
T-Cell Intracellular Antigen 1-Like Protein in Physiology and Pathology


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
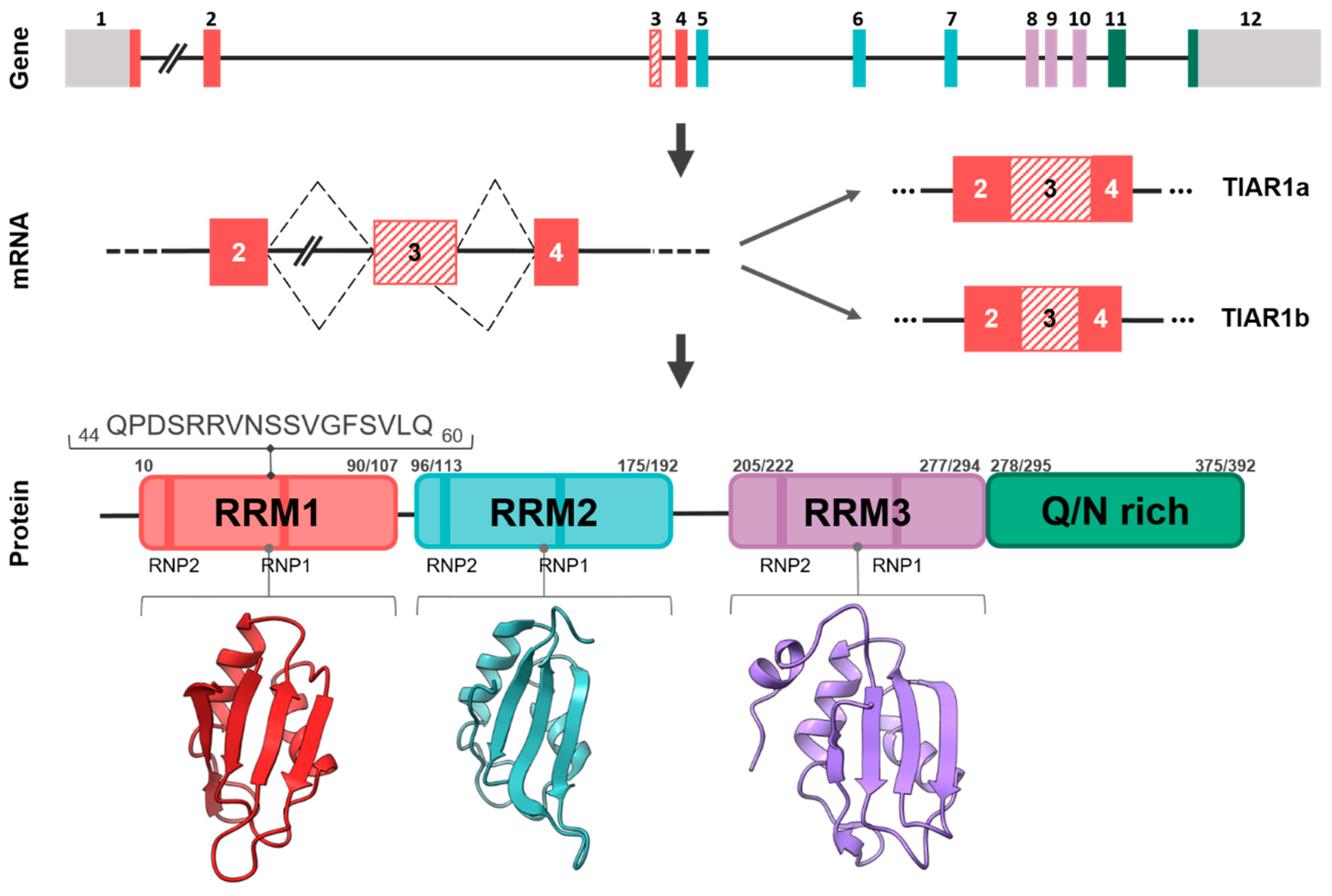
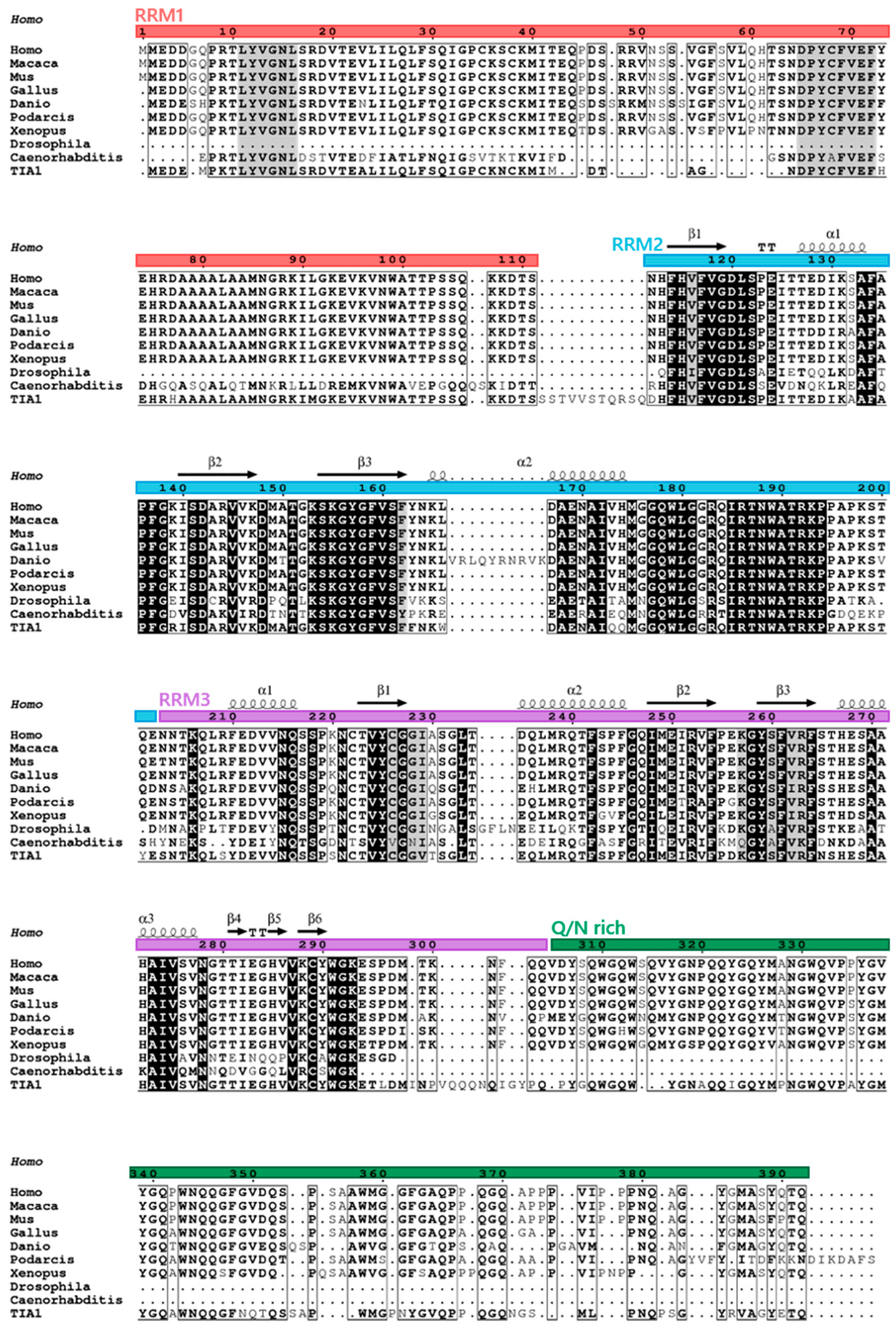
TIAR: One Gene, Two Main Isoforms and a Classical RBP Structure
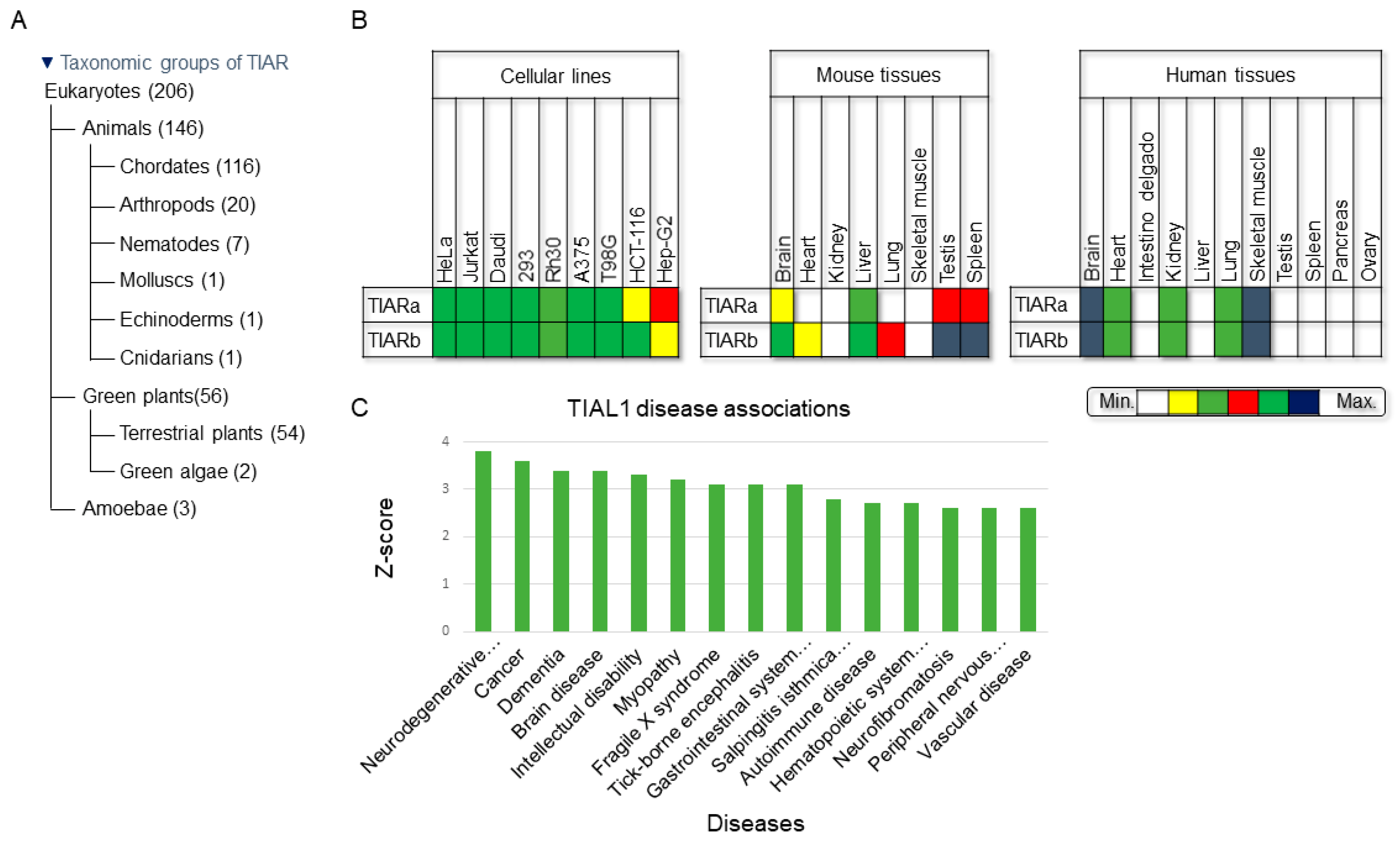
2. Phylogenetics and Cellular/Tissular Expression Profiling
2.1. Gene Expression Control
2.2. Transcription
2.3. Alternative Splicing
2.4. Translation
2.5. Turnover/Stability

2.6. lncRNA- and miRNA-Mediated Regulation
2.7. Tissular and Cellular Homeostasis
2.8. Autophagy
2.9. Apoptosis
2.10. Cell Cycle
2.11. Mitochondrial Function
2.12. Cellular Stress
2.13. Viral Biology
3. Physiology and Pathology
3.1. Inflammation
3.2. Embryogenesis
3.3. Carcinogenesis
3.4. Neurodegenerative Diseases
3.4.1. Neurofibromatosis Type I
3.4.2. Axon Regeneration
3.4.3. Alzheimer’s Disease
4. Future Challenges
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Kawakami, A.; Tian, Q.; Duan, X.; Streuli, M.; Schlossman, S.F.; Anderson, P. Identification and functional characterization of a TIA-1-related nucleolysin. Proc. Natl. Acad. Sci. USA 1992, 89, 8681–8685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, Q.; Streuli, M.; Saito, H.; Schlossman, S.F.; Anderson, P. A polyadenylate binding protein localized to the granules of cytolytic lymphocytes induces DNA fragmentation in target cells. Cell 1991, 67, 629–639. [Google Scholar] [CrossRef]
- Beck, A.R.; Medley, Q.G.; O’Brien, S.; Anderson, P.; Streuli, M. Structure, tissue distribution and genomic organization of the murine RRM-type RNA binding proteins TIA-1 and TIAR. Nucleic Acids Res. 1996, 24, 3829–3835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dember, L.M.; Kim, N.D.; Liu, K.Q.; Anderson, P. Individual RNA recognition motifs of TIA-1 and TIAR have different RNA binding specificities. J. Biol. Chem. 1996, 271, 2783–2788. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.S.; Headey, S.J.; Yoga, Y.M.; Scanlon, M.J.; Gorospe, M.; Wilce, M.C.; Wilce, J.A. Distinct binding properties of TIAR RRMs and linker region. RNA Biol. 2013, 10, 579–589. [Google Scholar] [CrossRef] [Green Version]
- Waris, S.; Wilce, M.C.; Wilce, J.A. RNA recognition and stress granule formation by TIA proteins. Int. J. Mol. Sci. 2014, 15, 23377–23388. [Google Scholar] [CrossRef] [Green Version]
- Taupin, J.L.; Tian, Q.; Kedersha, N.; Robertson, M.; Anderson, P. The RNA-binding protein TIAR is translocated from the nucleus to the cytoplasm during Fas-mediated apoptotic cell death. Proc. Natl. Acad. Sci. USA 1995, 92, 1629–1633. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Gómez, A.; Izquierdo, J.M. The multifunctional faces of T-cell Intracellular antigen 1 in health and disease. Int. J. Mol. Sci. 2022, 23, 1400. [Google Scholar] [CrossRef]
- Adam, S.A.; Nakagawa, T.; Swanson, M.S.; Woodruff, T.K.; Dreyfuss, G. mRNA polyadenylate-binding protein: Gene isolation and sequencing and identification of a ribonucleoprotein consensus sequence. Mol. Cell. Biol. 1986, 6, 2932–2943. [Google Scholar]
- Swanson, M.S.; Nakagawa, T.Y.; LeVan, K.; Dreyfuss, G. Primary structure of human nuclear ribonucleoprotein particle C proteins: Conservation of sequence and domain structures in heterogeneous nuclear RNA, mRNA, and pre-rRNA-binding proteins. Mol. Cell. Biol. 1987, 7, 173–1739. [Google Scholar]
- Dreyfuss, G.; Swanson, M.S.; Pinol-Roma, S. Heterogeneous nuclear ribonucleoprotein particles and the pathway of mRNA formation. Trends Biochem. Sci. 1988, 13, 86–91. [Google Scholar] [CrossRef]
- Mazan-Mamczarz, K.; Lal, A.; Martindale, J.L.; Kawai, T.; Gorospe, M. Translational repression by RNA-binding protein TIAR. Mol. Cell. Biol. 2006, 26, 2716–2727. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.S.; Kuwano, Y.; Zhan, M.; Pullmann, R., Jr.; Mazan-Mamczarz, K.; Li, H.; Kedersha, N.; Anderson, P.; Wilce, M.C.; Gorospe, M.; et al. Elucidation of a C-rich signature motif in target mRNAs of RNA-binding protein TIAR. Mol. Cell. Biol. 2007, 27, 6806–6817. [Google Scholar] [CrossRef] [Green Version]
- Izquierdo, J.M.; Valcárcel, J. Two isoforms of the T-cell intracellular antigen 1 (TIA-1) splicing factor display distinct splicing regulation activities. Control of TIA-1 isoform ratio by TIA-1-related protein. J. Biol. Chem. 2007, 282, 19410–19417. [Google Scholar] [CrossRef] [Green Version]
- The Human Protein Atlas. Available online: https://www.proteinatlas.org/ENSG00000151923-TIAL1 (accessed on 20 May 2022).
- Eraslan, G.; Drokhlyansky, E.; Anand, S.; Fiskin, E.; Subramanian, A.; Slyper, M.; Wang, J.; Van Wittenberghe, N.; Rouhana, J.M.; Julia Waldman, J.; et al. Single-nucleus cross-tissue molecular reference maps toward understanding disease gene function. Science 2022, 376, eabl4290. [Google Scholar] [CrossRef]
- Tabula Sapiens Consortium; Jones, R.C.; Karkanias, J.; Krasnow, M.A.; Oliveira Pisco, A.; Quake, S.R.; Salzman, J.; Yosef, N.; Bulthaup, B.; Brown, P.; et al. The Tabula Sapiens: A multiple-organ, single-cell transcriptomic atlas of humans. Science 2022, 376, eabl4896. [Google Scholar]
- Domínguez Conde, C.; Xu, C.; Jarvis, L.B.; Rainbow, D.B.; Wells, S.B.; Gomes, T.; Howlett, S.K.; Suchanek, O.; Polanski, K.; King, H.W.; et al. Cross-tissue immune cell analysis reveals tissue-specific features in humans. Science 2022, 376, eabl5197. [Google Scholar] [CrossRef]
- Pullmann, R., Jr.; Kim, H.H.; Abdelmohsen, K.; Lal, A.; Martindale, J.L.; Yang, X.; Gorospe, M. Analysis of turnover and translation regulatory RNA-binding protein expression through binding to cognate mRNAs. Mol. Cell. Biol. 2007, 27, 6265–6278. [Google Scholar] [CrossRef] [Green Version]
- Masuda, K.; Marasa, B.; Martindale, J.L.; Halushka, M.K.; Gorospe, M. Tissue- and age-dependent expression of RNA-binding proteins that influence mRNA turnover and translation. Aging 2009, 1, 681–698. [Google Scholar] [CrossRef] [Green Version]
- Cho, N.H.; Cheveralls, K.C.; Brunner, A.D.; Kim, K.; Michaelis, A.C.; Raghavan, P.; Kobayashi, H.; Savy, L.; Li, J.Y.; Canaj, H.; et al. OpenCell: Endogenous tagging for the cartography of human cellular organization. Science 2022, 375, eabi6983. [Google Scholar] [CrossRef]
- Das, R.; Yu, J.; Zhang, Z.; Gygi, M.P.; Krainer, A.R.; Gygi, S.P.; Reed, R. SR proteins function in coupling RNAP II transcription to pre-mRNA splicing. Mol. Cell 2007, 26, 867–881. [Google Scholar] [CrossRef]
- Suswam, E.A.; Li, Y.Y.; Mahtani, H.; King, P.H. Novel DNA-binding properties of the RNA-binding protein TIAR. Nucleic Acids Res. 2005, 33, 4507–4518. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.S.; Wilce, M.C.; Yoga, Y.M.; Pendini, N.R.; Gunzburg, M.J.; Cowieson, N.P.; Wilson, G.M.; Williams, B.R.; Gorospe, M.; Wilce, J.A. Different modes of interaction by TIAR and HuR with target RNA and DNA. Nucleic Acids Res. 2011, 39, 1117–1130. [Google Scholar] [CrossRef] [Green Version]
- Tominaga, A.; Sugawara, H.; Futagawa, T.; Inoue, K.; Sasaki, K.; Minamino, N.; Hatakeyama, M.; Handa, H.; Miyata, A. Characterization of the testis-specific promoter region in the human pituitary adenylate cyclase-activating polypeptide (PACAP) gene. Genes Cells 2010, 15, 595–606. [Google Scholar] [CrossRef]
- Reyes, R.; Alcalde, J.; Izquierdo, J.M. Depletion of T-cell intracellular antigen proteins promotes cell proliferation. Genome Biol. 2009, 10, R87. [Google Scholar] [CrossRef] [Green Version]
- Del Gatto-Konczak, F.; Bourgeois, C.F.; Le Guiner, C.; Kister, L.; Gesnel, M.C.; Stévenin, J.; Breathnach, R. The RNA-binding protein TIA-1 is a novel mammalian splicing regulator acting through intron sequences adjacent to a 5′ splice site. Mol. Cell. Biol. 2000, 20, 6287–6299. [Google Scholar] [CrossRef]
- Förch, P.; Puig, O.; Kedersha, N.; Martínez, C.; Granneman, S.; Séraphin, B.; Anderson, P.; Valcárcel, J. The apoptosis-promoting factor TIA-1 is a regulator of alternative pre-mRNA splicing. Mol. Cell 2000, 6, 1089–1098. [Google Scholar] [CrossRef]
- Le Guiner, C.; Lejeune, F.; Galiana, D.; Kister, L.; Breathnach, R.; Stévenin, J.; Del Gatto-Konczak, F. TIA-1 and TIAR activate splicing of alternative exons with weak 5’ splice sites followed by a U-rich stretch on their own pre-mRNAs. J. Biol. Chem. 2001, 276, 40638–40646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Förch, P.; Puig, O.; Martínez, C.; Séraphin, B.; Valcárcel, J. The splicing regulator TIA-1 interacts with U1-C to promote U1 snRNP recruitment to 5′ splice sites. EMBO J. 2002, 21, 6882–6892. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, J.M.; Majós, N.; Bonnal, S.; Martínez, C.; Castelo, R.; Guigó, R.; Bilbao, D.; Valcárcel, J. Regulation of Fas alternative splicing by antagonistic effects of TIA-1 and PTB on exon definition. Mol. Cell 2005, 19, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Hinman, M.N.; Hasman, R.A.; Mehta, P.; Lou, H. Regulation of neuron-specific alternative splicing of neurofibromatosis type 1 pre-mRNA. Mol. Cell. Biol. 2008, 28, 1240–1251. [Google Scholar] [CrossRef] [Green Version]
- Aznarez, I.; Barash, Y.; Shai, O.; He, D.; Zielenski, J.; Tsui, L.C.; Parkinson, J.; Frey, B.J.; Rommens, J.M.; Blencowe, B.J. A systematic analysis of intronic sequences downstream of 5′ splice sites reveals a widespread role for U-rich motifs and TIA1/TIAL1 proteins in alternative splicing regulation. Genome Res. 2008, 18, 1247–1258. [Google Scholar] [CrossRef] [Green Version]
- Gal-Mark, N.; Schwartz, S.; Ram, O.; Eyras, E.; Ast, G. The pivotal roles of TIA proteins in 5′ splice-site selection of alu exons and across evolution. PLoS Genet. 2009, 5, e1000717. [Google Scholar] [CrossRef]
- Wang, Z.; Kayikci, M.; Briese, M.; Zarnack, K.; Luscombe, N.M.; Rot, G.; Zupan, B.; Curk, T.; Ule, J. iCLIP predicts the dual splicing effects of TIA-RNA interactions. PLoS Biol. 2010, 8, 1000530. [Google Scholar] [CrossRef] [Green Version]
- Izquierdo, J.M. Heterogeneous ribonucleoprotein C displays a repressor activity mediated by T-cell intracellular antigen-1-related/like protein to modulate Fas exon 6 splicing through a mechanism involving Hu antigen R. Nucleic Acids Res. 2010, 38, 8001–8014. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.N.; Seo, J.; Ottesen, E.W.; Shishimorova, M.; Bhattacharya, D.; Singh, R.N. TIA1 prevents skipping of a critical exon associated with spinal muscular atrophy. Mol. Cell. Biol. 2011, 31, 935–954. [Google Scholar] [CrossRef] [Green Version]
- Meyer, C.; Garzia, A.; Mazzola, M.; Gerstberger, S.; Molina, H.; Tuschl, T. The TIA1 RNA-binding protein family regulates EIF2AK2-mediated stress response and cell cycle progression. Mol. Cell 2018, 69, 622–635. [Google Scholar] [CrossRef] [Green Version]
- Van Nostrand, E.L.; Freese, P.; Pratt, G.A.; Wang, X.; Wei, X.; Xiao, R.; Blue, S.M.; Chen, J.Y.; Cody, N.A.L.; Dominguez, D.; et al. A large-scale binding and functional map of human RNA-binding proteins. Nature 2020, 583, 711–719. [Google Scholar] [CrossRef]
- Zhao, W.; Zhao, J.; Hou, M.; Wang, Y.; Zhang, Y.; Zhao, X.; Zhang, C.; Guo, D. HuR and TIA1/TIAL1 are involved in regulation of alternative splicing of SIRT1 pre-mRNA. Int. J. Mol. Sci. 2014, 15, 2946–2958. [Google Scholar] [CrossRef] [Green Version]
- Královičová, J.; Borovská, I.; Pengelly, R.; Lee, E.; Abaffy, P.; Šindelka, R.; Grutzner, F.; Vořechovský, I. Restriction of an intron size en route to endothermy. Nucleic Acids Res. 2021, 49, 2460–2487. [Google Scholar] [CrossRef]
- Zhu, H.; Hasman, R.A.; Young, K.M.; Kedersha, N.L.; Hua, L. U1 snRNP-dependent function of TIAR in the regulation of alternative RNA processing of the human calcitonin/CGRP pre-mRNA. Mol. Cell. Biol. 2003, 23, 5959–5971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shukla, S.; Dirksen, W.P.; Joyce, K.M.; Le Guiner-Blanvillain, C.; Breathnach, R.; Fisher, S.A. TIA proteins are necessary but not sufficient for the tissue-specific splicing of the myosin phosphatase targeting subunit 1. J. Biol. Chem. 2004, 279, 13668–13676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; Delestienne, N.; Huez, G.; Kruys, V.; Gueydan, C. Identification of the sequence determinants mediating the nucleo-cytoplasmic shuttling of TIAR and TIA-1 RNA-binding proteins. J. Cell. Sci. 2005, 118, 5453–5463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gueydan, C.; Droogmans, L.; Chalon, P.; Huez, G.; Caput, D.; Kruys, V. Identification of TIAR as a protein binding to the translational regulatory AU-rich element of tumor necrosis factor alpha mRNA. J. Biol. Chem. 1999, 274, 2322–2326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piecyk, M.; Wax, S.; Beck, A.R.; Kedersha, N.; Gupta, M.; Maritim, B.; Chen, S.; Gueydan, C.; Kruys, V.; Streuli, M.; et al. TIA-1 is a translational silencer that selectively regulates the expression of TNF-alpha. EMBO J. 2000, 19, 4154–4163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Q.; Cok, S.J.; Zeng, C.; Morrison, A.R. Translational repression of human matrix metalloproteinases-13 by an alternatively spliced form of T-cell-restricted intracellular antigen-related protein (TIAR). J. Biol. Chem. 2003, 278, 1579–1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cok, S.J.; Acton, S.J.; Morrison, A.R. The proximal region of the 3’-untranslated region of cyclooxygenase-2 is recognized by a multimeric protein complex containing HuR, TIA-1, TIAR, and the heterogeneous nuclear ribonucleoprotein U. J. Biol. Chem. 2003, 278, 36157–36162. [Google Scholar] [CrossRef] [Green Version]
- Cok, S.J.; Acton, S.J.; Sexton, A.E.; Morrison, A.R. Identification of RNA-binding proteins in RAW 264.7 cells that recognize a lipopolysaccharide-responsive element in the 3-untranslated region of the murine cyclooxygenase-2 mRNA. J. Biol. Chem. 2004, 279, 8196–8205. [Google Scholar] [CrossRef] [Green Version]
- Kandasamy, K.; Joseph, K.; Subramaniam, K.; Raymond, J.R.; Tholanikunnel, B.G. Translational control of beta2-adrenergic receptor mRNA by T-cell-restricted intracellular antigen-related protein. J. Biol. Chem. 2005, 280, 1931–1943. [Google Scholar] [CrossRef] [Green Version]
- László, C.F.; Fayad, S.; Carpenter, O.; George, K.S.; Lu, W.; Saad, A.A.; Wu, S. The role of translational regulation in ultraviolet C light-induced cyclooxygenase-2 expression. Life Sci. 2009, 85, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Subramaniam, K.; Kandasamy, K.; Joseph, K.; Spicer, E.K.; Tholanikunnel, B.G. The 3′-untranslated region length and AU-rich RNA location modulate RNA-protein interaction and translational control of β2-adrenergic receptor mRNA. Mol. Cell. Biochem. 2011, 352, 125–141. [Google Scholar] [CrossRef]
- Colegrove-Otero, L.J.; Devaux, A.; Standart, N. The Xenopus ELAV protein ElrB represses Vg1 mRNA translation during oogenesis. Mol. Cell. Biol. 2005, 25, 9028–9039. [Google Scholar] [CrossRef] [Green Version]
- Lal, A.; Abdelmohsen, K.; Pullmann, R.; Kawai, T.; Galban, S.; Yang, X.; Brewer, G.; Gorospe, M. Posttranscriptional derepression of GADD45alpha by genotoxic stress. Mol. Cell. 2006, 22, 117–128. [Google Scholar] [CrossRef]
- Yu, C.; York, B.; Wang, S.; Feng, Q.; Xu, J.; O’Malley, B.W. An essential function of the SRC-3 coactivator in suppression of cytokine mRNA translation and inflammatory response. Mol. Cell 2007, 25, 765–778. [Google Scholar] [CrossRef] [Green Version]
- Liao, B.; Hu, Y.; Brewer, G. Competitive binding of AUF1 and TIAR to MYC mRNA controls its translation. Nat. Struct. Mol. Biol. 2007, 14, 511–518. [Google Scholar] [CrossRef]
- Damgaard, C.K.; Lykke-Andersen, J. Translational coregulation of 5’TOP mRNAs by TIA-1 and TIAR. Genes Dev. 2011, 25, 2057–2068. [Google Scholar] [CrossRef] [Green Version]
- Miloslavski, R.; Cohen, E.; Avraham, A.; Iluz, Y.; Hayouka, Z.; Kasir, J.; Mudhasani, R.; Jones, S.N.; Cybulski, N.; Rüegg, M.A.; et al. Oxygen sufficiency controls TOP mRNA translation via the TSC-Rheb-mTOR pathway in a 4E-BP-independent manner. J. Mol. Cell. Biol. 2014, 6, 255–266. [Google Scholar] [CrossRef] [Green Version]
- Fred, R.G.; Mehrabi, S.; Adams, C.M.; Welsh, N. PTB and TIAR binding to insulin mRNA 3’- and 5’-UTRs; implications for insulin biosynthesis and messenger stability. Heliyon 2016, 2, e00159. [Google Scholar] [CrossRef] [Green Version]
- Marchese, D.; Botta-Orfila, T.; Cirillo, D.; Rodriguez, J.A.; Livi, C.M.; Fernández-Santiago, R.; Ezquerra, M.; Martí, M.J.; Bechara, E.; Tartaglia, G.G.; et al. Registry (CMSAR). Discovering the 3′ UTR-mediated regulation of alpha-synuclein. Nucleic Acids Res. 2017, 45, 12888–12903. [Google Scholar] [CrossRef]
- Carrascoso, I.; Sánchez-Jiménez, C.; Izquierdo, J.M. Genome-wide profiling reveals a role for T-cell intracellular antigens TIA1 and TIAR in the control of translational specificity in HeLa cells. Biochem. J. 2014, 461, 43–50. [Google Scholar] [CrossRef]
- Carrascoso, I.; Sánchez-Jiménez, C.; Izquierdo, J.M. Long-term reduction of T-cell intracellular antigens leads to increased beta-actin expression. Mol. Cancer 2014, 13, 90. [Google Scholar] [CrossRef] [Green Version]
- Baez, M.V.; Boccaccio, G.L. Mammalian Smaug is a translational repressor that forms cytoplasmic foci similar to stress granules. J. Biol. Chem. 2005, 280, 43131–43140. [Google Scholar] [CrossRef] [Green Version]
- Connolly, E.P.; Thuillier, V.; Rouy, D.; Bouétard, G.; Schneider, R.J. Inhibition of Cap-initiation complexes linked to a novel mechanism of eIF4G depletion in acute myocardial ischemia. Cell Death. Differ. 2006, 13, 1586–1594. [Google Scholar] [CrossRef] [Green Version]
- Reinhardt, H.C.; Hasskamp, P.; Schmedding, I.; Morandell, S.; van Vugt, M.A.; Wang, X.; Linding, R.; Ong, S.E.; Weaver, D.; Carr, S.A.; et al. DNA damage activates a spatially distinct late cytoplasmic cell-cycle checkpoint network controlled by MK2-mediated RNA stabilization. Mol. Cell 2010, 40, 34–49. [Google Scholar] [CrossRef]
- Velázquez-Cruz, A.; Baños-Jaime, B.; Díaz-Quintana, A.; De la Rosa, M.A.; Díaz-Moreno, I. Post-translational control of RNA-binding proteins and disease-related dysregulation. Front. Mol. Biosci. 2021, 8, 658852. [Google Scholar] [CrossRef]
- Dean, J.L.; Sully, G.; Clark, A.R.; Saklatvala, J. The involvement of AU-rich element-binding proteins in p38 mitogen-activated protein kinase pathway-mediated mRNA stabilisation. Cell Signal 2004, 16, 1113–1121. [Google Scholar] [CrossRef]
- Duttagupta, R.; Tian, B.; Wilusz, C.J.; Khounh, D.T.; Soteropoulos, P.; Ouyang, M.; Dougherty, J.P.; Peltz, S.W. Global analysis of Pub1p targets reveals a coordinate control of gene expression through modulation of binding and stability. Mol. Cell. Biol. 2005, 25, 5499–5513. [Google Scholar] [CrossRef] [Green Version]
- García-Mauriño, S.M.; Rivero-Rodríguez, F.; Velázquez-Cruz, A.; Hernández-Vellisca, M.; Díaz-Quintana, A.; De la Rosa, M.A.; Díaz-Moreno, I. RNA binding protein regulation and cross-talk in the control of AU-rich mRNA fate. Front. Mol. Biosci. 2017, 4, 71. [Google Scholar] [CrossRef]
- Fechir, M.; Linker, K.; Pautz, A.; Hubrich, T.; Kleinert, H. The RNA binding protein TIAR is involved in the regulation of human iNOS expression. Cell. Mol. Biol. 2005, 51, 299–305. [Google Scholar]
- Motohashi, H.; Mukudai, Y.; Ito, C.; Kato, K.; Shimane, T.; Kondo, S.; Shirota, T. Tumor protein D52 expression is post-transcriptionally regulated by T-cell intercellular antigen (TIA) 1 and TIA-related protein via mRNA stability. Biochem. J. 2017, 474, 1669–1687. [Google Scholar] [CrossRef] [PubMed]
- Stoecklin, G.; Stubbs, T.; Kedersha, N.; Wax, S.; Rigby, W.F.; Blackwell, T.K.; Anderson, P. MK2-induced tristetraprolin: 14-3-3 complexes prevent stress granule association and ARE-mRNA decay. EMBO J. 2004, 23, 1313–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Sun, Y.; Azad, T.; Janse van Rensburg, H.J.; Luo, J.; Yang, S.; Liu, P.; Lv, Z.; Zhan, M.; Lu, L.; et al. Rox8 promotes microRNA-dependent yki messenger RNA decay. Proc. Natl. Acad. Sci. USA 2020, 117, 30520–30530. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Yue, H.; Liu, Q.; Yuan, J.; Li, J.; Wei, G.; Chen, X.; Lu, Y.; Guo, M.; Luo, J.; et al. LncRNA MT1JP functions as a tumor suppressor by interacting with TIAR to modulate the p53 pathway. Oncotarget 2016, 7, 15787–15800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, A.T.; Berasain, C.; Bhatia, S.; Rivera, K.; Liu, B.; Rigo, F.; Pappin, D.J.; Spector, D.L. PHAROH lncRNA regulates Myc translation in hepatocellular carcinoma via sequestering TIAR. eLife 2021, 10, e68263. [Google Scholar] [CrossRef]
- Yi, B.; Li, H.; Cai, H.; Lou, X.; Yu, M.; Li, Z. LOXL1-AS1 communicating with TIAR modulates vasculogenic mimicry in glioma via regulation of the miR-374b-5p/MMP14 axis. J. Cell. Mol. Med. 2022, 26, 475–490. [Google Scholar] [CrossRef]
- Suvanto, M.; Beesley, J.; Blomqvist, C.; Chenevix-Trench, G.; Khan, S.; Nevanlinna, H. SNPs in lncRNA regions and breast cancer risk. Front. Genet. 2020, 11, 550. [Google Scholar] [CrossRef]
- Wang, Q.; Yu, H.; Yu, H.; Ma, M.; Ma, Y.; Li, R. miR-223-3p/TIAL1 interaction is involved in the mechanisms associated with the neuroprotective effects of dexmedetomidine on hippocampal neuronal cells in vitro. Mol. Med. Rep. 2019, 19, 805–812. [Google Scholar] [CrossRef] [Green Version]
- El Gazzar, M.; McCall, C.E. MicroRNAs distinguish translational from transcriptional silencing during endotoxin tolerance. J. Biol. Chem. 2010, 285, 20940–20951. [Google Scholar] [CrossRef] [Green Version]
- Collino, F.; Deregibus, M.C.; Bruno, S.; Sterpone, L.; Aghemo, G.; Viltono, L.; Tetta, C.; Camussi, G. Microvesicles derived from adult human bone marrow and tissue specific mesenchymal stem cells shuttle selected pattern of miRNAs. PLoS ONE 2010, 5, e11803. [Google Scholar] [CrossRef]
- Beck, A.R.; Miller, I.J.; Anderson, P.; Streuli, M. RNA-binding protein TIAR is essential for primordial germ cell development. Proc. Natl. Acad. Sci. USA 1998, 95, 2331–2336. [Google Scholar] [CrossRef] [Green Version]
- Oleana, V.H.; Salehi, A.; Swaab, D.F. Increased expression of the TIAR protein in the hippocampus of Alzheimer patients. NeuroReport 1998, 9, 1451–1454. [Google Scholar] [CrossRef] [Green Version]
- Kedersha, N.L.; Gupta, M.; Li, W.; Miller, I.; Anderson, P. RNA-binding proteins TIA-1 and TIAR link the phosphorylation of eIF-2 alpha to the assembly of mammalian stress granules. J. Cell. Biol. 1999, 147, 1431–1442. [Google Scholar] [CrossRef]
- Kedersha, N.; Chen, S.; Gilks, N.; Li, W.; Miller, I.J.; Stahl, J.; Anderson, P. Evidence that ternary complex (eIF2-GTP-tRNA(i)(Met))-deficient preinitiation complexes are core constituents of mammalian stress granules. Mol. Biol. Cell. 2002, 13, 195–210. [Google Scholar] [CrossRef] [Green Version]
- Iseni, F.; Garcin, D.; Nishio, M.; Kedersha, N.; Anderson, P.; Kolakofsky, D. Sendai virus trailer RNA binds TIAR, a cellular protein involved in virus-induced apoptosis. EMBO J. 2002, 21, 5141–5150. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Li, Y.; Kedersha, N.; Anderson, P.; Emara, M.; Swiderek, K.M.; Moreno, G.T.; Brinton, M.A. Cell proteins TIA-1 and TIAR interact with the 3’ stem-loop of the West Nile virus complementary minus-strand RNA and facilitate virus replication. J. Virol. 2002, 76, 11989–12000. [Google Scholar] [CrossRef] [Green Version]
- Le Guiner, C.; Gesnel, M.C.; Breathnach, R. TIA-1 or TIAR is required for DT40 cell viability. J. Biol. Chem. 2003, 278, 10465–10476. [Google Scholar] [CrossRef] [Green Version]
- Emara, M.M.; Brinton, M.A. Interaction of TIA-1/TIAR with West Nile and dengue virus products in infected cells interferes with stress granule formation and processing body assembly. Proc. Natl. Acad. Sci. USA 2007, 104, 9041–9046. [Google Scholar] [CrossRef] [Green Version]
- Emara, M.M.; Liu, H.; Davis, W.G.; Brinton, M.A. Mutation of mapped TIA-1/TIAR binding sites in the 3’ terminal stem-loop of West Nile virus minus-strand RNA in an infectious clone negatively affects genomic RNA amplification. J. Virol. 2008, 82, 10657–10670. [Google Scholar] [CrossRef] [Green Version]
- Kharraz, Y.; Salmand, P.-A.; Camus, A.; Auriol, J.; Gueydan, C.; Kruys, V.; Morello, D. Impaired embryonic development in mice overexpressing the RNA-binding protein TIAR. PLoS ONE 2010, 5, e11352. [Google Scholar] [CrossRef] [Green Version]
- Izquierdo, J.M.; Alcalde, J.; Carrascoso, I.; Reyes, R.; Ludeña, M.D. Knockdown of T-cell intracellular antigens triggers cell proliferation, invasion and tumour growth. Biochem. J. 2011, 435, 337–344. [Google Scholar] [CrossRef]
- Garaigorta, U.; Heim, M.H.; Boyd, B.; Wieland, S.; Chisari, F.V. Hepatitis C virus (HCV) induces formation of stress granules whose proteins regulate HCV RNA replication and virus assembly and egress. J. Virol. 2012, 86, 11043–11056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Jiménez, C.; Izquierdo, J.M. T-cell intracellular antigen (TIA)-proteins deficiency in murine embryonic fibroblasts alters cell cycle progression and induces autophagy. PLoS ONE 2013, 8, e75127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ash, P.E.; Vanderweyde, T.E.; Youmans, K.L.; Apicco, D.J.; Wolozin, B. Pathological stress granules in Alzheimer’s disease. Brain Res. 2014, 1584, 52–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Jiménez, C.; Ludeña, M.D.; Izquierdo, J.M. T-cell intracellular antigens function as tumor suppressor genes. Cell Death Dis. 2015, 6, e1669. [Google Scholar] [CrossRef] [PubMed]
- Bley, N.; Lederer, M.; Pfalz, B.; Reinke, C.; Fuchs, T.; Glaß, M.; Möller, B.; Hüttelmaier, S. Stress granules are dispensable for mRNA stabilization during cellular stress. Nucleic Acids Res. 2015, 43, e26. [Google Scholar] [CrossRef]
- Geng, Z.; Li, P.; Tan, L.; Song, H. Targeted knockdown of RNA-binding protein TIAR for promoting self-renewal and attenuating differentiation of mouse embryonic stem cells. Stem Cells Int. 2015, 2015, 657325. [Google Scholar] [CrossRef] [Green Version]
- Kharraz, Y.; Lefort, A.; Libert, F.; Christopher, J.; Mann, C.J.; Gueydan, C.; Kruys, V. Genome-wide analysis of TIAR RNA ligands in mouse macrophages before and after LPS stimulation. Genom. Data 2016, 7, 297–300. [Google Scholar] [CrossRef]
- Tak, H.; Eun, J.W.; Kim, J.; Park, S.J.; Kim, C.; Ji, E.; Lee, H.; Kang, H.; Cho, D.H.; Lee, K.; et al. T-cell-restricted intracellular antigen 1 facilitates mitochondrial fragmentation by enhancing the expression of mitochondrial fission factor. Cell Death Differ. 2017, 24, 49–58. [Google Scholar] [CrossRef] [Green Version]
- Carrascoso, I.; Alcalde, J.; Sánchez-Jiménez, C.; González-Sánchez, P.; Izquierdo, J.M. T-cell intracellular antigens and Hu antigen R antagonistically modulate mitochondrial activity and dynamics by regulating optic atrophy 1 gene expression. Mol. Cell. Biol. 2017, 37, e0017417. [Google Scholar] [CrossRef] [Green Version]
- Hou, S.; Kumar, A.; Xu, Z.; Airo, A.M.; Stryapunina, I.; Wong, C.P.; Branton, W.; Tchesnokov, E.; Götte, M.; Power, C.; et al. Zika virus hijacks stress granule proteins and modulates the host stress response. J. Virol. 2017, 91, e00474-17. [Google Scholar] [CrossRef] [Green Version]
- Battu, S.; Afroz, S.; Giddaluru, J.; Naz, S.; Huang, W.; Khumukcham, S.S.; Khan, R.A.; Bhat, S.Y.; Qureshi, I.A.; Manavathi, B.; et al. Amino acid starvation sensing dampens IL-1β production by activating riboclustering and autophagy. PLoS Biol. 2018, 16, e2005317. [Google Scholar] [CrossRef]
- Naz, S.; Khan, R.A.; Giddaluru, J.; Battu, S.; Vishwakarma, S.K.; Subahan, M.; Satti, V.; Khan, N.; Khan, A.A. Transcriptome meta-analysis identifies immune signature comprising of RNA binding proteins in ulcerative colitis patients. Cell Immunol. 2018, 334, 42–48. [Google Scholar] [CrossRef]
- Lafarga, V.; Sung, H.-M.; Haneke, K.; Roessig, L.; Pauleau, A.-L.; Bruer, M.; Rodriguez-Acebes, S.; Lopez-Contreras, A.J.; Gruss, O.J.; Erhardt, S.; et al. TIAR marks nuclear G2/M transition granules and restricts CDK1 activity under replication stress. EMBO Rep. 2019, 20, e46224. [Google Scholar] [CrossRef]
- Andrusiak, M.G.; Sharifnia, P.; Lyu, X.; Wang, Z.; Dickey, A.M.; Wu, Z.; Chisholm, A.D.; Jin, Y. Inhibition of axon regeneration by liquid-like TIAR-2 granules. Neuron 2019, 104, 290–304. [Google Scholar] [CrossRef]
- Mahboubi, H.; Moujaber, O.; Kodiha, M.; Stochaj, U. The co-chaperone HspBP1 is a novel component of stress granules that regulates their formation. Cells 2020, 9, 825. [Google Scholar] [CrossRef] [Green Version]
- Carrascoso, I.; Alcalde, J.; Tabas-Madrid, D.; Oliveros, J.C.; Izquierdo, J.M. Transcriptome-wide analysis links the short-term expression of the b isoforms of TIA proteins to protective proteostasis-mediated cell quiescence response. PLoS ONE 2018, 13, e0208526. [Google Scholar] [CrossRef]
- Naz, S.; Battu, S.; Khan, R.A.; Afroz, S.; Giddaluru, J.; Vishwakarma, S.K.; Satti, V.; Habeeb, M.A.; Khan, A.A.; Khan, N. Activation of integrated stress response pathway regulates IL-1β production through posttranscriptional and translational reprogramming in macrophages. Eur. J. Immunol. 2019, 49, 277–289. [Google Scholar] [CrossRef] [Green Version]
- Ostareck, D.H.; Ostareck-Lederer, A. RNA-binding proteins in the control of LPS-induced macrophage response. Front. Genet. 2019, 10, 31. [Google Scholar] [CrossRef] [Green Version]
- Silva-García, C.G.; Estela Navarro, R. The C. elegans TIA-1/TIAR homolog TIAR-1 is required to induce germ cell apoptosis. Genesis 2013, 51, 690–707. [Google Scholar]
- Bossowski, A.; Czarnocka, B.; Bardadin, K.; Moniuszko, A.; Łyczkowska, A.; Czerwinska, J.; Dadan, J.; Bossowska, A. Identification of chosen apoptotic (TIAR and TIA1) markers expression in thyroid tissues from adolescents with immune and non-immune thyroid diseases. Folia Histochem. Cytobiol. 2010, 48, 178–184. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Wang, S.; Zheng, L.; Li, X.; Suswam, E.A.; Zhang, X.; Wheeler, C.G.; Nabors, L.B.; Filippova, N.; King, P.H. Amyotrophic lateral sclerosis-linked mutant SOD1 sequesters Hu antigen R (HuR) and TIA-1-related protein (TIAR): Implications for impaired post-transcriptional regulation of vascular endothelial growth factor. J. Biol. Chem. 2009, 284, 33989–33998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Sun, H.; Zhang, H.; Liu, J.; Fan, F.; Li, Y.; Ning, X.; Sun, Y.; Dai, S.; Liu, B.; et al. An shRNA based genetic screen identified Sesn2 as a potential tumor suppressor in lung cancer via suppression of Akt-mTOR-p70S6K signaling. PLoS ONE 2015, 10, e0124033. [Google Scholar] [CrossRef] [Green Version]
- Ravanidis, S.; Doxakis, E. RNA-binding proteins implicated in mitochondrial damage and mitophagy. Front. Cell. Dev. Biol. 2020, 8, 372. [Google Scholar] [CrossRef] [PubMed]
- Carrascoso, I.; Sánchez-Jiménez, C.; Silion, E.; Alcalde, J.; Izquierdo, J.M. A heterologous cell model for studying the role of T-cell intracellular antigen 1 in Welander distal myopathy. Mol. Cell. Biol. 2019, 39, e0029918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yongjie Cen, Y.; Zou, X.; Zhong, Q.; Chen, Y.; Lin, Y.; Feng, Q.; Wang, X.; Zheng, S. The TIAR-mediated Nrf2 response to oxidative stress is mediated through the Nrf2 noncoding 3’untranslated region in Spodoptera litura. Free Radic. Biol. Med. 2022, 184, 17–29. [Google Scholar]
- Dinkova-Kostovaa, A.T.; Abramovc, A.Y. The emerging role of Nrf2 in mitochondrial function. Free Radic Biol Med. 2015, 88, 179–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MSGP—Database. Available online: https://msgp.pt/ (accessed on 20 May 2022).
- RNP Granule Database. Available online: https://rnagranuledb.lunenfeld.ca/ (accessed on 20 May 2022).
- Esclatine, A.; Taddeo, B.; Roizman, B. Herpes simplex virus 1 induces cytoplasmic accumulation of TIA-1/TIAR and both synthesis and cytoplasmic accumulation of tristetraprolin, two cellular proteins that bind and destabilize AU-rich RNAs. J. Virol. 2004, 78, 8582–8592. [Google Scholar] [CrossRef] [Green Version]
- Thomas, M.G.; Martinez Tosar, L.J.; Loschi, M.; Pasquini, J.M.; Correale, J.; Kindler, S.; Boccaccio, G.L. Staufen recruitment into stress granules does not affect early mRNA transport in oligodendrocytes. Mol. Biol. Cell. 2005, 16, 405–420. [Google Scholar] [CrossRef] [Green Version]
- Sampuda, K.M.; Riley, M.; Boyd, L. Stress induced nuclear granules form in response to accumulation of misfolded proteins in Caenorhabditis elegans. BMC Cell Biol. 2017, 18, 18. [Google Scholar] [CrossRef] [Green Version]
- Wiegand, M.; Bossow, S.; Neubert, W.J. Sendai virus trailer RNA simultaneously blocks two apoptosis-inducing mechanisms in a cell type-dependent manner. J. Gen. Virol. 2005, 86, 2305–2314. [Google Scholar] [CrossRef]
- Moujaber, O.; Mahboubi, H.; Kodiha, M.; Bouttier, M.; Bednarz, K.; Bakshi, R.; White, J.; Larose, L.; Colmegna, I.; Stochaj, U. Dissecting the molecular mechanisms that impair stress granule formation in aging cells. Biochim. Biophys. Acta Mol. Cell. Res. 2017, 1864, 475–486. [Google Scholar] [CrossRef]
- Delestienne, N.; Wauquier, C.; Soin, R.; Dierick, J.F.; Gueydan, C.; Kruys, V. The splicing factor ASF/SF2 is associated with TIA-1-related/TIA-1-containing ribonucleoproteic complexes and contributes to post-transcriptional repression of gene expression. FEBS J. 2010, 277, 2496–2514. [Google Scholar] [CrossRef]
- Tessier, S.N.; Audas, T.E.; Wu, C.W.; Lee, S.; Storey, K.B. The involvement of mRNA processing factors TIA-1, TIAR, and PABP-1 during mammalian hibernation. Cell Stress Chaperones 2014, 19, 813–825. [Google Scholar] [CrossRef] [Green Version]
- Weeks, A.; Agnihotri, S.; Lymer, J.; Chalil, A.; Diaz, R.; Isik, S.; Smith, C.; Rutka, J.T. Epithelial cell transforming 2 and aurora kinase B modulate formation of stress granule-containing transcripts from diverse cellular pathways in astrocytoma cells. Am. J. Pathol. 2016, 186, 1674–1687. [Google Scholar] [CrossRef] [Green Version]
- Palangi, F.; Samuel, S.M.; Thompson, I.R.; Triggle, C.R.; Emara, M.M. Effects of oxidative and thermal stresses on stress granule formation in human induced pluripotent stem cells. PLoS ONE 2017, 12, e0182059. [Google Scholar] [CrossRef] [Green Version]
- Lechler, M.C.; David, D.C. More stressed out with age? Check your RNA granule aggregation. Prion 2017, 11, 313–322. [Google Scholar] [CrossRef] [Green Version]
- Kuo, C.T.; You, G.T.; Jian, Y.J.; Chen, T.S.; Siao, Y.C.; Hsu, A.L.; Ching, T.T. AMPK-mediated formation of stress granules is required for dietary restriction-induced longevity in Caenorhabditis elegans. Aging Cell 2020, 19, e13157. [Google Scholar] [CrossRef]
- Lo Bello, M.; Di Fini, F.; Notaro, A.; Spataro, R.; Conforti, F.L.; La Bella, V. ALS-related mutant FUS protein is mislocalized to cytoplasm and is recruited into stress granules of fibroblasts from asymptomatic FUS P525L mutation carriers. Neurodegener. Dis. 2017, 17, 292–303. [Google Scholar] [CrossRef]
- Buddika, K.; Ariyapala, I.S.; Hazuga, M.A.; Riffert, D.; Sokol, N.S. Canonical nucleators are dispensable for stress granule assembly in Drosophila intestinal progenitors. J. Cell. Sci. 2020, 133, jcs243451. [Google Scholar] [CrossRef]
- Aulas, A.; Finetti, P.; Lyons, S.M.; Bertucci, F.; Birnbaum, D.; Acquaviva, C.; Mamessier, E. Revisiting the concept of stress in the prognosis of solid tumors: A role for stress granules proteins? Cancers 2020, 12, 2470. [Google Scholar] [CrossRef]
- Asadi, M.R.; Rahmanpour, D.; Moslehian, M.S.; Sabaie, H.; Hassani, M.; Ghafouri-Fard, S.; Taheri, M.; Rezazadeh, M. Stress granules involved in formation, progression and metastasis of cancer: A scoping review. Front. Cell. Dev. Biol. 2021, 9, 745394. [Google Scholar] [CrossRef] [PubMed]
- Sidali, A.; Teotia, V.; Solaiman, N.S.; Bashir, N.; Kanagaraj, R.; Murphy, J.J.; Surendranath, K. AU-rich element RNA binding proteins: At the crossroads of post-Transcriptional regulation and genome integrity. Int. J. Mol. Sci. 2021, 23, 96. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, R.E. Nuclear proteins hijacked by mammalian cytoplasmic plus strand RNA viruses. Virology 2015, 479–480, 457–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCormick, C.; Khaperskyy, D.A. Translation inhibition and stress granules in the antiviral immune response. Nat. Rev. Immunol. 2017, 17, 647–660. [Google Scholar] [CrossRef]
- Le Sage, V.; Cinti, A.; McCarthy, S.; Amorim, R.; Rao, S.; Daino, G.L.; Tramontano, E.; Branch, D.R.; Mouland, A.J. Ebola virus VP35 blocks stress granule assembly. Virology 2017, 502, 73–83. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Dong, L.; Yu, S.; Wang, X.; Zheng, H.; Zhang, P.; Meng, C.; Zhan, Y.; Tan, L.; Song, C.; et al. Newcastle disease virus induces stable formation of bona fide stress granules to facilitate viral replication through manipulating host protein translation. FASEB J. 2017, 31, 1337–1353. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wang, H.; Li, Y.; Jin, Y.; Chu, Y.; Su, A.; Wu, Z. TIA-1 and TIAR interact with 5’-UTR of enterovirus 71 genome and facilitate viral replication. Biochem. Biophys. Res. Commun. 2015, 466, 254–259. [Google Scholar] [CrossRef]
- White, J.P.; Cardenas, A.M.; Marissen, W.E.; Lloyd, R.E. Inhibition of cytoplasmic mRNA stress granule formation by a viral proteinase. Cell Host Microbe 2007, 2, 295–305. [Google Scholar] [CrossRef] [Green Version]
- Soto-Rifo, R.; Valiente-Echeverria, F.; Rubilar, P.S.; Garcia-de-Gracia, F.; Ricci, E.P.; Limousin, T.; Décimo, D.; Mouland, A.J.; Ohlmann, T. HIV-2 genomic RNA accumulates in stress granules in the absence of active translation. Nucleic Acids Res. 2014, 42, 12861–12875. [Google Scholar] [CrossRef]
- Valiente-Echeverría, F.; Melnychuk, L.; Vyboh, K.; Ajamian, L.; Gallouzi, I.E.; Bernard, N.; Mouland, A.J. eEF2 and Ras-GAP SH3 domain-binding protein (G3BP1) modulate stress granule assembly during HIV-1 infection. Nat. Commun. 2014, 5, 4819. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Fang, L.; Wang, D.; Cai, K.; Chen, H.; Xiao, S. Porcine reproductive and respiratory syndrome virus infection induces stress granule formation depending on protein kinase R-like endoplasmic reticulum kinase (PERK) in MARC-145 cells. Front. Cell. Infect. Microbiol. 2017, 7, 111. [Google Scholar] [CrossRef] [Green Version]
- Albornoz, A.; Carletti, T.; Corazza, G.; Marcello, A. The stress granule component TIA-1 binds tick-borne encephalitis virus RNA and is recruited to perinuclear sites of viral replication to inhibit viral translation. J. Virol. 2014, 88, 6611–6622. [Google Scholar] [CrossRef] [Green Version]
- Bonenfant, G.; Williams, N.; Netzband, R.; Schwarz, M.C.; Evans, M.J.; Pager, C.T. Zika virus subverts stress granules to promote and restrict viral gene expression. J. Virol. 2019, 93, e0052019. [Google Scholar] [CrossRef] [Green Version]
- López-Urrutia, E.; Valdés, J.; Bonilla-Moreno, R.; Martínez-Salazar, M.; Martínez-Garcia, M.; Berumen, J.; Villegas-Sepúlveda, N. A few nucleotide polymorphisms are sufficient to recruit nuclear factors differentially to the intron 1 of HPV-16 intratypic variants. Virus Res. 2012, 166, 43–53. [Google Scholar] [CrossRef]
- Dinh, P.X.; Beura, L.K.; Das, P.B.; Panda, D.; Das, A.; Pattnaik, A.K. Induction of stress granule-like structures in vesicular stomatitis virus-infected cells. J. Virol. 2013, 87, 372–383. [Google Scholar] [CrossRef] [Green Version]
- Catanzaro, N.; Meng, X.J. Porcine reproductive and respiratory syndrome virus (PRRSV)-induced stress granules are associated with viral replication complexes and suppression of host translation. Virus Res. 2019, 265, 47–56. [Google Scholar] [CrossRef]
- Álvarez, E.; Castelló, A.; Carrasco, L.; Izquierdo, J.M. Poliovirus 2A protease triggers a selective nucleo-cytoplasmic redistribution of splicing factors to regulate alternative pre-mRNA splicing. PLoS ONE 2013, 8, e73723. [Google Scholar] [CrossRef] [Green Version]
- Choi, E.Y.; Pintel, D. Splicing of the large intron present in the nonstructural gene of minute virus of mice is governed by TIA-1/TIAR binding downstream of the nonconsensus donor. J. Virol. 2009, 83, 6306–6311. [Google Scholar] [CrossRef] [Green Version]
- Katsanou, V.; Papadaki, O.; Milatos, S.; Blackshear, P.J.; Anderson, P.; Kollias, G.; Kontoyiannis, D.L. HuR as a negative posttranscriptional modulator in inflammation. Mol. Cell 2005, 19, 777–789. [Google Scholar] [CrossRef]
- Akira, S.; Maeda, K. Control of RNA stability in immunity. Annu. Rev. Immunol. 2021, 39, 481–509. [Google Scholar] [CrossRef]
- Suswam, E.A.; Nabors, L.B.; Huang, Y.; Yang, X.; King, P.H. IL-1beta induces stabilization of IL-8 mRNA in malignant breast cancer cells via the 3’ untranslated region: Involvement of divergent RNA-binding factors HuR, KSRP and TIAR. Int. J. Cancer 2005, 113, 911–919. [Google Scholar] [CrossRef]
- Jimenez-Boj, E.; Kedersha, N.; Tohidast-Akrad, M.; Karlhofer, F.M.; Stummvoll, G.; Zimmermann, C.; Ulrich, W.; Guiducci, S.; Hoefler, E.; Aringer, M.; et al. Autoantibodies to the translational suppressors T cell intracytoplasmic antigen 1 and T cell intracytoplasmic antigen 1-related protein in patients with rheumatic diseases: Increased prevalence in systemic lupus erythematosus and systemic sclerosis and correlation with clinical features. Arthritis Rheum. 2008, 58, 1226–1236. [Google Scholar]
- Ravi, A.; Chowdhury, S.; Dijkhuis, A.; Bonta, P.I.; Sterk, P.J.; Lutter, R. Neutrophilic inflammation in asthma and defective epithelial translational control. Eur. Respir. J. 2019, 54, 1900547. [Google Scholar] [CrossRef]
- Navratilova, Z.; Novosadova, E.; Hagemann-Jensen, M.; Kullberg, S.; Kolek, V.; Grunewald, J.; Petrek, M. Expression profile of six RNA-binding proteins in pulmonary sarcoidosis. PLoS ONE 2016, 11, e0161669. [Google Scholar] [CrossRef] [Green Version]
- Yamaji, M.; Tanaka, T.; Shigeta, M.; Chuma, S.; Saga, Y.; Saitou, M. Functional reconstruction of NANOS3 expression in the germ cell lineage by a novel transgenic reporter reveals distinct subcellular localizations of NANOS3. Reproduction 2010, 139, 381–393. [Google Scholar] [CrossRef] [Green Version]
- Huelgas-Morales, G.; Silva-García, C.G.; Salinas, L.S.; Greenstein, D.; Navarro, R.E. The stress granule RNA-binding protein TIAR-1 protects female germ cells from heat shock in Caenorhabditis elegans. G3 Genes Genomes Genet. 2016, 6, 1031–1047. [Google Scholar]
- Chen, J.; Wang, J.; Qian, J.; Bao, M.; Zhang, X.; Huang, Z. MBNL1 suppressed cancer metastatic of skin squamous cell carcinoma via by TIAL1/MYOD1/caspase-9/3 signaling pathways. Technol. Cancer Res. Treat. 2021, 20. [Google Scholar] [CrossRef]
- Tian, K.; Qi, W.; Yan, Q.; Lv, M.; Song, D. Signature constructed by glycolysis-immune-related genes can predict the prognosis of osteosarcoma patients. Investig. New Drugs 2022, 40, 818–830. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Toomer, K.H.; Chen, Z.; Cai, X. Genome-wide analysis of alternative transcripts in human breast cancer. Breast Cancer Res. Treat. 2015, 151, 295–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottschald, O.R.; Malec, V.; Krasteva, G.; Hasan, D.; Kamlah, F.; Herold, S.; Rose, F.; Seeger, W.; Hänze, J. TIAR and TIA-1 mRNA-binding proteins co-aggregate under conditions of rapid oxygen decline and extreme hypoxia and suppress the HIF-1α pathway. J. Mol. Cell. Biol. 2010, 2, 345–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podszywalow-Bartnicka, P.; Wolczyk, M.; Kusio-Kobialka, M.; Wolanin, K.; Skowronek, K.; Nieborowska-Skorska, M.; Dasgupta, Y.; Skorski, T.; Piwocka, K. Downregulation of BRCA1 protein in BCR-ABL1 leukemia cells depends on stress-triggered TIAR-mediated suppression of translation. Cell Cycle 2014, 13, 3727–3741. [Google Scholar] [CrossRef] [Green Version]
- Abe, Y.; Mukudai, Y.; Kurihara, M.; Houri, A.; Chikuda, J.; Yaso, A.; Kato, K.; Shimane, T.; Shirota, T. Tumor protein D52 is upregulated in oral squamous carcinoma cells under hypoxia in a hypoxia-inducible-factor-independent manner and is involved in cell death resistance. Cell Biosci. 2021, 11, 122. [Google Scholar] [CrossRef]
- Ravanidis, S.; Kattan, F.G.; Doxakis, E. Unraveling the pathways to neuronal homeostasis and disease: Mechanistic insights into the role of RNA-binding proteins and associated factors. Int. J. Mol. Sci. 2018, 19, 2280. [Google Scholar] [CrossRef] [Green Version]
- Barron, V.A.; Zhu, H.; Hinman, M.N.; Ladd, A.N.; Lou, H. The neurofibromatosis type I pre-mRNA is a novel target of CELF protein-mediated splicing regulation. Nucleic Acids Res. 2010, 38, 253–264. [Google Scholar] [CrossRef] [Green Version]
- Barron, V.A.; Lou, H. Alternative splicing of the neurofibromatosis type I pre-mRNA. Biosci. Rep. 2012, 32, 131–138. [Google Scholar] [CrossRef] [Green Version]
- Fleming, V.A.; Geng, C.; Ladd, A.N.; Lou, H. Alternative splicing of the neurofibromatosis type 1 pre-mRNA is regulated by the muscleblind-like proteins and the CUG-BP and ELAV-like factors. BMC Mol. Biol. 2012, 13, 35. [Google Scholar] [CrossRef] [Green Version]
- Vanderweyde, T.; Yu, H.; Varnum, M.; Liu-Yesucevitz, L.; Citro, A.; Ikezu, T.; Duff, K.; Wolozin, B. Contrasting pathology of stress granule proteins TIA-1 and G3BP in tauopathies. J. Neurosci. 2012, 32, 8270–8283. [Google Scholar] [CrossRef] [Green Version]
- Vanderweyde, T.; Youmans, K.; Liu-Yesucevitz, L.; Wolozin, B. Role of stress granules and RNA-binding proteins in neurodegeneration: A mini-review. Gerontology 2013, 59, 524–533. [Google Scholar] [CrossRef] [Green Version]
- Jin, K.; Li, W.; Nagayama, T.; He, X.; Sinor, A.D.; Chang, J.; Mao, X.; Graham, S.H.; Simon, R.P.; Greenberg, D.A. Expression of the RNA-binding protein TIAR is increased in neurons after ischemic cerebral injury. J. Neurosci. Res. 2000, 59, 767–774. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Velasco, B.R.; Izquierdo, J.M. T-Cell Intracellular Antigen 1-Like Protein in Physiology and Pathology. Int. J. Mol. Sci. 2022, 23, 7836. https://doi.org/10.3390/ijms23147836
Velasco BR, Izquierdo JM. T-Cell Intracellular Antigen 1-Like Protein in Physiology and Pathology. International Journal of Molecular Sciences. 2022; 23(14):7836. https://doi.org/10.3390/ijms23147836
Chicago/Turabian StyleVelasco, Beatriz Ramos, and José M. Izquierdo. 2022. "T-Cell Intracellular Antigen 1-Like Protein in Physiology and Pathology" International Journal of Molecular Sciences 23, no. 14: 7836. https://doi.org/10.3390/ijms23147836
APA StyleVelasco, B. R., & Izquierdo, J. M. (2022). T-Cell Intracellular Antigen 1-Like Protein in Physiology and Pathology. International Journal of Molecular Sciences, 23(14), 7836. https://doi.org/10.3390/ijms23147836