
222-Nanometer Far-UVC Exposure Results in DNA Damage and Transcriptional Changes to Mammalian Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
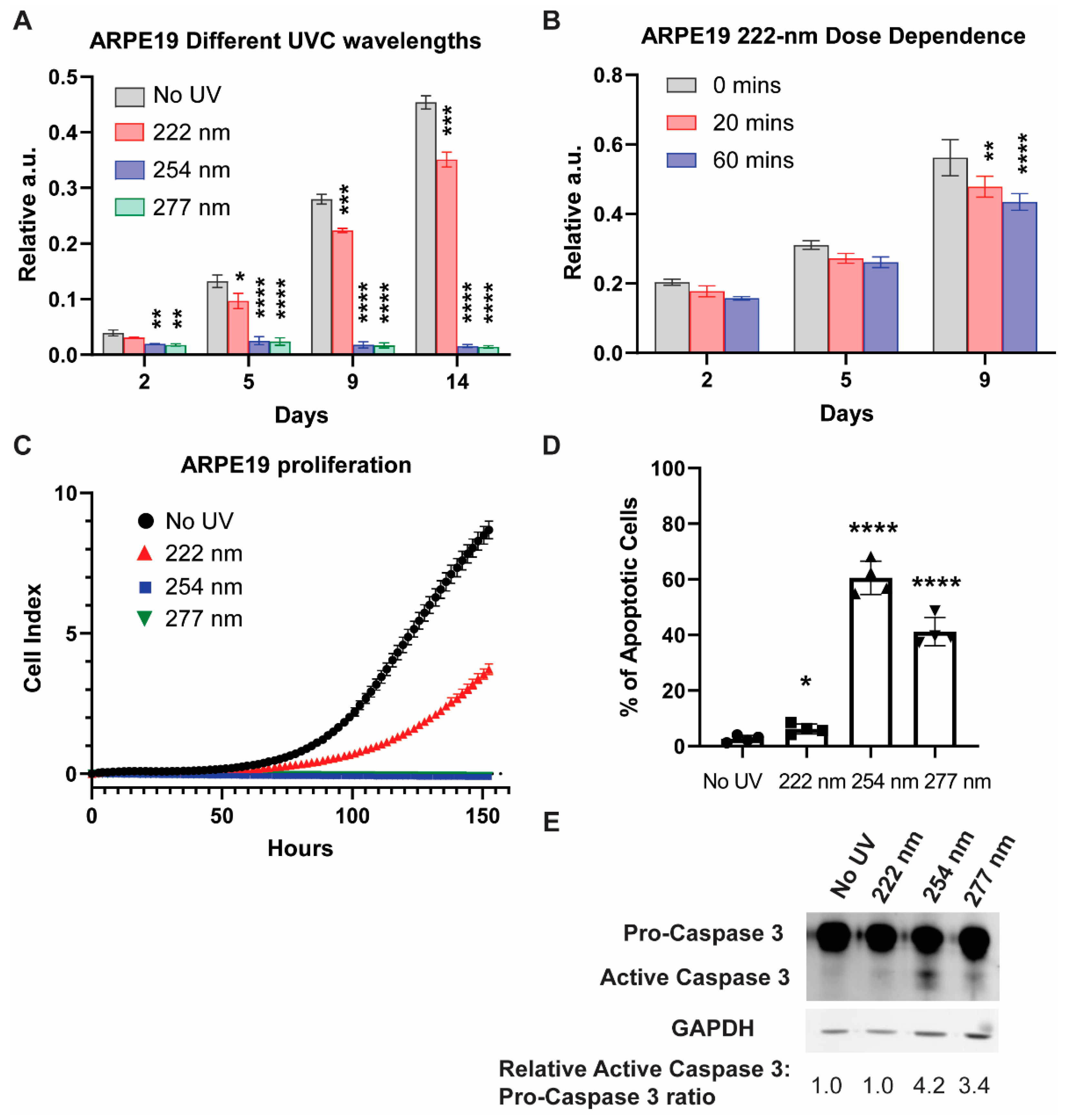
2.1. UVC-Induced Cell Viability Reduction and Apoptotic Processes
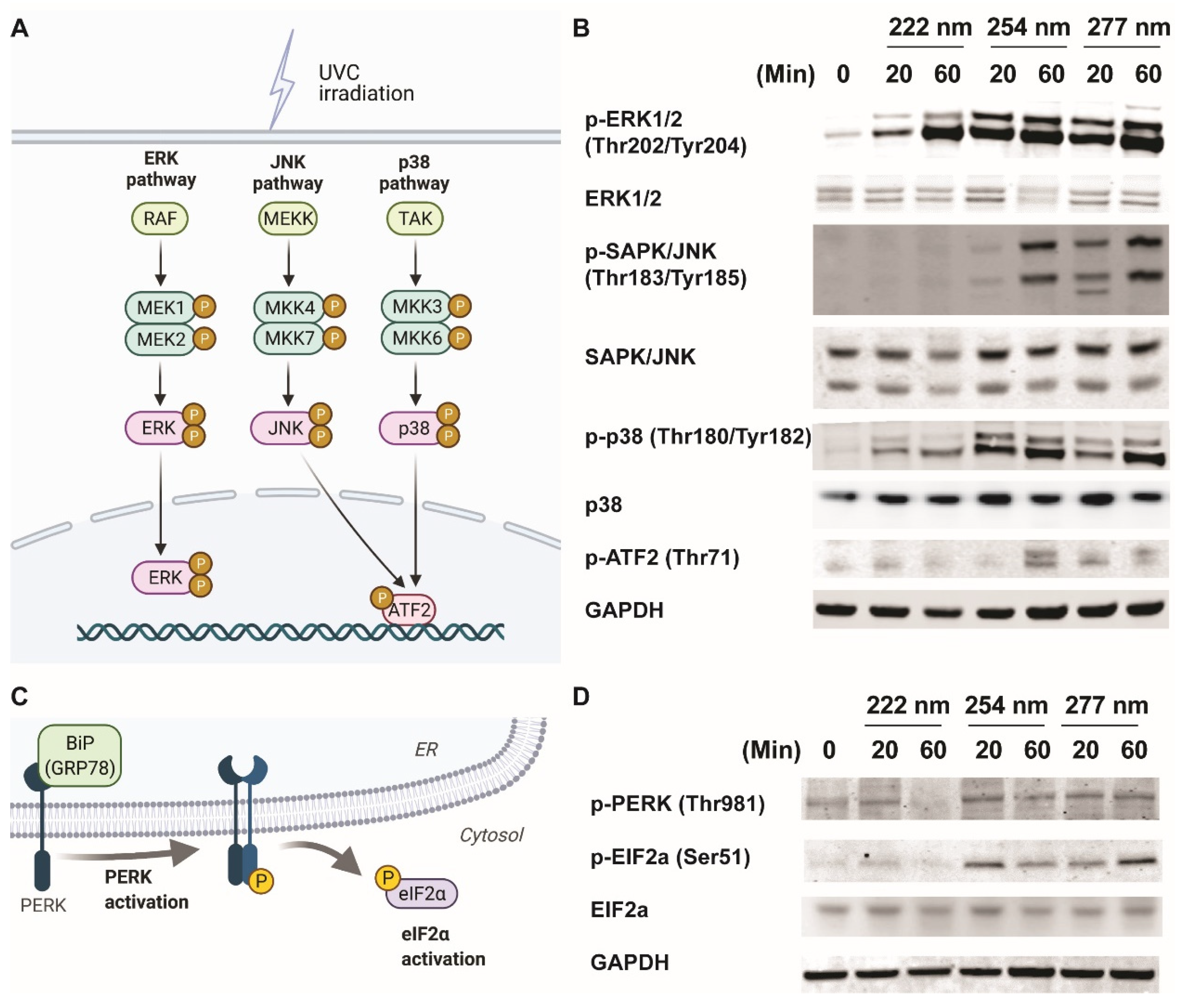
2.2. Differential Signaling Pathway Activation under Varied UVC Wavelengths
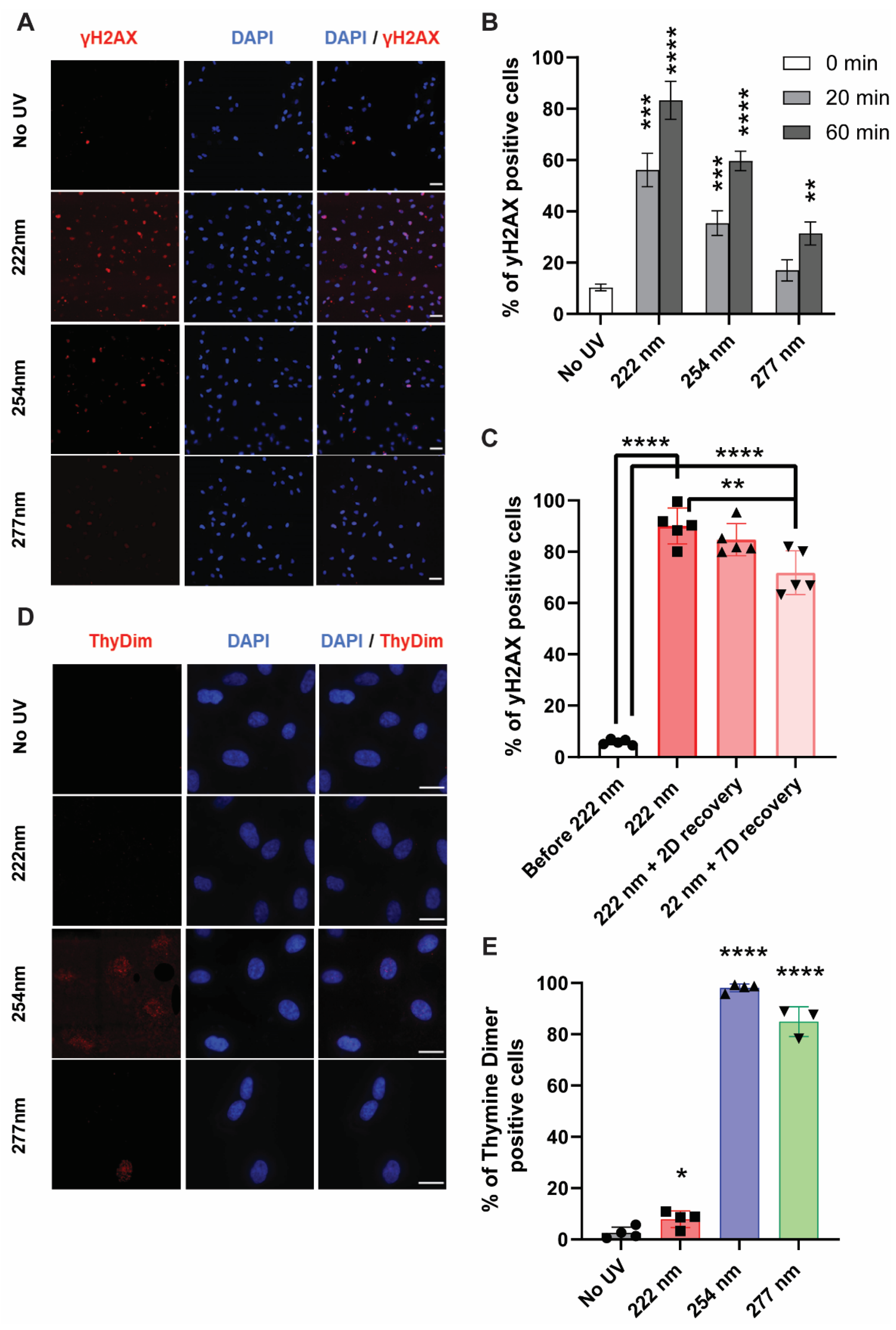
2.3. DNA-Associated Damage or Associated Repair Mechanisms Were Observed in 222-Nanometer-Lit Cells
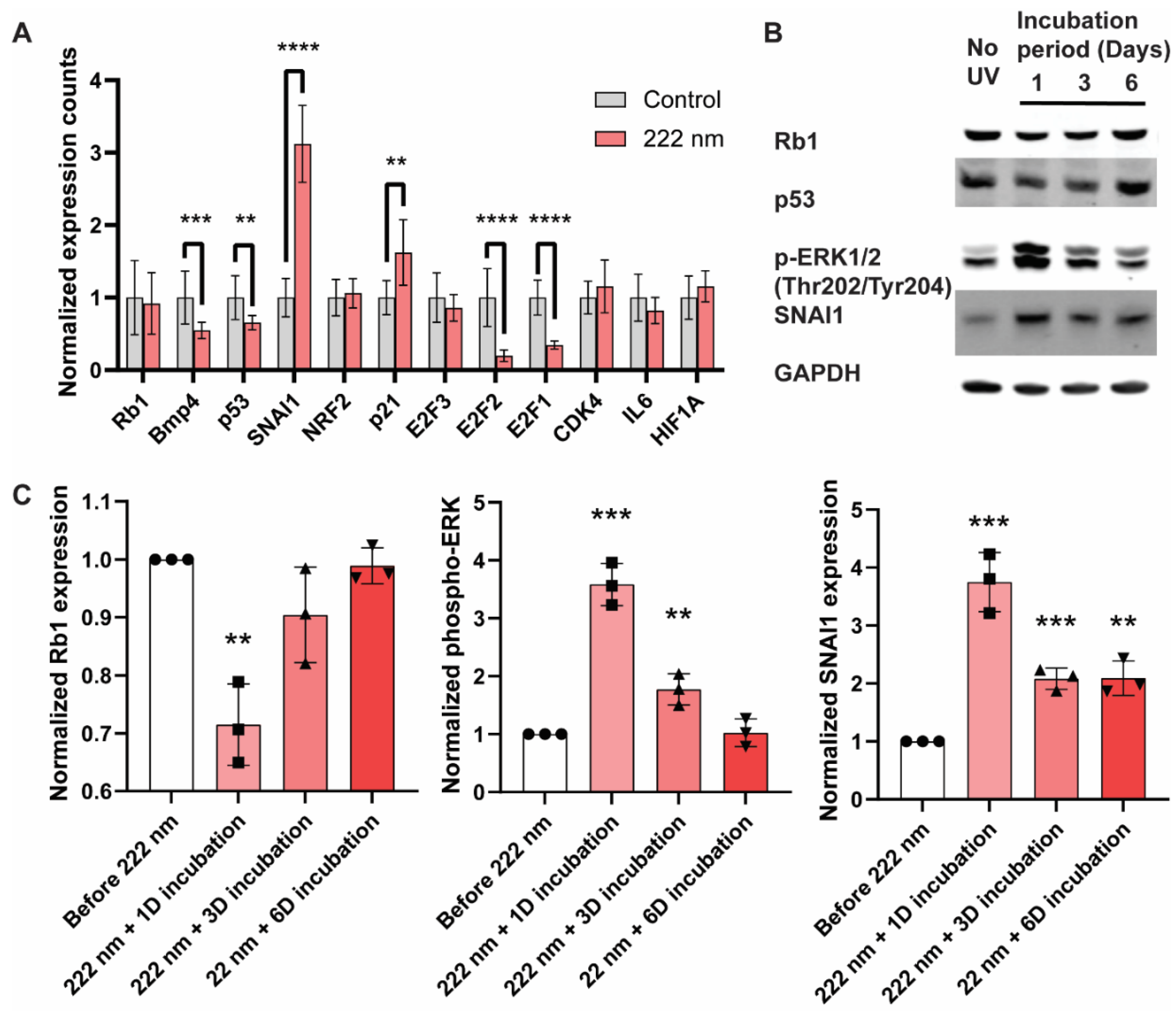
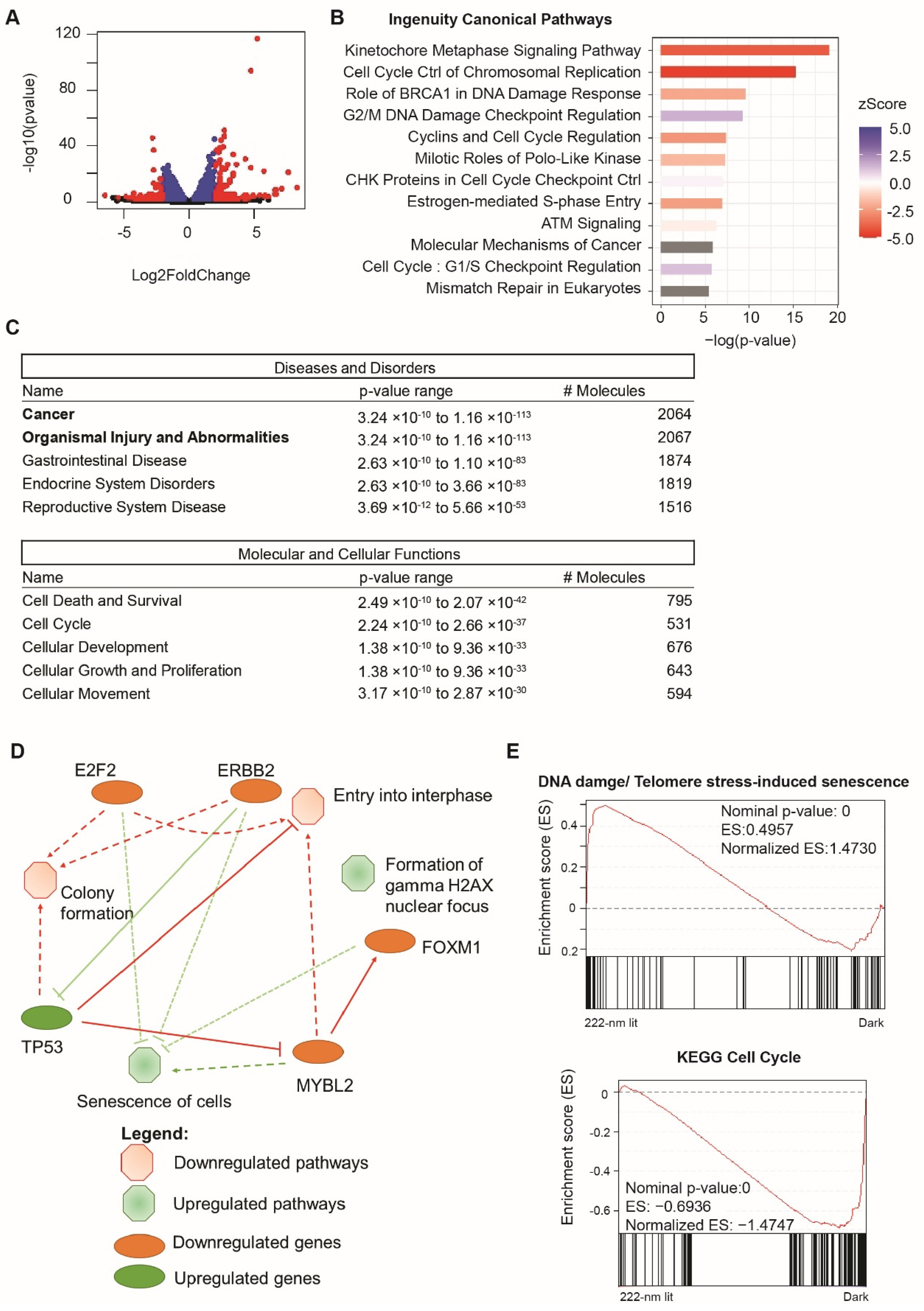
2.4. Perturbation of Transcription and Cellular Signaling Events Was Observed in 222-Nanometer-Lit Cells
2.5. Analysis of Global RNA Transcripts One Week after 222-Nanometer Exposure
3. Discussion
4. Materials and Methods
4.1. Key Resources Table
4.2. UVC Sources and Irradiance Measurements
4.3. Cell Culture
4.4. Cell Viability Assays
4.5. XCELLigence Platform
4.6. SRB Assay
4.7. Immunofluorescence Experiments
4.8. Western Blot Experiments
4.9. RNA Extraction and Analysis
4.10. RNA Sequencing
4.11. Quantification of Gene Expression
4.12. Pathway Analyses
4.13. Quantification and Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shaulian, E.; Schreiber, M.; Piu, F.; Beeche, M.; Wagner, E.F.; Karin, M. The Mammalian UV Response: C-Jun Induction Is Required for Exit from P53-Imposed Growth Arrest. Cell 2000, 103, 897–908. [Google Scholar] [CrossRef]
- Dunkern, T.R.; Fritz, G.; Kaina, B. Ultraviolet Light-Induced DNA Damage Triggers Apoptosis in Nucleotide Excision Repair-Deficient Cells via Bcl-2 Decline and Caspase-3/-8 Activation. Oncogene 2001, 20, 6026–6038. [Google Scholar] [CrossRef] [PubMed]
- Kollias, N.; Baqer, A.H. Quantitative Assessment of UV-Induced Pigmentation and Erythema. Photodermatol 1988, 5, 53–60. [Google Scholar] [PubMed]
- Armstrong, B.K.; Kricker, A. The Epidemiology of UV Induced Skin Cancer. J. Photochem. Photobiol. B Biol. 2001, 63, 8–18. [Google Scholar] [CrossRef]
- Development and Repair of Cataract Induced by Ultraviolet Radiation—ProQuest. Available online: https://www.proquest.com/openview/4ee61a6faf429bda7bf10549a195b1e6/1?cbl=2026366&diss=y&pq-origsite=gscholar (accessed on 5 August 2022).
- Wegener, A.R. In Vivo Studies on the Effect of UV-Radiation on the Eye Lens in Animals. Doc. Ophthalmol. 1994, 88, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Haag, R.; Sieber, N.; Heßling, M. Cataract Development by Exposure to Ultraviolet and Blue Visible Light in Porcine Lenses. Medicina 2021, 57, 535. [Google Scholar] [CrossRef]
- Ziegelberger, G. Icnirp Guidelines on Limits of Exposure to Laser Radiation of Wavelengths between 180 Nm and 1000 Μm. Health Phys. 2013, 105, 271–295. [Google Scholar] [CrossRef]
- Bruls, W.A.G.; Slaper, H.; van der Leun, J.C.; Berrens, L. Transmission of Human Epidermis and Stratum Corneum as A Function of Thickness in the Ultraviolet and Visible Wavelengths. Photochem. Photobiol. 1984, 40, 485–494. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Richa; Kumar, A.; Tyagi, M.B.; Sinha, R.P. Molecular Mechanisms of Ultraviolet Radiation-Induced DNA Damage and Repair. J. Nucleic Acids 2010, 2010, 592980. [Google Scholar] [CrossRef]
- Bode, A.M.; Dong, Z. Mitogen-Activated Protein Kinase Activation in UV-Induced Signal Transduction. Sci. Signal. 2003, 2003, re2. [Google Scholar] [CrossRef]
- Dhanwada, K.R.; Dickens, M.; Neades, R.; Davis, R.; Pelling, J.C. Differential Effects of UV-B and UV-C Components of Solar Radiation on MAP Kinase Signal Transduction Pathways in Epidermal Keratinocytes. Oncogene 1995, 11, 1947–1953. [Google Scholar] [PubMed]
- Kim, D.K.; Kang, D.H. UVC LED Irradiation Effectively Inactivates Aerosolized Viruses, Bacteria, and Fungi in a Chamber-Type Air Disinfection System. Appl. Environ. Microbiol. 2018, 84, e00944-18. [Google Scholar] [CrossRef] [PubMed]
- Buonanno, M.; Ponnaiya, B.; Welch, D.; Stanislauskas, M.; Randers-Pehrson, G.; Smilenov, L.; Lowy, F.D.; Owens, D.M.; Brenner, D.J. Germicidal Efficacy and Mammalian Skin Safety of 222-Nm UV Light. Radiat. Res. 2017, 187, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, H.; Nomura, T.; Nazmul, T.; Omori, K.; Shigemoto, N.; Sakaguchi, T.; Ohge, H. Effectiveness of 222-Nm Ultraviolet Light on Disinfecting SARS-CoV-2 Surface Contamination. Am. J. Infect. Control 2020, 49, 299–301. [Google Scholar] [CrossRef] [PubMed]
- Woods, J.A.; Evans, A.; Forbes, P.D.; Coates, P.J.; Gardner, J.; Valentine, R.M.; Ibbotson, S.H.; Ferguson, J.; Fricker, C.; Moseley, H. The Effect of 222-Nm UVC Phototesting on Healthy Volunteer Skin: A Pilot Study. Photodermatol. Photoimmunol. Photomed. 2015, 31, 159–166. [Google Scholar] [CrossRef]
- Zhong, J.L.; Yang, L.; Lü, F.; Xiao, H.; Xu, R.; Wang, L.; Zhu, F.; Zhang, Y. UVA, UVB and UVC Induce Differential Response Signaling Pathways Converged on the EIF2α Phosphorylation. Photochem. Photobiol. 2011, 87, 1092–1104. [Google Scholar] [CrossRef] [PubMed]
- Rozpedek, W.; Pytel, D.; Mucha, B.; Leszczynska, H.; Diehl, J.A.; Majsterek, I. The Role of the PERK/EIF2α/ATF4/CHOP Signaling Pathway in Tumor Progression During Endoplasmic Reticulum Stress. Curr. Mol. Med. 2016, 16, 533–544. [Google Scholar] [CrossRef]
- Dhanasekaran, D.N.; Reddy, E.P. JNK Signaling in Apoptosis. Oncogene 2008, 27, 6245–6251. [Google Scholar] [CrossRef]
- Mah, L.J.; El-Osta, A.; Karagiannis, T.C. Γh2AX: A Sensitive Molecular Marker of DNA Damage and Repair. Leukemia 2010, 24, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Welch, D.; Spotnitz, H.M.; Brenner, D.J.; Randers-Pehrson, G.; Buonanno, M.; Shuryak, I. Far-UVC Light Applications: Sterilization of MRSA on a Surface and Inactivation of Aerosolized Influenza Virus. In Light-Based Diagnosis and Treatment of Infectious Diseases; Dai, T., Ed.; SPIE: Bellingham, WA, USA, 2018; Volume 10479, p. 60. [Google Scholar]
- Buonanno, M.; Welch, D.; Shuryak, I.; Brenner, D.J. Far-UVC Light (222 Nm) Efficiently and Safely Inactivates Airborne Human Coronaviruses. Sci. Rep. 2020, 10, 10285. [Google Scholar] [CrossRef]
- Goh, J.C.; Fisher, D.; Hing, E.C.H.; Hanjing, L.; Lin, Y.Y.; Lim, J.; Chen, O.W.; Chye, L.T. Disinfection Capabilities of a 222nm Wavelength Ultraviolet Lighting Device: A Pilot Study. J. Wound Care 2021, 30, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Fukui, T.; Niikura, T.; Oda, T.; Kumabe, Y.; Ohashi, H.; Sasaki, M.; Igarashi, T.; Kunisada, M.; Yamano, N.; Oe, K.; et al. Exploratory Clinical Trial on the Safety and Bactericidal Effect of 222-Nm Ultraviolet C Irradiation in Healthy Humans. PLoS ONE 2020, 15, e0235948. [Google Scholar] [CrossRef] [PubMed]
- Ong, Q.; Ronnie Teo, J.W.; dela Cruz, J.; Wee, E.; Wee, W. Irradiation of UVC LED at 277 Nm Inactivates Coronaviruses by Photodegradation of Spike Protein. 1 2. Biorxiv 2021. [Google Scholar] [CrossRef]
- Vichai, V.; Kirtikara, K. Sulforhodamine B Colorimetric Assay for Cytotoxicity Screening. Nat. Protoc. 2006, 1, 1112–1116. [Google Scholar] [CrossRef]
- FASTQC. A Quality Control Tool for High Throughput Sequence Data | BibSonomy. Available online: https://www.bibsonomy.org/bibtex/f230a919c34360709aa298734d63dca3 (accessed on 3 June 2021).





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ong, Q.; Wee, W.; Dela Cruz, J.; Teo, J.W.R.; Han, W. 222-Nanometer Far-UVC Exposure Results in DNA Damage and Transcriptional Changes to Mammalian Cells. Int. J. Mol. Sci. 2022, 23, 9112. https://doi.org/10.3390/ijms23169112
Ong Q, Wee W, Dela Cruz J, Teo JWR, Han W. 222-Nanometer Far-UVC Exposure Results in DNA Damage and Transcriptional Changes to Mammalian Cells. International Journal of Molecular Sciences. 2022; 23(16):9112. https://doi.org/10.3390/ijms23169112
Chicago/Turabian StyleOng, Qunxiang, Winson Wee, Joshua Dela Cruz, Jin Wah Ronnie Teo, and Weiping Han. 2022. "222-Nanometer Far-UVC Exposure Results in DNA Damage and Transcriptional Changes to Mammalian Cells" International Journal of Molecular Sciences 23, no. 16: 9112. https://doi.org/10.3390/ijms23169112
APA StyleOng, Q., Wee, W., Dela Cruz, J., Teo, J. W. R., & Han, W. (2022). 222-Nanometer Far-UVC Exposure Results in DNA Damage and Transcriptional Changes to Mammalian Cells. International Journal of Molecular Sciences, 23(16), 9112. https://doi.org/10.3390/ijms23169112