
Different Sources of Mesenchymal Stem Cells for Tissue Regeneration: A Guide to Identifying the Most Favorable One in Orthopedics and Dentistry Applications

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Mesenchymal Stem Cell Sources
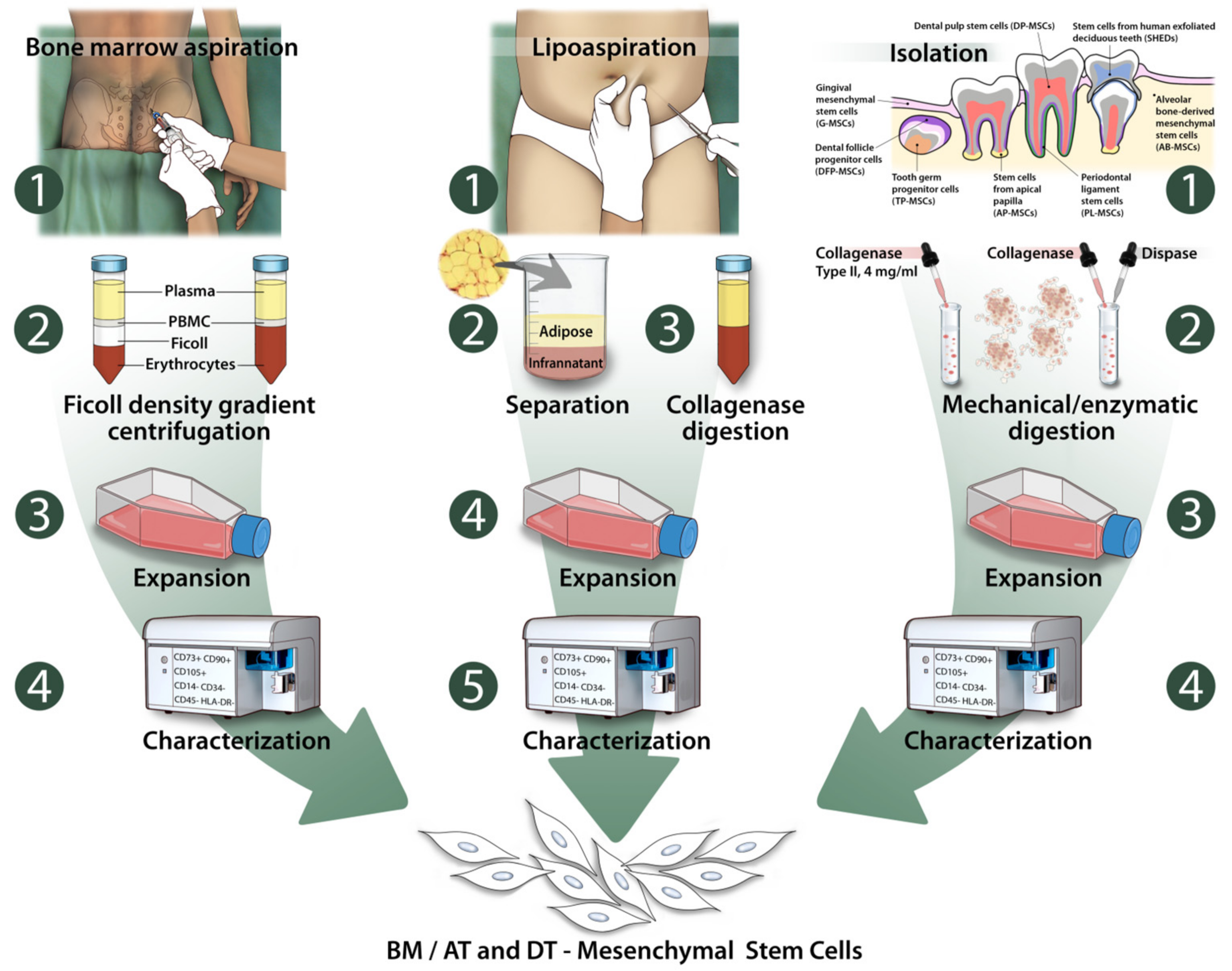
2.1. Bone Marrow Stromal Cells (BMSCs)

{kind=link}
{kind=link}
| MSC Type | Markers | References |
|---|---|---|
| BM-MSCs | CD73, THY1/CD90, CD105, CD146/MCAM, CD29, CD44, STRO-1, OCT4, NANOG | Dominici et al., Samsonraj et al. [11,43]. |
| AT-MSCs | DPP4/CD26, PDGFRa, CD29, CD34, SCA1, CD55, THY1/C90, CD24, BMP7, PI16, WNT2, ANXA3 | Merrick et al. [37]. |
| DPSCs | CD44, CD90, CD105, CD73, STRO-1 | Mattei et al. [52] |
2.2. Adipose Tissue Derived Stem Cells (AT-MSCs)
2.3. Dental Tissue Derived Stem Cells (DT-MSCs)
2.3.1. Dental Pulp Stem Cells (DP-MSCs)
2.3.2. Stem Cells from Human Exfoliated Deciduous Teeth (SHEDs)
2.3.3. Periodontal Ligament Stem Cells (PL-MSCs)
2.3.4. Dental Follicle Progenitor Cells (DFP-MSCs)
2.3.5. Alveolar Bone-Derived Mesenchymal Stem Cells (AB-MSCs)
2.3.6. Stem Cells from Apical Papilla (AP-MSCs)
2.3.7. Tooth Germ Progenitor Cells (TP-MSCs)
2.3.8. Gingival Mesenchymal Stem Cells (G-MSCs)
3. Source Comparison
3.1. Proliferation
3.2. Migration Capacity
3.3. Clonogenicity
3.4. Angiogenic Capacity
3.5. Differentiation Potential
3.6. Secretome
4. Methods
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Goujon, E. Recherches Experimentales Sur Les Proprietes Physiologiques de La Moelle Des Os. J. Anat. Physiol. Norm. Pathol. Homme Anim. 1869, 6, 399–412. [Google Scholar]
- Friedenstein, A.J.; Chailakhjan, R.K.; Lalykina, K.S. The Development of Fibroblast Colonies in Monolayer Cultures of Guinea-Pig Bone Marrow and Spleen Cells. Cell Tissue Kinet. 1970, 3, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Friedenstein, A.J.; Gorskaja, J.F.; Kulagina, N.N. Fibroblast Precursors in Normal and Irradiated Mouse Hematopoietic Organs. Exp. Hematol. 1976, 4, 267–274. [Google Scholar]
- Friedenstein, A.J.; Petrakova, K.V.; Kurolesova, A.I.; Frolova, G.P. Heterotopic of Bone Marrow. Analysis of Precursor Cells for Osteogenic and Hematopoietic Tissues. Transplantation 1968, 6, 230–247. [Google Scholar] [CrossRef] [PubMed]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage Potential of Adult Human Mesenchymal Stem Cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caplan, A.I. Mesenchymal Stem Cells. J. Orthop. Res. 1991, 9, 641–650. [Google Scholar] [CrossRef]
- Caplan, A.I. Mesenchymal Stem Cells: Time to Change the Name! Stem Cells Transl. Med. 2017, 6, 1445–1451. [Google Scholar] [CrossRef] [Green Version]
- Bianco, P.; Robey, P.G. Skeletal Stem Cells. Development 2015, 142, 1023–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambrosi, T.H.; Longaker, M.T.; Chan, C.K.F. A Revised Perspective of Skeletal Stem Cell Biology. Front. Cell Dev. Biol. 2019, 7, 189. [Google Scholar] [CrossRef] [Green Version]
- Viswanathan, S.; Shi, Y.; Galipeau, J.; Krampera, M.; Leblanc, K.; Martin, I.; Nolta, J.; Phinney, D.G.; Sensebe, L. Mesenchymal Stem versus Stromal Cells: International Society for Cell & Gene Therapy (ISCT®) Mesenchymal Stromal Cell Committee Position Statement on Nomenclature. Cytotherapy 2019, 21, 1019–1024. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal Criteria for Defining Multipotent Mesenchymal Stromal Cells. The International Society for Cellular Therapy Position Statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Zuk, P.A.; Zhu, M.; Mizuno, H.; Huang, J.; Futrell, J.W.; Katz, A.J.; Benhaim, P.; Lorenz, H.P.; Hedrick, M.H. Multilineage Cells from Human Adipose Tissue: Implications for Cell-Based Therapies. Tissue Eng. 2001, 7, 211–228. [Google Scholar] [CrossRef] [Green Version]
- Beltrami, A.P.; Cesselli, D.; Bergamin, N.; Marcon, P.; Rigo, S.; Puppato, E.; D’Aurizio, F.; Verardo, R.; Piazza, S.; Pignatelli, A.; et al. Multipotent Cells Can Be Generated in Vitro from Several Adult Human Organs (Heart, Liver, and Bone Marrow). Blood 2007, 110, 3438–3446. [Google Scholar] [CrossRef] [PubMed]
- De Coppi, P.; Bartsch, G.; Siddiqui, M.M.; Xu, T.; Santos, C.C.; Perin, L.; Mostoslavsky, G.; Serre, A.C.; Snyder, E.Y.; Yoo, J.J.; et al. Isolation of Amniotic Stem Cell Lines with Potential for Therapy. Nat. Biotechnol. 2007, 25, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Erices, A.; Conget, P.; Minguell, J.J. Mesenchymal Progenitor Cells in Human Umbilical Cord Blood: Mesenchymal Progenitors in Cord Blood. Br. J. Haematol. 2000, 109, 235–242. [Google Scholar] [CrossRef]
- Gronthos, S.; Mankani, M.; Brahim, J.; Robey, P.G.; Shi, S. Postnatal Human Dental Pulp Stem Cells (DPSCs) in Vitro and in Vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 13625–13630. [Google Scholar] [CrossRef] [Green Version]
- Bi, Y.; Ehirchiou, D.; Kilts, T.M.; Inkson, C.A.; Embree, M.C.; Sonoyama, W.; Li, L.; Leet, A.I.; Seo, B.-M.; Zhang, L.; et al. Identification of Tendon Stem/Progenitor Cells and the Role of the Extracellular Matrix in Their Niche. Nat. Med. 2007, 13, 1219–1227. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Menicanin, D.; Gronthos, S.; Bartold, P.M. Stem Cells, Tissue Engineering and Periodontal Regeneration. Aust. Dent. J. 2014, 59, 117–130. [Google Scholar] [CrossRef]
- Zhang, Q.Z.; Nguyen, A.L.; Yu, W.H.; Le, A.D. Human Oral Mucosa and Gingiva. J. Dent. Res. 2012, 91, 1011–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopa, S.; Colombini, A.; Stanco, D.; de Girolamo, L.; Sansone, V.; Moretti, M. Donor-Matched Mesenchymal Stem Cells from Knee Infrapatellar and Subcutaneous Adipose Tissue of Osteoarthritic Donors Display Differential Chondrogenic and Osteogenic Commitment. Eur. Cells Mater. 2014, 27, 298–311. [Google Scholar] [CrossRef]
- Stanco, D.; Viganò, M.; Perucca Orfei, C.; Di Giancamillo, A.; Peretti, G.M.; Lanfranchi, L.; de Girolamo, L. Multidifferentiation Potential of Human Mesenchymal Stem Cells from Adipose Tissue and Hamstring Tendons for Musculoskeletal Cell-Based Therapy. Regen. Med. 2015, 10, 729–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musina, R.A.; Bekchanova, E.S.; Belyavskii, A.V.; Sukhikh, G.T. Differentiation Potential of Mesenchymal Stem Cells of Different Origin. Bull. Exp. Biol. Med. 2006, 141, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Wagner, W.; Wein, F.; Seckinger, A.; Frankhauser, M.; Wirkner, U.; Krause, U.; Blake, J.; Schwager, C.; Eckstein, V.; Ansorge, W.; et al. Comparative Characteristics of Mesenchymal Stem Cells from Human Bone Marrow, Adipose Tissue, and Umbilical Cord Blood. Exp. Hematol. 2005, 33, 1402–1416. [Google Scholar] [CrossRef]
- Sensebe, L.; Krampera, M.; Schrezenmeier, H.; Bourin, P.; Giordano, R. Mesenchymal Stem Cells for Clinical Application. Vox Sang. 2010, 98, 93–107. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, S.; Pittenger, M.F. Human Mesenchymal Stem Cells Modulate Allogeneic Immune Cell Responses. Blood 2005, 105, 1815–1822. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, S.A.; Pinxteren, J.; Roobrouck, V.D.; Luyckx, A.; Van’t Hof, W.; Deans, R.; Verfaillie, C.M.; Waer, M.; Billiau, A.D.; Van Gool, S.W. Human Multipotent Adult Progenitor Cells Are Nonimmunogenic and Exert Potent Immunomodulatory Effects on Alloreactive T-Cell Responses. Cell Transplant. 2013, 22, 1915–1928. [Google Scholar] [CrossRef]
- Uccelli, A.; Moretta, L.; Pistoia, V. Immunoregulatory Function of Mesenchymal Stem Cells. Eur. J. Immunol. 2006, 36, 2566–2573. [Google Scholar] [CrossRef]
- López-García, L.; Castro-Manrreza, M.E. TNF-α and IFN-γ Participate in Improving the Immunoregulatory Capacity of Mesenchymal Stem/Stromal Cells: Importance of Cell–Cell Contact and Extracellular Vesicles. Int. J. Mol. Sci. 2021, 22, 9531. [Google Scholar] [CrossRef]
- Shi, Y.; Wang, Y.; Li, Q.; Liu, K.; Hou, J.; Shao, C.; Wang, Y. Immunoregulatory Mechanisms of Mesenchymal Stem and Stromal Cells in Inflammatory Diseases. Nat. Rev. Nephrol. 2018, 14, 493–507. [Google Scholar] [CrossRef]
- Ding, G.; Du, J.; Hu, X.; Ao, Y. Mesenchymal Stem Cells From Different Sources in Meniscus Repair and Regeneration. Front. Bioeng. Biotechnol. 2022, 10, 796367. [Google Scholar] [CrossRef]
- Estrela, C.; de Alencar, A.H.G.; Kitten, G.T.; Vencio, E.F.; Gava, E. Mesenchymal Stem Cells in the Dental Tissues: Perspectives for Tissue Regeneration. Braz. Dent. J. 2011, 22, 91–98. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Lee, A.E.; Xu, Q.; Zhang, Q.; Le, A.D. Gingiva-Derived Mesenchymal Stem Cells: Potential Application in Tissue Engineering and Regenerative Medicine—A Comprehensive Review. Front. Immunol. 2021, 12, 1282. [Google Scholar] [CrossRef]
- Sonoyama, W.; Liu, Y.; Yamaza, T.; Tuan, R.S.; Wang, S.; Shi, S.; Huang, G.T.-J. Characterization of the Apical Papilla and Its Residing Stem Cells from Human Immature Permanent Teeth: A Pilot Study. J. Endod. 2008, 34, 166–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soliman, H.; Theret, M.; Scott, W.; Hill, L.; Underhill, T.M.; Hinz, B.; Rossi, F.M.V. Multipotent Stromal Cells: One Name, Multiple Identities. Cell Stem Cell 2021, 28, 1690–1707. [Google Scholar] [CrossRef]
- Pagella, P.; de Vargas Roditi, L.; Stadlinger, B.; Moor, A.E.; Mitsiadis, T.A. A Single-Cell Atlas of Human Teeth. iScience 2021, 24, 102405. [Google Scholar] [CrossRef] [PubMed]
- Vijay, J.; Gauthier, M.-F.; Biswell, R.L.; Louiselle, D.A.; Johnston, J.J.; Cheung, W.A.; Belden, B.; Pramatarova, A.; Biertho, L.; Gibson, M.; et al. Single-Cell Analysis of Human Adipose Tissue Identifies Depot- and Disease-Specific Cell Types. Nat. Metab. 2020, 2, 97–109. [Google Scholar] [CrossRef]
- Tikhonova, A.N.; Dolgalev, I.; Hu, H.; Sivaraj, K.K.; Hoxha, E.; Cuesta-Domínguez, Á.; Pinho, S.; Akhmetzyanova, I.; Gao, J.; Witkowski, M.; et al. The Bone Marrow Microenvironment at Single-Cell Resolution. Nature 2019, 569, 222–228. [Google Scholar] [CrossRef]
- Merrick, D.; Sakers, A.; Irgebay, Z.; Okada, C.; Calvert, C.; Morley, M.P.; Percec, I.; Seale, P. Identification of a Mesenchymal Progenitor Cell Hierarchy in Adipose Tissue. Science 2019, 364, eaav2501. [Google Scholar] [CrossRef]
- Miltenyi, S.; Müller, W.; Weichel, W.; Radbruch, A. High Gradient Magnetic Cell Separation with MACS. Cytometry 1990, 11, 231–238. [Google Scholar] [CrossRef]
- Gronthos, S.; Zannettino, A.C.W.; Hay, S.J.; Shi, S.; Graves, S.E.; Kortesidis, A.; Simmons, P.J. Molecular and Cellular Characterisation of Highly Purified Stromal Stem Cells Derived from Human Bone Marrow. J. Cell Sci. 2003, 116, 1827–1835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grisendi, G.; Annerén, C.; Cafarelli, L.; Sternieri, R.; Veronesi, E.; Cervo, G.L.; Luminari, S.; Maur, M.; Frassoldati, A.; Palazzi, G.; et al. GMP-Manufactured Density Gradient Media for Optimized Mesenchymal Stromal/Stem Cell Isolation and Expansion. Cytotherapy 2010, 12, 466–477. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.I. Adult Mesenchymal Stem Cells for Tissue Engineering versus Regenerative Medicine. J. Cell. Physiol. 2007, 213, 341–347. [Google Scholar] [CrossRef]
- Siegel, G.; Kluba, T.; Hermanutz-Klein, U.; Bieback, K.; Northoff, H.; Schäfer, R. Phenotype, Donor Age and Gender Affect Function of Human Bone Marrow-Derived Mesenchymal Stromal Cells. BMC Med. 2013, 11, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samsonraj, R.M.; Raghunath, M.; Nurcombe, V.; Hui, J.H.; van Wijnen, A.J.; Cool, S.M. Concise Review: Multifaceted Characterization of Human Mesenchymal Stem Cells for Use in Regenerative Medicine: Characterization of Human Mesenchymal Stem Cells. Stem Cells Transl. Med. 2017, 6, 2173–2185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolock, S.L.; Krishnan, I.; Tenen, D.E.; Matkins, V.; Camacho, V.; Patel, S.; Agarwal, P.; Bhatia, R.; Tenen, D.G.; Klein, A.M.; et al. Mapping Distinct Bone Marrow Niche Populations and Their Differentiation Paths. Cell Rep. 2019, 28, 302–311.e5. [Google Scholar] [CrossRef] [Green Version]
- Woods, K.; Guezguez, B. Dynamic Changes of the Bone Marrow Niche: Mesenchymal Stromal Cells and Their Progeny During Aging and Leukemia. Front. Cell Dev. Biol. 2021, 9, 714716. [Google Scholar] [CrossRef]
- Liu, S.; Liu, D.; Chen, C.; Hamamura, K.; Moshaverinia, A.; Yang, R.; Liu, Y.; Jin, Y.; Shi, S. MSC Transplantation Improves Osteopenia via Epigenetic Regulation of Notch Signaling in Lupus. Cell Metab. 2015, 22, 606–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lian, J.B.; Stein, G.S.; van Wijnen, A.J.; Stein, J.L.; Hassan, M.Q.; Gaur, T.; Zhang, Y. MicroRNA Control of Bone Formation and Homeostasis. Nat. Rev. Endocrinol. 2012, 8, 212–227. [Google Scholar] [CrossRef] [Green Version]
- Zaidi, M. Skeletal Remodeling in Health and Disease. Nat. Med. 2007, 13, 791–801. [Google Scholar] [CrossRef]
- Pittenger, M.F.; Discher, D.E.; Péault, B.M.; Phinney, D.G.; Hare, J.M.; Caplan, A.I. Mesenchymal Stem Cell Perspective: Cell Biology to Clinical Progress. Npj Regen. Med. 2019, 4, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salamanna, F.; Contartese, D.; Borsari, V.; Pagani, S.; Barbanti Brodano, G.; Griffoni, C.; Ricci, A.; Gasbarrini, A.; Fini, M. Two Hits for Bone Regeneration in Aged Patients: Vertebral Bone Marrow Clot as a Biological Scaffold and Powerful Source of Mesenchymal Stem Cells. Front. Bioeng. Biotechnol. 2021, 9, 807679. [Google Scholar] [CrossRef]
- Mattei, V.; Santacroce, C.; Tasciotti, V.; Martellucci, S.; Santilli, F.; Manganelli, V.; Piccoli, L.; Misasi, R.; Sorice, M.; Garofalo, T. Role of Lipid Rafts in Neuronal Differentiation of Dental Pulp-Derived Stem Cells. Exp. Cell Res. 2015, 339, 231–240. [Google Scholar] [CrossRef]
- Boquest, A.C.; Shahdadfar, A.; Brinchmann, J.E.; Collas, P. Isolation of Stromal Stem Cells from Human Adipose Tissue. Methods Mol. Biol. 2006, 325, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.T.-J.; Gronthos, S.; Shi, S. Mesenchymal Stem Cells Derived from Dental Tissues vs. Those from Other Sources. J. Dent. Res. 2009, 88, 792–806. [Google Scholar] [CrossRef]
- Hashemi-Beni, B.; Khoroushi, M.; Foroughi, M.R.; Karbasi, S.; Khademi, A.A. Tissue Engineering: Dentin—Pulp Complex Regeneration Approaches (A Review). Tissue Cell 2017, 49, 552–564. [Google Scholar] [CrossRef]
- Liu, H.; Gronthos, S.; Shi, S. Dental Pulp Stem Cells. Methods Enzymol. 2006, 419, 99–113. [Google Scholar] [CrossRef]
- Huang, C.-C.; Narayanan, R.; Alapati, S.; Ravindran, S. Exosomes as Biomimetic Tools for Stem Cell Differentiation: Applications in Dental Pulp Tissue Regeneration. Biomaterials 2016, 111, 103–115. [Google Scholar] [CrossRef] [Green Version]
- Graziano, A.; d’Aquino, R.; Laino, G.; Papaccio, G. Dental Pulp Stem Cells: A Promising Tool for Bone Regeneration. Stem Cell Rev. 2008, 4, 21–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrúa, C.P.; Centeno, E.G.Z.; da Rosa, L.C.; do Amaral, C.C.; Severo, R.F.; Sarkis-Onofre, R.; Nascimento, G.G.; Cordenonzi, G.; Bast, R.K.; Demarco, F.F.; et al. How Has Dental Pulp Stem Cells Isolation Been Conducted? A Scoping Review. Braz. Oral Res. 2017, 31, e87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferro, F.; Spelat, R.; Baheney, C.S. Dental Pulp Stem Cell (DPSC) Isolation, Characterization, and Differentiation. Methods Mol. Biol. 2014, 1210, 91–115. [Google Scholar] [CrossRef]
- Mattei, V.; Martellucci, S.; Pulcini, F.; Santilli, F.; Sorice, M.; Delle Monache, S. Regenerative Potential of DPSCs and Revascularization: Direct, Paracrine or Autocrine Effect? Stem Cell Rev. Rep. 2021, 17, 1635–1646. [Google Scholar] [CrossRef] [PubMed]
- Ledesma-Martínez, E.; Mendoza-Núñez, V.M.; Santiago-Osorio, E. Mesenchymal Stem Cells Derived from Dental Pulp: A Review. Stem Cells Int. 2016, 2016, 4709572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miura, M.; Gronthos, S.; Zhao, M.; Lu, B.; Fisher, L.W.; Robey, P.G.; Shi, S. SHED: Stem Cells from Human Exfoliated Deciduous Teeth. Proc. Natl. Acad. Sci. USA 2003, 100, 5807–5812. [Google Scholar] [CrossRef] [Green Version]
- Rosa, V.; Dubey, N.; Islam, I.; Min, K.-S.; Nör, J.E. Pluripotency of Stem Cells from Human Exfoliated Deciduous Teeth for Tissue Engineering. Stem Cells Int. 2016, 2016, 5957806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishino, Y.; Yamada, Y.; Ebisawa, K.; Nakamura, S.; Okabe, K.; Umemura, E.; Hara, K.; Ueda, M. Stem Cells from Human Exfoliated Deciduous Teeth (SHED) Enhance Wound Healing and the Possibility of Novel Cell Therapy. Cytotherapy 2011, 13, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Collado-González, M.; Pecci-Lloret, M.P.; García-Bernal, D.; Aznar-Cervantes, S.; Oñate-Sánchez, R.E.; Moraleda, J.M.; Cenis, J.L.; Rodríguez-Lozano, F.J. Biological Effects of Silk Fibroin 3D Scaffolds on Stem Cells from Human Exfoliated Deciduous Teeth (SHEDs). Odontology 2018, 106, 125–134. [Google Scholar] [CrossRef]
- Yamaza, T.; Kentaro, A.; Chen, C.; Liu, Y.; Shi, Y.; Gronthos, S.; Wang, S.; Shi, S. Immunomodulatory Properties of Stem Cells from Human Exfoliated Deciduous Teeth. Stem Cell Res. Ther. 2010, 1, 5. [Google Scholar] [CrossRef] [Green Version]
- Seo, B.-M.; Miura, M.; Gronthos, S.; Bartold, P.M.; Batouli, S.; Brahim, J.; Young, M.; Robey, P.G.; Wang, C.-Y.; Shi, S. Investigation of Multipotent Postnatal Stem Cells from Human Periodontal Ligament. Lancet 2004, 364, 149–155. [Google Scholar] [CrossRef]
- Chamila Prageeth Pandula, P.K.; Samaranayake, L.P.; Jin, L.J.; Zhang, C. Periodontal Ligament Stem Cells: An Update and Perspectives. J. Investig. Clin. Dent. 2014, 5, 81–90. [Google Scholar] [CrossRef]
- Mrozik, K.; Gronthos, S.; Shi, S.; Bartold, P.M. A Method to Isolate, Purify, and Characterize Human Periodontal Ligament Stem Cells. Methods Mol. Biol. 2017, 1537, 413–427. [Google Scholar] [CrossRef]
- Morsczeck, C.; Schmalz, G. Transcriptomes and Proteomes of Dental Follicle Cells. J. Dent. Res. 2010, 89, 445–456. [Google Scholar] [CrossRef]
- Yang, H.; Li, J.; Sun, J.; Guo, W.; Li, H.; Chen, J.; Hu, Y.; Tian, W.; Li, S. Cells Isolated from Cryopreserved Dental Follicle Display Similar Characteristics to Cryopreserved Dental Follicle Cells. Cryobiology 2017, 78, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, T.; Suardita, K.; Ishii, M.; Sugiyama, M.; Igarashi, A.; Oda, R.; Nishimura, M.; Saito, M.; Nakagawa, K.; Yamanaka, K.; et al. Alveolar Bone Marrow as a Cell Source for Regenerative Medicine: Differences between Alveolar and Iliac Bone Marrow Stromal Cells. J. Bone Miner. Res. 2005, 20, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-C.; Oh, S.-Y.; Lee, J.-S.; Park, S.-Y.; Choi, E.-Y.; Cho, K.-S.; Kim, C.-S. In Vivo Bone Formation by Human Alveolar-Bone-Derived Mesenchymal Stem Cells Obtained during Implant Osteotomy Using Biphasic Calcium Phosphate Ceramics or Bio-Oss as Carriers. J. Biomed. Mater. Res. Part B Appl. Biomater. 2016, 104, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yu, F.; Sun, Y.; Jiang, B.; Zhang, W.; Yang, J.; Xu, G.-T.; Liang, A.; Liu, S. Concise Reviews: Characteristics and Potential Applications of Human Dental Tissue-Derived Mesenchymal Stem Cells. Stem Cells 2015, 33, 627–638. [Google Scholar] [CrossRef]
- Sonoyama, W.; Liu, Y.; Fang, D.; Yamaza, T.; Seo, B.-M.; Zhang, C.; Liu, H.; Gronthos, S.; Wang, C.-Y.; Wang, S.; et al. Mesenchymal Stem Cell-Mediated Functional Tooth Regeneration in Swine. PLoS ONE 2006, 1, e79. [Google Scholar] [CrossRef] [Green Version]
- Kang, J.; Fan, W.; Deng, Q.; He, H.; Huang, F. Stem Cells from the Apical Papilla: A Promising Source for Stem Cell-Based Therapy. Biomed. Res. Int. 2019, 2019, 6104738. [Google Scholar] [CrossRef] [Green Version]
- Nada, O.A.; El Backly, R.M. Stem Cells From the Apical Papilla (SCAP) as a Tool for Endogenous Tissue Regeneration. Front. Bioeng. Biotechnol. 2018, 6, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, E.; Yagi, K.; Kojima, M.; Yagyuu, T.; Ohshima, A.; Sobajima, S.; Tadokoro, M.; Katsube, Y.; Isoda, K.; Kondoh, M.; et al. Multipotent Cells from the Human Third Molar: Feasibility of Cell-Based Therapy for Liver Disease. Differentiation 2008, 76, 495–505. [Google Scholar] [CrossRef]
- Doğan, A.; Demirci, S.; Şahin, F. In Vitro Differentiation of Human Tooth Germ Stem Cells into Endothelial- and Epithelial-like Cells. Cell Biol. Int. 2015, 39, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Chalisserry, E.P.; Nam, S.Y.; Park, S.H.; Anil, S. Therapeutic Potential of Dental Stem Cells. J. Tissue Eng. 2017, 8, 2041731417702531. [Google Scholar] [CrossRef]
- Jiang, Q.; Yu, Y.; Ruan, H.; Luo, Y.; Guo, X. Morphological and Functional Characteristics of Human Gingival Junctional Epithelium. BMC Oral Health 2014, 14, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fawzy El-Sayed, K.M.; Dörfer, C.E. Gingival Mesenchymal Stem/Progenitor Cells: A Unique Tissue Engineering Gem. Stem Cells Int. 2016, 2016, 7154327. [Google Scholar] [CrossRef] [Green Version]
- Huang, F.; Liu, Z.M.; Zheng, S.G. Updates on GMSCs Treatment for Autoimmune Diseases. Curr. Stem Cell Res. Ther. 2018, 13, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Riekstina, U.; Cakstina, I.; Parfejevs, V.; Hoogduijn, M.; Jankovskis, G.; Muiznieks, I.; Muceniece, R.; Ancans, J. Embryonic Stem Cell Marker Expression Pattern in Human Mesenchymal Stem Cells Derived from Bone Marrow, Adipose Tissue, Heart and Dermis. Stem Cell Rev. 2009, 5, 378–386. [Google Scholar] [CrossRef]
- Heo, J.S.; Choi, Y.; Kim, H.-S.; Kim, H.O. Comparison of Molecular Profiles of Human Mesenchymal Stem Cells Derived from Bone Marrow, Umbilical Cord Blood, Placenta and Adipose Tissue. Int. J. Mol. Med. 2016, 37, 115–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zong, C.; Zhao, L.; Huang, C.; Chen, Y.; Tian, L. Isolation and Culture of Bone Marrow Mesenchymal Stem Cells from the Human Mandible. JoVE J. Vis. Exp. 2022, 182, e63811. [Google Scholar] [CrossRef]
- Dmitrieva, R.I.; Minullina, I.R.; Bilibina, A.A.; Tarasova, O.V.; Anisimov, S.V.; Zaritskey, A.Y. Bone Marrow- and Subcutaneous Adipose Tissue-Derived Mesenchymal Stem Cells: Differences and Similarities. Cell Cycle 2012, 11, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, O.; Katsube, Y.; Hirose, M.; Ohgushi, H.; Ito, H. Comparison of Osteogenic Ability of Rat Mesenchymal Stem Cells from Bone Marrow, Periosteum, and Adipose Tissue. Calcif. Tissue Int. 2008, 82, 238–247. [Google Scholar] [CrossRef]
- Waldner, M.; Zhang, W.; James, I.B.; Allbright, K.; Havis, E.; Bliley, J.M.; Almadori, A.; Schweizer, R.; Plock, J.A.; Washington, K.M.; et al. Characteristics and Immunomodulating Functions of Adipose-Derived and Bone Marrow-Derived Mesenchymal Stem Cells Across Defined Human Leukocyte Antigen Barriers. Front. Immunol. 2018, 9, 1642. [Google Scholar] [CrossRef] [Green Version]
- Alssum, L.; Eubank, T.D.; Roy, S.; Erdal, B.S.; Yildiz, V.O.; Tatakis, D.N.; Leblebicioglu, B. Gingival Perfusion and Tissue Biomarkers During Early Healing of Postextraction Regenerative Procedures: A Prospective Case Series. J. Periodontol. 2017, 88, 1163–1172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Xing, Y.; Jia, L.; Ji, Y.; Zhao, B.; Wen, Y.; Xu, X. An In Vitro Comparative Study of Multisource Derived Human Mesenchymal Stem Cells for Bone Tissue Engineering. Stem Cells Dev. 2018, 27, 1634–1645. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, M.E.; Emons, J.A.M.; Karperien, M.; Nauta, A.J.; Willemze, R.; Roelofs, H.; Romeo, S.; Marchini, A.; Rappold, G.A.; Vukicevic, S.; et al. Human Mesenchymal Stem Cells Derived from Bone Marrow Display a Better Chondrogenic Differentiation Compared with Other Sources. Connect. Tissue Res. 2007, 48, 132–140. [Google Scholar] [CrossRef]
- Stanko, P.; Kaiserova, K.; Altanerova, V.; Altaner, C. Comparison of Human Mesenchymal Stem Cells Derived from Dental Pulp, Bone Marrow, Adipose Tissue, and Umbilical Cord Tissue by Gene Expression. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc Czech Repub. 2014, 158, 373–377. [Google Scholar] [CrossRef] [Green Version]
- Danisovic, L.; Oravcova, L.; Krajciova, L.; Varchulova Novakova, Z.; Bohac, M.; Varga, I.; Vojtassak, J. Effect of Long-Term Culture on the Biological and Morphological Characteristics of Human Adipose Tissue-Derived Stem Cells. J. Physiol. Pharmacol. 2017, 68, 149–158. [Google Scholar] [PubMed]
- Hakki, S.S.; Turaç, G.; Bozkurt, S.B.; Kayis, S.A.; Hakki, E.E.; Şahin, E.; Subaşı, C.; Karaoz, E. Comparison of Different Sources of Mesenchymal Stem Cells: Palatal versus Lipoaspirated Adipose Tissue. Cells Tissues Organs 2017, 204, 228–240. [Google Scholar] [CrossRef]
- Stanko, P.; Altanerova, U.; Jakubechova, J.; Repiska, V.; Altaner, C. Dental Mesenchymal Stem/Stromal Cells and Their Exosomes. Stem Cells Int. 2018, 2018, 8973613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angelopoulos, I.; Brizuela, C.; Khoury, M. Gingival Mesenchymal Stem Cells Outperform Haploidentical Dental Pulp-Derived Mesenchymal Stem Cells in Proliferation Rate, Migration Ability, and Angiogenic Potential. Cell Transplant. 2018, 27, 967–978. [Google Scholar] [CrossRef] [Green Version]
- Winning, L.; El Karim, I.A.; Lundy, F.T. A Comparative Analysis of the Osteogenic Potential of Dental Mesenchymal Stem Cells. Stem Cells Dev. 2019, 28, 1050–1058. [Google Scholar] [CrossRef]
- Oh, S.-L. Attached Gingiva: Histology and Surgical Augmentation. Gen. Dent. 2009, 57, 381–385. [Google Scholar] [PubMed]
- Hatayama, T.; Nakada, A.; Nakamura, H.; Mariko, W.; Tsujimoto, G.; Nakamura, T. Regeneration of Gingival Tissue Using in Situ Tissue Engineering with Collagen Scaffold. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2017, 124, 348–354.e1. [Google Scholar] [CrossRef] [Green Version]
- Jeon, Y.-J.; Kim, J.; Cho, J.H.; Chung, H.-M.; Chae, J.-I. Comparative Analysis of Human Mesenchymal Stem Cells Derived From Bone Marrow, Placenta, and Adipose Tissue as Sources of Cell Therapy. J. Cell. Biochem. 2016, 117, 1112–1125. [Google Scholar] [CrossRef] [PubMed]
- Eramo, S.; Natali, A.; Pinna, R.; Milia, E. Dental Pulp Regeneration via Cell Homing. Int. Endod. J. 2018, 51, 405–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadlapati, M.; Biguetti, C.; Cavalla, F.; Nieves, F.; Bessey, C.; Bohluli, P.; Garlet, G.P.; Letra, A.; Fakhouri, W.D.; Silva, R.M. Characterization of a Vascular Endothelial Growth Factor-Loaded Bioresorbable Delivery System for Pulp Regeneration. J. Endod. 2017, 43, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, X.; Yu, W.; Zhang, Y.; Shi, C.; Ni, S.; Liu, Q.; Li, X.; Sun, Y.; Zheng, C.; et al. Effects of Human Vascular Endothelial Growth Factor on Reparative Dentin Formation. Mol. Med. Rep. 2016, 13, 705–712. [Google Scholar] [CrossRef] [Green Version]
- Oryan, A.; Kamali, A.; Moshiri, A.; Baghaban Eslaminejad, M. Role of Mesenchymal Stem Cells in Bone Regenerative Medicine: What Is the Evidence? Cells Tissues Organs 2017, 204, 59–83. [Google Scholar] [CrossRef] [PubMed]
- Gregory, C.A.; Gunn, W.G.; Peister, A.; Prockop, D.J. An Alizarin Red-Based Assay of Mineralization by Adherent Cells in Culture: Comparison with Cetylpyridinium Chloride Extraction. Anal. Biochem. 2004, 329, 77–84. [Google Scholar] [CrossRef]
- Puchtler, H.; Meloan, S.N.; Terry, M.S. On the History and Mechanism of Alizarin and Alizarin Red S Stains for Calcium. J. Histochem. Cytochem. 1969, 17, 110–124. [Google Scholar] [CrossRef]
- Zajdel, A.; Kałucka, M.; Kokoszka-Mikołaj, E.; Wilczok, A. Osteogenic Differentiation of Human Mesenchymal Stem Cells from Adipose Tissue and Wharton’s Jelly of the Umbilical Cord. Acta Biochim. Pol. 2017, 64, 365–369. [Google Scholar] [CrossRef]
- Adolpho, L.F.; Lopes, H.B.; Freitas, G.P.; Weffort, D.; Campos Totoli, G.G.; Loyola Barbosa, A.C.; Freire Assis, R.I.; Silverio Ruiz, K.G.; Andia, D.C.; Rosa, A.L.; et al. Human Periodontal Ligament Stem Cells with Distinct Osteogenic Potential Induce Bone Formation in Rat Calvaria Defects. Regen. Med. 2022, 17, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.I.; Correa, D. The MSC: An Injury Drugstore. Cell Stem Cell 2011, 9, 11–15. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Wu, J.; Li, D.; Hao, L.; Li, Y.; Yi, D.; Yeung, K.W.K.; Chen, D.; Lu, W.W.; Pan, H.; et al. Engineering Stem Cells to Produce Exosomes with Enhanced Bone Regeneration Effects: An Alternative Strategy for Gene Therapy. J. Nanobiotechnol. 2022, 20, 135. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, M.; Luong, J.C.; Rodriguez, S.M.; Cao, R.; Wheeler, A.E.; Lau, H.; Li, S.; Shabestari, S.K.; Chadarevian, J.P.; Alexander, M.; et al. Controlled Release of Stem Cell Secretome Attenuates Inflammatory Response against Implanted Biomaterials. Adv. Healthc. Mater. 2020, 9, e1901874. [Google Scholar] [CrossRef] [PubMed]
- Marx, C.; Gardner, S.; Harman, R.M.; Van de Walle, G.R. The Mesenchymal Stromal Cell Secretome Impairs Methicillin-Resistant Staphylococcus Aureus Biofilms via Cysteine Protease Activity in the Equine Model. Stem Cells Transl. Med. 2020, 9, 746–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsiao, S.T.-F.; Asgari, A.; Lokmic, Z.; Sinclair, R.; Dusting, G.J.; Lim, S.Y.; Dilley, R.J. Comparative Analysis of Paracrine Factor Expression in Human Adult Mesenchymal Stem Cells Derived from Bone Marrow, Adipose, and Dermal Tissue. Stem Cells Dev. 2012, 21, 2189–2203. [Google Scholar] [CrossRef] [Green Version]


| MSC Source | MSC Property | Key Findings | References |
|---|---|---|---|
| BM-MSCs | Proliferation | BM-MSCs have the potential to double in a 24–72 h period. | Riekstina et al. [85] Heo et al. [86] Zong et al. [87] |
| Migration capacity | The migration capacity of BM-MSCs and AT-MSCs is similar. | Jeon et al. [88] | |
| Clonogenicity | The CFU capacity of BM-MSCs does not remain stable, entering senescence after passage 3–4. | Dmitrieva et al. [89] | |
| The CFU of BM-MSCs is lower than that of AT-MSCs. | Hayashi et al. [90] | ||
| Angiogenic capacity | VEGF expression has increased alongside differentiation of BM-MSCs | Waldner et al. [91] | |
| Differentiation potential | BM-MSCs are able to constitutively express alkaline phosphatase (ALP) activity with no necessity of induction of differentiation. | Riekstina et al. [85] | |
| BM-MSCs extracted from human mandibles showed calcium deposition in the extracellular matrix after 7 days of osteogenic induction and mineralization nodes after 21 days of induction. | Zong et al. [87] | ||
| BM-MSCs showed higher osteogenic capacity compared to PL-MSCs and DF-MSCs. | Zhang et al. [92] | ||
| BM-MSCs carried a higher expression of osteogenic markers than subcutaneous AT-MSCs. Similarly, there were higher calcium depositions in BM-MSCs than in AT-MSC-related sources. | Waldner et al. [91] | ||
| The ability of BM-MSCs (fetal and adult) to differentiate into chondrocytes, adipocytes, and osteoblasts was found to decrease over the passages. | Bernardo et al. [93] | ||
| The osteogenic differentiation capacity of BM-MSCs decreased along the passages. | Stanko et al. [94] | ||
| AT-MSCs | Proliferation | AT-MSCs have the potential to double in a 24–48 h period. | Riekstina et al. [85] Heo et al. [86] |
| The proliferation rate of human AT-MSCs increases up to passage 10, finding a 32% reduction in proliferation at passage 30. | Danisovic et al. [95] | ||
| There is no significant difference between the proliferation rate of BM-MSCs obtained from lipoaspirate compared to PAT-MSCs. | Hakki et al. [96] | ||
| Clonogenicity | The CFU capacity of AT-MSCs remains stable along the passages. | Dmitrieva et al. [89] | |
| Angiogenic capacity | VEGF expression has increased alongside differentiation of AT-MSCs (subcutaneous and omental). | Waldner et al. [91] | |
| Differentiation potential | AT-MSCs are able to constitutively express alkaline phosphatase (ALP) activity with no necessity of induction of differentiation. | Riekstina et al. [85] | |
| ALP expression was revealed to be higher in PAT-MSCs than in MSCs derived from lipoaspiration. | Hakki et al. [96] | ||
| AT-MSCs showed higher osteogenic capacity than PL-MSCs and DF-MSCs. | Zhang et al. [92] | ||
| AT-MSCs from omental tissue had higher expression of osteogenic markers than subcutaneous AT-MSCs. | Waldner et al. [91] | ||
| The osteogenic differentiation capacity of AT-MSCs decreased along the passages. | Stanko et al. [94] | ||
| DT-MSCs | Proliferation | TP-MSCs can be expanded and maintained for almost 60 doubling populations with a high rate of proliferation. | Pandula et al. [69] |
| DT-MSCs proliferated faster than BM-MSCs and AT-MSCs | Zhang et al. [92] | ||
| DT-MSCs have a high cell proliferative potential. | Stanko et al. [97] | ||
| G-MSCs showed higher proliferation rates compared to DP-MSCs. | Angelopoulos et al. [98] | ||
| Migration capacity | The migration capacity of G-MSCs is higher than that of DP-MSCs. | Angelopoulos et al. [98] | |
| Clonogenicity | CFU was higher in G-MSCs compared to DP-MSCs. | Angelopoulos et al. [98] | |
| Angiogenic capacity | MSCs obtained from gingival tissue showed higher angiogenic capacity than cells from DP. | Angelopoulos et al. [98] | |
| Differentiation potential | The osteogenic differentiation capacity of DP-MSCs increased along the passages. | Stanko et al. [94] | |
| PL-MSCs showed increased expression of ALP, calcium deposits, and an early expression of differentiation genes (ALP and COL1A1) compared to SHEDs and DP-MSCs. | Winning et al. [99] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costela-Ruiz, V.J.; Melguizo-Rodríguez, L.; Bellotti, C.; Illescas-Montes, R.; Stanco, D.; Arciola, C.R.; Lucarelli, E. Different Sources of Mesenchymal Stem Cells for Tissue Regeneration: A Guide to Identifying the Most Favorable One in Orthopedics and Dentistry Applications. Int. J. Mol. Sci. 2022, 23, 6356. https://doi.org/10.3390/ijms23116356
Costela-Ruiz VJ, Melguizo-Rodríguez L, Bellotti C, Illescas-Montes R, Stanco D, Arciola CR, Lucarelli E. Different Sources of Mesenchymal Stem Cells for Tissue Regeneration: A Guide to Identifying the Most Favorable One in Orthopedics and Dentistry Applications. International Journal of Molecular Sciences. 2022; 23(11):6356. https://doi.org/10.3390/ijms23116356
Chicago/Turabian StyleCostela-Ruiz, Victor J., Lucía Melguizo-Rodríguez, Chiara Bellotti, Rebeca Illescas-Montes, Deborah Stanco, Carla Renata Arciola, and Enrico Lucarelli. 2022. "Different Sources of Mesenchymal Stem Cells for Tissue Regeneration: A Guide to Identifying the Most Favorable One in Orthopedics and Dentistry Applications" International Journal of Molecular Sciences 23, no. 11: 6356. https://doi.org/10.3390/ijms23116356
APA StyleCostela-Ruiz, V. J., Melguizo-Rodríguez, L., Bellotti, C., Illescas-Montes, R., Stanco, D., Arciola, C. R., & Lucarelli, E. (2022). Different Sources of Mesenchymal Stem Cells for Tissue Regeneration: A Guide to Identifying the Most Favorable One in Orthopedics and Dentistry Applications. International Journal of Molecular Sciences, 23(11), 6356. https://doi.org/10.3390/ijms23116356