
Antifungal Effect of Penicillamine Due to the Selective Targeting of L-Homoserine O-Acetyltransferase




Abstract
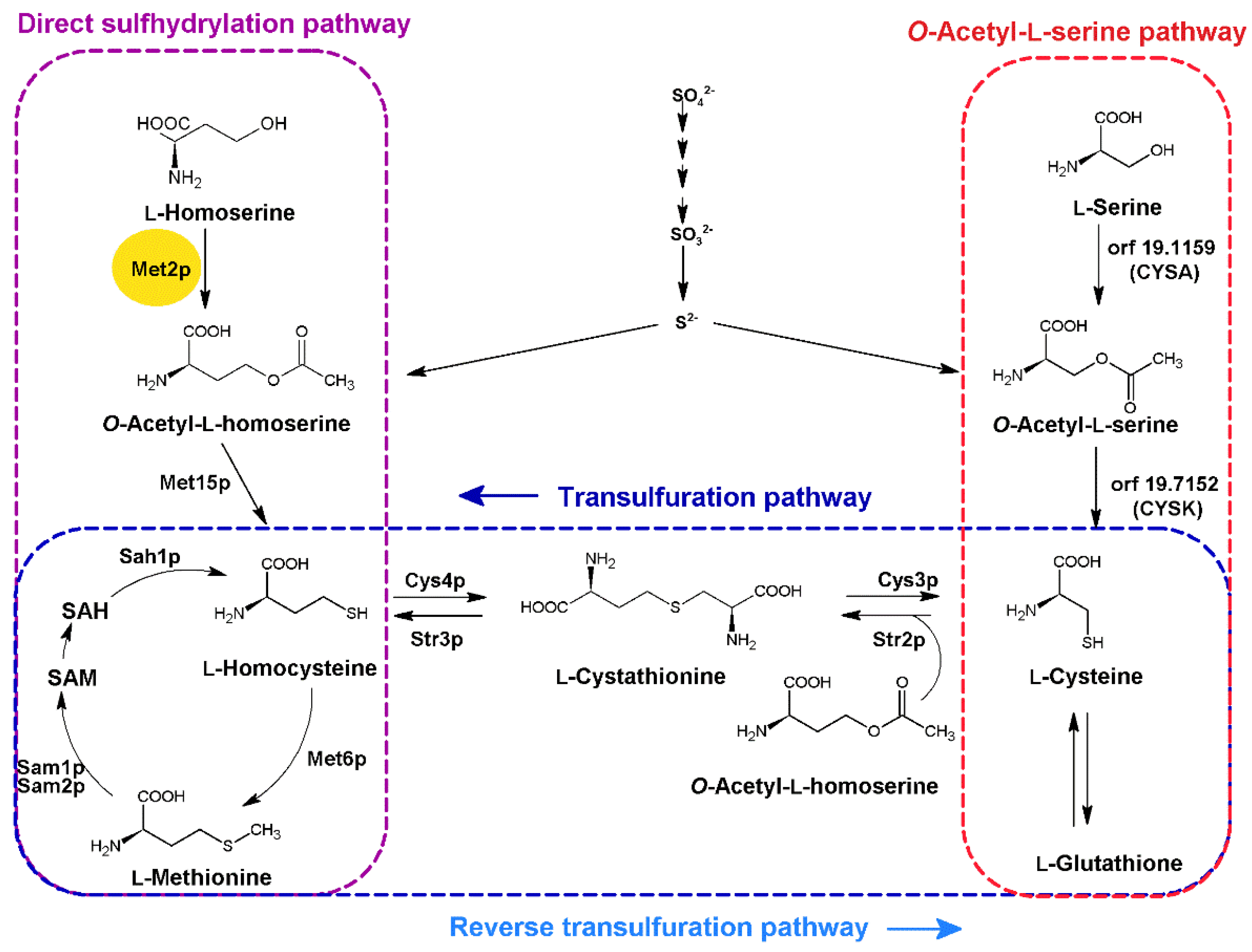
:1. Introduction
2. Results and Discussion
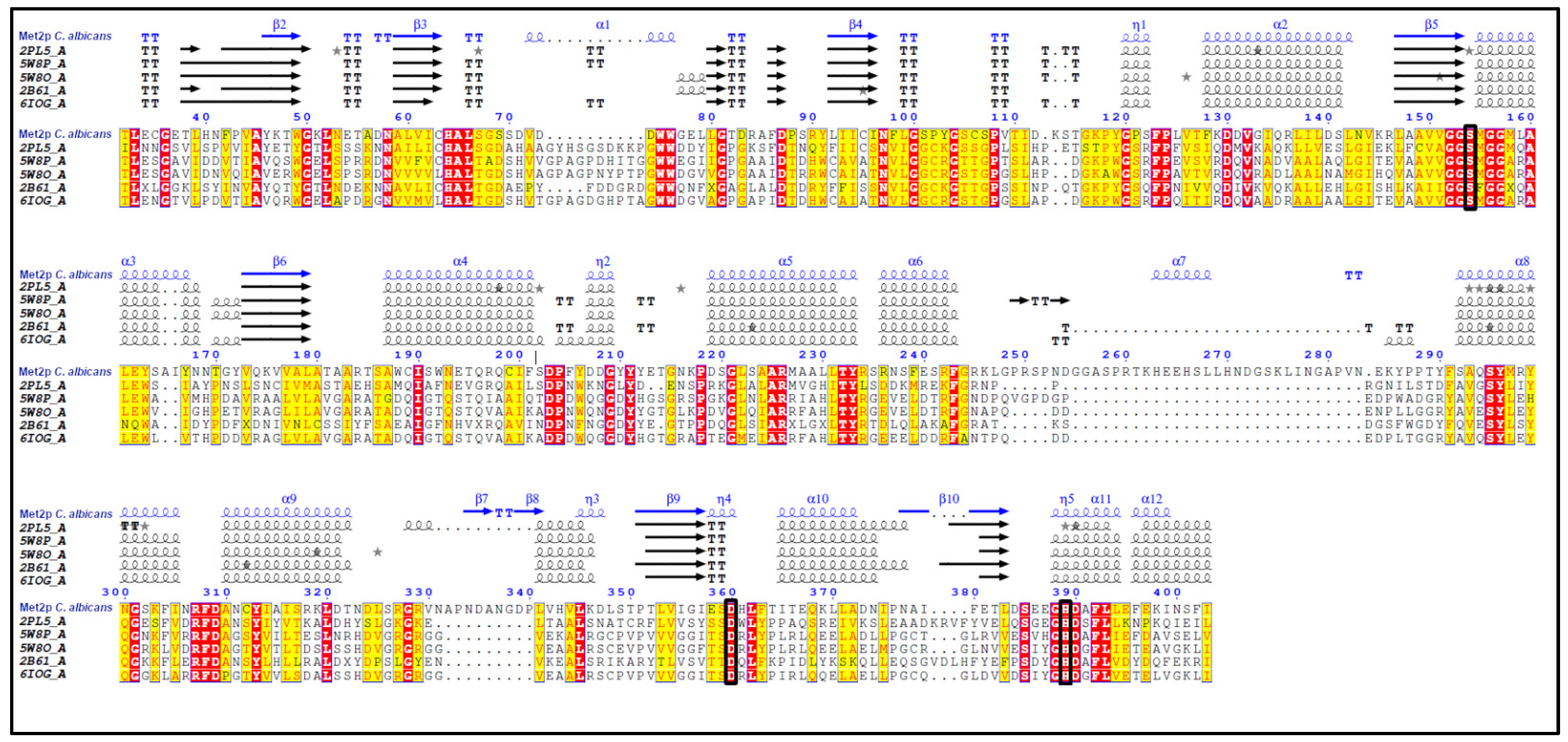
2.1. Identification of CaMET2 Gene and Bioinformatical Analysis of the Predicted Gene Product
2.2. Subcloning, Protein Expression, and Purification
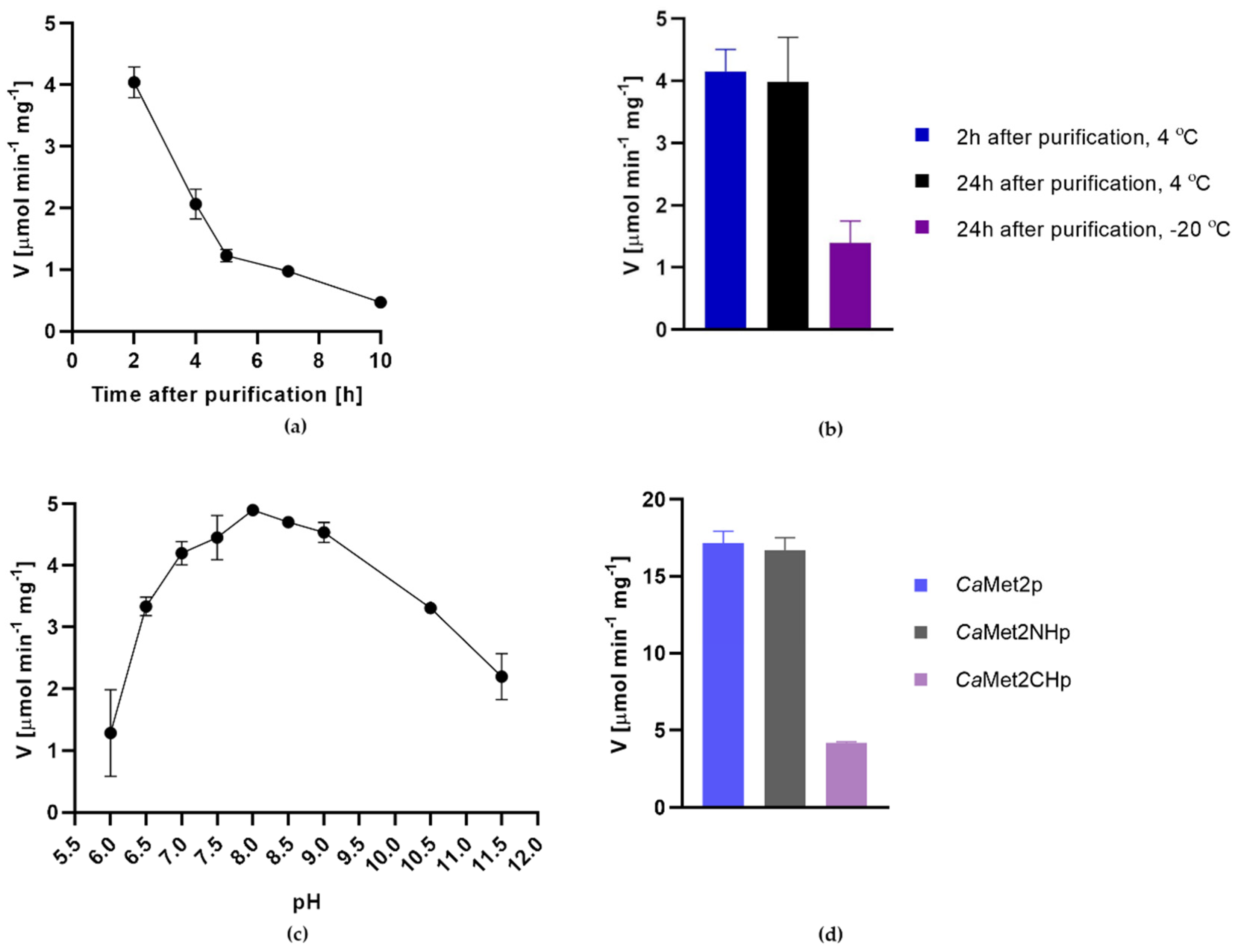
2.3. Characterization of CaMet2p Properties
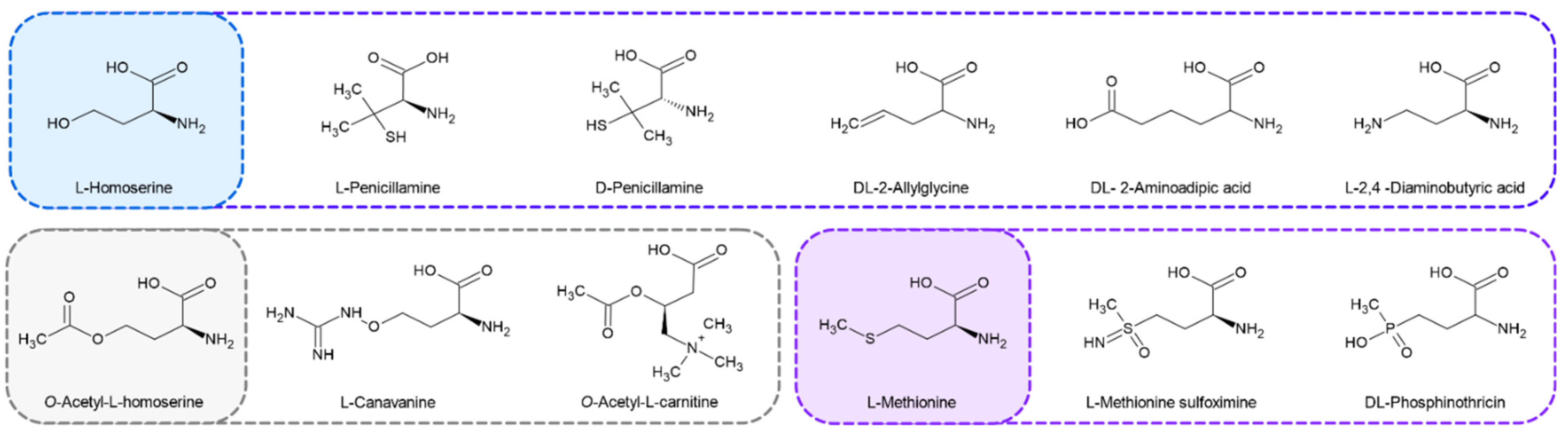
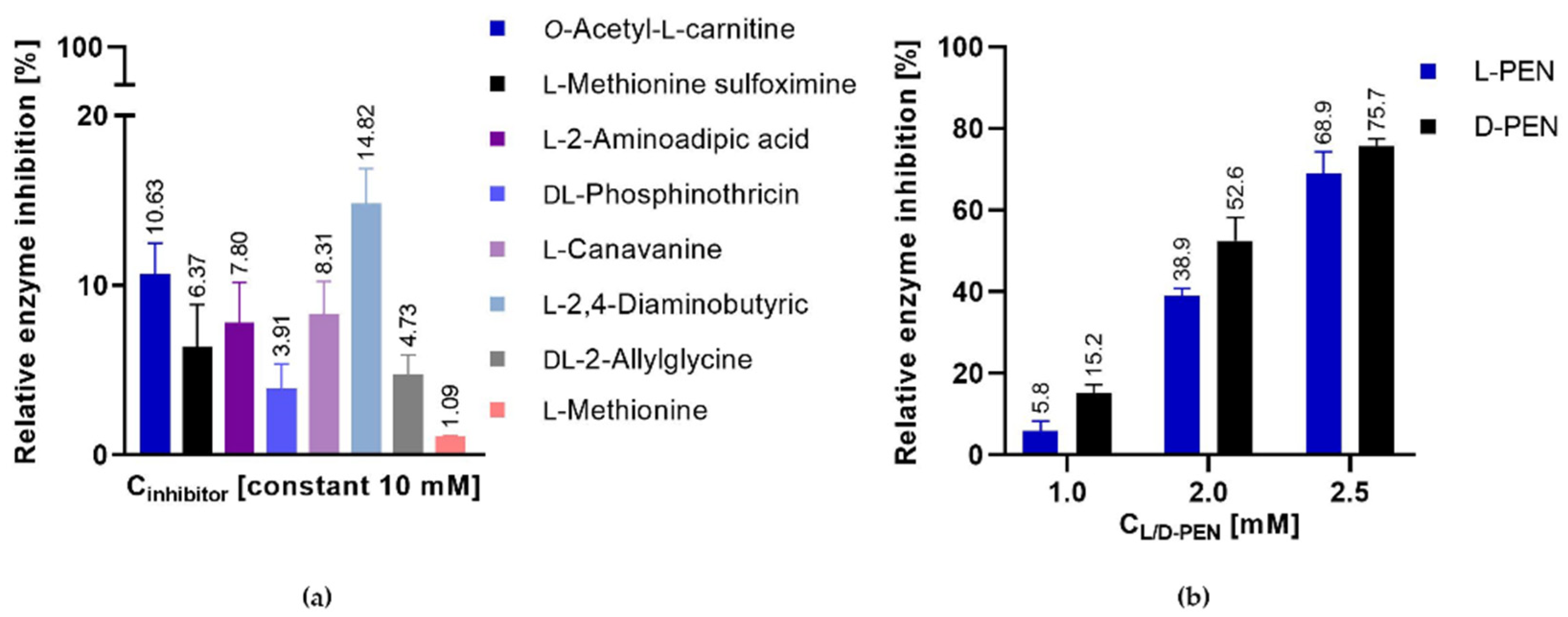
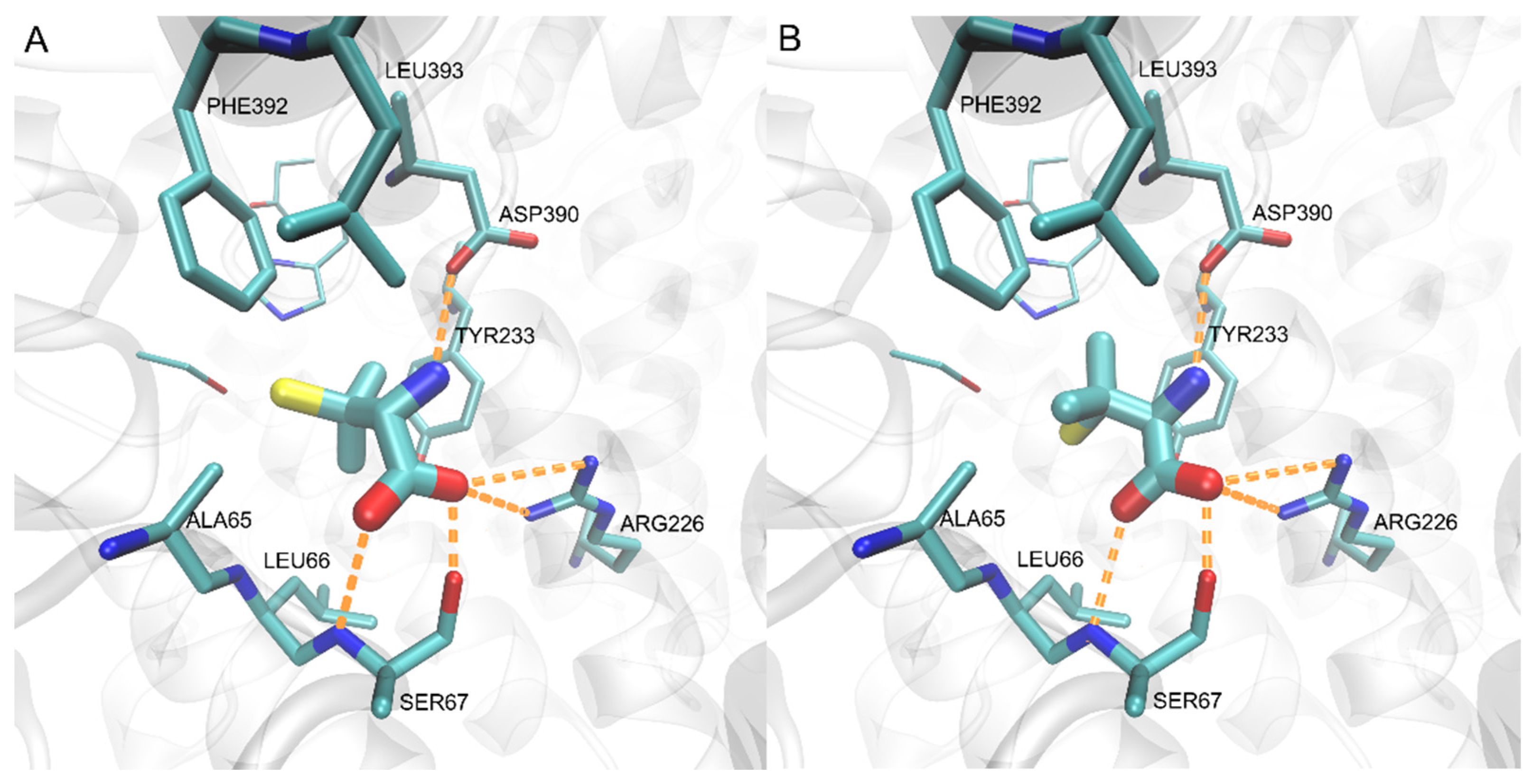
2.4. Penicillamine Inhibits CaMet2p
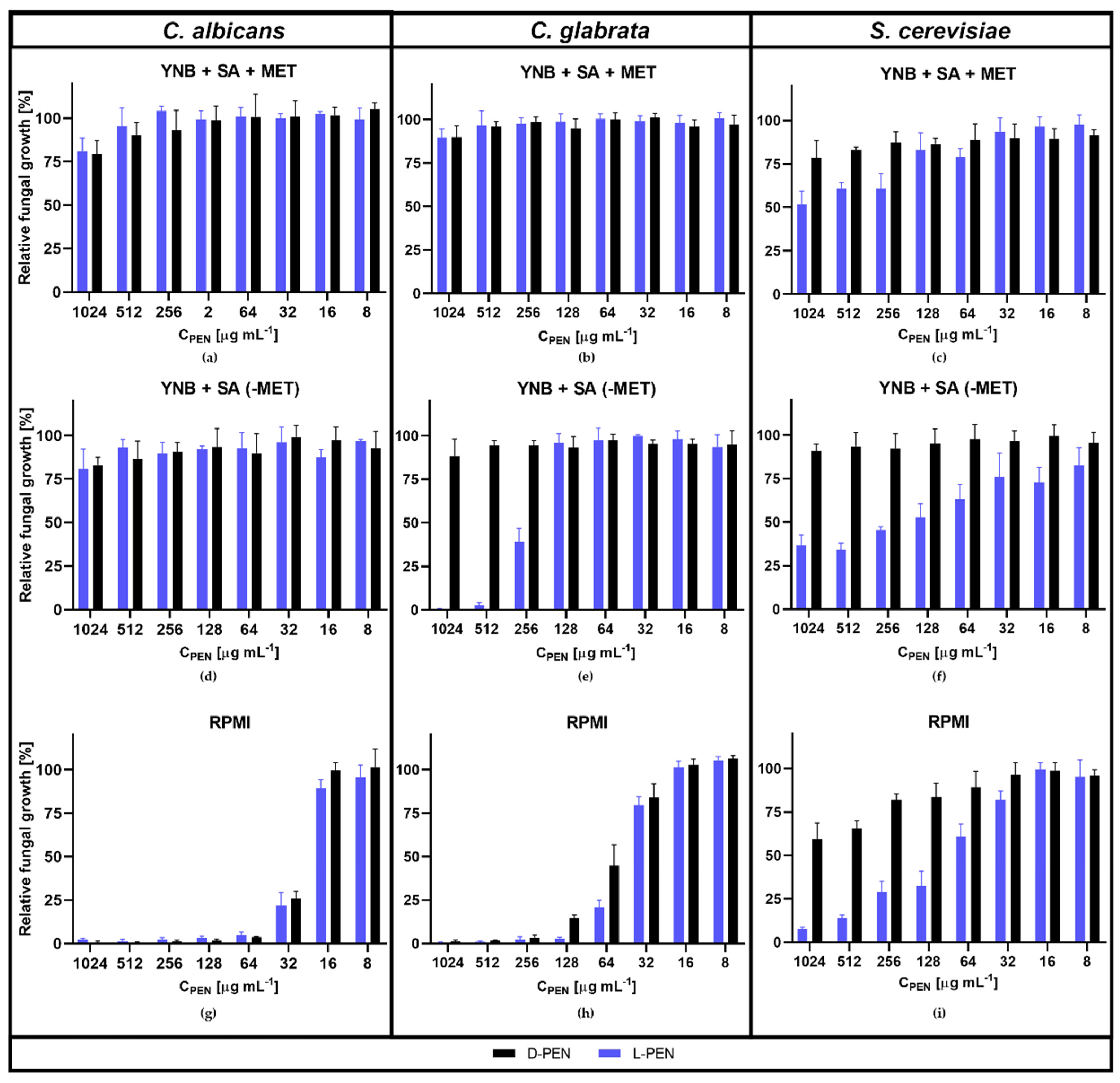
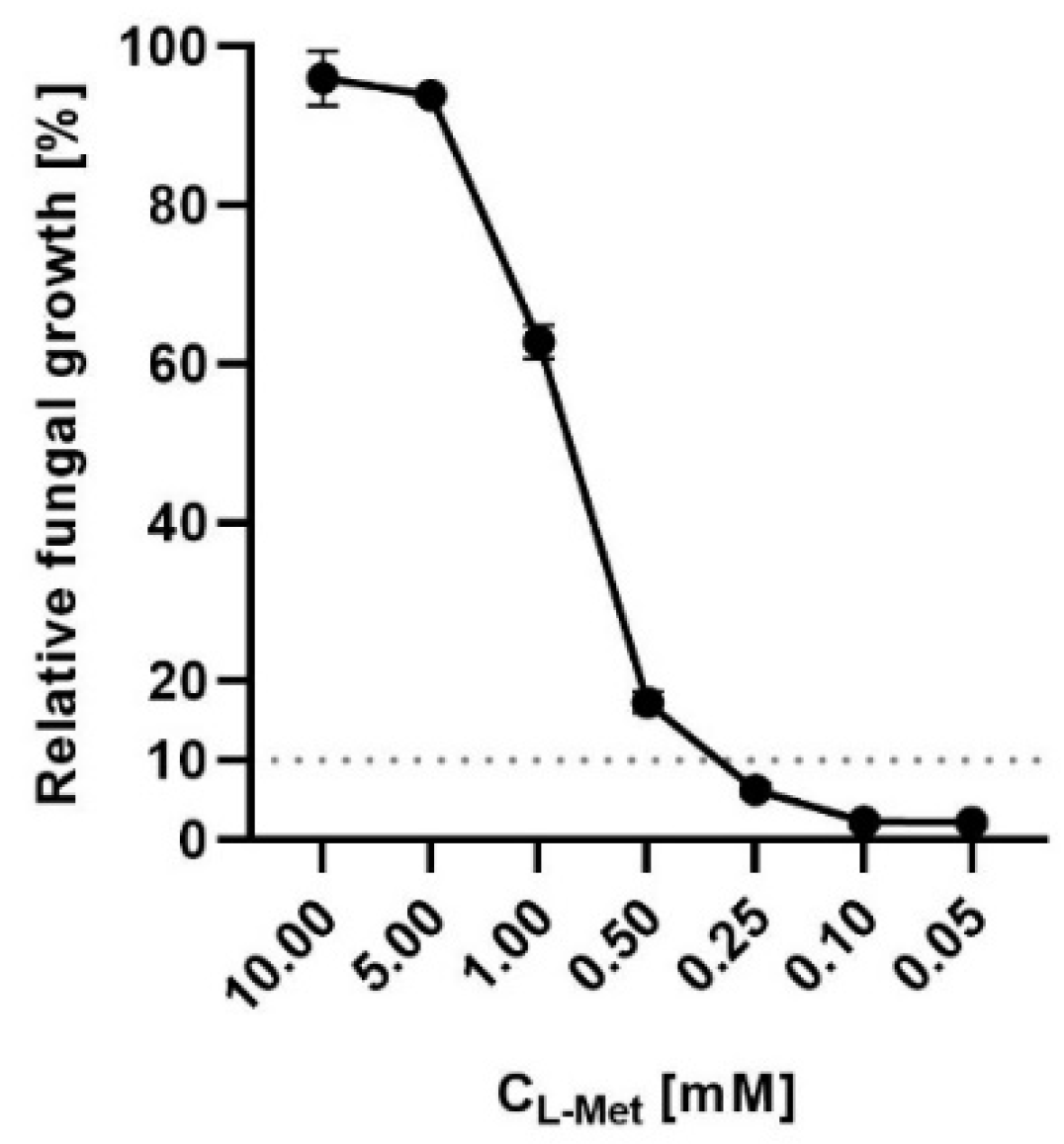
2.5. Penicillamine Exhibits Antifungal In Vitro Activity
3. Materials and Methods
3.1. Reagents
3.2. Microbial Strains and Growth Conditions
3.3. Cloning of MET2 Gene and Plasmids Construction
3.4. Mutagenesis
3.5. Western Blot Analysis
3.6. Expression of MET2 Gene
3.7. Enzyme Purification
3.8. Determination of Met2p Activity
3.9. Determination of the Kinetic Parameters
3.10. Determination of the Optimal pH and Reaction Buffer
3.11. Inhibitory Assay
3.12. Determination of Minimum Inhibitory Concentration
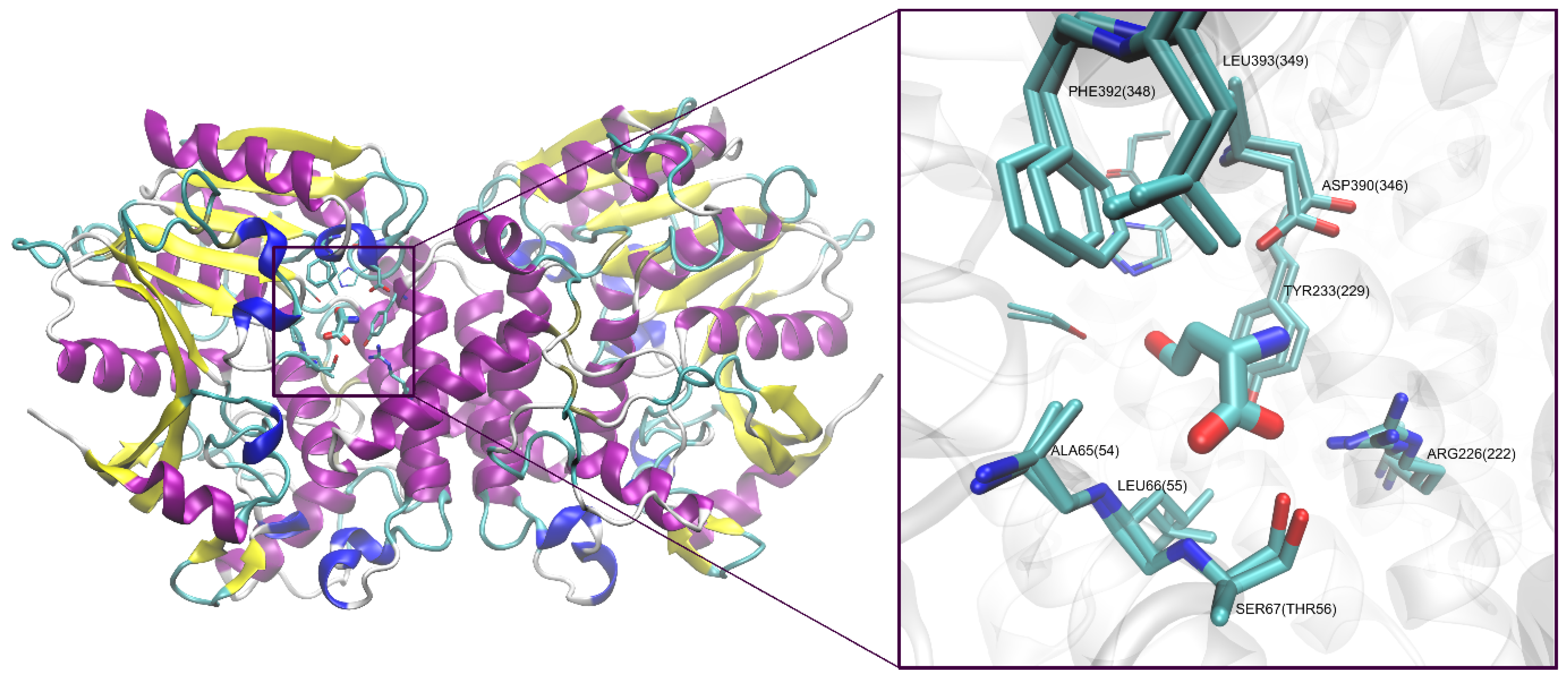
3.13. Homology Modeling and Docking Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brown, G.D.; Denning, D.W.; Gow, N.A.R.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden killers: Human fungal infections. Sci. Transl. Med. 2012, 4, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Patterson, T.F. Advances and challenges in management of invasive mycoses. Lancet 2005, 366, 1013–1025. [Google Scholar] [CrossRef]
- Almeida, F.; Rodrigues, M.L.; Coelho, C. The still underestimated problem of fungal diseases worldwide. Front. Microbiol. 2019, 10, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farmakiotis, D.; Kontoyiannis, D.P. Epidemiology of antifungal resistance in human pathogenic yeasts: Current viewpoint and practical recommendations for management. Int. J. Antimicrob. Agents 2017, 50, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Segal, E.S.; Gritsenko, V.; Levitan, A.; Yadav, B.; Dror, N.; Steenwyk, J.L.; Silberberg, Y.; Mielich, K.; Rokas, A.; Gow, N.A.R.; et al. Gene essentiality analyzed by in vivo transposon mutagenesis and machine learning in a stable haploid isolate of candida albicans. MBio 2018, 9, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuplińska, A.; Rząd, K. Molecular targets for antifungals in amino acid and protein biosynthetic pathways. Amino Acids 2021, 1, 3. [Google Scholar] [CrossRef]
- Rząd, K.; Milewski, S.; Gabriel, I. Versatility of putative aromatic aminotransferases from Candida albicans. Fungal Genet. Biol. 2018, 110, 26–37. [Google Scholar] [CrossRef]
- Stegink, L.D.; Filer, L.J.; Brummel, M.C.; Baker, G.L.; Krause, W.L.; Bell, E.F.; Ziegler, E.E. Plasma amino acid concentrations and amino acid ratios in normal adults and adults heterozygous for phenylketonuria ingesting a hamburger and milk shake meal. Am. J. Clin. Nutr. 1991, 53, 670–675. [Google Scholar] [CrossRef]
- Nazi, I.; Scott, A.; Sham, A.; Rossi, L.; Williamson, P.R.; Kronstad, J.W.; Wright, G.D. Role of homoserine transacetylase as a new target for antifungal agents. Antimicrob. Agents Chemother. 2007, 51, 1731–1736. [Google Scholar] [CrossRef] [Green Version]
- Kingsbury, J.M.; McCusker, J.H. Cytocidal amino acid starvation of Saccharomyces cerevisiae and Candida albicans acetolactate synthase (ilv2Δ) mutants is influenced by the carbon source and rapamycin. Microbiology 2010, 156, 929–939. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Sherman, F. Characteristics and relationships of mercury resistant mutants and methionine auxotrophs of yeast. J. Bacteriol. 1974, 118, 911–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obando Montoya, E.J.; Mélin, C.; Blanc, N.; Lanoue, A.; Foureau, E.; Boudesocque, L.; Prie, G.; Simkin, A.J.; Crèche, J.; Atehortùa, L.; et al. Disrupting the methionine biosynthetic pathway in Candida guilliermondii: Characterization of the MET2 gene as counter-selectable marker. Yeast 2014, 31, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Seyran, E. Fungal homoserine transacetylase: A potential antifungal target. İstanb. J. Pharm. 2021, 51, 137–140. [Google Scholar] [CrossRef]
- Berney, M.; Berney-Meyer, L.; Wong, K.W.; Chen, B.; Chen, M.; Kim, J.; Wang, J.; Harris, D.; Parkhill, J.; Chan, J.; et al. Essential roles of methionine and S-adenosylmethionine in the autarkic lifestyle of Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 2015, 112, 10008–10013. [Google Scholar] [CrossRef] [Green Version]
- Chaton, C.T.; Rodriguez, E.S.; Reed, R.W.; Li, J.; Kenner, C.W.; Korotkov, K.V. Structural analysis of mycobacterial homoserine transacetylases central to methionine biosynthesis reveals druggable active site. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Skrzypek, M.S.; Binkley, J.; Binkley, G.; Miyasato, S.R.; Simison, M.; Sherlock, G. The Candida Genome Database (CGD): Incorporation of Assembly 22, systematic identifiers and visualization of high throughput sequencing data. Nucleic Acids Res. 2017, 45, D592–D596. [Google Scholar] [CrossRef] [Green Version]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server. In The Proteomics Protocols Handbook; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar]
- Nazi, I.; Wright, G.D. Catalytic mechanism of fungal homoserine transacetylase. Biochemistry 2005, 44, 13560–13566. [Google Scholar] [CrossRef]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef] [Green Version]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Mirza, I.A.; Nazi, I.; Korczynska, M.; Wright, G.D.; Berghuis, A.M. Crystal structure of homoserine transacetylase from Haemophilus influenzae reveals a new family of α/β-hydrolases. Biochemistry 2005, 44, 15768–15773. [Google Scholar] [CrossRef]
- Bailey, T.L.; Gribskov, M. Combining evidence using p-values: Application to sequence homology searches. Bioinformatics 1998, 14, 48–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, S.; Stamatoyannopoulos, J.A.; Bailey, T.L.; Noble, W. Quantifying similarity between motifs. Genome Biol. 2007, 8, R24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, C.E.; Bailey, T.L.; Noble, W.S. FIMO: Scanning for occurrences of a given motif. Bioinformatics 2011, 27, 1017–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyman, A.; Paulus, H. Purification and properties of homoserine transacetylase from Bacillus polymyxa. J. Biol. Chem. 1975, 250, 3897–3903. [Google Scholar] [CrossRef]
- Shiio, I.; Ozaki, H. Feedback inhibition by methionine and S-adenosylmethionine, and desensitization of homoserine O-acetyltransferase in Brevibacterium flavum. J. Biochem. 1981, 89, 1493–1500. [Google Scholar] [CrossRef]
- Yamagata, S. Partial purification and some properties of homoserine O-acetyltransferase of a methionine auxotroph of Saccharomyces cerevisiae. J. Bacteriol. 1987, 169, 3458–3463. [Google Scholar] [CrossRef] [Green Version]
- Sagong, H.Y.; Hong, J.; Kim, K.J. Crystal structure and biochemical characterization of O-acetylhomoserine acetyltransferase from Mycobacterium smegmatis ATCC 19420. Biochem. Biophys. Res. Commun. 2019, 517, 399–406. [Google Scholar] [CrossRef]
- Lowther, J.; Beattie, A.E.; Langridge-Smith, P.R.R.; Clarke, D.J.; Campopiano, D.J. L-Penicillamine is a mechanism-based inhibitor of serine palmitoyltransferase by forming a pyridoxal-5′-phosphate-thiazolidine adduct. Medchemcomm 2012, 3, 1003–1008. [Google Scholar] [CrossRef] [Green Version]
- Weiss, K.H.; Thurik, F.; Gotthardt, D.N.; Schäfer, M.; Teufel, U.; Wiegand, F.; Merle, U.; Ferenci-Foerster, D.; Maieron, A.; Stauber, R.; et al. Efficacy and safety of oral chelators in treatment of patients with wilson disease. Clin. Gastroenterol. Hepatol. 2013, 11, 1028–1035.e2. [Google Scholar] [CrossRef]
- Eidelman, C.; Lowry, J.A. D-Penicillamine. In Critical Care Toxicology; Springer International Publishing: Cham, Switzerland, 2016; pp. 1–7. ISBN 9783319207902. [Google Scholar]
- Borzyszkowska-Bukowska, J.; Górska, J.; Szczeblewski, P.; Laskowski, T.; Gabriel, I.; Jurasz, J.; Kozłowska-Tylingo, K.; Szweda, P.; Milewski, S. Quest for the molecular basis of improved selective toxicity of all-trans isomers of aromatic heptaene macrolide antifungal antibiotics. Int. J. Mol. Sci. 2021, 22, 10108. [Google Scholar] [CrossRef]
- Li, Y.Y.; Jiao, P.; Li, Y.Y.; Gong, Y.; Chen, X.; Sun, S. The synergistic antifungal effect and potential mechanism of D-Penicillamine combined with fluconazole against Candida albicans. Front. Microbiol. 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marzluf, G.A. Molecular genetics of sulfur assimilation in filamentous fungi and yeast. Annu. Rev. Microbiol. 1997, 51, 73–96. [Google Scholar] [CrossRef] [PubMed]
- Hébert, A.; Casaregola, S.; Beckerich, J.M. Biodiversity in sulfur metabolism in hemiascomycetous yeasts. FEMS Yeast Res. 2011, 11, 366–378. [Google Scholar] [CrossRef] [Green Version]
- Cherest, H.; Surdin-Kerjan, Y. Genetic analysis of a new mutation conferring cysteine auxotrophy in Saccharomyces cerevisiae: Updating of the sulfur metabolism pathway. Genetics 1992, 130, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.K.; Kumar Bachhawat, A.; Bachhawat, A.K. CgCYN1, a plasma membrane cystine-specific transporter of Candida glabrata with orthologues prevalent among pathogenic yeast and fungi. J. Biol. Chem. 2011, 286, 19714–19723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Murillo, L.A.; Newport, G.; Lan, C.Y.; Habelitz, S.; Dungan, J.; Agabian, N.M. Genome-wide transcription profiling of the early phase of biofilm formation by Candida albicans. Eukaryot. Cell 2005, 4, 1562–1573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jauniaux, J.-C.; Grenson, M. GAP1, the general amino acid permease gene of Saccharomyces cerevisiae Nucleotide sequence, protein similarity with the other bakers yeast amino acid permeases, and nitrogen catabolite repression. Eur. J. Biochem. 1990, 190, 39–44. [Google Scholar] [CrossRef]
- Regenberg, B.; Düring-Olsen, L.; Kielland-Brandt, M.C.; Holmberg, S. Substrate specificity and gene expression of the amino acid permeases in Saccharomyces cerevisiae. Curr. Genet. 1999, 36, 317–328. [Google Scholar] [CrossRef]
- Martho, K.F.C.; De Melo, A.T.; Takahashi, J.P.F.; Guerra, J.M.; Da Silva Santos, D.C.; Purisco, S.U.; Melhem, M.D.S.C.; Dos Anjos Fazioli, R.; Phanord, C.; Sartorelli, P.; et al. Amino acid permeases and virulence in Cryptococcus neoformans. PLoS ONE 2016, 11, e0163919. [Google Scholar] [CrossRef]
- Corso, G.; Cristofano, A.; Sapere, N.; La Marca, G.; Angiolillo, A.; Vitale, M.; Fratangelo, R.; Lombardi, T.; Porcile, C.; Intrieri, M.; et al. Serum amino acid profiles in normal subjects and in patients with or at risk of Alzheimer dementia. Dement. Geriatr. Cogn. Dis. Extra 2017, 7, 143–159. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.A.; Rinaldi, S.; Scalbert, A.; Ferrari, P.; Achaintre, D.; Gunter, M.J.; Appleby, P.N.; Key, T.J.; Travis, R.C. Plasma concentrations and intakes of amino acids in male meat-eaters, fish-eaters, vegetarians and vegans: A cross-sectional analysis in the EPIC-Oxford cohort. Eur. J. Clin. Nutr. 2016, 70, 306–312. [Google Scholar] [CrossRef] [Green Version]
- Sato, K.; Fukushima, D.; Doi, H.; Satomi, S. Higher serum methionine levels as a predictive factor in patients with irreversible fulminant hepatic failure. Transplant. Proc. 2013, 45, 1904–1906. [Google Scholar] [CrossRef] [PubMed]
- Szweda, P.; Gucwa, K.; Romanowska, E.; Dzierżanowska-Fangrat, K.; Naumiuk, Ł.; Brillowska-Dąbrowska, A.; Wojciechowska-Koszko, I.; Milewski, S. Mechanisms of azole resistance among clinical isolates of Candida glabrata in Poland. J. Med. Microbiol. 2015, 64, 610–619. [Google Scholar] [CrossRef] [PubMed]
- Lazar, I., Jr.; Lazar, I., Sr. GelAnalyzer 19.1. Available online: www.gelanalyzer.com (accessed on 10 January 2022).
- Foyn, H.; Thompson, P.R.; Arnesen, T. DTNB-based quantification of in vitro enzymatic N-terminal acetyltransferase activity. In Methods in Molecular Biology; Schilling, O., Ed.; Humana Press Inc.: New York, NY, USA, 2017; Volume 1574, pp. 9–15. ISBN 978-1-4939-6849-7. [Google Scholar]
- Wayne, P. Reference method for broth dilution antifungal susceptibility testing of yeasts; Approved standar—Third Edition. CLSI Doc. 2008, 28, M27-A3. [Google Scholar]
- Froimowitz, M. HyperChem: A software package for computational chemistry and molecular modeling. Biotechniques 1993, 14, 1010–1013. [Google Scholar]
- Morris, G.M.; Ruth, H.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. Software news and updates AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wojciechowski, M. Simplified AutoDock force field for hydrated binding sites. J. Mol. Graph. Model. 2017, 78, 74–80. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | KM [mM] | kcat [s−1] | kcat/KM [M−1·s−1] | Vmax [µmol·min−1·mg−1] |
|---|---|---|---|---|
| CaMet2p | ||||
| L-Hom | 0.405 ± 0.050 | 13.0 ± 0.405 | 32.1 × 103 | 17.1 ± 0.536 |
| AcCoA | 0.906 ± 0.119 | 15.2 ± 0.572 | 16.8 × 103 | 20.1 ± 0.756 |
| CaMet2NHp | ||||
| L-Hom | 0.578 ± 0.118 | 13.1 ± 0.723 | 22.7 × 103 | 17.2 ± 0.956 |
| AcCoA | 1.13 ± 0.150 | 15.0 ± 0.635 | 13.3 × 103 | 19.8 ± 0.839 |
| MIC50 (MIC90) [µg·mL−1] | |||||
|---|---|---|---|---|---|
| L-Pen | Flu | AmB | |||
| Strain | YNB + SA + L-Met | YNB + SA − L-Met | RPMI1640 | RPMI1640 | RPMI1640 |
| Candida albicans ATCC 10231 | >1024 | >1024 | 32 (64) | (4) | (0.5) |
| Candida parapsilosis ATCC 22019 | >1024 | >1024 | 256 (1024) | (8) | (1) |
| Candida krusei ATCC 6258 | >1024 | >1024 | >1024 | (64) | (1) |
| Candida glabrata ATCC 90030 | >1024 | 256 (512) | 64 (128) | (32) | (1) |
| Candida famata DSM 3428 | >1024 | >1024 | 1024 (>1024) | (16) | (1.0) * |
| Candida rugosa DSM 2031 | >1024 | >1024 | 1024 (>1024) | (16) | (2.0) * |
| Candida dublinensis CBS 7987 | >1024 | >1024 | >1024 | (4) | (0.25) * |
| Saccharomyces cerevisiae ATCC 9763 | 512 (>1024) | 128 (>1024) | 128 (1024) | (8) | (0.5) |
| D-Pen | Flu | AmB | |||
| YNB + SA + L-Met | YNB + SA − L-Met | RPMI1640 | RPMI1640 | RPMI1640 | |
| Candida albicans ATCC 10231 | >1024 | >1024 | 32 (64) | (4) | (0.5) |
| Candida glabrata ATCC 90030 | >1024 | >1024 | 128 (256) | (32) | (1) |
| Saccharomyces cerevisiae ATCC 9763 | >1024 | >1024 | >1024 | (8) | (0.5) |
| Susceptibility | MIC90 (MIC50) [µg·mL−1] | ||
|---|---|---|---|
| Strain | Flu | Flu | L-Pen |
| C. glabrata CZD 6 | S | 32 | 512 (256) |
| C. glabrata CZD 209 | R | >256 | 512 |
| C. glabrata GD 211 | R | 256 | 512 (256) |
| C. glabrata GD 310 | R | >256 | 512 |
| C. glabrata CZD 342 | S | 16 | 512 |
| C. glabrata CZD 373 | R | 256 | 512 (256) |
| C. glabrata CZD 377 | R | >256 | 512 |
| C. glabrata CZD 513 | R | >256 | 512 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuplińska, A.; Rząd, K.; Wojciechowski, M.; Milewski, S.; Gabriel, I. Antifungal Effect of Penicillamine Due to the Selective Targeting of L-Homoserine O-Acetyltransferase. Int. J. Mol. Sci. 2022, 23, 7763. https://doi.org/10.3390/ijms23147763
Kuplińska A, Rząd K, Wojciechowski M, Milewski S, Gabriel I. Antifungal Effect of Penicillamine Due to the Selective Targeting of L-Homoserine O-Acetyltransferase. International Journal of Molecular Sciences. 2022; 23(14):7763. https://doi.org/10.3390/ijms23147763
Chicago/Turabian StyleKuplińska, Aleksandra, Kamila Rząd, Marek Wojciechowski, Sławomir Milewski, and Iwona Gabriel. 2022. "Antifungal Effect of Penicillamine Due to the Selective Targeting of L-Homoserine O-Acetyltransferase" International Journal of Molecular Sciences 23, no. 14: 7763. https://doi.org/10.3390/ijms23147763
APA StyleKuplińska, A., Rząd, K., Wojciechowski, M., Milewski, S., & Gabriel, I. (2022). Antifungal Effect of Penicillamine Due to the Selective Targeting of L-Homoserine O-Acetyltransferase. International Journal of Molecular Sciences, 23(14), 7763. https://doi.org/10.3390/ijms23147763