
A Type Ib Crustin from Deep-Sea Shrimp Possesses Antimicrobial and Immunomodulatory Activity

Abstract
:1. Introduction
2. Results
2.1. Crus2 Is a Type 1b Crustin
2.2. Crus2 Binds and Kills Gram-Positive and Gram-Negative Bacteria in a Manner That Is Affected in Degree by the C-Terminal P58 Region
2.3. Crus2 Kills Bacteria by Inducing Damage in the Bacteria Membrane
2.4. The Bactericidal Activity of Crus2 Depends on the Conserved WAP Domain and Is Affected by P58
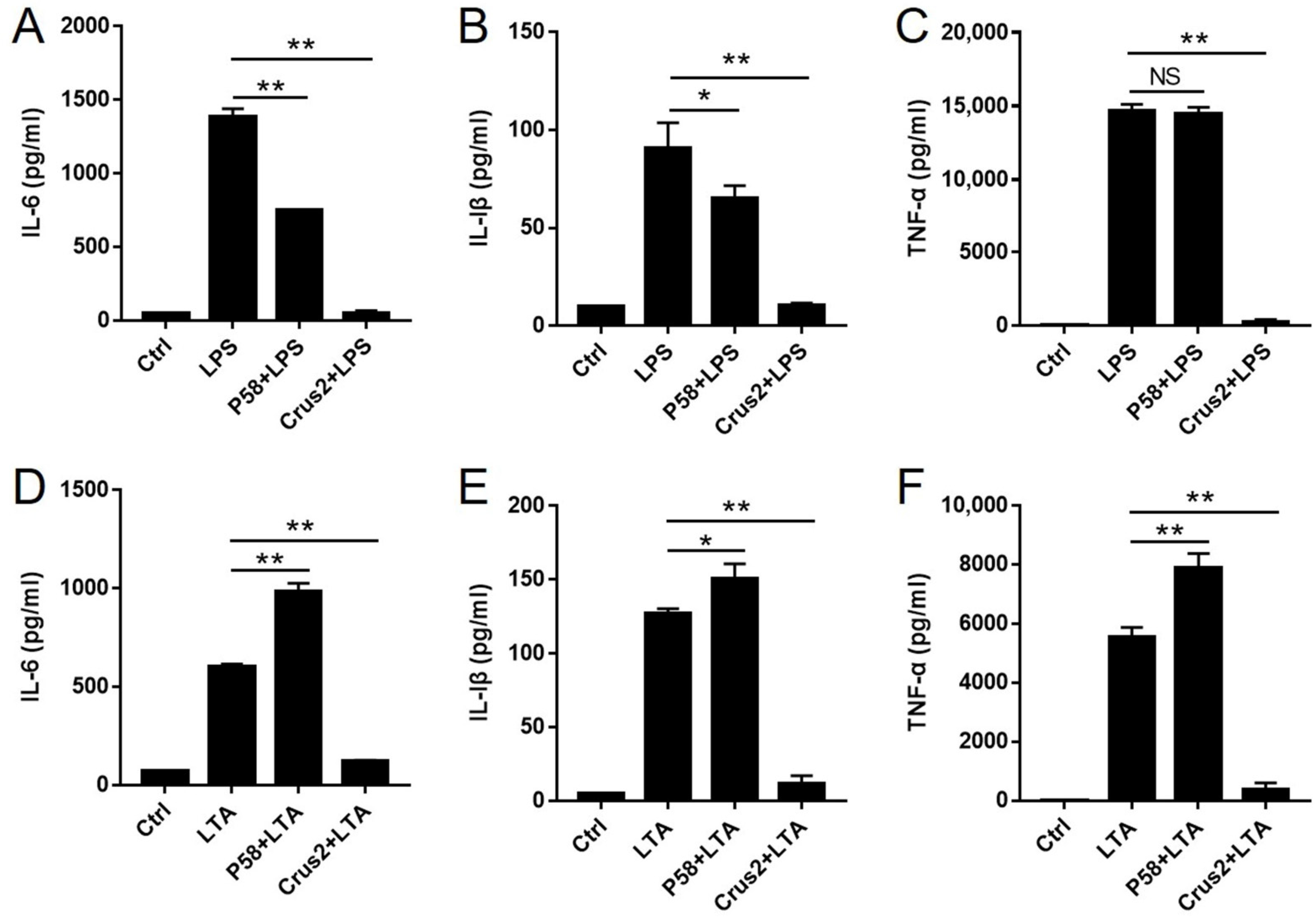
2.5. Crus2 Blocks LPS- and LTA-Induced Inflammatory Response
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
4.2. Bioinformatics and Structural Characterization
4.3. Preparation of Recombinant Proteins
4.4. Peptide Synthesis
4.5. Antibacterial Assay
4.6. Binding of Crus2 and Crus2DC to Bacteria
4.7. PI staining Assay and Electron Microscopy
4.8. Membrane Depolarization Assay
4.9. Pro-inflammatory Cytokine Detection
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Izadpanah, A.; Gallo, R.L. Antimicrobial peptides. J. Am. Acad. Dermatol. 2005, 52, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Haney, E.F.; Vogel, H.J. The expanding scope of antimicrobial peptide structures and their modes of action. Trends Biotechnol. 2011, 29, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Papo, N.; Shai, Y. Can we predict biological activity of antimicrobial peptides from their interactions with model phospholipid membranes? Peptides 2003, 24, 1693–1703. [Google Scholar] [CrossRef] [PubMed]
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Hadley, E.B.; Hancock, R.E. Strategies for the discovery and advancement of novel cationic antimicrobial peptides. Curr. Top. Med. Chem. 2010, 10, 1872–1881. [Google Scholar] [CrossRef]
- Omardien, S.; Brul, S.; Zaat, S.A. Antimicrobial Activity of Cationic Antimicrobial Peptides against Gram-Positives: Current Progress Made in Understanding the Mode of Action and the Response of Bacteria. Front. Cell Dev. Biol. 2016, 4, 111. [Google Scholar] [CrossRef]
- Tennessen, J.A. Molecular evolution of animal antimicrobial peptides: Widespread moderate positive selection. J. Evol. Biol. 2005, 18, 1387–1394. [Google Scholar] [CrossRef]
- Bandeira, P.T.; Vernal, J.; Matos, G.M.; Farias, N.D.; Terenzi, H.; Pinto, A.R.; Barracco, M.A.; Rosa, R.D. A Type IIa crustin from the pink shrimp Farfantepenaeus paulensis (crusFpau) is constitutively synthesized and stored by specific granule-containing hemocyte subpopulations. Fish Shellfish. Immunol. 2020, 97, 294–299. [Google Scholar] [CrossRef]
- Smith, V.J.; Fernandes, J.M.; Kemp, G.D.; Hauton, C. Crustins: Enigmatic WAP domain-containing antibacterial proteins from crustaceans. Dev. Comp. Immunol. 2008, 32, 758–772. [Google Scholar] [CrossRef] [Green Version]
- Ranganathan, S.; Simpson, K.J.; Shaw, D.C.; Nicholas, K.R. The whey acidic protein family: A new signature motif and three-dimensional structure by comparative modeling. J. Mol. Graph. Model. 1999, 17, 106–113. [Google Scholar] [CrossRef]
- Fjell, C.D.; Hiss, J.A.; Hancock, R.E.; Schneider, G. Designing antimicrobial peptides: Form follows function. Nat. Rev. Drug Discov. 2011, 11, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.F.; Wang, J.X. The antimicrobial peptides of the immune response of shrimp. Invertebr. Surviv. J. 2008, 5, 4. [Google Scholar]
- Krusong, K.; Poolpipat, P.; Supungul, P.; Tassanakajon, A. A comparative study of antimicrobial properties of crustinPm1 and crustinPm7 from the black tiger shrimp Penaeus monodon. Dev. Comp. Immunol. 2012, 36, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Amparyup, P.; Kondo, H.; Hirono, I.; Aoki, T.; Tassanakajon, A. Molecular cloning, genomic organization and recombinant expression of a crustin-like antimicrobial peptide from black tiger shrimp Penaeus monodon. Mol. Immunol. 2008, 45, 1085–1093. [Google Scholar] [CrossRef]
- Arockiaraj, J.; Gnanam, A.J.; Muthukrishnan, D.; Gudimella, R.; Milton, J.; Singh, A.; Muthupandian, S.; Kasi, M.; Bhassu, S. Crustin, a WAP domain containing antimicrobial peptide from freshwater prawn Macrobrachium rosenbergii: Immune characterization. Fish Shellfish. Immunol. 2013, 34, 109–118. [Google Scholar] [CrossRef]
- Tassanakajon, A.; Somboonwiwat, K.; Amparyup, P. Sequence diversity and evolution of antimicrobial peptides in invertebrates. Dev. Comp. Immunol. 2015, 48, 324–341. [Google Scholar] [CrossRef]
- Li, S.; Lv, X.; Yu, Y.; Zhang, X.; Li, F. Molecular and Functional Diversity of Crustin-Like Genes in the Shrimp Litopenaeus vannamei. Mar. Drugs 2020, 18, 361. [Google Scholar] [CrossRef]
- Thornburg, C.C.; Zabriskie, T.M.; McPhail, K.L. Deep-sea hydrothermal vents: Potential hot spots for natural products discovery? J. Nat. Prod. 2010, 73, 489–499. [Google Scholar] [CrossRef]
- Zhang, J.; Sun, Q.L.; Luan, Z.D.; Lian, C.; Sun, L. Comparative transcriptome analysis of Rimicaris sp. reveals novel molecular features associated with survival in deep-sea hydrothermal vent. Sci. Rep. 2017, 7, 2000. [Google Scholar] [CrossRef]
- Gu, H.J.; Sun, Q.L.; Jiang, S.; Zhang, J.; Sun, L. First characterization of an anti-lipopolysaccharide factor (ALF) from hydrothermal vent shrimp: Insights into the immune function of deep-sea crustacean ALF. Dev. Comp. Immunol. 2018, 84, 382–395. [Google Scholar] [CrossRef]
- Luo, J.C.; Zhang, J.; Sun, L. A g-Type Lysozyme from Deep-Sea Hydrothermal Vent Shrimp Kills Selectively Gram-Negative Bacteria. Molecules 2021, 26, 7624. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, J.; Sun, Y.; Sun, L. A Crustin from Hydrothermal Vent Shrimp: Antimicrobial Activity and Mechanism. Mar. Drugs 2021, 19, 176. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Kim, M.; Kim, A.R.; Yi, M.; Choi, J.H.; Park, H.; Park, W.; Kim, H.W. Differences in gene organization between type I and type II crustins in the morotoge shrimp, Pandalopsis japonica. Fish Shellfish. Immunol. 2013, 35, 1176–1184. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.L.; Wang, S.L.; Zhu, F.C.; Xue, F.; He, L.S. Characteristics of Two Crustins from Alvinocaris longirostris in Hydrothermal Vents. Mar. Drugs 2021, 19, 600. [Google Scholar] [CrossRef] [PubMed]
- Le Bloa, S.; Boidin-Wichlacz, C.; Cueff-Gauchard, V.; Rosa, R.D.; Cuvillier-Hot, V.; Durand, L.; Methou, P.; Pradillon, F.; Cambon-Bonavita, M.A.; Tasiemski, A. Antimicrobial Peptides and Ectosymbiotic Relationships: Involvement of a Novel Type IIa Crustin in the Life Cycle of a Deep-Sea Vent Shrimp. Front. Immunol. 2020, 11, 1511. [Google Scholar] [CrossRef]
- Donpudsa, S.; Rimphanitchayakit, V.; Tassanakajon, A.; Söderhäll, I.; Söderhäll, K. Characterization of two crustin antimicrobial peptides from the freshwater crayfish Pacifastacus leniusculus. J. Invertebr. Pathol. 2010, 104, 234–238. [Google Scholar] [CrossRef]
- Supungul, P.; Tang, S.; Maneeruttanarungroj, C.; Rimphanitchayakit, V.; Hirono, I.; Aoki, T.; Tassanakajon, A. Cloning, expression and antimicrobial activity of crustinPm1, a major isoform of crustin, from the black tiger shrimp Penaeus monodon. Dev. Comp. Immunol. 2008, 32, 61–70. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, C.; Wang, H.; Ma, H.; Huang, Y.Q.; Lu, J.X.; Li, X.C.; Zhang, X.W. Involvement of a newly identified atypical type II crustin (SpCrus5) in the antibacterial immunity of mud crab Scylla paramamosain. Fish Shellfish. Immunol. 2018, 75, 346–356. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, J.X.; Wang, Y.; Fang, W.H.; Wang, Y.; Zhou, J.F.; Zhao, S.; Li, X.C. Newly identified type II crustin (SpCrus2) in Scylla paramamosain contains a distinct cysteine distribution pattern exhibiting broad antimicrobial activity. Dev. Comp. Immunol. 2018, 84, 1–13. [Google Scholar] [CrossRef]
- Scocchi, M.; Mardirossian, M.; Runti, G.; Benincasa, M. Non-Membrane Permeabilizing Modes of Action of Antimicrobial Peptides on Bacteria. Curr. Top. Med. Chem. 2016, 16, 76–88. [Google Scholar] [CrossRef]
- Malanovic, N.; Lohner, K. Antimicrobial Peptides Targeting Gram-Positive Bacteria. Pharmaceuticals 2016, 9, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Y.P.; Sun, Y.D.; Wang, Z.H.; Wang, Q.; Wang, X.W.; Zhao, X.F.; Wang, J.X. A single whey acidic protein domain (SWD)-containing peptide from fleshy prawn with antimicrobial and proteinase inhibitory activities. Aquaculture 2008, 284, 246–259. [Google Scholar] [CrossRef]
- van der Lee, R.; Buljan, M.; Lang, B.; Weatheritt, R.J.; Daughdrill, G.W.; Dunker, A.K.; Fuxreiter, M.; Gough, J.; Gsponer, J.; Jones, D.T.; et al. Classification of intrinsically disordered regions and proteins. Chem. Rev. 2014, 114, 6589–6631. [Google Scholar] [CrossRef] [PubMed]
- Dunker, A.K.; Babu, M.M.; Barbar, E.; Blackledge, M.; Bondos, S.E.; Dosztányi, Z.; Dyson, H.J.; Forman-Kay, J.; Fuxreiter, M.; Gsponer, J.; et al. What’s in a name? Why these proteins are intrinsically disordered: Why these proteins are intrinsically disordered. Intrinsically Disord. Proteins 2013, 1, e24157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.J.; Gallo, R.L. Antimicrobial peptides. Curr. Biol. CB 2016, 26, R14–R19. [Google Scholar] [CrossRef]
- Wu, S.H.; Lin, H.J.; Lin, W.F.; Wu, J.L.; Gong, H.Y. A potent tilapia secreted granulin peptide enhances the survival of transgenic zebrafish infected by Vibrio vulnificus via modulation of innate immunity. Fish Shellfish. Immunol. 2018, 75, 74–90. [Google Scholar] [CrossRef]
- Su, B.C.; Chen, J.Y. Epinecidin-1: An orange-spotted grouper antimicrobial peptide that modulates Staphylococcus aureus lipoteichoic acid-induced inflammation in macrophage cells. Fish Shellfish. Immunol. 2020, 99, 362–367. [Google Scholar] [CrossRef]
- Essig, A.; Hofmann, D.; Münch, D.; Gayathri, S.; Künzler, M.; Kallio, P.T.; Sahl, H.G.; Wider, G.; Schneider, T.; Aebi, M. Copsin, a novel peptide-based fungal antibiotic interfering with the peptidoglycan synthesis. J. Biol. Chem. 2014, 289, 34953–34964. [Google Scholar] [CrossRef] [Green Version]
- Bellemare, A.; Vernoux, N.; Morin, S.; Gagné, S.M.; Bourbonnais, Y. Structural and antimicrobial properties of human pre-elafin/trappin-2 and derived peptides against Pseudomonas aeruginosa. BMC Microbiol. 2010, 10, 253. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria | MIC (μm) | MBC (μm) | ||
|---|---|---|---|---|
| Crus2 | Crus2DC | Crus2 | Crus2DC | |
| Gram-positive bacteria | ||||
| Bacillus subtilis G7 | 16 | 32 | 32 | 64 |
| Bacillus cereus MB1 | 16 | 32 | 32 | 64 |
| Bacillus cereus H2 | 32 | 16 | 32 | 32 |
| Bacillus wiedmannii SR52 | 16 | 32 | 16 | 32 |
| Micrococcus luteus | 16 | 16 | 32 | 16 |
| Staphylococcus aureus | 16 | 16 | 32 | 32 |
| Streptococcus iniae | 32 | 32 | 32 | 32 |
| Gram-negative bacteria | ||||
| Vibrio harveyi | 16 | 32 | 16 | 32 |
| Edwardsiella tarda TX1 | 32 | 32 | 32 | 32 |
| Vibrio anguillarum | 32 | 32 | — | — |
| Escherichia coli | — | — | — | — |
| Pseudomonas fluorescens | — | — | — | — |
| Marinobacter algicola | — | — | — | — |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.-J.; Sun, L. A Type Ib Crustin from Deep-Sea Shrimp Possesses Antimicrobial and Immunomodulatory Activity. Int. J. Mol. Sci. 2022, 23, 6444. https://doi.org/10.3390/ijms23126444
Wang Y-J, Sun L. A Type Ib Crustin from Deep-Sea Shrimp Possesses Antimicrobial and Immunomodulatory Activity. International Journal of Molecular Sciences. 2022; 23(12):6444. https://doi.org/10.3390/ijms23126444
Chicago/Turabian StyleWang, Yu-Jian, and Li Sun. 2022. "A Type Ib Crustin from Deep-Sea Shrimp Possesses Antimicrobial and Immunomodulatory Activity" International Journal of Molecular Sciences 23, no. 12: 6444. https://doi.org/10.3390/ijms23126444
APA StyleWang, Y. -J., & Sun, L. (2022). A Type Ib Crustin from Deep-Sea Shrimp Possesses Antimicrobial and Immunomodulatory Activity. International Journal of Molecular Sciences, 23(12), 6444. https://doi.org/10.3390/ijms23126444