
Overexpression of OsGF14f Enhances Quantitative Leaf Blast and Bacterial Blight Resistance in Rice

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Expression Patterns of OsGF14f and Subcellular Localization of OsGF14f
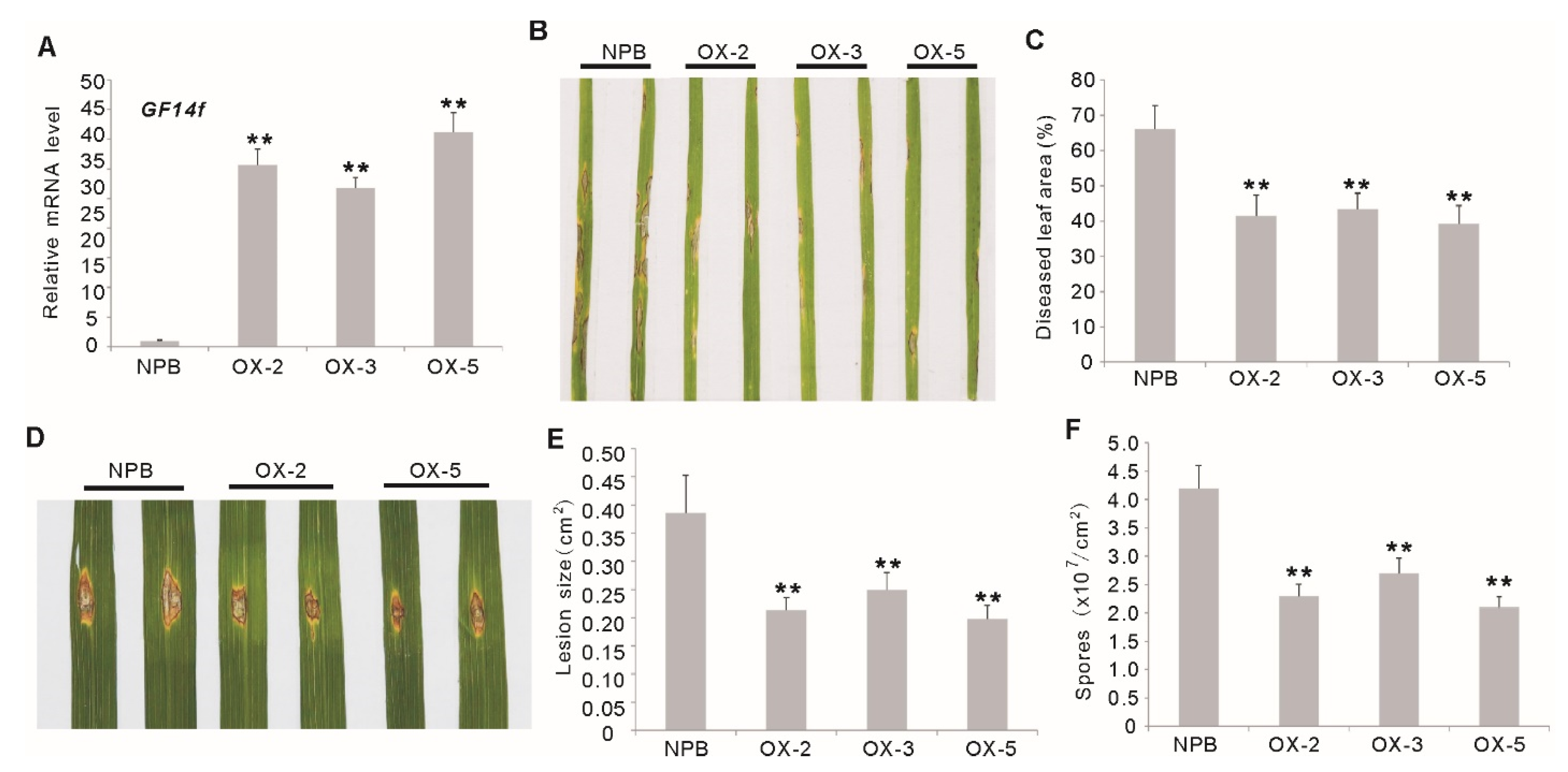
2.2. Overexpression of OsGF14f Enhances Resistance to Leaf Blast in Rice
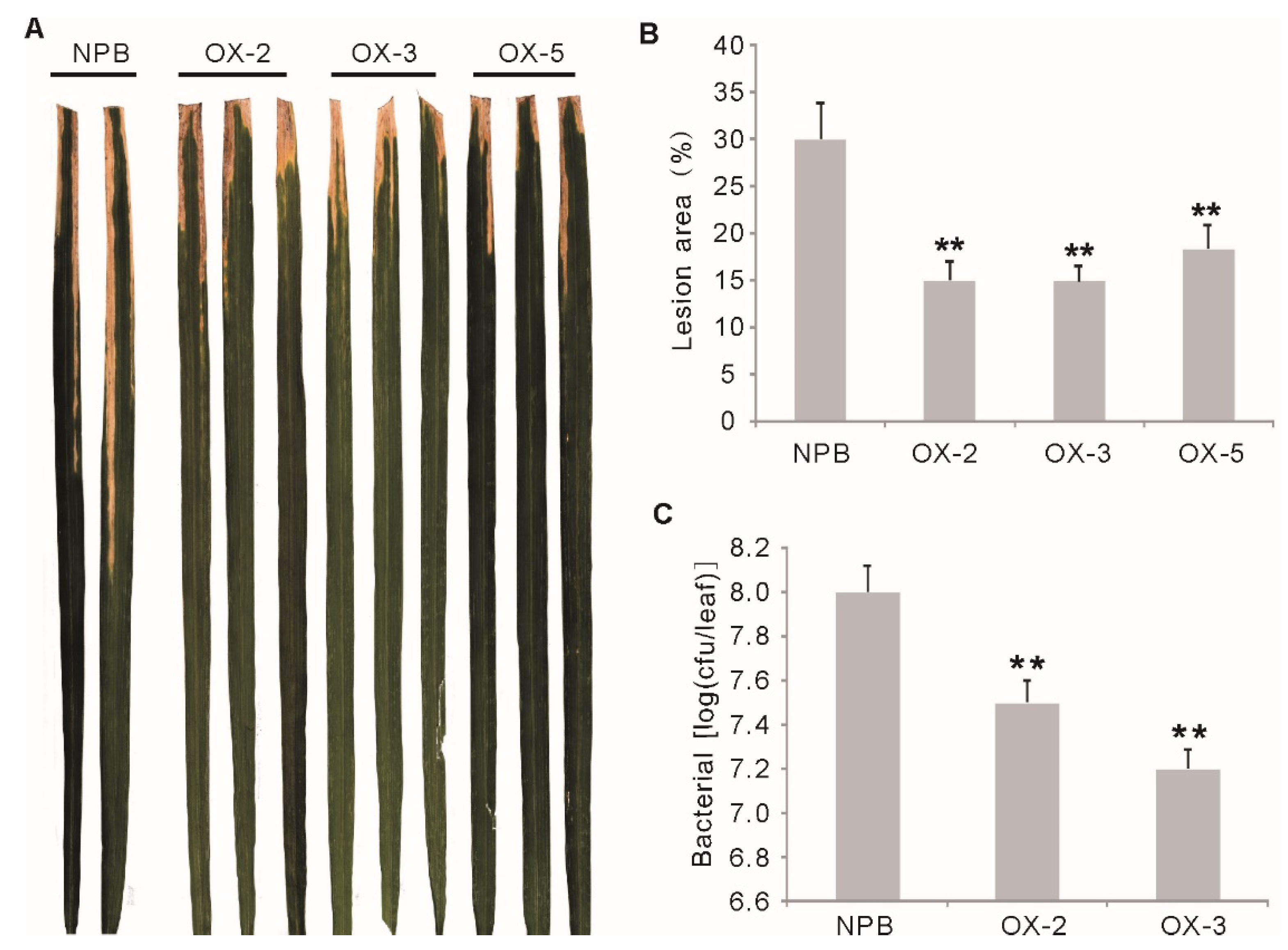
2.3. Overexpression of OsGF14f Enhances Resistance to Bacterial Blight in Rice
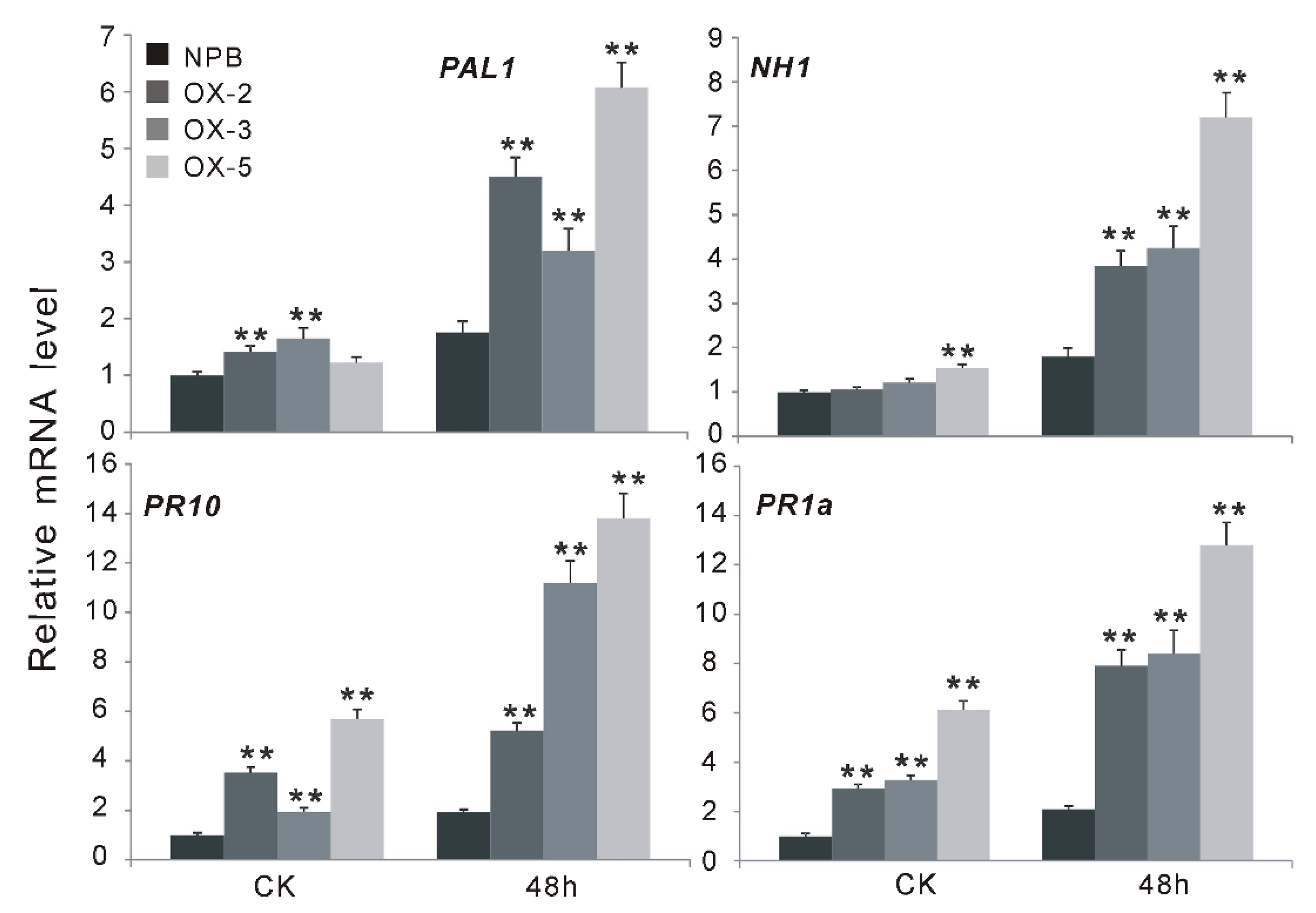
2.4. OsGF14f Positively Regulates the SA-Dependent Pathway Instead of JA Pathway
2.5. OsGF14f Is Induced by SA Treatment and Overexpression of OsGF14f Can Increase Endogenous SA Levels
3. Discussion
3.1. OsGF14f Enhances Blast and Bacterial Blight Resistance in Rice in a Quantitative Manner
3.2. OsGF14f-Mediated Disease Resistance Is Involved in the SA-Dependent Pathway
4. Materials and Methods
4.1. Plant Materials and Pathogens
4.2. Pathogen Inoculation and Evaluation of Disease Resistance
4.3. Plasmid Construction and Rice Transformation
4.4. Subcellular Localization
4.5. Gene Expression Analysis
4.6. SA Treatment and Measurement
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sharma, R.; De Vleesschauwer, D.; Sharma, M.K.; Ronald, P.C. Recent Advances in Dissecting Stress-Regulatory Crosstalk in Rice. Mol. Plant 2013, 6, 250–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinozaki, K.; Yamaguchi-Shinozaki, K.; Seki, M. Regulatory network of gene expression in the drought and cold stress responses. Curr. Opin. Plant Biol. 2003, 6, 410–417. [Google Scholar] [CrossRef]
- Capel, J.; Leyva, A.; Salinas, J. Two related low-temperature-inducible genes of Arabidopsis encode proteins showing high homology to 14-3-3 proteins, a family of putative kinase regulators. Plant Mol. Biol. 1994, 25, 693–704. [Google Scholar] [CrossRef]
- Chen, F.; Li, Q.; Sun, L.; He, Z. The Rice 14-3-3 Gene Family and its Involvement in Responses to Biotic and Abiotic Stress. DNA Res. 2006, 13, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Rosenquist, M.; Sehnke, P.; Ferl, R.J.; Sommarin, M.; Larsson, C. Evolution of the 14-3-3 Protein Family: Does the Large Number of Isoforms in Multicellular Organisms Reflect Functional Specificity? J. Mol. Evol. 2000, 51, 446–458. [Google Scholar] [CrossRef]
- Roberts, M.; Bowles, D.J. Fusicoccin, 14-3-3 Proteins, and Defense Responses in Tomato Plants1. Plant Physiol. 1999, 119, 1243–1250. [Google Scholar] [CrossRef] [Green Version]
- Cooper, B.; Clarke, J.D.; Budworth, P.; Kreps, J.; Hutchison, D.; Park, S.; Guimil, S.; Dunn, M.; Luginbühl, P.; Ellero, C.; et al. A network of rice genes associated with stress response and seed development. Proc. Natl. Acad. Sci. USA 2003, 100, 4945–4950. [Google Scholar] [CrossRef] [Green Version]
- Oh, C.-S.; Pedley, K.F.; Martin, G.B. Tomato 14-3-3 Protein 7 Positively Regulates Immunity-Associated Programmed Cell Death by Enhancing Protein Abundance and Signaling Ability of MAPKKK α. Plant Cell 2010, 22, 260–272. [Google Scholar] [CrossRef] [Green Version]
- Manosalva, P.M.; Bruce, M.; Leach, J.E. Rice 14-3-3 protein (GF14e) negatively affects cell death and disease resistance. Plant J. 2011, 68, 777–787. [Google Scholar] [CrossRef]
- Teper, D.; Salomon, D.; Sunitha, S.; Kim, J.-G.; Mudgett, M.B.; Sessa, G. Xanthomonas euvesicatoria type III effector XopQ interacts with tomato and pepper 14-3-3 isoforms to suppress effector-triggered immunity. Plant J. 2013, 77, 297–309. [Google Scholar] [CrossRef]
- Liu, Q.; Zhang, S.; Liu, B. 14-3-3 proteins: Macro-regulators with great potential for improving abiotic stress tolerance in plants. Biochem. Biophys. Res. Commun. 2016, 477, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Seehaus, K.; Tenhaken, R. Cloning of genes by mRNA differential display induced during the hypersensitive reaction of soybean after inoculation with Pseudomonas syringae pv. glycinea. Plant Mol. Biol. 1998, 38, 1225–1234. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wang, W.-M.; Coleman, M.; Orgil, U.; Feng, J.; Ma, X.; Ferl, R.; Turner, J.G.; Xiao, S. Arabidopsis 14-3-3 lambda is a positive regulator of RPW8-mediated disease resistance. Plant J. 2009, 60, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Denison, F.C.; Paul, A.-L.; Zupanska, A.K.; Ferl, R.J. 14-3-3 proteins in plant physiology. Semin. Cell Dev. Biol. 2011, 22, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Elmayan, T.; Fromentin, J.; Riondet, C.; Alcaraz, G.; Blein, J.-P.; Simon-Plas, F. Regulation of reactive oxygen species production by a 14-3-3 protein in elicited tobacco cells. Plant Cell Environ. 2007, 30, 722–732. [Google Scholar] [CrossRef]
- Oh, C.-S.; Martin, G.B. Tomato 14-3-3 Protein TFT7 Interacts with a MAP Kinase Kinase to Regulate Immunity-associated Programmed Cell Death Mediated by Diverse Disease Resistance Proteins. J. Biol. Chem. 2011, 286, 14129–14136. [Google Scholar] [CrossRef] [Green Version]
- Taylor, K.W.; Kim, J.-G.; Su, X.B.; Aakre, C.D.; Roden, J.A.; Adams, C.M.; Mudgett, M.B. Tomato TFT1 Is Required for PAMP-Triggered Immunity and Mutations that Prevent T3S Effector XopN from Binding to TFT1 Attenuate Xanthomonas Virulence. PLOS Pathog. 2012, 8, e1002768. [Google Scholar] [CrossRef] [Green Version]
- Lozano-Durán, R.; Robatzek, S. 14-3-3 Proteins in Plant-Pathogen Interactions. Mol. Plant-Microbe Interact. 2015, 28, 511–518. [Google Scholar] [CrossRef] [Green Version]
- Ashkani, S.; Rafii, M.Y.; Shabanimofrad, M.; Miah, G.; Sahebi, M.; Azizi, P.; Tanweer, F.A.; Akhtar, M.S.; Nasehi, A. Molecular Breeding Strategy and Challenges Towards Improvement of Blast Disease Resistance in Rice Crop. Front. Plant Sci. 2015, 6, 886. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Yang, J.; Zhang, S.; Zhao, J.; Feng, A.; Yang, T.; Wang, X.; Mao, X.; Dong, J.; Zhu, X.; et al. OsGF14e positively regulates panicle blast resistance in rice. Biochem. Biophys. Res. Commun. 2016, 471, 247–252. [Google Scholar] [CrossRef]
- Liu, Q.; Yang, J.; Zhang, S.; Zhao, J.; Feng, A.; Yang, T.; Wang, X.; Mao, X.; Dong, J.; Zhu, X.; et al. OsGF14b Positively Regulates Panicle Blast Resistance but Negatively Regulates Leaf Blast Resistance in Rice. Mol. Plant-Microbe Interact. 2016, 29, 46–56. [Google Scholar] [CrossRef] [Green Version]
- Yan, S.; Liu, Q.; Naake, T.; Huang, W.; Chen, M.; Kong, Q.; Zhang, S.; Li, W.; Li, X.; Liu, Q.; et al. OsGF14b modulates defense signaling pathways in rice panicle blast response. Crop J. 2020, 9, 725–738. [Google Scholar] [CrossRef]
- Hu, K.-M.; Qiu, D.-Y.; Shen, X.-L.; Li, X.-H.; Wang, S.-P. Isolation and Manipulation of Quantitative Trait Loci for Disease Resistance in Rice Using a Candidate Gene Approach. Mol. Plant 2008, 1, 786–793. [Google Scholar] [CrossRef] [Green Version]
- Thaler, J.S.; Humphrey, P.T.; Whiteman, N.K. Evolution of jasmonate and salicylate signal crosstalk. Trends Plant Sci. 2012, 17, 260–270. [Google Scholar] [CrossRef]
- Deng, H.; Liu, H.; Li, X.; Xiao, J.; Wang, S. A CCCH-Type Zinc Finger Nucleic Acid-Binding Protein Quantitatively Confers Resistance against Rice Bacterial Blight Disease. Plant Physiol. 2011, 158, 876–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kou, Y.; Wang, S. Broad-spectrum and durability: Understanding of quantitative disease resistance. Curr. Opin. Plant Biol. 2010, 13, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Liu, H.; Li, Y.; Yu, H.; Li, X.; Xiao, J.; Wang, S. Manipulating Broad-Spectrum Disease Resistance by Suppressing Pathogen-Induced Auxin Accumulation in Rice. Plant Physiol. 2011, 155, 589–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, B.A.; Linde, C. Pathogen population genetics, evolutionary potential, and durable resistance. Annu. Rev. Phytopathol. 2002, 40, 349–379. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, N.; Inoue, H.; Kato, T.; Funao, T.; Shirota, M.; Shimizu, T.; Kanamori, H.; Yamane, H.; Hayano-Saito, Y.; Matsumoto, T.; et al. Durable panicle blast-resistance gene Pb1 encodes an atypical CC-NBS-LRR protein and was generated by acquiring a promoter through local genome duplication. Plant J. 2010, 64, 498–510. [Google Scholar] [CrossRef]
- De Vleesschauwer, D.; Gheysen, G.; Höfte, M. Hormone defense networking in rice: Tales from a different world. Trends Plant Sci. 2013, 18, 555–565. [Google Scholar] [CrossRef]
- Bari, R.; Jones, J.D.G. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2008, 69, 473–488. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bioassays with Tobacco Tissue Cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Liu, Q.; Yan, S.; Huang, W.; Yang, J.; Dong, J.; Zhang, S.; Zhao, J.; Yang, T.; Mao, X.; Zhu, X.; et al. NAC transcription factor ONAC066 positively regulates disease resistance by suppressing the ABA signaling pathway in rice. Plant Mol. Biol. 2018, 98, 289–302. [Google Scholar] [CrossRef] [PubMed]
- Ono, E.; Wong, H.L.; Kawasaki, T.; Hasegawa, M.; Kodama, O.; Shimamoto, K. Essential role of the small GTPase Rac in disease resistance of rice. Proc. Natl. Acad. Sci. USA 2001, 98, 759–764. [Google Scholar] [CrossRef]
- Ding, B.; Bellizzi, M.D.R.; Ning, Y.; Meyers, B.; Wang, G.-L. HDT701, a Histone H4 Deacetylase, Negatively Regulates Plant Innate Immunity by Modulating Histone H4 Acetylation of Defense-Related Genes in Rice. Plant Cell 2012, 24, 3783–3794. [Google Scholar] [CrossRef] [Green Version]
- Kauffman, H.E.; Reddy, A.P.K.; Hsieh, S.P.Y.; Merca, S.D.; U.S. Agricultural Research Service Plant Science Research Division. An improved technique for evaluating resistance of rice varieties to Xanthomonas oryzae. Plant Dis. Rep. 1973, 57, 537–541. [Google Scholar]
- Toki, S.; Hara, N.; Ono, K.; Onodera, H.; Tagiri, A.; Oka, S.; Tanaka, H. Early infection of scutellum tissue with Agrobacterium allows high-speed transformation of rice. Plant J. 2006, 47, 969–976. [Google Scholar] [CrossRef]
- Yang, C.; Ma, Y.; He, Y.; Tian, Z.; Li, J. OsOFP19 modulates plant architecture by integrating the cell division pattern and brassinosteroid signaling. Plant J. 2017, 93, 489–501. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Welti, R.; Wang, X. Quantitative analysis of major plant hormones in crude plant extracts by high-performance liquid chromatography–mass spectrometry. Nat. Protoc. 2010, 5, 986–992. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Y.; Yang, J.; Dong, J.; Zhang, S.; Yang, W.; Zhao, J.; Yang, T.; Chen, L.; Zhou, L.; Wang, J.; et al. Overexpression of OsGF14f Enhances Quantitative Leaf Blast and Bacterial Blight Resistance in Rice. Int. J. Mol. Sci. 2022, 23, 7440. https://doi.org/10.3390/ijms23137440
Ma Y, Yang J, Dong J, Zhang S, Yang W, Zhao J, Yang T, Chen L, Zhou L, Wang J, et al. Overexpression of OsGF14f Enhances Quantitative Leaf Blast and Bacterial Blight Resistance in Rice. International Journal of Molecular Sciences. 2022; 23(13):7440. https://doi.org/10.3390/ijms23137440
Chicago/Turabian StyleMa, Yamei, Jianyuan Yang, Jingfang Dong, Shaohong Zhang, Wu Yang, Junliang Zhao, Tifeng Yang, Luo Chen, Lian Zhou, Jian Wang, and et al. 2022. "Overexpression of OsGF14f Enhances Quantitative Leaf Blast and Bacterial Blight Resistance in Rice" International Journal of Molecular Sciences 23, no. 13: 7440. https://doi.org/10.3390/ijms23137440
APA StyleMa, Y., Yang, J., Dong, J., Zhang, S., Yang, W., Zhao, J., Yang, T., Chen, L., Zhou, L., Wang, J., Chen, J., Li, W., Wu, W., Liu, Q., & Liu, B. (2022). Overexpression of OsGF14f Enhances Quantitative Leaf Blast and Bacterial Blight Resistance in Rice. International Journal of Molecular Sciences, 23(13), 7440. https://doi.org/10.3390/ijms23137440