
Molecular Mapping of Antifungal Mechanisms Accessing Biomaterials and New Agents to Target Oral Candidiasis

 , ,
, , 
 , and
, and 
Abstract
:1. Introduction
2. Oral Health and Candidiasis Development
3. Antifungal Therapy and Multidrug Resistance
4. Biomaterials and Nanotechnological Approaches
4.1. Nanocolloids for Old Molecules, but with Enhanced Antifungal Activity
4.1.1. Microemulsions
4.1.2. Nanoemulsions
4.1.3. Nanosuspensions
4.1.4. Nanostructured Lipid Carriers (NSLc)
4.2. Inorganic and Organic Nanoparticles
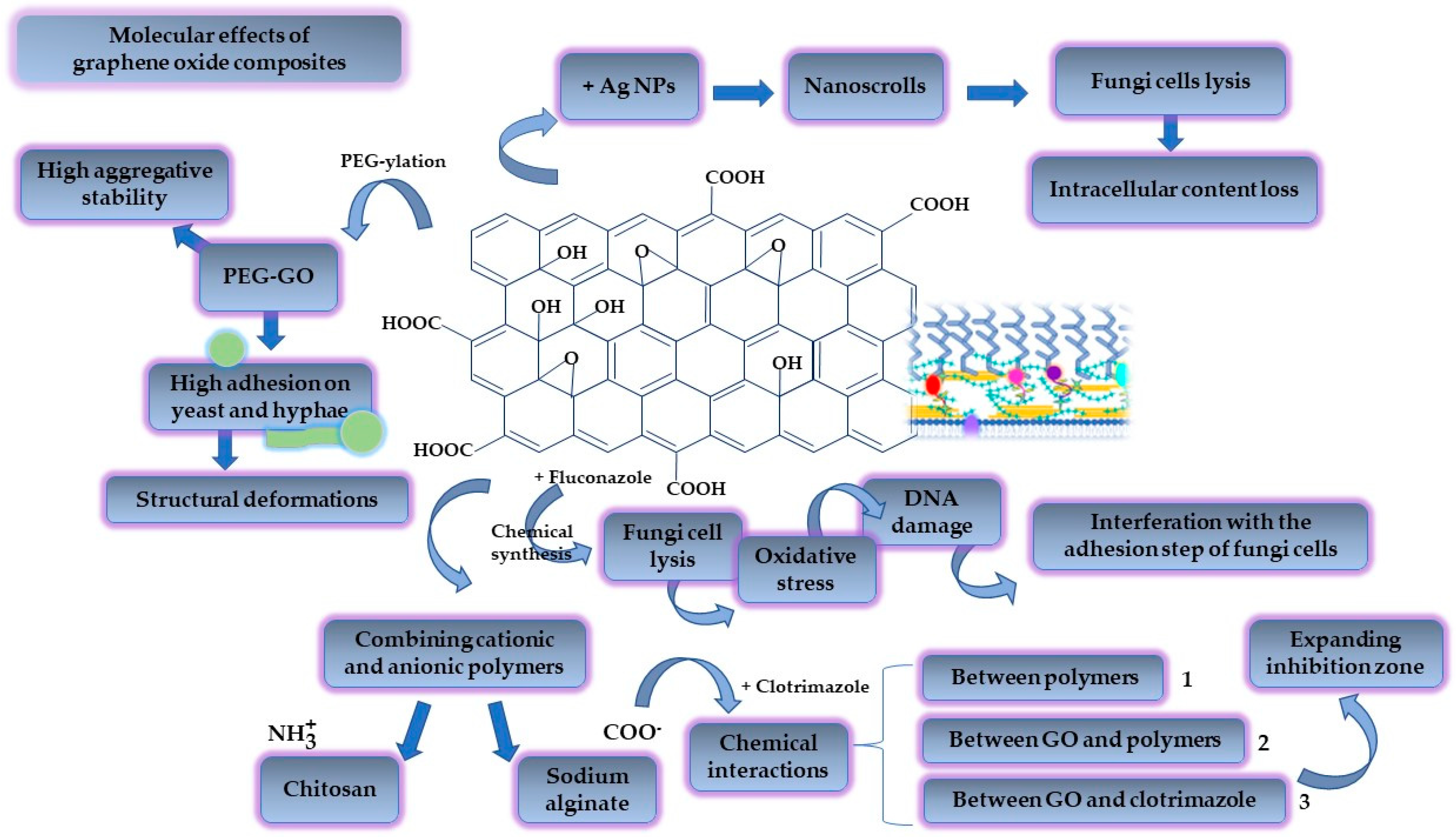
4.3. Graphene-Oxide-Based Biomaterials
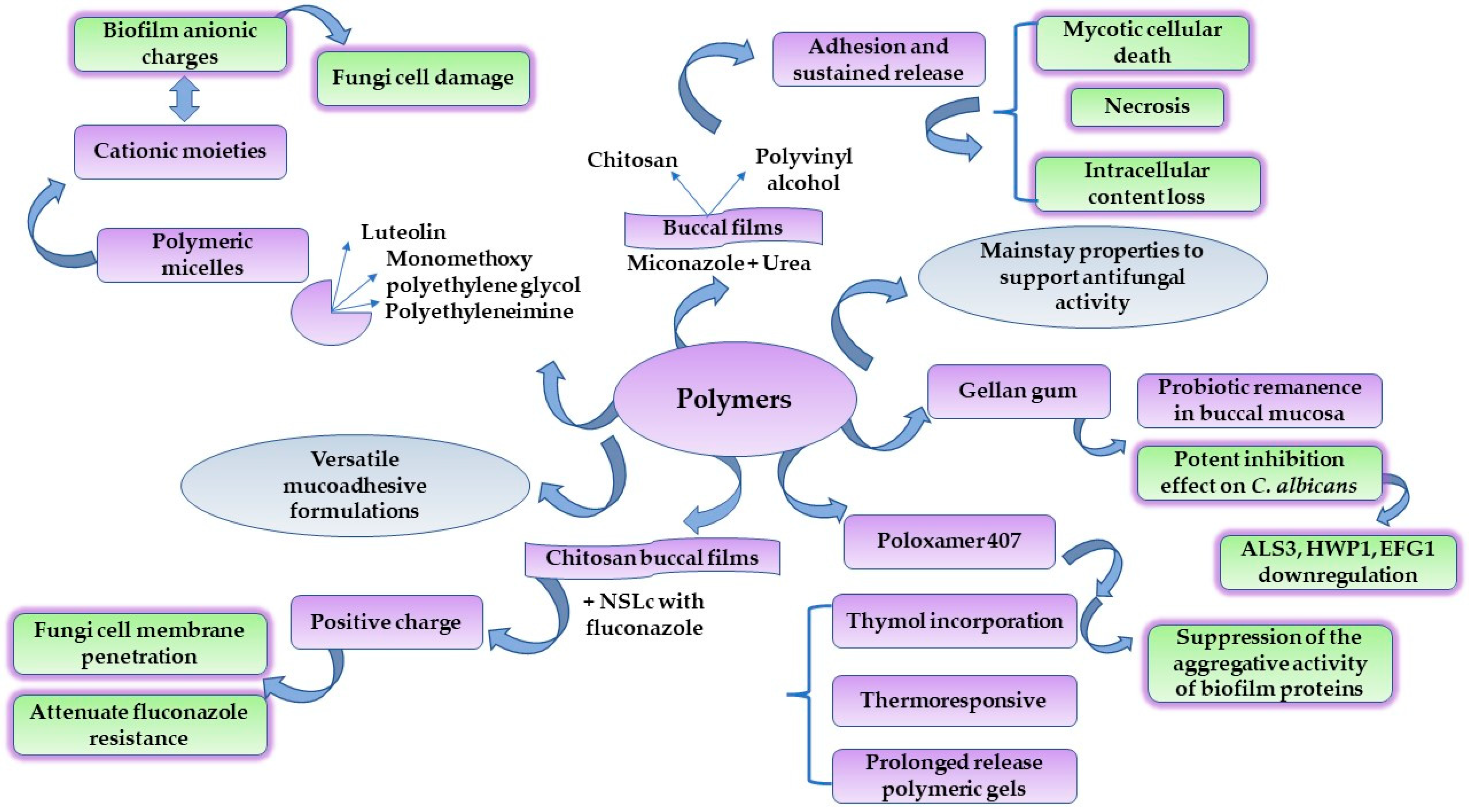
4.4. Polymers for Tailored DDS with Antifungal Activity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Oral Health. Available online: https://www.who.int/news-room/fact-sheets/detail/oral-health (accessed on 2 June 2022).
- Hujoel, P.P.; Lingström, P. Nutrition, dental caries and periodontal disease: A narrative review. J. Clin. Periodontol. 2017, 44 (Suppl. 1), S79–S84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhadse, P.; Gattani, D.; Mishra, R. The link between periodontal disease and cardiovascular disease: How far we have come in last two decades? J. Indian Soc. Periodontol. 2010, 14, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Onose, G.; Anghelescu, A.; Blendea, D.; Ciobanu, V.; Daia, C.; Firan, F.C.; Oprea, M.; Spinu, A.; Popescu, C.; Ionescu, A.; et al. Cellular and Molecular Targets for Non-Invasive, Non-Pharmacological Therapeutic/Rehabilitative Interventions in Acute Ischemic Stroke. Int. J. Mol. Sci. 2022, 23, 907. [Google Scholar] [CrossRef]
- Busnatu, S.-S.; Salmen, T.; Pana, M.-A.; Rizzo, M.; Stallone, T.; Papanas, N.; Popovic, D.; Tanasescu, D.; Serban, D.; Stoian, A.P. The Role of Fructose as a Cardiovascular Risk Factor: An Update. Metabolites 2022, 12, 67. [Google Scholar] [CrossRef] [PubMed]
- Irani, S.; Barati, I.; Badiei, M. Periodontitis and oral cancer—Current concepts of the etiopathogenesis. Oncol. Rev. 2020, 14, 465. [Google Scholar] [CrossRef] [Green Version]
- Gulati, M.; Nobile, C.J. Candida albicans biofilms: Development, regulation, and molecular mechanisms. Microbes Infect. 2016, 18, 310–321. [Google Scholar] [CrossRef] [Green Version]
- Cowen, L.E.; Sanglard, D.; Howard, S.J.; Rogers, P.D.; Perlin, D.S. Mechanisms of Antifungal Drug Resistance. Cold Spring Harb. Perspect. Med. 2014, 5, a019752. [Google Scholar] [CrossRef]
- Reyna-Beltrán, E.; Bazán Méndez, C.I.; Iranzo, M.; Mormeneo, S.; Luna-Arias, J.P. The Cell Wall of Candida albicans: A Proteomics View. In Candida albicans; Sandai, D., Ed.; IntechOpen: London, UK, 2019; pp. 71–92. ISBN 978-1-83880-160-1. [Google Scholar]
- Al-Adham, I.S.I.; Ashour, H.; Al-Kaissi, E.; Khalil, E.; Kierans, M.; Collier, P.J. Studies on the kinetics of killing and the proposed mechanism of action of microemulsions against fungi. Int. J. Pharm. 2013, 454, 226–232. [Google Scholar] [CrossRef]
- Tran, P.H.L.; Duan, W.; Tran, T.T.D. Recent developments of nanoparticle-delivered dosage forms for buccal delivery. Int. J. Pharm. 2019, 571, 118697. [Google Scholar] [CrossRef]
- Zangeneh, M.M.; Bovandi, S.; Gharehyakheh, S.; Zangeneh, A.; Irani, P. Green synthesis and chemical characterization of silver nanoparticles obtained using Allium saralicum aqueous extract and survey of in vitro antioxidant, cytotoxic, antibacterial and antifungal properties. Appl. Organomet. Chem. 2019, 33, e4961. [Google Scholar] [CrossRef]
- Jia, D.; Sun, W. Silver nanoparticles offer a synergistic effect with fluconazole against fluconazole-resistant Candida albicans by abrogating drug efflux pumps and increasing endogenous ROS. Infect. Genet. Evol. 2021, 93, 104937. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Jacobsen, J.; Larsen, S.W.; Genina, N.; van de Weert, M.; Müllertz, A.; Nielsen, H.M.; Mu, H. Graphene oxide as a functional excipient in buccal films for delivery of clotrimazole: Effect of molecular interactions on drug release and antifungal activity in vitro. Int. J. Pharm. 2020, 589, 119811. [Google Scholar] [CrossRef] [PubMed]
- Mady, O.Y.; Donia, A.M.; Al-Madboly, L.A. Miconazole-Urea in a Buccal Film as a New Trend for Treatment of Resistant Mouth Fungal White Patches. Front. Microbiol. 2018, 9, 837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patil, S.; Rao, R.S.; Majumdar, B.; Anil, S. Clinical Appearance of Oral Candida Infection and Therapeutic Strategies. Front. Microbiol. 2015, 6, 1391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.-H.; Ma, Y.-Y.; Ding, Y.; Chen, X.-Q.; Gao, G.-X. An insight into new strategies to combat antifungal drug resistance. Drug Des. Dev. Ther. 2018, 12, 3807–3816. [Google Scholar] [CrossRef] [Green Version]
- Taverne-Ghadwal, L.; Kuhns, M.; Buhl, T.; Schulze, M.H.; Mbaitolum, W.J.; Kersch, L.; Weig, M.; Bader, O.; Groß, U. Epidemiology and Prevalence of Oral Candidiasis in HIV Patients From Chad in the Post-HAART Era. Front. Microbiol. 2022, 13, 844069. [Google Scholar] [CrossRef]
- Akpan, A.; Morgan, R. Oral candidiasis. Postgrad. Med. J. 2002, 78, 455–459. [Google Scholar] [CrossRef]
- Quindós, G.; Gil-Alonso, S.; Marcos-Arias, C.; Sevillano, E.; Mateo, E.; Jauregizar, N.; Eraso, E. Therapeutic tools for oral candidiasis: Current and new antifungal drugs. Med. Oral Patol. Oral Cir. Bucal 2019, 24, e172–e180. [Google Scholar] [CrossRef]
- Vila, T.; Sultan, A.S.; Montelongo-Jauregui, D.; Jabra-Rizk, M.A. Oral Candidiasis: A Disease of Opportunity. J. Fungi 2020, 6, 15. [Google Scholar] [CrossRef] [Green Version]
- Centers of Disease Control and Prevention. Candida Infections of the Mouth, Throat, and Esophagus. Available online: https://www.cdc.gov/fungal/diseases/candidiasis/thrush/index.html (accessed on 5 April 2022).
- Santiso, G.M.; Messina, F.; Gallo, A.; Marín, E.; Depardo, R.; Arechavala, A.; Walker, L.; Negroni, R.; Romero, M.M. Tongue lesion due to Cryptococcus neoformans as the first finding in an HIV-positive patient. Rev. Iberoam. Micol. 2021, 38, 19–22. [Google Scholar] [CrossRef]
- Stone, N.; Gupta, N.; Schwartz, I. Mucormycosis: Time to address this deadly fungal infection. Lancet Microbe 2021, 2, e343–e344. [Google Scholar] [CrossRef]
- Ahmed, E.; Abou-Bakr, A.; Hussein, R.R.; El-Gawish, A.A.; Ras, A.E.; Ghalwash, D.M. Oral mucormycosis in post-COVID-19 patients: A case series. Oral Dis. 2021. [Google Scholar] [CrossRef] [PubMed]
- Nambiar, M.; Varma, S.R.; Jaber, M.; Sreelatha, S.V.; Thomas, B.; Nair, A.S. Mycotic infections—Mucormycosis and oral candidiasis associated with COVID-19: A significant and challenging association. J. Oral Microbiol. 2021, 13, 1967699. [Google Scholar] [CrossRef] [PubMed]
- Nadig, S.D.; Ashwathappa, D.T.; Manjunath, M.; Krishna, S.; Annaji, A.G.; Shivaprakash, P.K. A relationship between salivary flow rates and Candida counts in patients with xerostomia. J. Oral Maxillofac. Pathol. 2017, 21, 316. [Google Scholar] [CrossRef] [Green Version]
- Gleiznys, A.; Zdanavičienė, E.; Žilinskas, J. Candida albicans importance to denture wearers. A literature review. Stomatologija 2015, 17, 54–66. [Google Scholar]
- Lafuente-Ibáñez de Mendoza, I.; Cayero-Garay, A.; Quindós-Andrés, G.; Aguirre-Urizar, J.M. A systematic review on the implication of Candida in peri-implantitis. Int. J. Implant Dent. 2021, 7, 73. [Google Scholar] [CrossRef] [PubMed]
- Santana, I.L.; Gonçalves, L.M.; de Vasconcellos, A.A.; da Silva, W.J.; Cury, J.A.; Del Bel Cury, A.A. Dietary carbohydrates modulate Candida albicans biofilm development on the denture surface. PLoS ONE 2013, 8, e64645. [Google Scholar] [CrossRef] [Green Version]
- Weindl, G.; Wagener, J.; Schaller, M. Epithelial cells and innate antifungal defense. J. Dent. Res. 2010, 89, 666–675. [Google Scholar] [CrossRef] [Green Version]
- Villar, C.C.; Dongari-Bagtzoglou, A. Fungal diseases: Oral dysbiosis in susceptible hosts. Periodontology 2000 2021, 87, 166–180. [Google Scholar] [CrossRef]
- Bertolini, M.; Vazquez Munoz, R.; Archambault, L.; Shah, S.; Souza, J.G.S.; Costa, R.C.; Thompson, A.; Zhou, Y.; Sobue, T.; Dongari-Bagtzoglou, A. Mucosal Bacteria Modulate Candida albicans Virulence in Oropharyngeal Candidiasis. mBio 2021, 12, e0193721. [Google Scholar] [CrossRef]
- Dekhuijzen, P.N.R.; Batsiou, M.; Bjermer, L.; Bosnic-Anticevich, S.; Chrystyn, H.; Papi, A.; Rodríguez-Roisin, R.; Fletcher, M.; Wood, L.; Cifra, A.; et al. Incidence of oral thrush in patients with COPD prescribed inhaled corticosteroids: Effect of drug, dose, and device. Respir. Med. 2016, 120, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Gunasegar, S.; Himratul-Aznita, W.H. Nicotine enhances the thickness of biofilm and adherence of Candida albicans ATCC 14053 and Candida parapsilosis ATCC 22019. FEMS Yeast Res. 2019, 19, foy123. [Google Scholar] [CrossRef]
- Salerno, C.; Pascale, M.; Contaldo, M.; Esposito, V.; Busciolano, M.; Milillo, L.; Guida, A.; Petruzzi, M.; Serpico, R. Candida-associated denture stomatitis. Med. Oral Patol. Oral Cir. Bucal 2011, 16, e139–e143. [Google Scholar] [CrossRef]
- Lu, S.-Y. Perception of iron deficiency from oral mucosa alterations that show a high prevalence of Candida infection. J. Formos. Med. Assoc. 2016, 115, 619–627. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, F.; Javaheri, M.R.; Nekoeian, S.; Dehghan, P. Identification of Candida species in the oral cavity of diabetic patients. Curr. Med. Mycol. 2016, 2, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Ciesielska, A.; Kusiak, A.; Ossowska, A.; Grzybowska, M.E. Changes in the Oral Cavity in Menopausal Women-A Narrative Review. Int. J. Environ. Res. Public Health 2021, 19, 253. [Google Scholar] [CrossRef]
- Warrier, S.A.; Sathasivasubramanian, S. Human immunodeficiency virus induced oral candidiasis. J. Pharm. Bioallied Sci. 2015, 7, S812–S814. [Google Scholar] [CrossRef]
- Aslani, N.; Janbabaei, G.; Abastabar, M.; Meis, J.F.; Babaeian, M.; Khodavaisy, S.; Boekhout, T.; Badali, H. Identification of uncommon oral yeasts from cancer patients by MALDI-TOF mass spectrometry. BMC Infect. Dis. 2018, 18, 24. [Google Scholar] [CrossRef]
- Lalla, R.V.; Treister, N.; Sollecito, T.; Schmidt, B.; Patton, L.L.; Mohammadi, K.; Hodges, J.S.; Brennan, M.T. Oral complications at 6 months after radiation therapy for head and neck cancer. Oral Dis. 2017, 23, 1134–1143. [Google Scholar] [CrossRef]
- Buranarom, N.; Komin, O.; Matangkasombut, O. Hyposalivation, oral health, and Candida colonization in independent dentate elders. PLoS ONE 2020, 15, e0242832. [Google Scholar] [CrossRef]
- Lynge Pedersen, A.M.; Belstrøm, D. The role of natural salivary defences in maintaining a healthy oral microbiota. J. Dent. 2019, 80, S3–S12. [Google Scholar] [CrossRef]
- Carniello, V.; Peterson, B.W.; van der Mei, H.C.; Busscher, H.J. Physico-chemistry from initial bacterial adhesion to surface-programmed biofilm growth. Adv. Colloid Interface Sci. 2018, 261, 1–14. [Google Scholar] [CrossRef]
- Bars, P.L.; Soueidan, A. Distribution Patterns of E-Cadherin, Type VII Collagen and Fibronectin in Denture-Related Stomatitis: A Preliminary Study. Open Dent. J. 2012, 6, 14–22. [Google Scholar] [CrossRef] [Green Version]
- Komatsu, T.; Watanabe, K.; Hamada, N.; Helmerhorst, E.; Oppenheim, F.; Lee, M.C. Association between Antimicrobial Peptide Histatin 5 Levels and Prevalence of Candida in Saliva of Patients with Down Syndrome. Antibiotics 2021, 10, 494. [Google Scholar] [CrossRef]
- Buda De Cesare, G.; Cristy, S.A.; Garsin, D.A.; Lorenz, M.C. Antimicrobial Peptides: A New Frontier in Antifungal Therapy. mBio 2022, 11, e02123-20. [Google Scholar] [CrossRef]
- Kilian, M.; Chapple, I.L.C.; Hannig, M.; Marsh, P.D.; Meuric, V.; Pedersen, A.M.L.; Tonetti, M.S.; Wade, W.G.; Zaura, E. The oral microbiome—An update for oral healthcare professionals. Br. Dent. J. 2016, 221, 657–666. [Google Scholar] [CrossRef]
- Buu, L.-M.; Chen, Y.-C. Impact of glucose levels on expression of hypha-associated secreted aspartyl proteinases in Candida albicans. J. Biomed. Sci. 2014, 21, 22. [Google Scholar] [CrossRef] [Green Version]
- Correia, I.; Prieto, D.; Román, E.; Wilson, D.; Hube, B.; Alonso-Monge, R.; Pla, J. Cooperative Role of MAPK Pathways in the Interaction of Candida albicans with the Host Epithelium. Microorganisms 2019, 8, 48. [Google Scholar] [CrossRef] [Green Version]
- Fourie, R.; Kuloyo, O.O.; Mochochoko, B.M.; Albertyn, J.; Pohl, C.H. Iron at the Centre of Candida albicans Interactions. Front. Cell. Infect. Microbiol. 2018, 8, 185. [Google Scholar] [CrossRef] [Green Version]
- Popoviciu, M.S.; Marin, V.N.; Vesa, C.M.; Stefan, S.D.; Stoica, R.A.; Serafinceanu, C.; Merlo, E.M.; Rizvi, A.A.; Rizzo, M.; Busnatu, S.; et al. Correlations between Diabetes Mellitus Self-Care Activities and Glycaemic Control in the Adult Population: A Cross-Sectional Study. Healthcare 2022, 10, 174. [Google Scholar] [CrossRef]
- Cheng, X.; He, F.; Si, M.; Sun, P.; Chen, Q. Effects of Antibiotic Use on Saliva Antibody Content and Oral Microbiota in Sprague Dawley Rats. Front. Cell. Infect. Microbiol. 2022, 12, 721691. [Google Scholar] [CrossRef]
- Salvatori, O.; Puri, S.; Tati, S.; Edgerton, M. Innate Immunity and Saliva in Candida albicans—Mediated Oral Diseases. J. Dent. Res. 2016, 95, 365–371. [Google Scholar] [CrossRef] [Green Version]
- Lafleur, M.D.; Qi, Q.; Lewis, K. Patients with long-term oral carriage harbor high-persister mutants of Candida albicans. Antimicrob. Agents Chemother. 2010, 54, 39–44. [Google Scholar] [CrossRef] [Green Version]
- Rajendra Santosh, A.B.; Muddana, K.; Bakki, S.R. Fungal Infections of Oral Cavity: Diagnosis, Management, and Association with COVID-19. SN Compr. Clin. Med. 2021, 3, 1373–1384. [Google Scholar] [CrossRef]
- Jain, A.; Taneja, S. Post-COVID fungal infections of maxillofacial region: A systematic review. Oral Maxillofac. Surg. 2021, 1–7. [Google Scholar] [CrossRef]
- Noble, S.M.; Gianetti, B.A.; Witchley, J.N. Candida albicans cell-type switching and functional plasticity in the mammalian host. Nat. Rev. Microbiol. 2017, 15, 96–108. [Google Scholar] [CrossRef] [Green Version]
- Wächtler, B.; Citiulo, F.; Jablonowski, N.; Förster, S.; Dalle, F.; Schaller, M.; Wilson, D.; Hube, B. Candida albicans-epithelial interactions: Dissecting the roles of active penetration, induced endocytosis and host factors on the infection process. PLoS ONE 2012, 7, e36952. [Google Scholar] [CrossRef] [Green Version]
- Swidergall, M.; Filler, S.G. Oropharyngeal Candidiasis: Fungal Invasion and Epithelial Cell Responses. PLOS Pathog. 2017, 13, e1006056. [Google Scholar] [CrossRef]
- Şenel, S. An Overview of Physical, Microbiological and Immune Barriers of Oral Mucosa. Int. J. Mol. Sci. 2021, 22, 7821. [Google Scholar] [CrossRef]
- Wu, R.-Q.; Zhang, D.-F.; Tu, E.; Chen, Q.-M.; Chen, W. The mucosal immune system in the oral cavity—An orchestra of T cell diversity. Int. J. Oral Sci. 2014, 6, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Barua, S.; Kim, H.; Jo, K.; Seo, C.W.; Park, T.J.; Lee, K.B.; Yun, G.; Oh, K.; Lee, J. Drug delivery techniques for buccal route: Formulation strategies and recent advances in dosage form design. J. Pharm. Investig. 2016, 46, 593–613. [Google Scholar] [CrossRef]
- Fábián, T.K.; Hermann, P.; Beck, A.; Fejérdy, P.; Fábián, G. Salivary defense proteins: Their network and role in innate and acquired oral immunity. Int. J. Mol. Sci. 2012, 13, 4295–4320. [Google Scholar] [CrossRef] [PubMed]
- Colombo, P.; Cagnani, S.; Buttini, F.; Santi, P.; Sonvico, F.; Russo, P.; Colombo, G. Biological In Vitro Models for Absorption by Non-Oral Routes. In Reference Module in Chemistry, Molecular Sciences and Chemical Engineering; Elsevier: Oxford, UK, 2013; ISBN 9780124095472. [Google Scholar]
- Squier, C.A. The permeability of oral mucosa. Crit. Rev. Oral Biol. Med. Off. Publ. Am. Assoc. Oral Biol. 1991, 2, 13–32. [Google Scholar] [CrossRef]
- Tsui, C.; Kong, E.F.; Jabra-Rizk, M.A. Pathogenesis of Candida albicans biofilm. Pathog. Dis. 2016, 74, ftw018. [Google Scholar] [CrossRef] [Green Version]
- Knapp, K.M.; Flynn, P.M. Candidiasis. In Feigin and Cherry’s Textbook of Pediatric Infectious Diseases; Feigin, R.D., Cherry, J.D., Demmler-Harrison, G.J., Kaplan, S.L., Eds.; W.B. Saunders: Philadelphia, PA, USA, 2009; pp. 2741–2751. ISBN 978-1-4160-4044-6. [Google Scholar]
- Lalla, R.V.; Latortue, M.C.; Hong, C.H.; Ariyawardana, A.; D’Amato-Palumbo, S.; Fischer, D.J.; Martof, A.; Nicolatou-Galitis, O.; Patton, L.L.; Elting, L.S.; et al. A systematic review of oral fungal infections in patients receiving cancer therapy. Support. Care Cancer 2010, 18, 985–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allert, S.; Förster, T.M.; Svensson, C.-M.; Richardson, J.P.; Pawlik, T.; Hebecker, B.; Rudolphi, S.; Juraschitz, M.; Schaller, M.; Blagojevic, M.; et al. Candida albicans-Induced Epithelial Damage Mediates Translocation through Intestinal Barriers. mBio 2018, 9, e00915-18. [Google Scholar] [CrossRef] [Green Version]
- Chow, E.W.; Pang, L.M.; Wang, Y. From Jekyll to Hyde: The Yeast–Hyphal Transition of Candida albicans. Pathogens 2021, 10, 859. [Google Scholar] [CrossRef]
- Sudbery, P.E. Growth of Candida albicans hyphae. Nat. Rev. Microbiol. 2011, 9, 737–748. [Google Scholar] [CrossRef]
- Bernard, C.; Girardot, M.; Imbert, C. Candida albicans interaction with Gram-positive bacteria within interkingdom biofilms. J. Mycol. Med. 2020, 30, 100909. [Google Scholar] [CrossRef]
- Rapala-Kozik, M.; Zawrotniak, M.; Gogol, M.; Bartnicka, D.; Satala, D.; Smolarz, M.; Karkowska-Kuleta, J.; Kozik, A. Interactions of Candida albicans Cells with Aerobic and Anaerobic Bacteria during Formation of Mixed Biofilms in the Oral Cavity; Zawrotniak, M., Ed.; IntechOpen: Rijeka, Croatia, 2019; ISBN 978-1-83880-160-1. [Google Scholar]
- Li, Z.; Nielsen, K. Morphology Changes in Human Fungal Pathogens upon Interaction with the Host. J. Fungi 2017, 3, 66. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.; Cowen, L.E.; Griffin, A.M.; Chan, L.; Köhler, J.R. The Candida albicans pescadillo homolog is required for normal hypha-to-yeast morphogenesis and yeast proliferation. Proc. Natl. Acad. Sci. USA 2008, 105, 20918–20923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
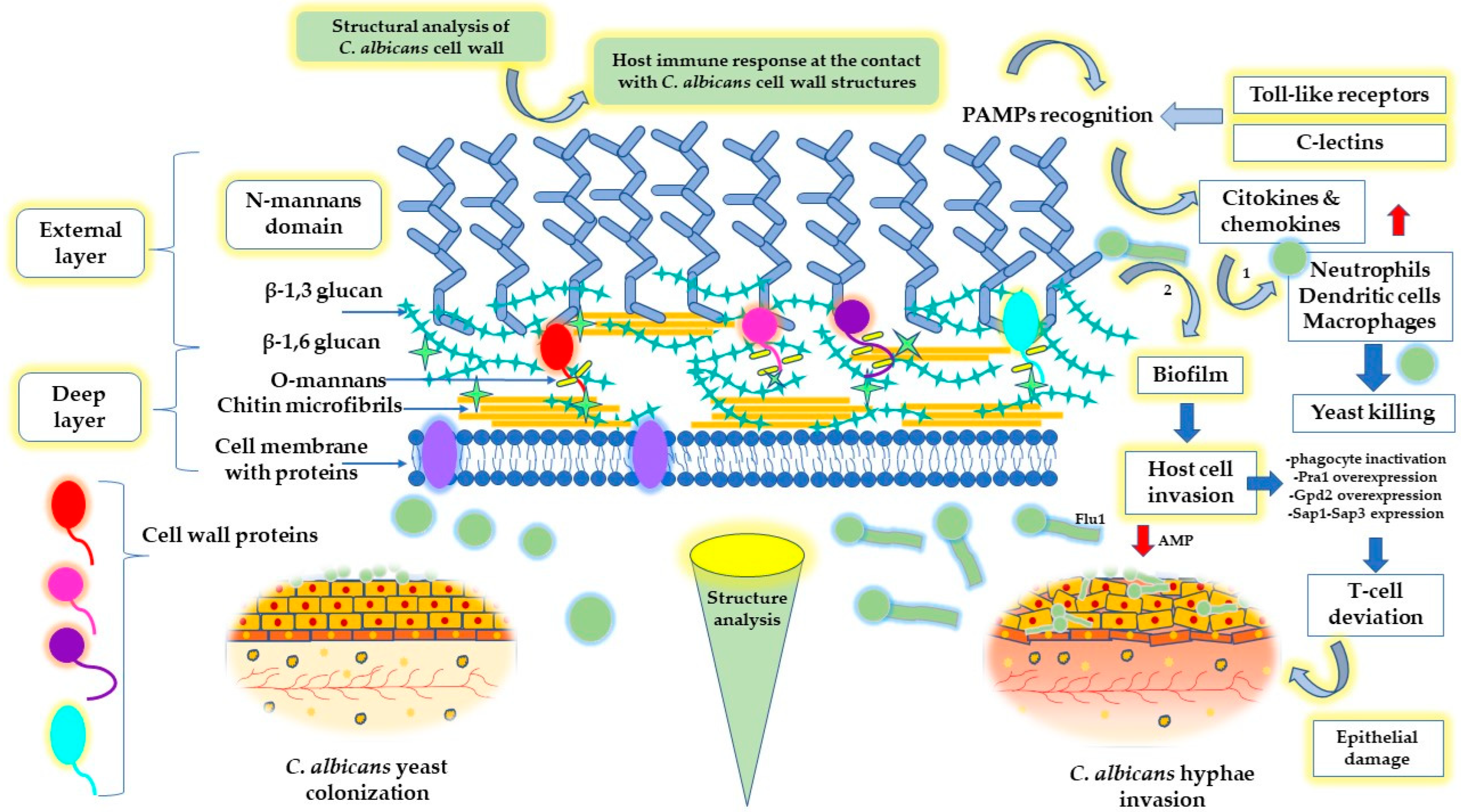
- Garcia-Rubio, R.; de Oliveira, H.C.; Rivera, J.; Trevijano-Contador, N. The Fungal Cell Wall: Candida, Cryptococcus, and Aspergillus Species. Front. Microbiol. 2020, 10, 2993. [Google Scholar] [CrossRef] [PubMed]
- Ene, I.V.; Walker, L.A.; Schiavone, M.; Lee, K.K.; Martin-Yken, H.; Dague, E.; Gow, N.A.; Munro, C.A.; Brown, A.J.P. Cell Wall Remodeling Enzymes Modulate Fungal Cell Wall Elasticity and Osmotic Stress Resistance. mBio 2015, 6, e00986-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Childers, D.S.; Avelar, G.M.; Bain, J.M.; Larcombe, D.E.; Pradhan, A.; Budge, S.; Heaney, H.; Brown, A.J.P. Impact of the Environment upon the Candida albicans Cell Wall and Resultant Effects upon Immune Surveillance BT—The Fungal Cell Wall: An Armour and a Weapon for Human Fungal Pathogens; Latgé, J.-P., Ed.; Springer: Cham, Switzerland, 2020; pp. 297–330. ISBN 978-3-030-49928-0. [Google Scholar]
- Lenardon, M.D.; Sood, P.; Dorfmueller, H.C.; Brown, A.J.P.; Gow, N.A.R. Scalar nanostructure of the Candida albicans cell wall; a molecular, cellular and ultrastructural analysis and interpretation. Cell Surf. 2020, 6, 100047. [Google Scholar] [CrossRef]
- Ruiz-Herrera, J.; Victoria Elorza, M.; Valentín, E.; Sentandreu, R. Molecular organization of the cell wall of Candida albicans and its relation to pathogenicity. FEMS Yeast Res. 2006, 6, 14–29. [Google Scholar] [CrossRef] [Green Version]
- Alim, D.; Sircaik, S.; Panwar, S.L. The Significance of Lipids to Biofilm Formation in Candida albicans: An Emerging Perspective. J. Fungi 2018, 4, 140. [Google Scholar] [CrossRef] [Green Version]
- Dwivedi, P.; Thompson, A.; Xie, Z.; Kashleva, H.; Ganguly, S.; Mitchell, A.P.; Dongari-Bagtzoglou, A. Role of Bcr1-activated genes Hwp1 and Hyr1 in Candida albicans oral mucosal biofilms and neutrophil evasion. PLoS ONE 2011, 6, e16218. [Google Scholar] [CrossRef] [Green Version]
- Murciano, C.; Moyes, D.L.; Runglall, M.; Tobouti, P.; Islam, A.; Hoyer, L.L.; Naglik, J.R. Evaluation of the role of Candida albicans agglutinin-like sequence (Als) proteins in human oral epithelial cell interactions. PLoS ONE 2012, 7, e33362. [Google Scholar] [CrossRef] [Green Version]
- Glazier, V.E.; Murante, T.; Murante, D.; Koselny, K.; Liu, Y.; Kim, D.; Koo, H.; Krysan, D.J. Genetic analysis of the Candida albicans biofilm transcription factor network using simple and complex haploinsufficiency. PLOS Genet. 2017, 13, e1006948. [Google Scholar] [CrossRef] [Green Version]
- Chandra, J.; Mukherjee, P.K. Candida Biofilms: Development, Architecture, and Resistance. Microbiol. Spectr. 2015, 3, 3–4. [Google Scholar] [CrossRef] [Green Version]
- Cavalheiro, M.; Teixeira, M.C. Candida Biofilms: Threats, Challenges, and Promising Strategies. Front. Med. 2018, 5, 28. [Google Scholar] [CrossRef] [Green Version]
- Childers, D.S.; Kadosh, D. Filament condition-specific response elements control the expression of NRG1 and UME6, key transcriptional regulators of morphology and virulence in Candida albicans. PLoS ONE 2015, 10, e0122775. [Google Scholar] [CrossRef] [Green Version]
- Gong, Y.; Li, T.; Yu, C.; Sun, S. Candida albicans Heat Shock Proteins and Hsps-Associated Signaling Pathways as Potential Antifungal Targets. Front. Cell. Infect. Microbiol. 2017, 7, 520. [Google Scholar] [CrossRef]
- McCall, A.D.; Pathirana, R.U.; Prabhakar, A.; Cullen, P.J.; Edgerton, M. Candida albicans biofilm development is governed by cooperative attachment and adhesion maintenance proteins. NPJ Biofilms Microbiomes 2019, 5, 21. [Google Scholar] [CrossRef] [Green Version]
- Dalle, F.; Wächtler, B.; L’Ollivier, C.; Holland, G.; Bannert, N.; Wilson, D.; Labruère, C.; Bonnin, A.; Hube, B. Cellular interactions of Candida albicans with human oral epithelial cells and enterocytes. Cell. Microbiol. 2010, 12, 248–271. [Google Scholar] [CrossRef]
- Phan, Q.T.; Myers, C.L.; Fu, Y.; Sheppard, D.C.; Yeaman, M.R.; Welch, W.H.; Ibrahim, A.S.; Edwards, J.E., Jr.; Filler, S.G. Als3 is a Candida albicans invasin that binds to cadherins and induces endocytosis by host cells. PLoS Biol. 2007, 5, e64. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.N.; Solis, N.V.; Phan, Q.T.; Bajwa, J.S.; Kashleva, H.; Thompson, A.; Liu, Y.; Dongari-Bagtzoglou, A.; Edgerton, M.; Filler, S.G. Host cell invasion and virulence mediated by Candida albicans Ssa1. PLoS Pathog. 2010, 6, e1001181. [Google Scholar] [CrossRef] [Green Version]
- Mukaremera, L.; Lee, K.K.; Mora-Montes, H.M.; Gow, N.A.R. Candida albicans Yeast, Pseudohyphal, and Hyphal Morphogenesis Differentially Affects Immune Recognition. Front. Immunol. 2017, 8, 629. [Google Scholar] [CrossRef] [Green Version]
- Netea, M.G.; Joosten, L.A.B.; van der Meer, J.W.M.; Kullberg, B.-J.; van de Veerdonk, F.L. Immune defence against Candida fungal infections. Nat. Rev. Immunol. 2015, 15, 630–642. [Google Scholar] [CrossRef]
- Atriwal, T.; Azeem, K.; Husain, F.M.; Hussain, A.; Khan, M.N.; Alajmi, M.F.; Abid, M. Mechanistic Understanding of Candida albicans Biofilm Formation and Approaches for Its Inhibition. Front. Microbiol. 2021, 12, 638609. [Google Scholar] [CrossRef]
- Harpf, V.; Rambach, G.; Würzner, R.; Lass-Flörl, C.; Speth, C. Candida and Complement: New Aspects in an Old Battle. Front. Immunol. 2020, 11, 1471. [Google Scholar] [CrossRef]
- Swidergall, M.; Ernst, J.F. Interplay between Candida albicans and the antimicrobial peptide armory. Eukaryot. Cell 2014, 13, 950–957. [Google Scholar] [CrossRef] [Green Version]
- Puri, S.; Kumar, R.; Chadha, S.; Tati, S.; Conti, H.R.; Hube, B.; Cullen, P.J.; Edgerton, M. Secreted Aspartic Protease Cleavage of Candida albicans Msb2 Activates Cek1 MAPK Signaling Affecting Biofilm Formation and Oropharyngeal Candidiasis. PLoS ONE 2012, 7, e46020. [Google Scholar] [CrossRef] [Green Version]
- Kavanaugh, N.L.; Zhang, A.Q.; Nobile, C.J.; Johnson, A.D.; Ribbeck, K. Mucins suppress virulence traits of Candida albicans. mBio 2014, 5, e01911. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.; Lewis, M. Pathogenesis and treatment of oral candidosis. J. Oral Microbiol. 2011, 3, 5771. [Google Scholar] [CrossRef] [Green Version]
- Hellstein, J.W.; Marek, C.L. Candidiasis: Red and White Manifestations in the Oral Cavity. Head Neck Pathol. 2019, 13, 25–32. [Google Scholar] [CrossRef]
- Li, B.; Fang, X.; Hu, X.; Hua, H.; Wei, P. Successful treatment of chronic hyperplastic candidiasis with 5-aminolevulinic acid photodynamic therapy: A case report. Photodiagnosis Photodyn. Ther. 2022, 37, 102633. [Google Scholar] [CrossRef]
- Lu, S.-Y. Oral Candidosis: Pathophysiology and Best Practice for Diagnosis, Classification, and Successful Management. J. Fungi 2021, 7, 555. [Google Scholar] [CrossRef]
- Garcia-Cuesta, C.; Sarrion-Pérez, M.-G.; Bagán, J.V. Current treatment of oral candidiasis: A literature review. J. Clin. Exp. Dent. 2014, 6, e576–e582. [Google Scholar] [CrossRef]
- Scorzoni, L.; de Paula E Silva, A.C.A.; Marcos, C.M.; Assato, P.A.; de Melo, W.C.M.A.; de Oliveira, H.C.; Costa-Orlandi, C.B.; Mendes-Giannini, M.J.S.; Fusco-Almeida, A.M. Antifungal Therapy: New Advances in the Understanding and Treatment of Mycosis. Front. Microbiol. 2017, 8, 36. [Google Scholar] [CrossRef] [Green Version]
- Fernández Campos, F.; Calpena Campmany, A.C.; Rodríguez Delgado, G.; López Serrano, O.; Clares Naveros, B. Development and characterization of a novel nystatin-loaded nanoemulsion for the buccal treatment of candidosis: Ultrastructural effects and release studies. J. Pharm. Sci. 2012, 101, 3739–3752. [Google Scholar] [CrossRef]
- Stone, N.R.H.; Bicanic, T.; Salim, R.; Hope, W. Liposomal Amphotericin B (AmBisome®): A Review of the Pharmacokinetics, Pharmacodynamics, Clinical Experience and Future Directions. Drugs 2016, 76, 485–500. [Google Scholar] [CrossRef] [Green Version]
- Lalla, R.V.; Bensadoun, R.-J. Miconazole mucoadhesive tablet for oropharyngeal candidiasis. Expert Rev. Anti. Infect. Ther. 2011, 9, 13–17. [Google Scholar] [CrossRef] [Green Version]
- Kofla, G.; Ruhnke, M. Pharmacology and metabolism of anidulafungin, caspofungin and micafungin in the treatment of invasive candidosis—Review of the literature. Eur. J. Med. Res. 2011, 16, 159. [Google Scholar] [CrossRef] [Green Version]
- Coad, B.R.; Lamont-Friedrich, S.J.; Gwynne, L.; Jasieniak, M.; Griesser, S.S.; Traven, A.; Peleg, A.Y.; Griesser, H.J. Surface coatings with covalently attached caspofungin are effective in eliminating fungal pathogens. J. Mater. Chem. B 2015, 3, 8469–8476. [Google Scholar] [CrossRef]
- Perlin, D.S.; Rautemaa-Richardson, R.; Alastruey-Izquierdo, A. The global problem of antifungal resistance: Prevalence, mechanisms, and management. Lancet Infect. Dis. 2017, 17, e383–e392. [Google Scholar] [CrossRef]
- Gilhotra, R.M.; Ikram, M.; Srivastava, S.; Gilhotra, N. A clinical perspective on mucoadhesive buccal drug delivery systems. J. Biomed. Res. 2014, 28, 81–97. [Google Scholar] [CrossRef]
- Jacob, S.; Nair, A.B.; Boddu, S.H.S.; Gorain, B.; Sreeharsha, N.; Shah, J. An Updated Overview of the Emerging Role of Patch and Film-Based Buccal Delivery Systems. Pharmaceutics 2021, 13, 1206. [Google Scholar] [CrossRef]
- Uzunoğlu, B.; Wilson, C.G.; Sağıroğlu, M.; Yüksel, S.; Şenel, S. Mucoadhesive bilayered buccal platform for antifungal drug delivery into the oral cavity. Drug Deliv. Transl. Res. 2021, 11, 318–327. [Google Scholar] [CrossRef]
- Vazquez, J.A.; Sobel, J.D. Miconazole Mucoadhesive Tablets: A Novel Delivery System. Clin. Infect. Dis. 2012, 54, 1480–1484. [Google Scholar] [CrossRef] [Green Version]
- Mathé, L.; Van Dijck, P. Recent insights into Candida albicans biofilm resistance mechanisms. Curr. Genet. 2013, 59, 251–264. [Google Scholar] [CrossRef] [Green Version]
- Grover, N.D. Echinocandins: A ray of hope in antifungal drug therapy. Indian J. Pharmacol. 2010, 42, 9–11. [Google Scholar] [CrossRef] [Green Version]
- Delma, F.Z.; Al-Hatmi, A.M.S.; Brüggemann, R.J.M.; Melchers, W.J.G.; de Hoog, S.; Verweij, P.E.; Buil, J.B. Molecular Mechanisms of 5-Fluorocytosine Resistance in Yeasts and Filamentous Fungi. J. Fungi 2021, 7, 909. [Google Scholar] [CrossRef]
- Prasadh, S.; Ratheesh, V.; Wong, R. Chapter 6—Impact of biomaterial mechanics on cellular and molecular responses. In Handbook of Nanomaterials Biocompatibility; Mozafari, M., Ed.; Woodhead Publishing: Duckford, UK, 2020; pp. 85–109. ISBN 978-0-08-102967-1. [Google Scholar]
- Augustine, R.; Hasan, A. Chapter 21—Cellular response to nanobiomaterials. In Handbook of Nanomaterials Biocompatibility; Mozafari, M., Ed.; Woodhead Publishing: Duckford, UK, 2020; pp. 473–504. ISBN 978-0-08-102967-1. [Google Scholar]
- Joyce, K.; Fabra, G.T.; Bozkurt, Y.; Pandit, A. Bioactive potential of natural biomaterials: Identification, retention and assessment of biological properties. Signal Transduct. Target. Ther. 2021, 6, 122. [Google Scholar] [CrossRef]
- Augustine, R.; Hasan, A.; Primavera, R.; Wilson, R.J.; Thakor, A.S.; Kevadiya, B.D. Cellular uptake and retention of nanoparticles: Insights on particle properties and interaction with cellular components. Mater. Today Commun. 2020, 25, 101692. [Google Scholar] [CrossRef]
- Rodríguez-Arco, L.; Poma, A.; Ruiz-Pérez, L.; Scarpa, E.; Ngamkham, K.; Battaglia, G. Molecular bionics—Engineering biomaterials at the molecular level using biological principles. Biomaterials 2019, 192, 26–50. [Google Scholar] [CrossRef]
- Coad, B.R.; Kidd, S.E.; Ellis, D.H.; Griesser, H.J. Biomaterials surfaces capable of resisting fungal attachment and biofilm formation. Biotechnol. Adv. 2014, 32, 296–307. [Google Scholar] [CrossRef]
- Vera-González, N.; Shukla, A. Advances in Biomaterials for the Prevention and Disruption of Candida Biofilms. Front. Microbiol. 2020, 11, 538602. [Google Scholar] [CrossRef]
- Wen, J.; Jiang, F.; Yeh, C.-K.; Sun, Y. Controlling fungal biofilms with functional drug delivery denture biomaterials. Colloids Surf. B. Biointerfaces 2016, 140, 19–27. [Google Scholar] [CrossRef] [Green Version]
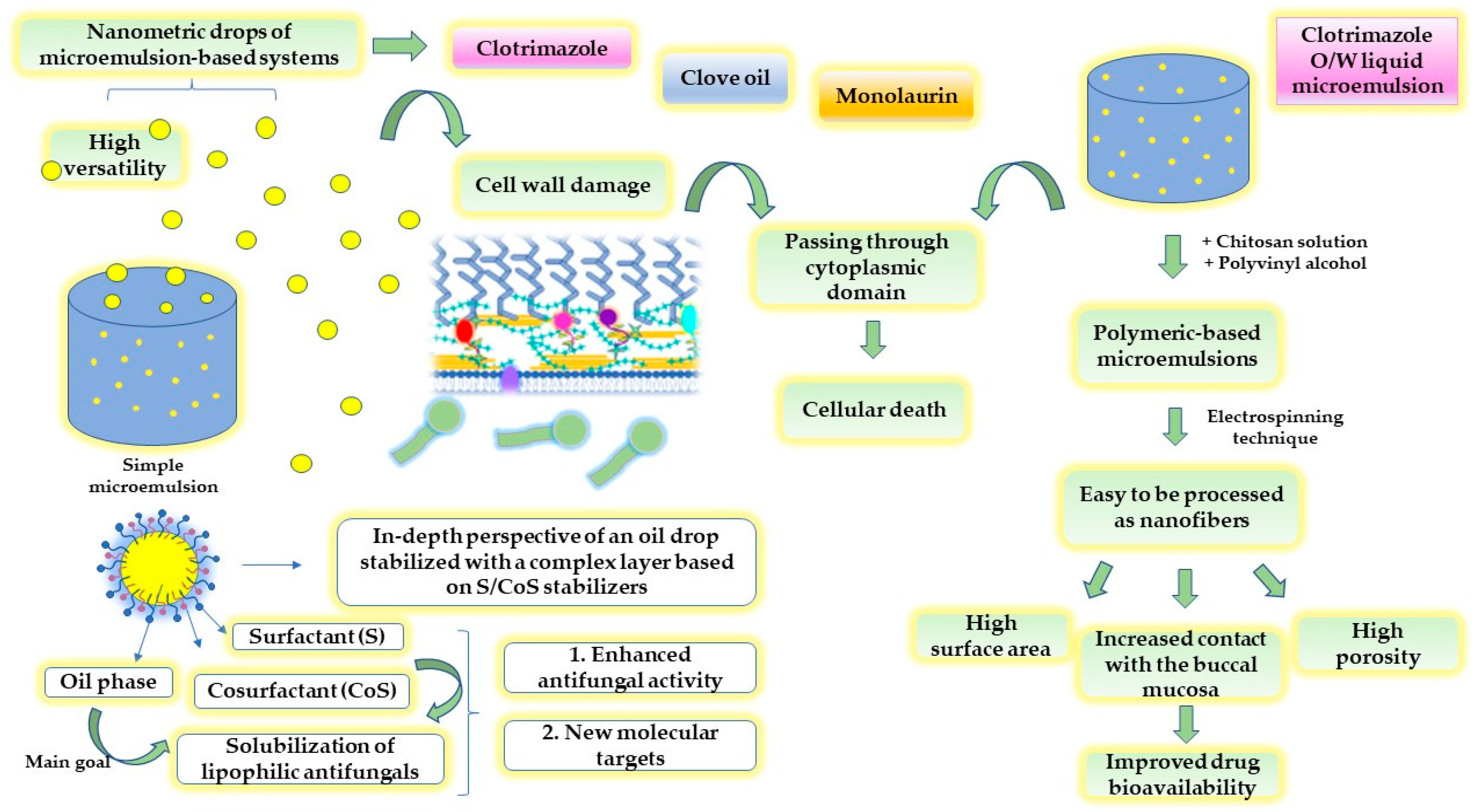
- Tartaro, G.; Mateos, H.; Schirone, D.; Angelico, R.; Palazzo, G. Microemulsion Microstructure(s): A Tutorial Review. Nanomaterials 2020, 10, 1657. [Google Scholar] [CrossRef]
- Gradzielski, M.; Duvail, M.; de Molina, P.M.; Simon, M.; Talmon, Y.; Zemb, T. Using Microemulsions: Formulation Based on Knowledge of Their Mesostructure. Chem. Rev. 2021, 121, 5671–5740. [Google Scholar] [CrossRef] [PubMed]
- Talianu, M.T.; Dinu-Pîrvu, C.E.; Ghica, M.V.; Anuţa, V.; Jinga, V.; Popa, L. Foray into Concepts of Design and Evaluation of Microemulsions as a Modern Approach for Topical Applications in Acne Pathology. Nanomaterials 2020, 10, 2292. [Google Scholar] [CrossRef]
- Anicescu, M.-C.; Dinu-Pîrvu, C.-E.; Talianu, M.-T.; Ghica, M.V.; Anuța, V.; Prisada, R.-M.; Nicoară, A.C.; Popa, L. Insights from a Box–Behnken Optimization Study of Microemulsions with Salicylic Acid for Acne Therapy. Pharmaceutics 2022, 14, 174. [Google Scholar] [CrossRef]
- Kaewbanjong, J.; Wan Sia Heng, P.; Boonme, P. Clotrimazole microemulsion and microemulsion-based gel: Evaluation of buccal drug delivery and irritancy using chick chorioallantoic membrane as the model. J. Pharm. Pharmacol. 2017, 69, 1716–1723. [Google Scholar] [CrossRef]
- Tonglairoum, P.; Ngawhirunpat, T.; Rojanarata, T.; Kaomongkolgit, R.; Opanasopit, P. Fabrication of a novel scaffold of clotrimazole-microemulsion-containing nanofibers using an electrospinning process for oral candidiasis applications. Colloids Surf. B. Biointerfaces 2015, 126, 18–25. [Google Scholar] [CrossRef]
- Edmans, J.G.; Clitherow, K.H.; Murdoch, C.; Hatton, P.V.; Spain, S.G.; Colley, H.E. Mucoadhesive Electrospun Fibre-Based Technologies for Oral Medicine. Pharmaceutics 2020, 12, 504. [Google Scholar] [CrossRef]
- Monton, C.; Settharaksa, S.; Suksaeree, J.; Chusut, T. The preparation, characterization, and stability evaluation of a microemulsion-based oral spray containing clove oil for the treatment of oral candidiasis. J. Drug Deliv. Sci. Technol. 2020, 57, 101735. [Google Scholar] [CrossRef]
- Rajkowska, K.; Nowicka-Krawczyk, P.; Kunicka-Styczyńska, A. Effect of Clove and Thyme Essential Oils on Candida Biofilm Formation and the Oil Distribution in Yeast Cells. Molecules 2019, 24, 1954. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, A.; Wani, M.Y.; Khan, A.; Manzoor, N.; Molepo, J. Synergistic Interactions of Eugenol-tosylate and Its Congeners with Fluconazole against Candida albicans. PLoS ONE 2015, 10, e0145053. [Google Scholar] [CrossRef]
- Tubtimsri, S.; Weerapol, Y.; Soontaranon, S.; Limmatvapirat, C.; Limmatvapirat, S. Monolaurin-Loaded Gel-Like Microemulsion for Oropharyngeal Candidiasis Treatment: Structural Characterisation and In Vitro Antifungal Property. AAPS PharmSciTech 2022, 23, 87. [Google Scholar] [CrossRef]
- McClements, D.J. Nanoemulsions versus microemulsions: Terminology, differences, and similarities. Soft Matter 2012, 8, 1719–1729. [Google Scholar] [CrossRef]
- Chime, S.A.; Kenechukwu, F.C.; Attama, A.A. Nanoemulsions—Advances in Formulation, Characterization and Applications in Drug Delivery. In Application of Nanotechnology in Drug Delivery; Sezer, A.D., Ed.; IntechOpen: Rijeka, Croatia, 2014; pp. 77–126. [Google Scholar]
- Kotta, S.; Khan, A.W.; Pramod, K.; Ansari, S.H.; Sharma, R.K.; Ali, J. Exploring oral nanoemulsions for bioavailability enhancement of poorly water-soluble drugs. Expert Opin. Drug Deliv. 2012, 9, 585–598. [Google Scholar] [CrossRef] [PubMed]
- Sabjan, K.B.; Munawar, S.M.; Rajendiran, D.; Vinoji, S.K.; Kasinathan, K. Nanoemulsion as Oral Drug Delivery—A Review. Curr. Drug Res. Rev. Former. Curr. Drug Abus. Rev. 2020, 12, 4–15. [Google Scholar] [CrossRef] [PubMed]
- Souto, E.B.; Cano, A.; Martins-Gomes, C.; Coutinho, T.E.; Zielińska, A.; Silva, A.M. Microemulsions and Nanoemulsions in Skin Drug Delivery. Bioengineering 2022, 9, 158. [Google Scholar] [CrossRef] [PubMed]
- Garcia, A.; Fan, Y.Y.; Vellanki, S.; Huh, E.Y.; Vanegas, D.; Wang, S.H.; Lee, S.C. Nanoemulsion as an Effective Treatment against Human-Pathogenic Fungi. mSphere 2019, 4, e00729-19. [Google Scholar] [CrossRef] [Green Version]
- Narang, J.K.; Narang, R.S. Emerging role of nanoemulsions in oral health management. Int. J. Pharm. Investig. 2017, 7, 1–3. [Google Scholar] [CrossRef]
- Soriano-Ruiz, J.L.; Calpena-Capmany, A.C.; Cañadas-Enrich, C.; Febrer, N.B.; Suñer-Carbó, J.; Souto, E.B.; Clares-Naveros, B. Biopharmaceutical profile of a clotrimazole nanoemulsion: Evaluation on skin and mucosae as anticandidal agent. Int. J. Pharm. 2019, 554, 105–115. [Google Scholar] [CrossRef]
- Hosny, K.M.; Aldawsari, H.M.; Bahmdan, R.H.; Sindi, A.M.; Kurakula, M.; Alrobaian, M.M.; Aldryhim, A.Y.; Alkhalidi, H.M.; Bahmdan, H.H.; Khallaf, R.A.; et al. Preparation, Optimization, and Evaluation of Hyaluronic Acid-Based Hydrogel Loaded with Miconazole Self-Nanoemulsion for the Treatment of Oral Thrush. AAPS PharmSciTech 2019, 20, 297. [Google Scholar] [CrossRef]
- Hosny, K.; Asfour, H.; Rizg, W.; Alhakamy, N.A.; Sindi, A.; Alkhalidi, H.; Abualsunun, W.; Bakhaidar, R.; Almehmady, A.M.; Akeel, S.; et al. Formulation, Optimization, and Evaluation of Oregano Oil Nanoemulsions for the Treatment of Infections Due to Oral Microbiota. Int. J. Nanomed. 2021, 16, 5465–5478. [Google Scholar] [CrossRef]
- Hosseini, S.S.; Yadegari, M.H.; Rajabibazl, M.; Ghaemi, E.A. Inhibitory effects of carvacrol on the expression of secreted aspartyl proteinases 1–3 in fluconazole-resistant Candida albicans isolates. Iran. J. Microbiol. 2016, 8, 401–409. [Google Scholar]
- Shaban, S.; Patel, M.; Ahmad, A. Improved efficacy of antifungal drugs in combination with monoterpene phenols against Candida auris. Sci. Rep. 2020, 10, 1162. [Google Scholar] [CrossRef] [PubMed]
- Rocha, M.F.G.; Sales, J.A.; da Rocha, M.G.; Galdino, L.M.; de Aguiar, L.; Pereira-Neto, W.d.A.; de Aguiar Cordeiro, R.; Castelo-Branco, D.d.S.C.M.; Sidrim, J.J.C.; Brilhante, R.S.N. Antifungal effects of the flavonoids kaempferol and quercetin: A possible alternative for the control of fungal biofilms. Biofouling 2019, 35, 320–328. [Google Scholar] [CrossRef]
- Khanzada, B.; Akhtar, N.; Okla, M.K.; Alamri, S.A.; Al-Hashimi, A.; Baig, M.W.; Rubnawaz, S.; AbdElgawad, H.; Hirad, A.H.; Haq, I.-U.; et al. Profiling of Antifungal Activities and In Silico Studies of Natural Polyphenols from Some Plants. Molecules 2021, 26, 7164. [Google Scholar] [CrossRef] [PubMed]
- Lotfi, M.; Kazemi, S.; Ebrahimpour, A.; Shirafkan, F.; Pirzadeh, M.; Hosseini, M.; Moghadamnia, A.A. Protective Effect of Quercetin Nanoemulsion on 5-Fluorouracil-Induced Oral Mucositis in Mice. J. Oncol. 2021, 2021, 5598230. [Google Scholar] [CrossRef]
- Ingrosso, G.; Saldi, S.; Marani, S.; Wong, A.Y.W.; Bertelli, M.; Aristei, C.; Zelante, T. Breakdown of Symbiosis in Radiation-Induced Oral Mucositis. J. Fungi 2021, 7, 290. [Google Scholar] [CrossRef]
- Jacob, S.; Nair, A.B.; Shah, J. Emerging role of nanosuspensions in drug delivery systems. Biomater. Res. 2020, 24, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melkoumov, A.; Goupil, M.; Louhichi, F.; Raymond, M.; de Repentigny, L.; Leclair, G. Nystatin nanosizing enhances in vitro and in vivo antifungal activity against Candida albicans. J. Antimicrob. Chemother. 2013, 68, 2099–2105. [Google Scholar] [CrossRef] [Green Version]
- Authimoolam, S.P.; Dziubla, T.D. Biopolymeric Mucin and Synthetic Polymer Analogs: Their Structure, Function and Role in Biomedical Applications. Polymers 2016, 8, 71. [Google Scholar] [CrossRef] [Green Version]
- Pornpitchanarong, C.; Rojanarata, T.; Opanasopit, P.; Ngawhirunpat, T.; Patrojanasophon, P. Clotrimazole nanosuspensions-loaded hyaluronic acid-catechol/polyvinyl alcohol mucoadhesive films for oral candidiasis treatment. J. Drug Deliv. Sci. Technol. 2020, 60, 101927. [Google Scholar] [CrossRef]
- Kraisit, P.; Hirun, N.; Mahadlek, J.; Limmatvapirat, S. Fluconazole-loaded solid lipid nanoparticles (SLNs) as a potential carrier for buccal drug delivery of oral candidiasis treatment using the Box-Behnken design. J. Drug Deliv. Sci. Technol. 2021, 63, 102437. [Google Scholar] [CrossRef]
- Mendes, A.I.; Silva, A.C.; Catita, J.A.M.; Cerqueira, F.; Gabriel, C.; Lopes, C.M. Miconazole-loaded nanostructured lipid carriers (NLC) for local delivery to the oral mucosa: Improving antifungal activity. Colloids Surf. B Biointerfaces 2013, 111, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Hosny, K.M.; Sindi, A.M.; Ali, S.; Alharbi, W.S.; Hajjaj, M.S.; Bukhary, H.A.; Badr, M.Y.; Mushtaq, R.Y.; Murshid, S.S.A.; Almehmady, A.M.; et al. Development, optimization, and evaluation of a nanostructured lipid carrier of sesame oil loaded with miconazole for the treatment of oral candidiasis. Drug Deliv. 2022, 29, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Castillo, H.A.P.; Castellanos, L.N.M.; Chamoro, R.M.; Martinez, R.R.; Borunda, E. Nanoparticles as New Therapeutic Agents against Candida albicans. In Candida albicans; Sandai, D., Ed.; IntechOpen: London, UK, 2018; pp. 145–171. ISBN 978-1-83880-160-1. [Google Scholar]
- McCarron, P.A.; Donnelly, R.F.; Marouf, W.; Calvert, D.E. Anti-adherent and antifungal activities of surfactant-coated poly(ethylcyanoacrylate) nanoparticles. Int. J. Pharm. 2007, 340, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Lara, H.H.; Guisbiers, G.; Mendoza, J.; Mimun, L.C.; Vincent, B.A.; Lopez-Ribot, J.L.; Nash, K.L. Synergistic antifungal effect of chitosan-stabilized selenium nanoparticles synthesized by pulsed laser ablation in liquids against Candida albicans biofilms. Int. J. Nanomed. 2018, 13, 2697–2708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Alcântara Sica de Toledo, L.; Rosseto, H.C.; dos Santos, R.S.; Spizzo, F.; Del Bianco, L.; Montanha, M.C.; Esposito, E.; Kimura, E.; Bonfim-Mendonça, P.d.S.; Svidzinski, T.I.E.; et al. Thermal Magnetic Field Activated Propolis Release From Liquid Crystalline System Based on Magnetic Nanoparticles. AAPS PharmSciTech 2018, 19, 3258–3271. [Google Scholar] [CrossRef] [PubMed]
- Niemirowicz, K.; Durnaś, B.; Tokajuk, G.; Piktel, E.; Michalak, G.; Gu, X.; Kułakowska, A.; Savage, P.B.; Bucki, R. Formulation and candidacidal activity of magnetic nanoparticles coated with cathelicidin LL-37 and ceragenin CSA-13. Sci. Rep. 2017, 7, 4610. [Google Scholar] [CrossRef]
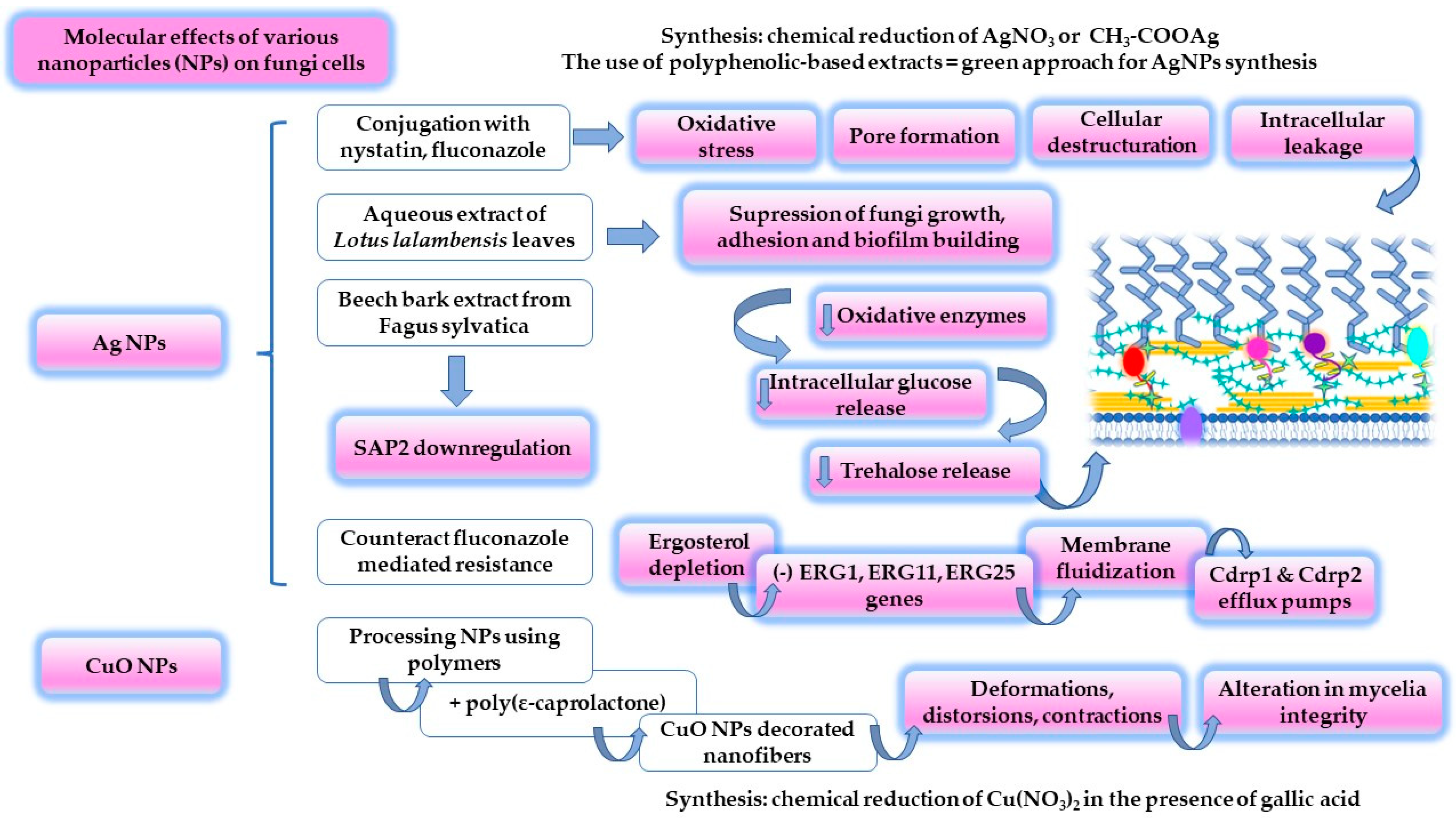
- Hussain, M.A.; Ahmed, D.; Anwar, A.; Perveen, S.; Ahmed, S.; Anis, I.; Shah, M.R.; Khan, N.A. Combination Therapy of Clinically Approved Antifungal Drugs Is Enhanced by Conjugation with Silver Nanoparticles. Int. Microbiol. 2019, 22, 239–246. [Google Scholar] [CrossRef]
- Pimentel, B.N.A.d.S.; de Foggi, C.C.; Barbugli, P.A.; de Oliveira, R.C.; de Avila, E.D.; Longo, E.; Vergani, C.E. Antifungal activity and biocompatibility of α-AgVO(3) microcrystals: A promising material against oral Candida disease. Mater. Sci. Eng. C Mater. Biol. Appl. 2020, 108, 110405. [Google Scholar] [CrossRef]
- Naikoo, G.A.; Mustaqeem, M.; Hassan, I.U.; Awan, T.; Arshad, F.; Salim, H.; Qurashi, A. Bioinspired and green synthesis of nanoparticles from plant extracts with antiviral and antimicrobial properties: A critical review. J. Saudi Chem. Soc. 2021, 25, 101304. [Google Scholar] [CrossRef]
- Adam, R.Z.; Khan, S.B. Antimicrobial efficacy of silver nanoparticles against Candida albicans: A systematic review protocol. PLoS ONE 2021, 16, e0245811. [Google Scholar] [CrossRef]
- Jalalvand, A.R.; Zhaleh, M.; Goorani, S.; Zangeneh, M.M.; Seydi, N.; Zangeneh, A.; Moradi, R. Chemical characterization and antioxidant, cytotoxic, antibacterial, and antifungal properties of ethanolic extract of Allium Saralicum R.M. Fritsch leaves rich in linolenic acid, methyl ester. J. Photochem. Photobiol. B Biol. 2019, 192, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, B.M.; Ali, E.M. Green Synthesis of Silver Nanoparticles Using the Lotus lalambensis Aqueous Leaf Extract and Their Anti-Candidal Activity against Oral Candidiasis. ACS Omega 2021, 6, 8151–8162. [Google Scholar] [CrossRef] [PubMed]
- Mare, A.D.; Ciurea, C.N.; Man, A.; Mareș, M.; Toma, F.; Berța, L.; Tanase, C. In Vitro Antifungal Activity of Silver Nanoparticles Biosynthesized with Beech Bark Extract. Plants 2021, 10, 2153. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Escobar, A.; Reyes-López, S.Y. Antifungal susceptibility of Candida species to copper oxide nanoparticles on polycaprolactone fibers (PCL-CuONPs). PLoS ONE 2020, 15, e0228864. [Google Scholar] [CrossRef]
- Ahmadpour Kermani, S.; Salari, S.; Ghasemi Nejad Almani, P. Comparison of antifungal and cytotoxicity activities of titanium dioxide and zinc oxide nanoparticles with amphotericin B against different Candida species: In vitro evaluation. J. Clin. Lab. Anal. 2021, 35, e23577. [Google Scholar] [CrossRef]
- Gerstner, E. Nobel Prize 2010: Andre Geim & Konstantin Novoselov. Nat. Phys. 2010, 6, 836. [Google Scholar] [CrossRef]
- Armano, A.; Agnello, S. Two-Dimensional Carbon: A Review of Synthesis Methods, and Electronic, Optical, and Vibrational Properties of Single-Layer Graphene. C 2019, 5, 67. [Google Scholar] [CrossRef] [Green Version]
- Papageorgiou, D.G.; Kinloch, I.A.; Young, R.J. Mechanical properties of graphene and graphene-based nanocomposites. Prog. Mater. Sci. 2017, 90, 75–127. [Google Scholar] [CrossRef]
- Chung, H.-C.; Chang, C.-P.; Lin, C.-Y.; Lin, M.-F. Electronic and optical properties of graphene nanoribbons in external fields. Phys. Chem. Chem. Phys. 2016, 18, 7573–7616. [Google Scholar] [CrossRef] [Green Version]
- Shang, L.; Qi, Y.; Lu, H.; Pei, H.; Li, Y.; Qu, L.; Wu, Z.; Zhang, W. 7—Graphene and Graphene Oxide for Tissue Engineering and Regeneration. In Theranostic Bionanomaterials Micro and Nano Technologies; Cui, W., Zhao, X., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 165–185. ISBN 978-0-12-815341-3. [Google Scholar]
- Park, C.; Park, S.; Lee, D.; Choi, K.S.; Lim, H.-P.; Kim, J. Graphene as an Enabling Strategy for Dental Implant and Tissue Regeneration. Tissue Eng. Regen. Med. 2017, 14, 481–493. [Google Scholar] [CrossRef]
- Peña-Bahamonde, J.; Nguyen, H.N.; Fanourakis, S.K.; Rodrigues, D.F. Recent advances in graphene-based biosensor technology with applications in life sciences. J. Nanobiotechnol. 2018, 16, 75. [Google Scholar] [CrossRef] [Green Version]
- Shu, M.; Gao, F.; Zeng, M.; Yu, C.; Wang, X.; Huang, R.; Yang, J.; Su, Y.; Hu, N.; Zhou, Z.; et al. Microwave-Assisted Chitosan-Functionalized Graphene Oxide as Controlled Intracellular Drug Delivery Nanosystem for Synergistic Antitumour Activity. Nanoscale Res. Lett. 2021, 16, 75. [Google Scholar] [CrossRef] [PubMed]
- Farzanegan, A.; Roudbary, M.; Falahati, M.; Khoobi, M.; Gholibegloo, E.; Farahyar, S.; Karimi, P.; Khanmohammadi, M. Synthesis, characterization and antifungal activity of a novel formulated nanocomposite containing Indolicidin and Graphene oxide against disseminated candidiasis. J. Mycol. Med. 2018, 28, 628–636. [Google Scholar] [CrossRef] [PubMed]
- Hummers, W.S.; Offeman, R.E. Preparation of Graphitic Oxide. J. Am. Chem. Soc. 1958, 80, 1339. [Google Scholar] [CrossRef]
- Cui, J.; Yang, Y.; Zheng, M.; Liu, Y.; Xiao, Y.; Lei, B.; Chen, W. Facile fabrication of graphene oxide loaded with silver nanoparticles as antifungal materials. Mater. Res. Express 2014, 1, 45007. [Google Scholar] [CrossRef]
- Liu, C.; Tan, D.; Chen, X.; Liao, J.; Wu, L. Research on Graphene and Its Derivatives in Oral Disease Treatment. Int. J. Mol. Sci. 2022, 23, 4737. [Google Scholar] [CrossRef]
- Kashyap, S.; Mishra, S.; Behera, S.K. Aqueous Colloidal Stability of Graphene Oxide and Chemically Converted Graphene. J. Nanoparticles 2014, 2014, 640281. [Google Scholar] [CrossRef] [Green Version]
- Bregnocchi, A.; Zanni, E.; Uccelletti, D.; Marra, F.; Cavallini, D.; De Angelis, F.; De Bellis, G.; Bossù, M.; Ierardo, G.; Polimeni, A.; et al. Graphene-based dental adhesive with anti-biofilm activity. J. Nanobiotechnol. 2017, 15, 89. [Google Scholar] [CrossRef]
- Alnatheer, M.; Alqerban, A.; Alhazmi, H. Graphene oxide-modified dental adhesive for bonding orthodontic brackets. Int. J. Adhes. Adhes. 2021, 110, 102928. [Google Scholar] [CrossRef]
- Radhi, A.; Mohamad, D.; Abdul Rahman, F.S.; Abdullah, A.M.; Hasan, H. Mechanism and factors influence of graphene-based nanomaterials antimicrobial activities and application in dentistry. J. Mater. Res. Technol. 2021, 11, 1290–1307. [Google Scholar] [CrossRef]
- Seifi, T.; Kamali, A.R. Anti-pathogenic activity of graphene nanomaterials: A review. Colloids Surf. B Biointerfaces 2021, 199, 111509. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wang, X.; Chen, F.; Zhang, C.; Zhi, X.; Wang, K.; Cui, D. The antifungal activity of graphene oxide-silver nanocomposites. Biomaterials 2013, 34, 3882–3890. [Google Scholar] [CrossRef] [PubMed]
- Cheong, Y.-K.; Arce, M.P.; Benito, A.; Chen, D.; Luengo Crisóstomo, N.; Kerai, L.V.; Rodríguez, G.; Valverde, J.L.; Vadalia, M.; Cerpa-Naranjo, A.; et al. Synergistic Antifungal Study of PEGylated Graphene Oxides and Copper Nanoparticles against Candida albicans. Nanomaterials 2020, 10, 819. [Google Scholar] [CrossRef] [PubMed]
- Asadi Shahi, S.; Roudbar Mohammadi, S.; Roudbary, M.; Delavari, H. A new formulation of graphene oxide/fluconazole compound as a promising agent against Candida albicans. Prog. Biomater. 2019, 8, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Jacobsen, J.; Genina, N.; Larsen, S.W.; Nielsen, H.M.; Müllertz, A.; Mu, H. Investigating the effect of graphene oxide in chitosan/alginate-based foams on the release and antifungal activity of clotrimazole in vitro. Eur. J. Pharm. Sci. Off. J. Eur. Fed. Pharm. Sci. 2022, 174, 106204. [Google Scholar] [CrossRef] [PubMed]
- Velazco-Medel, M.A.; Camacho-Cruz, L.A.; Lugo-González, J.C.; Bucio, E. Antifungal polymers for medical applications. Med. Devices Sens. 2021, 4, e10134. [Google Scholar] [CrossRef]
- Maddu, N. Functions of Saliva. In Functions of Saliva, Saliva and Salivary Diagnostics; Gokul, S., Ed.; IntechOpen: Rijeka, Croatia, 2019; ISBN 978-1-78984-309-5. [Google Scholar]
- Punj, A. Secretions of Human Salivary Gland. In Salivary Glands: New Approaches in Diagnostics and Treatment; Güvenç, I.A., Ed.; IntechOpen: London, UK, 2019; ISBN 978-1-78984-988-2. [Google Scholar]
- Madhav, N.V.S.; Shakya, A.K.; Shakya, P.; Singh, K. Orotransmucosal drug delivery systems: A review. J. Control. Release 2009, 140, 2–11. [Google Scholar] [CrossRef]
- Khutoryanskiy, V.V. Advances in mucoadhesion and mucoadhesive polymers. Macromol. Biosci. 2011, 11, 748–764. [Google Scholar] [CrossRef]
- Brahmbhatt, D. Bioadhesive drug delivery systems: Overview and recent advances. Int. J. Chem. Life Sci. 2017, 6, 2016. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.; Pal, K.; Anis, A.; Pramanik, K.; Prabhakar, B. Polymers in Mucoadhesive Drug-Delivery Systems: A Brief Note. Des. Monomers Polym. 2009, 12, 483–495. [Google Scholar] [CrossRef] [Green Version]
- Alkhalidi, H.M.; Hosny, K.M.; Rizg, W.Y. Oral Gel Loaded by Fluconazole—Sesame Oil Nanotransfersomes: Development, Optimization, and Assessment of Antifungal Activity. Pharmaceutics 2020, 13, 27. [Google Scholar] [CrossRef] [PubMed]
- Rank, L.A.; Walsh, N.M.; Lim, F.Y.; Gellman, S.H.; Keller, N.P.; Hull, C.M. Peptide-Like Nylon-3 Polymers with Activity against Phylogenetically Diverse, Intrinsically Drug-Resistant Pathogenic Fungi. mSphere 2018, 3, e00223-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ntow-Boahene, W.; Cook, D.; Good, L. Antifungal Polymeric Materials and Nanocomposites. Front. Bioeng. Biotechnol. 2021, 9, 780328. [Google Scholar] [CrossRef]
- Liu, R.; Chen, X.; Falk, S.P.; Mowery, B.P.; Karlsson, A.J.; Weisblum, B.; Palecek, S.P.; Masters, K.S.; Gellman, S.H. Structure–Activity Relationships among Antifungal Nylon-3 Polymers: Identification of Materials Active against Drug-Resistant Strains of Candida albicans. J. Am. Chem. Soc. 2014, 136, 4333–4342. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Ling, P.; Zhang, T. Polymeric micelles, a promising drug delivery system to enhance bioavailability of poorly water-soluble drugs. J. Drug Deliv. 2013, 2013, 340315. [Google Scholar] [CrossRef]
- Suksiriworapong, J.; Mingkwan, T.; Chantasart, D. Enhanced transmucosal delivery of itraconazole by thiolated d-ɑ-tocopheryl poly(ethylene glycol) 1000 succinate micelles for the treatment of Candida albicans. Eur. J. Pharm. Biopharm. 2017, 120, 107–115. [Google Scholar] [CrossRef]
- Shahid, A.; Aslam, B.; Muzammil, S.; Aslam, N.; Shahid, M.; Almatroudi, A.; Allemailem, K.S.; Saqalein, M.; Nisar, M.A.; Rasool, M.H.; et al. The prospects of antimicrobial coated medical implants. J. Appl. Biomater. Funct. Mater. 2021, 19, 22808000211040304. [Google Scholar] [CrossRef]
- Barros, J.; Dias, A.; Rodrigues, M.A.; Pina-Vaz, C.; Lopes, M.A.; Pina-Vaz, I. Antibiofilm and Antimicrobial Activity of Polyethylenimine: An Interesting Compound for Endodontic Treatment. J. Contemp. Dent. Pract. 2015, 16, 427–432. [Google Scholar] [CrossRef]
- Zhou, L.; Li, A.; Wang, H.; Sun, W.; Zuo, S.; Li, C. Preparation and characterization of luteolin-loaded MPEG-PCL-g-PEI micelles for oral Candida albicans infection. J. Drug Deliv. Sci. Technol. 2021, 63, 102454. [Google Scholar] [CrossRef]
- Kraisit, P.; Yonemochi, E.; Furuishi, T.; Mahadlek, J.; Limmatvapirat, S. Chitosan film containing antifungal agent-loaded SLNs for the treatment of candidiasis using a Box-Behnken design. Carbohydr. Polym. 2022, 283, 119178. [Google Scholar] [CrossRef]
- Irimia, T.; Dinu-Pîrvu, C.-E.; Ghica, M.V.; Lupuleasa, D.; Muntean, D.-L.; Udeanu, D.I.; Popa, L. Chitosan-Based In Situ Gels for Ocular Delivery of Therapeutics: A State-of-the-Art Review. Mar. Drugs 2018, 16, 373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popa, L.; Ghica, M.V.; Popescu, R.; Irimia, T.; Dinu-Pîrvu, C.-E. Development and Optimization of Chitosan-Hydroxypropyl Methylcellulose In Situ Gelling Systems for Ophthalmic Delivery of Bupivacaine Hydrochloride. Processes 2021, 9, 1694. [Google Scholar] [CrossRef]
- Jia, R.; Duan, Y.; Fang, Q.; Wang, X.; Huang, J. Pyridine-grafted chitosan derivative as an antifungal agent. Food Chem. 2016, 196, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Tabriz, A.; Ur Rehman Alvi, M.A.; Khan Niazi, M.B.; Batool, M.; Bhatti, M.F.; Khan, A.L.; Khan, A.U.; Jamil, T.; Ahmad, N.M. Quaternized trimethyl functionalized chitosan based antifungal membranes for drinking water treatment. Carbohydr. Polym. 2019, 207, 17–25. [Google Scholar] [CrossRef]
- Al-Ani, E.; Heaselgrave, W. The Investigation of Thymol Formulations Containing Poloxamer 407 and Hydroxypropyl Methylcellulose to Inhibit Candida Biofilm Formation and Demonstrate Improved Bio-Compatibility. Pharmaceuticals 2022, 15, 71. [Google Scholar] [CrossRef]
- An, S.-H.; Ban, E.; Chung, I.-Y.; Cho, Y.-H.; Kim, A. Antimicrobial Activities of Propolis in Poloxamer Based Topical Gels. Pharmaceutics 2021, 13, 2021. [Google Scholar] [CrossRef]
- Kianfar, F.; Ayensu, I.; Boateng, J.S. Development and physico-mechanical characterization of carrageenan and poloxamer-based lyophilized matrix as a potential buccal drug delivery system. Drug Dev. Ind. Pharm. 2014, 40, 361–369. [Google Scholar] [CrossRef]
- Chen, Y.; Lee, J.-H.; Meng, M.; Cui, N.; Dai, C.-Y.; Jia, Q.; Lee, E.-S.; Jiang, H.-B. An Overview on Thermosensitive Oral Gel Based on Poloxamer 407. Materials 2021, 14, 4522. [Google Scholar] [CrossRef]
- Ribeiro, F.d.C.; Junqueira, J.C.; Dos Santos, J.D.; de Barros, P.P.; Rossoni, R.D.; Shukla, S.; Fuchs, B.B.; Shukla, A.; Mylonakis, E. Development of Probiotic Formulations for Oral Candidiasis Prevention: Gellan Gum as a Carrier To Deliver Lactobacillus paracasei 28.4. Antimicrob. Agents Chemother. 2020, 64, e02323-19. [Google Scholar] [CrossRef]
- Rossoni, R.D.; de Barros, P.P.; de Alvarenga, J.A.; Ribeiro, F.d.C.; Velloso, M.d.S.; Fuchs, B.B.; Mylonakis, E.; Jorge, A.O.C.; Junqueira, J.C. Antifungal activity of clinical Lactobacillus strains against Candida albicans biofilms: Identification of potential probiotic candidates to prevent oral candidiasis. Biofouling 2018, 34, 212–225. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
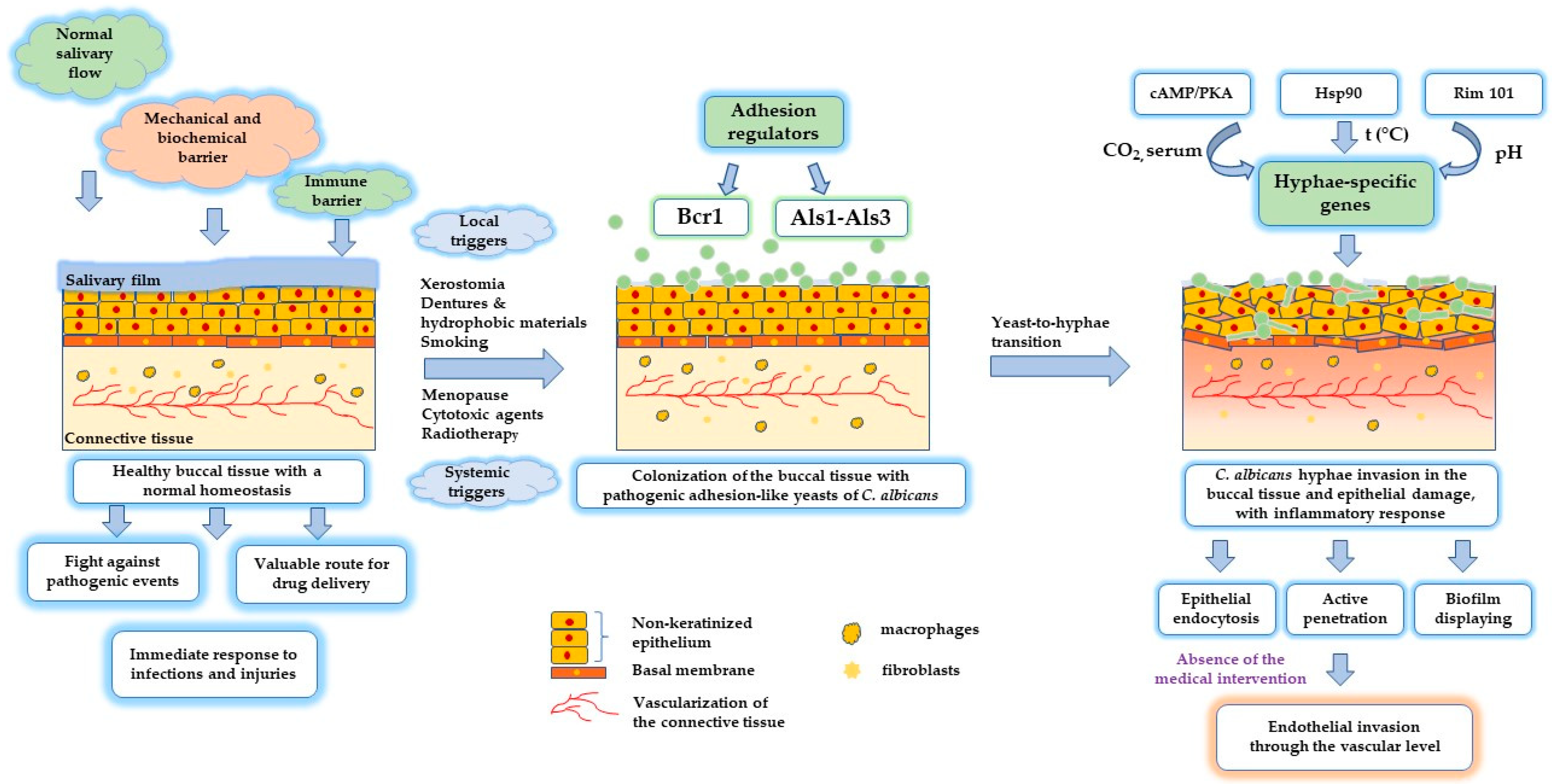
| Local Factors | ||
|---|---|---|
| Factors | Surface and Molecular Effects | Ref. |
| Imbalances in salivary flow | A high number of C. albicans colonies were found in patients with xerostomia, which indicates alterations in normal microbiota. These changes are signaled by a modified pH in oral mucosa and the loss of antimicrobial proteins such as lysozyme, lactoperoxidase, immunoglobulin, histatins, and lactoferrin. | [27,43,44] |
| Dentures | Dentures decrease O2 supply at the epithelial level and reduce the salivary flow, creating a favorable acidic medium for yeast development. The fungi have an increased affinity for the roughened hydrophobic surfaces of acrylic resins of dentures, encouraging biofilm generation and the initiation of stomatitis. The adhesion process is promoted by a gradual replacement of interfacial water. Intimate mechanisms involved in biofilm formation are the roughness of the surface, hydrophobicity, and electrostatic nature of interactions prior to promoting protein adsorption and adhesion, Lifshitz–van der Waals forces, Brownian motion, and receptor–ligand binding. | [28,45,46] |
| Prosthesis | Dental implants favor the localization of C. albicans in the subgingival sulcus and the initiation of pathogenic periodontitis (peri-implantitis). Poor oral hygiene acts in the sense of promoting colonization of periodontal pathogens. The virulence of C. albicans is related to the activity of aspartyl proteinase as a promotor for adhesion. In addition, secretion of candidalysin will harm the epithelial cells and bind the epithelial growth factor 1 ErbB1 (Her1). The damage to the tissues of the peri-implant area is favored by metallopeptidase (95 kDa), targeting type I collagen, type IV collagen, fibronectin, and basement membrane. | [29] |
| Pre-existing oral pathologies | Denture stomatitis is promoted by modification in E-cadherin, collagen VII and fibronectin, combined with the presence of C. albicans colonies in the oral mucosa. Biopsy studies highlighted disorganization of the epithelial cells, with an irregular arrangement of keratinocytes and inflammatory phenomena in connective tissue. | [36,46] |
| Poor epithelial local defense | Reduced response of the host immune defense elements (Toll-like receptors, C-type lectin receptors, and 2 NOD-like receptors) induces dissemination of virulence factors, specific for C. albicans growth: dimorphism, adhesion, phenotypic switching, polymorphism, and secretion of hydrolytic enzymes such as lipases, phospholipases, and proteinases. A deficiency of epithelial antimicrobial peptides (AMP) was correlated with candidiasis development. | [31,47,48] |
| Oral dysbiosis | The oral microbiome covers a large number of microorganisms, up to 700 species, of which more than 60 are fungi species. A reduction in the number of native fungi that normally harbor the buccal mucosa was associated with the risk of developing oral infections. The interactions between fungal entities and bacteria such as streptococci favor the development of mixed biofilms and modulate the mechanisms implied in polymorphism and host immune response. The interactions of streptococci with C. albicans are made through the cell wall surface proteins of the Csh protein family and streptococcal surface proteins A and B. For its part, C. albicans supports the interactions through 3 ALS1, 4 ALS3, and 5 HWP1. Other interactions are governed by carbohydrate and extracellular polysaccharides, quorum-sensing molecules, and metabolic events. Dietary sucrose increases C. albicans virulence and the symbiotic relationship with streptococcus species. | [32,33,49] |
| Inhalator corticosteroids | The treatment with inhalator corticosteroids causes a poor epithelial local defense of the immune system and was thought to elevate salivary glucose levels as a substrate for fungi growth. Oral candidiasis development was dependent on dose and the device used in administration. | [34,50] |
| Smoking | Several theories consider the epithelial damage induced by smoking. In a more profound understanding, a concentration of 1–2 mg/mL of nicotine was found to assure the process of fungi cell multiplication, and it was correlated with an increase in HWP1 and ALS3 expression, implied in hyphae expansion and biofilm formation. | [16,35] |
| Carbohydrate-based diet | The intake of dietary and sugar-based foods represents a substrate for candidiasis development. High glucose levels in diabetic patients influence oral candidiasis development as well. Low blood glucose levels (~0.1%) stimulate hyphae growing as the invasive form of candidiasis. This is the result of the regulation of secreted aspartyl proteinases (SAP) genes under the signals of the 6 cAMP/PKA pathway and 7 MAP kinase cascade. MAP kinase pathway was recognized as a trigger for adhesion, invasion, and reorganization of the epithelial level. | [16,50,51] |
| Systemic Factors | ||
| Factors | Surface and Molecular Effects | Ref. |
| Vitamin and mineral deficiencies | Candidiasis development can be promoted by vitamin A, B6, and B12 deficiency, iron deficiency, or a reduced level of essential fatty acids, folic acid, magnesium, selenium, or zinc. Concerning iron metabolism, it represents an essential element for cell differentiation, oxygen transport, and the normal activity of immune cells. The atrophy of oral mucosa and candidiasis were frequently discovered in anemic patients. | [16,37,52] |
| Metabolic disorders | Diabetic patients are highly predisposed to oral candidiasis, which can be seen as a pathological result of an accumulation of factors: poor oral hygiene, xerostomia, pH imbalances, increase in serum glucose levels, and poor epithelial local defense alike. | [38,53] |
| Menopause | A decrease in estrogen hormone levels in menopausal women can counteract the normal state of the oral mucosa. A hormonal change can induce a cascade of local effects involving the modification in salivary secretion, lysozyme decrease, poor local immune activity, and an increase in oxidative stress. | [39] |
| HIV immunodeficiency | Immunosuppression that can be quantified by a decreased number of CD4+ immune cells entails an increased risk of developing candidiasis. A decrease in histatins level contributes to the severity of the pathology. | [18,21,40] |
| Prolonged antibiotherapy | Administration of broad-spectrum antibiotics yields dysbiosis, affecting the normal oral flora and transforming the commensal microorganisms into pathogenic entities. Imbalances in oral microbiota were related to a decrease in salivary antibody content. Salivary proteins expressed as mucins, salivary IgA, cystatin S, basic proline-rich proteins, or statherins are implied in a dynamic process concerning adhesion/aggregation/clearance of fungal cells. | [21,54,55] |
| Immunosuppressive treatments | Immunosuppressive and cytotoxic treatments of malignancies promote a weakening of the immune system, with repercussions for epithelial cell defense. Resistance to antifungal therapy was observed due to the formation of biofilms with persistent C. albicans or C. glabrata cells. | [41,42,56] |
| COVID-19 | COVID-19 induces immunosuppression by decreasing CD4+ and CD8+ T immune cells. In addition, candidiasis development in its invasive form has a multifactorial pattern, drawn by the presence of comorbidities (diabetes mellitus, pulmonary disorders, and malignancies) and concomitant treatments with immunosuppressants, corticosteroids, or antibiotics. Once more, a decrease in the salivary level of AMP was considered to be a robust marker for both superficial and intrusive infections. | [57,58] |
| Therapeutic Class | Antifungal Drugs— Main Compounds | Mechanisms of Action | Mechanisms of Resistance | Drug-Related Drawbacks | Ref. |
|---|---|---|---|---|---|
| Polyenes | Nystatin Amphotericin B Natamycin | Target the ergosterol sites and disrupt the cellular membrane, promoting fungicidal effects. Fungicidal action is enhanced by inducing oxidative stress. | Modification of enzymes with catalytic activity (C8-sterol isomerase and Δ5,6-desaturase), implied in ergosterol biosynthesis, via ERG2 and ERG3 gene alteration. Increase catalase function and deviate ROS generation. | Nystatin is used only for its local effect, without systemic absorption. It has an unpleasant taste in buccal administration. | [16,20,107,116] |
| Amphotericin B is nephrotoxic and preferred as second-line therapy. | |||||
| Poor bioavailability. | |||||
| Azoles | Clotrimazole Miconazole Fluconazole Posaconazole Itraconazole Voriconazole | Inhibition of cytochrome P450 enzymes implied in the biosynthesis of ergosterol from lanosterol, namely 14-α demethylase. The global effect is based on morphological alteration of fungi cells and inhibition of fungi growth. | Overexpression of CDR1 and CDR2 of the ATP-binding cassette superfamily, and MDR1 (major facilitator superfamily) genes. Use of efflux pumps by regulation of Tac1 and Mrr1 ATP binding cassette genes. Structural modification of demethylase enzyme. Overexpression of ERG11 gene. Cross-resistance with polyenes via ERG3 gene alteration. Drug engulfment into a complex glucan-based matrix of the biofilm. | Poor bioavailability, and buccal delivery of azoles is less efficient (multiple dosings and short local retention). | [113,117,118] |
| Echinocandins | Micafungin Caspofungin Anidulafungin | Fungicidal effects induced via inhibition of β (1–3) glucan synthase. Drug molecules target cell wall proteins implied in β (1–3) glucan synthesis. | Mutation of FKS1 gene specific for the catalytic unit of glucan synthase. Increased chitin synthesis. Drug engulfment into a complex glucan-based matrix of the biofilm. Imposition of adaptive mechanisms of fungi cells via Hsp90, cell wall integrity pathway, or high-osmolarity glycerol pathway. | Poor bioavailability, and the intravenous route (i.v.) is only accepted for systemic treatment. | [8,119] |
| Pyrimidines antimetabolite-like | 5-fluorocytosine | Cytosine permease entraps drug molecules into the cells. 5-fluorocytosine is bio transformed in 5-fluorouracil in the presence of cytosine deaminase. The new compound alters RNA synthesis and consequently protein synthesis. A second conversion through fluoro-deoxyuridylic acid determines DNA alteration. | Mutations in cytosine permease and cytosine deaminase enzymes. | Drug monitoring is mandatory for oral and i.v. route to avoid immunosuppression and hepatotoxicity. | [107,119,120] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anuța, V.; Talianu, M.-T.; Dinu-Pîrvu, C.-E.; Ghica, M.V.; Prisada, R.M.; Albu Kaya, M.G.; Popa, L. Molecular Mapping of Antifungal Mechanisms Accessing Biomaterials and New Agents to Target Oral Candidiasis. Int. J. Mol. Sci. 2022, 23, 7520. https://doi.org/10.3390/ijms23147520
Anuța V, Talianu M-T, Dinu-Pîrvu C-E, Ghica MV, Prisada RM, Albu Kaya MG, Popa L. Molecular Mapping of Antifungal Mechanisms Accessing Biomaterials and New Agents to Target Oral Candidiasis. International Journal of Molecular Sciences. 2022; 23(14):7520. https://doi.org/10.3390/ijms23147520
Chicago/Turabian StyleAnuța, Valentina, Marina-Theodora Talianu, Cristina-Elena Dinu-Pîrvu, Mihaela Violeta Ghica, Răzvan Mihai Prisada, Mădălina Georgiana Albu Kaya, and Lăcrămioara Popa. 2022. "Molecular Mapping of Antifungal Mechanisms Accessing Biomaterials and New Agents to Target Oral Candidiasis" International Journal of Molecular Sciences 23, no. 14: 7520. https://doi.org/10.3390/ijms23147520
APA StyleAnuța, V., Talianu, M. -T., Dinu-Pîrvu, C. -E., Ghica, M. V., Prisada, R. M., Albu Kaya, M. G., & Popa, L. (2022). Molecular Mapping of Antifungal Mechanisms Accessing Biomaterials and New Agents to Target Oral Candidiasis. International Journal of Molecular Sciences, 23(14), 7520. https://doi.org/10.3390/ijms23147520