
The Consequences of Mitochondrial T10432C Mutation in Cika Cattle: A “Potential” Model for Leber’s Hereditary Optic Neuropathy

 , , ,
, , , 
 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
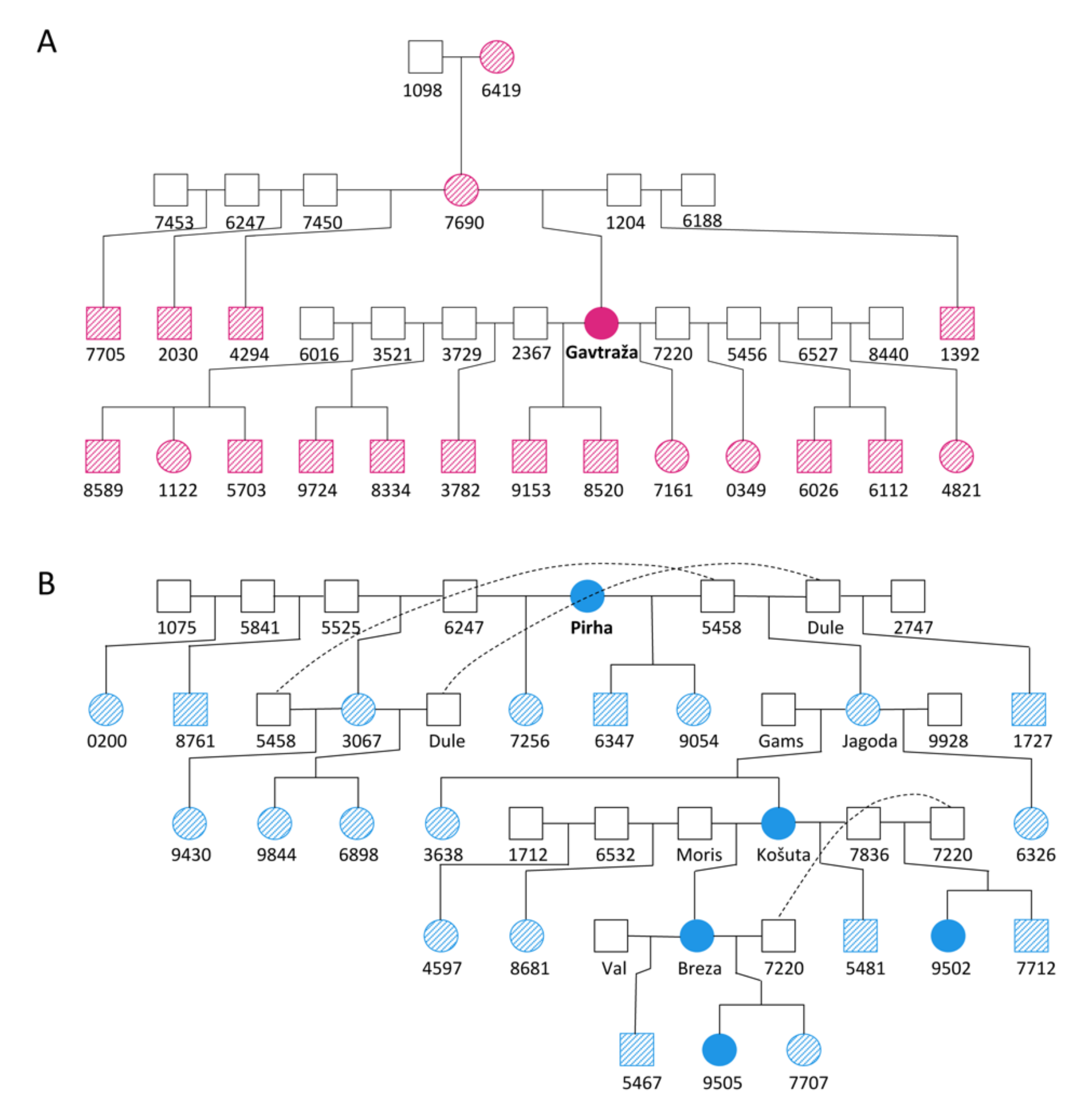
2.1. Identification of “Mutant” Individuals and Homoplasmic Maternal Inheritance
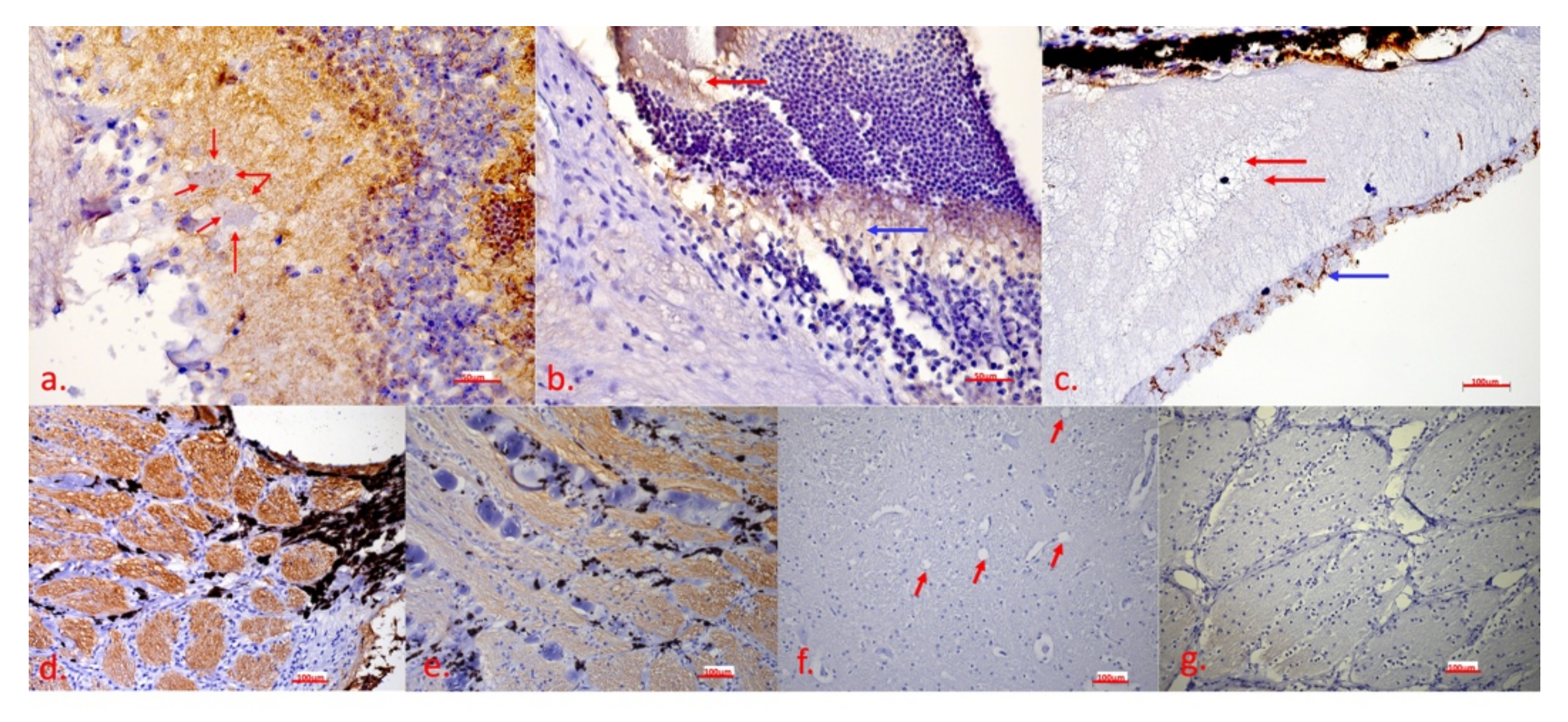
2.2. Clinical and Histopathological Examinations
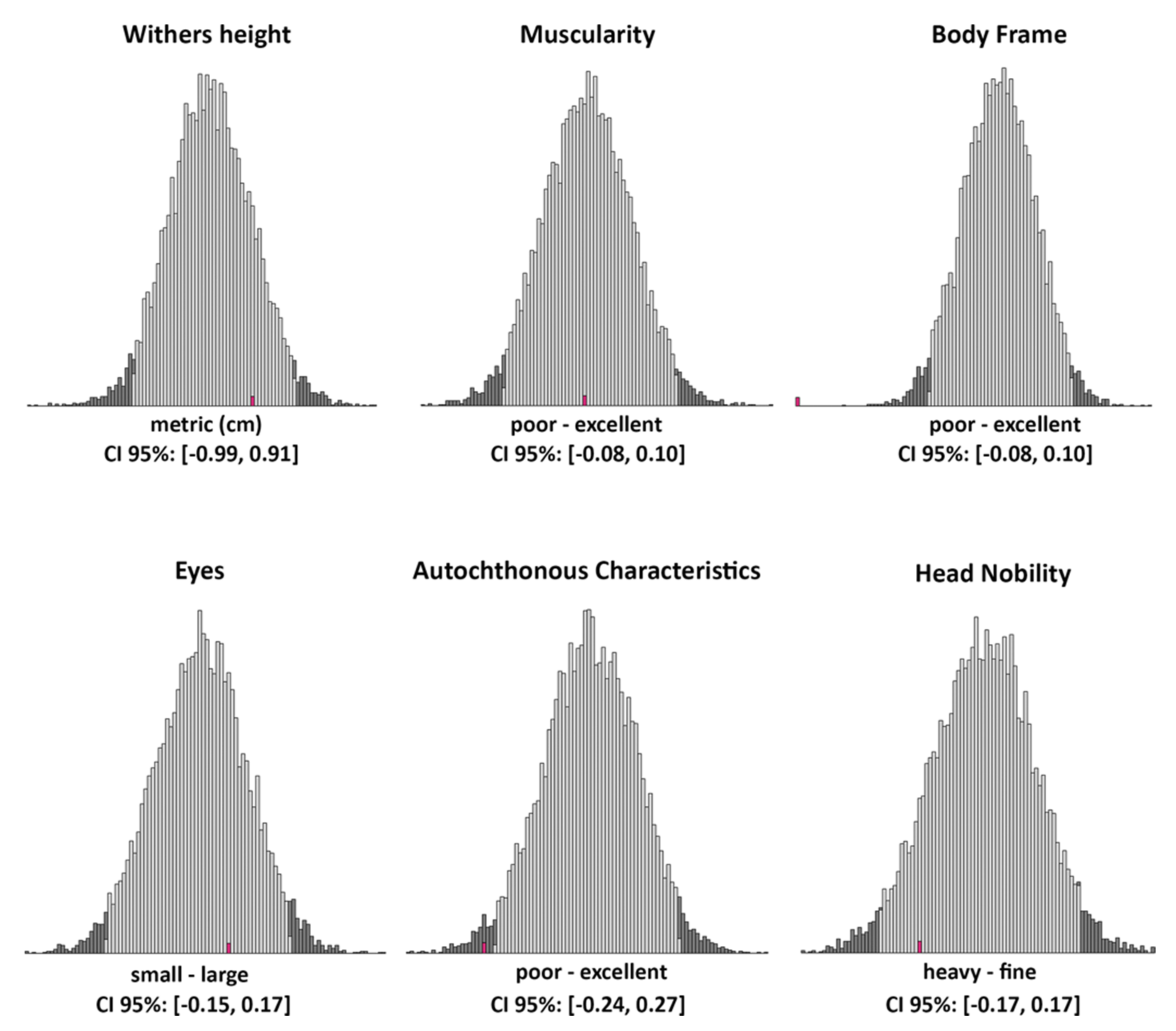
2.3. Suitability of “Mutant” Cattle for Cika Cattle Breeding Goals
3. Discussion
4. Materials and Methods
4.1. Identification of the T10432C Mutation, Histopathology Sampling, and Pedigree Analysis
4.2. Molecular Analyses
4.2.1. DNA Extraction
4.2.2. Primer Selection and Polymerase Chain Reaction
4.2.3. Sanger DNA Sequencing
4.3. Clinical and Histopathological Examinations
4.4. Suitability of “Mutant” Cattle for Cika Breeding Policy
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wallace, D.C. Mitochondrial diseases in man and mouse. Science 1999, 283, 1482–1488. [Google Scholar] [CrossRef] [Green Version]
- Dimauro, S.; Davidzon, G. Mitochondrial DNA and disease. Ann. Med. 2005, 37, 222–232. [Google Scholar] [CrossRef] [PubMed]
- Finsterer, J. Mitochondriopathies. Eur. J. Neurol. 2004, 11, 163–186. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-M. Mitochondrial diseases. J. Epilepsy Res. 2012, 2, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Carelli, V.; Ross-Cisneros, F.N.; Sadun, A.A. Mitochondrial dysfunction as a cause of optic neuropathies. Prog. Retin. Eye Res. 2004, 23, 53–89. [Google Scholar] [CrossRef]
- Newman, N.J. Hereditary Optic Neuropathies: From the Mitochondria to the Optic Nerve. Am. J. Ophthalmol. 2005, 140, 517.e1–517.e9. [Google Scholar] [CrossRef]
- Yu-Wai-Man, P.; Griffiths, P.G.; Chinnery, P.F. Mitochondrial optic neuropathies—Disease mechanisms and therapeutic strategies. Prog. Retin. Eye Res. 2011, 30, 81–114. [Google Scholar] [CrossRef] [Green Version]
- Tsang, S.H.; Aycinena, A.R.P.; Sharma+, T. Mitochondrial Disorder: Kearns-Sayre Syndrome. In Atlas of Inherited Retinal Diseases; Tsang, S.H., Sharma, T., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 161–162. ISBN 978-3-319-95046-4. [Google Scholar]
- Baranowska, I.; Jäderlund, K.H.; Nennesmo, I.; Holmqvist, E.; Heidrich, N.; Larsson, N.-G.; Andersson, G.; Wagner, E.G.H.; Hedhammar, Å.; Wibom, R.; et al. Sensory Ataxic Neuropathy in Golden Retriever Dogs Is Caused by a Deletion in the Mitochondrial tRNATyr Gene. PLoS Genet. 2009, 5, e1000499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trifunovic, A.; Wredenberg, A.; Falkenberg, M.; Spelbrink, J.N.; Rovio, A.T.; Bruder, C.E.; Bohlooly, Y.M.; Gidlöf, S.; Oldfors, A.; Wibom, R.; et al. Premature ageing in mice expressing defective mitochondrial DNA polymerase. Nature 2004, 429, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Wredenberg, A.; Wibom, R.; Wilhelmsson, H.; Graff, C.; Wiener, H.H.; Burden, S.J.; Oldfors, A.; Westerblad, H.; Larsson, N.-G. Increased mitochondrial mass in mitochondrial myopathy mice. Proc. Natl. Acad. Sci. USA 2002, 99, 15066–15071. [Google Scholar] [CrossRef] [Green Version]
- Ingraham, C.A.; Burwell, L.S.; Skalska, J.; Brookes, P.S.; Howell, R.L.; Sheu, S.-S.; Pinkert, C.A. NDUFS4: Creation of a mouse model mimicking a Complex I disorder. Mitochondrion 2009, 9, 204–210. [Google Scholar] [CrossRef] [Green Version]
- Dunn, S.; Cowling, V.H.; Irwin, M.H.; Pinkert, C.A. Animal models of human mitochondrial DNA mutations. Biochim. Biophys. Acta 2012, 1820, 601–607. [Google Scholar] [CrossRef] [Green Version]
- Novosel, D.; Cubric-Curik, V.; Simicic, M.; Dovc, P.; Curik, I. Two detrimental mutations in cattle mitogenome indicate the presence of Leber’s hereditary optic neuropathy. J. Cent. Eur. Agric. 2019, 20, 19–24. [Google Scholar] [CrossRef]
- Behbehani, R.; Melhem, M.; Alghanim, G.; Behbehani, K.; Alsmadi, O. ND4L gene concurrent 10609T>C and 10663T>C mutations are associated with Leber’s hereditary optic neuropathy in a large pedigree from Kuwait. Br. J. Ophthalmol. 2014, 98, 826–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu-Amero, K.K.; Bosley, T.M. Mitochondrial abnormalities in patients with LHON-like optic neuropathies. Invest. Ophthalmol. Vis. Sci. 2006, 47, 4211–4220. [Google Scholar] [CrossRef] [Green Version]
- Simčič, M.; Luštrek, B.; Štepec, M.; Logar, B.; Potočnik, K. Estimation of Genetic Parameters of Type Traits in First Parity Cows of the Autochthonous Cika Cattle in Slovenia. Front. Genet. 2021, 12, 724058. [Google Scholar] [CrossRef]
- Zeviani, M.; Carelli, V. Mitochondrial Retinopathies. Int. J. Mol. Sci. 2021, 23, 210. [Google Scholar] [CrossRef]
- Simčič, M.; Smetko, A.; Sölkner, J.; Seichter, D.; Gorjanc, G.; Kompan, D.; Medugorac, I. Recovery of Native Genetic Background in Admixed Populations Using Haplotypes, Phenotypes, and Pedigree Information—Using Cika Cattle as a Case Breed. PLoS ONE 2015, 10, e0123253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selle, M.L.; Steinsland, I.; Lindgren, F.; Brajkovic, V.; Cubric-Curik, V.; Gorjanc, G. Hierarchical Modelling of Haplotype Effects on a Phylogeny. Front. Genet. 2021, 11, 531218. [Google Scholar] [CrossRef]
- Drögemüller, C.; Reichart, U.; Seuberlich, T.; Oevermann, A.; Baumgartner, M.; Kühni Boghenbor, K.; Stoffel, M.H.; Syring, C.; Meylan, M.; Müller, S.; et al. An Unusual Splice Defect in the Mitofusin 2 Gene (MFN2) Is Associated with Degenerative Axonopathy in Tyrolean Grey Cattle. PLoS ONE 2011, 6, e18931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leber, T. Ueber hereditäre und congenital-angelegte Sehnervenleiden. Albr. Graefes Arch. Ophthalmol. 2005, 17, 249–291. [Google Scholar] [CrossRef]
- Giles, R.E.; Blanc, H.; Cann, H.M.; Wallace, D.C. Maternal inheritance of human mitochondrial DNA. Proc. Natl. Acad. Sci. USA 1980, 77, 6715–6719. [Google Scholar] [CrossRef] [Green Version]
- Wallace, D.C.; Singh, G.; Lott, M.T.; Hodge, J.A.; Schurr, T.G.; Lezza, A.M.; Elsas, L.J., 2nd; Nikoskelainen, E.K. Mitochondrial DNA mutation associated with Leber’s hereditary optic neuropathy. Science 1988, 242, 1427–1430. [Google Scholar] [CrossRef]
- Sadun, A.A.; Carelli, V.; Bose, S.; Ross-Cisneros, F.N.; Barboni, P.; Ahrens, E.T. First application of extremely high-resolution magnetic resonance imaging to study microscopic features of normal and LHON human optic nerve. Ophthalmology 2002, 109, 1085–1091. [Google Scholar] [CrossRef]
- Delettre, C.; Lenaers, G.; Griffoin, J.M.; Gigarel, N.; Lorenzo, C.; Belenguer, P.; Pelloquin, L.; Grosgeorge, J.; Turc-Carel, C.; Perret, E.; et al. Nuclear gene OPA1, encoding a mitochondrial dynamin-related protein, is mutated in dominant optic atrophy. Nat. Genet. 2000, 26, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Alexander, C.; Votruba, M.; Pesch, U.E.; Thiselton, D.L.; Mayer, S.; Moore, A.; Rodriguez, M.; Kellner, U.; Leo-Kottler, B.; Auburger, G.; et al. OPA1, encoding a dynamin-related GTPase, is mutated in autosomal dominant optic atrophy linked to chromosome 3q28. Nat. Genet. 2000, 26, 211–215. [Google Scholar] [CrossRef]
- Chinnery, P.F.; Johnson, M.A.; Wardell, T.M.; Singh-Kler, R.; Hayes, C.; Brown, D.T.; Taylor, R.W.; Bindoff, L.A.; Turnbull, D.M. The epidemiology of pathogenic mitochondrial DNA mutations. Ann. Neurol. 2000, 48, 188–193. [Google Scholar] [CrossRef]
- Wallace, D.C. Mitochondria, bioenergetics, and the epigenome in eukaryotic and human evolution. Cold Spring Harb. Symp. Quant. Biol. 2009, 74, 383–393. [Google Scholar] [CrossRef]
- Wallace, D.C. A Mitochondrial Paradigm of Metabolic and Degenerative Diseases, Aging, and Cancer: A Dawn for Evolutionary Medicine. Annu. Rev. Genet. 2005, 39, 359–407. [Google Scholar] [CrossRef] [Green Version]
- Cubric-Curik, V.; Novosel, D.; Brajkovic, V.; Rota Stabelli, O.; Krebs, S.; Sölkner, J.; Šalamon, D.; Ristov, S.; Berger, B.; Trivizaki, S.; et al. Large-scale mitogenome sequencing reveals consecutive expansions of domestic taurine cattle and supports sporadic aurochs introgression. Evol. Appl. 2021, 15, 663–678. [Google Scholar] [CrossRef]
- Ristov, S.; Brajkovic, V.; Cubric-Curik, V.; Michieli, I.; Curik, I. MaGelLAn 1.0: A software to facilitate quantitative and population genetic analysis of maternal inheritance by combination of molecular and pedigree information. Genet. Sel. Evol. 2016, 48, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wray, N.R.; Ripke, S.; Mattheisen, M.; Trzaskowski, M.; Byrne, E.M.; Abdellaoui, A.; Adams, M.J.; Agerbo, E.; Air, T.M.; Andlauer, T.M.F.; et al. Genome-wide association analyses identify 44 risk variants and refine the genetic architecture of major depression. Nat. Genet. 2018, 50, 668–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crouch, D.J.M.; Bodmer, W.F. Polygenic inheritance, GWAS, polygenic risk scores, and the search for functional variants. Proc. Natl. Acad. Sci. USA 2020, 117, 18924–18933. [Google Scholar] [CrossRef]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.; Francois, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Couch, S.P.; Bray, A.P.; Ismay, C.; Chasnovski, E.; Baumer, B.S.; Cetinkaya-Rundel, M. infer: An R package for tidyverse-friendly statistical inference. J. Open Source Softw. 2021, 6, 3661. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Novosel, D.; Brajković, V.; Simčič, M.; Zorc, M.; Svara, T.; Cakanic, K.B.; Jungić, A.; Logar, B.; Cubric-Curik, V.; Dovc, P.; et al. The Consequences of Mitochondrial T10432C Mutation in Cika Cattle: A “Potential” Model for Leber’s Hereditary Optic Neuropathy. Int. J. Mol. Sci. 2022, 23, 6335. https://doi.org/10.3390/ijms23116335
Novosel D, Brajković V, Simčič M, Zorc M, Svara T, Cakanic KB, Jungić A, Logar B, Cubric-Curik V, Dovc P, et al. The Consequences of Mitochondrial T10432C Mutation in Cika Cattle: A “Potential” Model for Leber’s Hereditary Optic Neuropathy. International Journal of Molecular Sciences. 2022; 23(11):6335. https://doi.org/10.3390/ijms23116335
Chicago/Turabian StyleNovosel, Dinko, Vladimir Brajković, Mojca Simčič, Minja Zorc, Tanja Svara, Karmen Branovic Cakanic, Andreja Jungić, Betka Logar, Vlatka Cubric-Curik, Peter Dovc, and et al. 2022. "The Consequences of Mitochondrial T10432C Mutation in Cika Cattle: A “Potential” Model for Leber’s Hereditary Optic Neuropathy" International Journal of Molecular Sciences 23, no. 11: 6335. https://doi.org/10.3390/ijms23116335
APA StyleNovosel, D., Brajković, V., Simčič, M., Zorc, M., Svara, T., Cakanic, K. B., Jungić, A., Logar, B., Cubric-Curik, V., Dovc, P., & Curik, I. (2022). The Consequences of Mitochondrial T10432C Mutation in Cika Cattle: A “Potential” Model for Leber’s Hereditary Optic Neuropathy. International Journal of Molecular Sciences, 23(11), 6335. https://doi.org/10.3390/ijms23116335