycarotenoid dioxygenase | 3.01 × 10−4 | 1.875411 |
| LOC_Os01g46760 | K14497 | PP2C | Protein serine/threonine phosphatase activity | 4.4 × 10−3 | 1.37321 |
| LOC_Os03g27280 | K14498 | OsSAPK1 | Serine/threonine-protein kinase SRK2 | 4.40 × 10−5 | −1.33978 |
| FC, fold change |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Chen, L.; Ding, X.; Fan, W.; Liu, J. Transcriptome Analysis Reveals Crosstalk between the Abscisic Acid and Jasmonic Acid Signaling Pathways in Rice-Mediated Defense against Nilaparvata lugens. Int. J. Mol. Sci. 2022, 23, 6319. https://doi.org/10.3390/ijms23116319
Li J, Chen L, Ding X, Fan W, Liu J. Transcriptome Analysis Reveals Crosstalk between the Abscisic Acid and Jasmonic Acid Signaling Pathways in Rice-Mediated Defense against Nilaparvata lugens. International Journal of Molecular Sciences. 2022; 23(11):6319. https://doi.org/10.3390/ijms23116319
Chicago/Turabian StyleLi, Jitong, Lin Chen, Xu Ding, Wenyan Fan, and Jinglan Liu. 2022. "Transcriptome Analysis Reveals Crosstalk between the Abscisic Acid and Jasmonic Acid Signaling Pathways in Rice-Mediated Defense against Nilaparvata lugens" International Journal of Molecular Sciences 23, no. 11: 6319. https://doi.org/10.3390/ijms23116319
APA StyleLi, J., Chen, L., Ding, X., Fan, W., & Liu, J. (2022). Transcriptome Analysis Reveals Crosstalk between the Abscisic Acid and Jasmonic Acid Signaling Pathways in Rice-Mediated Defense against Nilaparvata lugens. International Journal of Molecular Sciences, 23(11), 6319. https://doi.org/10.3390/ijms23116319