
Genetic Diversity of Tomato Black Ring Virus Satellite RNAs and Their Impact on Virus Replication

 , , , ,
, , , ,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Analysis of Genetic Diversity of satRNAs
2.2. Recombination and Selective Preasure Analysis of satRNAs
2.3. Phylogenetic Analysis of satRNAs
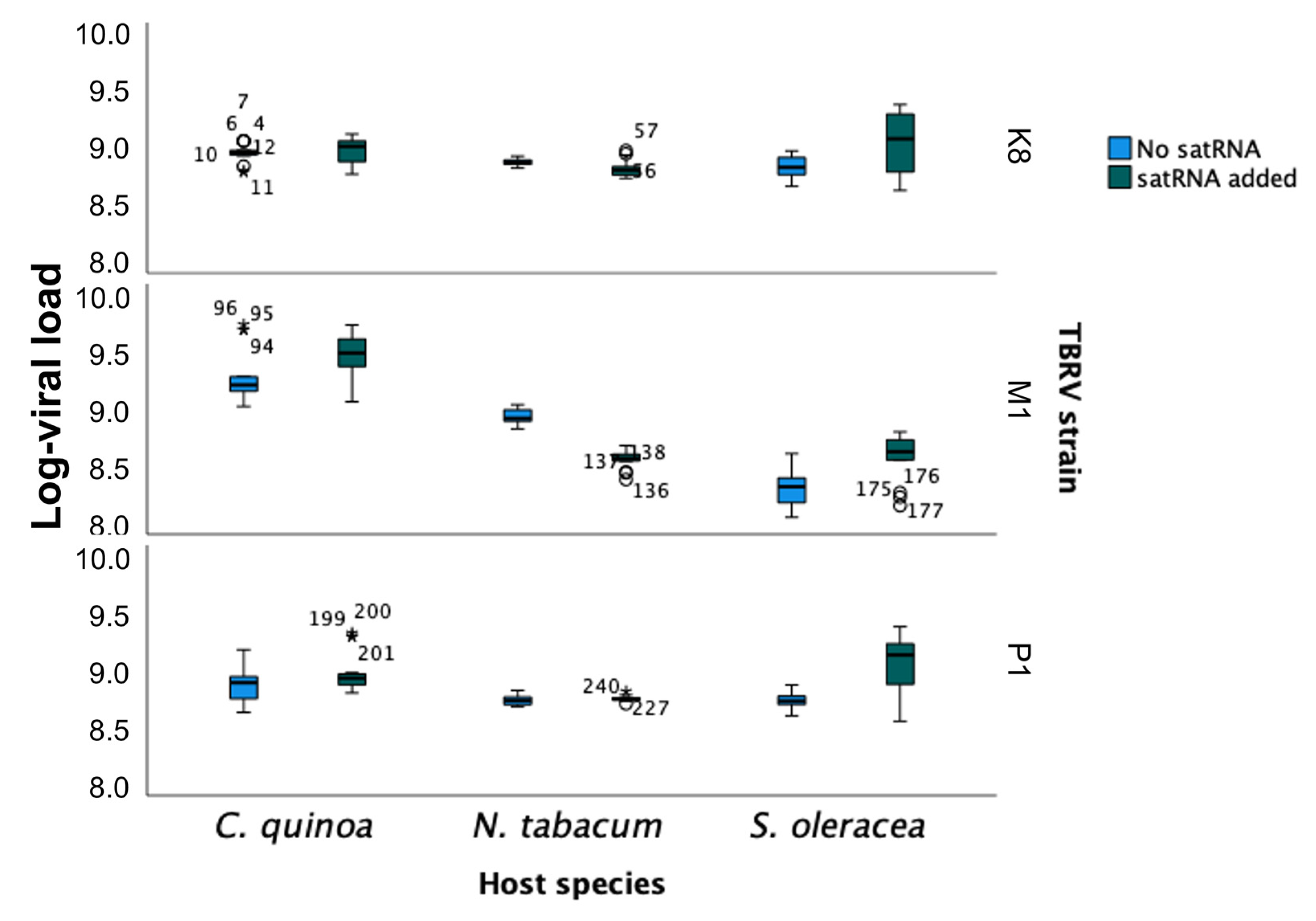
2.4. Analysis of the Absolute Quantification of TBRV Genomic RNA in Presence and Absence of satRNAs
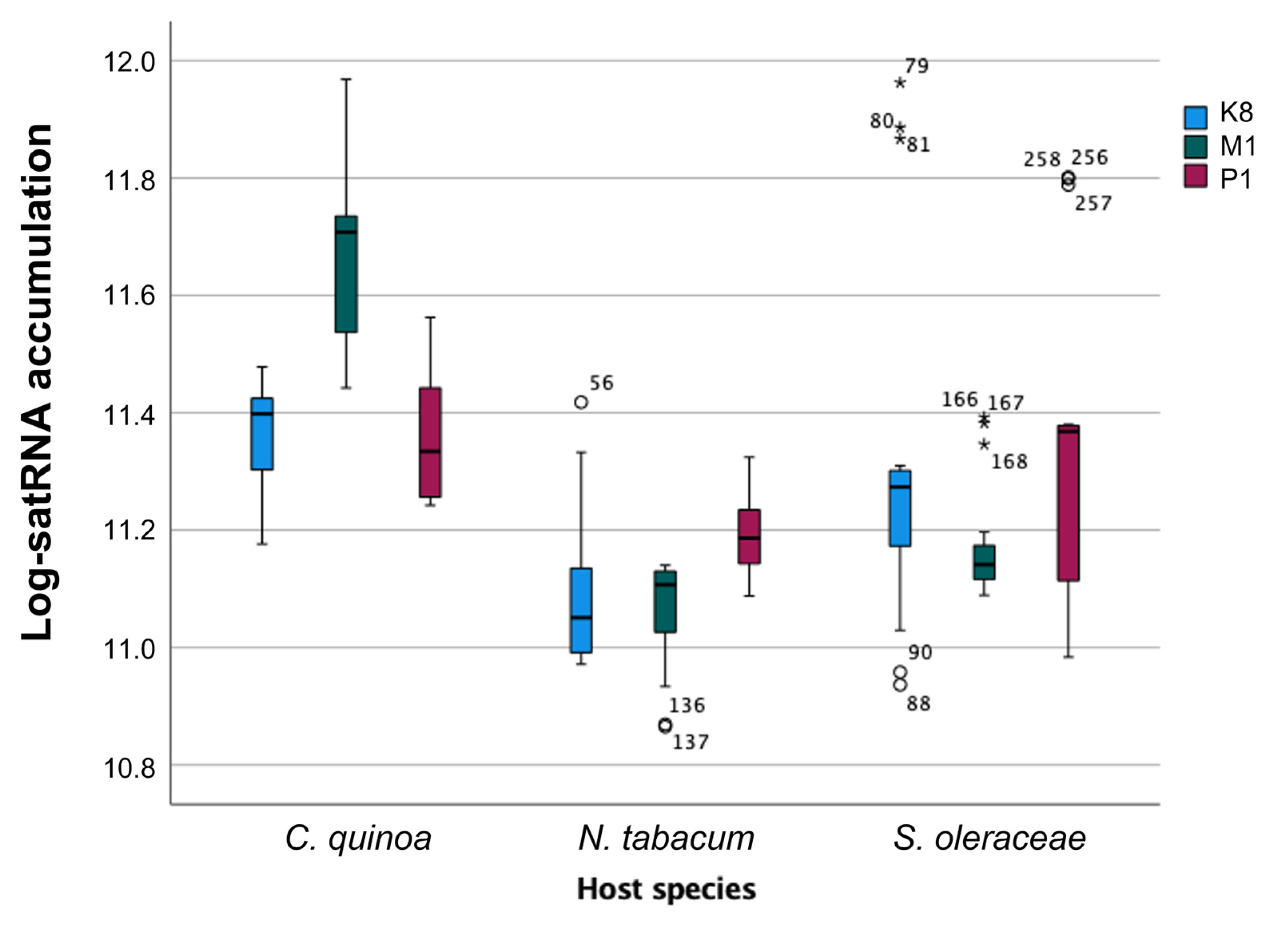
2.5. Analysis of the Absolute Quantification of satRNAs
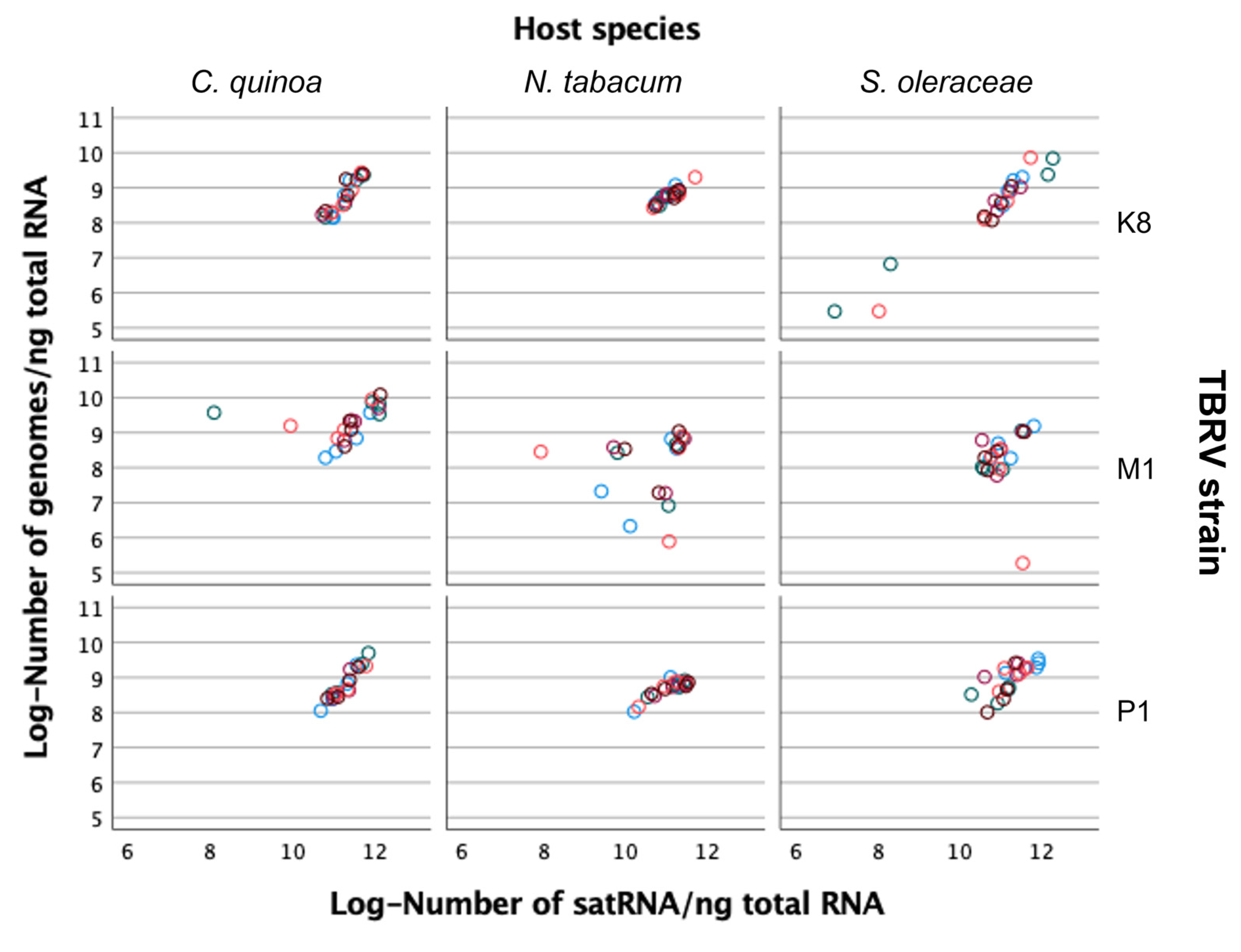
2.6. Association between Viral RNA and satRNA Accumulations
3. Discussion
4. Materials and Methods
4.1. Analysis of the Genetic Diversity of satRNAs Population
4.1.1. Virus Isolates
4.1.2. Amplification, Cloning and Sequencing of Satellite RNAs
4.1.3. Recombination and Selective Pressure Analysis
4.1.4. Phylogenetic Analysis of satRNAs
4.2. Obtaining an Infectious Clone of satRNA
4.3. Accumulation Experiments
4.3.1. Absolute Quantification of gRNA by RT-qPCR
4.3.2. Absolute Quantification of satRNAs by RT-qPCR
4.3.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Simon, A.E.; Roossinck, M.J.; Havelda, Z. Plant virus satellite and defective interfering RNAs: New paradigms for a new century. Annu. Rev. Phytopathol. 2004, 42, 415–437. [Google Scholar] [CrossRef] [PubMed]
- Pathak, K.B.; Nagy, P.D. Defective Interfering RNAs: Foes of Viruses and Friends of Virologists. Viruses 2009, 1, 895–919. [Google Scholar] [CrossRef] [PubMed]
- Graves, M.V.; Pogany, J.; Romero, J. Defective interfering RNAs and defective viruses associated with multipartite RNA viruses of plants. Sem. Virol. 1996, 7, 399–408. [Google Scholar] [CrossRef]
- Hasiów-Jaroszewska, B.; Minicka, J.; Zarzyńska-Nowak, A.; Budzyńska, D.; Elena, S.F. Defective RNA particles derived from Tomato black ring virus genome interfere with replication of parental virus. Virus Res. 2018, 250, 87–94. [Google Scholar] [CrossRef]
- Obrępalska-Stęplowska, A.; Wieczorek, P.; Budziszewska, M.; Jeszke, A.; Renaut, J. How can plant virus satellite RNAs alter the effects of plant virus infection? A study of the changes in the Nicotiana benthamiana proteome after infection by Peanut stunt virus in the presence or absence of its satellite RNA. Proteomics 2013, 13, 2162–2175. [Google Scholar] [CrossRef]
- Hu, C.C.; Yau-Heiu Hsu, Y.-H.; Linet, N.-S. Satellite RNAs and Satellite Viruses of Plants. Viruses 2009, 1, 1325–1350. [Google Scholar] [CrossRef]
- Fuchs, M.; Schmitt-Keichinger, C.; Sanfaçon, H. A Renaissance in Nepovirus Research Provides New Insights Into Their Molecular Interface With Hosts and Vectors. Adv. Virus Res. 2017, 97, 61–105. [Google Scholar]
- Brunt, A.A.; Crabtree, K.; Dallwitz, M.J.; Gibba, A.J.; Watson, L. Viruses of Plants. In Descriptions and Lists from the VIDE Database; CAB International: Wallingford, UK, 1966. [Google Scholar]
- Sneideris, D.; Staniulis, J. Phylogenetic analysis of Lithuanian tomato black ring virus isolates. Zemdirb.-Agric. 2014, 101, 193–198. [Google Scholar] [CrossRef]
- Rymelska, N.; Borodynko, N.; Pospieszny, H.; Hasiów-Jaroszewska, B. Analysis of the biological and molecular variability of the Polish isolates of Tomato black ring virus (TBRV). Virus Genes 2013, 47, 338–346. [Google Scholar] [CrossRef]
- Budzyńska, D.; Minicka, J.; Hasiów-Jaroszewska, B.; Elena, S.F. Molecular evolution of tomato black ring virus and de novo generation of a new type of defective RNAs during long-term passaging in different hosts. Plant Pathol. 2021, 69, 1767–1776. [Google Scholar] [CrossRef]
- Digiaro, M.; Yahyaoui, E.; Martelli, G.P.; Elbeaino, T. The sequencing of the complete genome of a Tomato black ring virus (TBRV) and of the RNA2 of three Grapevine chrome mosaic virus (GCMV) isolates from grapevine reveals the possible recombinant origin of GCMV. Virus Genes 2015, 50, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Oncino, C.; Hemmer, O.; Fritsch, O. Specificity in the association of Tomato black ring virus satellite RNA with helper virus. Virology 1995, 213, 87–96. [Google Scholar] [CrossRef]
- Jończyk, M.; Le Gall, O.L.; Pałucha, A.; Borodynko, N.; Pospieszny, H. Cloning and sequencing of full-length cDNAs of RNA1 and RNA2 of a Tomato black ring virus isolate from Poland. Arch. Virol. 2004, 149, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Zarzyńska-Nowak, A.; Hasiów-Jaroszewska, B.; Budzyńska, D.; Trzmiel, K. Genetic variability of Polish tomato black ring virus isolates and their satellite RNAs. Plant Pathol. 2020, 69, 1034–1041. [Google Scholar] [CrossRef]
- Fritsch, C.; Mayo, M.A.; Hemmer, O. Properties of the satellite RNA of nepoviruses. Biochimie 1993, 75, 561–567. [Google Scholar] [CrossRef]
- Roossinck, M.J.; Sleat, D.; Palukaitis, P. Satellite RNAs of plant viruses: Structures and biological effects. Microbiol. Rev. 1992, 56, 265–279. [Google Scholar] [CrossRef] [PubMed]
- Gottula, J.; Lapato, D.; Cantilina, K.; Saito, S.; Bartlett, B.; Fuchs, M. Genetic variability, evolution, and biological effects of Grapevine fanleaf virus satellite RNAs. Phytopathology 2013, 103, 1180–1187. [Google Scholar] [CrossRef]
- Fuchs, M.; Pinck, M.; Serghini, M.A.; Ravelonandro, M.; Walter, B.; Pinck, L. The Nucleotide Sequence of Satellite RNA in Grapevine Fanleaf Virus, Strain F13. J. Gen. Virol. 1989, 70, 955–962. [Google Scholar] [CrossRef]
- Cillo, F.; Finetti-Sialer, M.M.; Papanice, M.A.; Gallitelli, D. Analysis of mechanisms involved in the Cucumber mosaic virus satellite RNA-mediated transgenic resistance in tomato plants. Mol. Plant Microbe Interact. 2004, 17, 98–108. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evolut. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Hemmer, O.; Meyer, M.; Greif, C.; Fritsch, C. Comparison of the nucleotide sequences of five tomato black ring virus satellite RNAs. J. Gen. Virol. 1987, 68, 1823–1833. [Google Scholar] [CrossRef]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef] [PubMed]
- Kosakovsky Pond, S.L.; Posada, D.; Gravenor, M.B.; Woelk, C.H.; Frost, S.D.W. GARD: A genetic algorithm for recombination detection. Bioinformatics 2006, 22, 3096–3098. [Google Scholar] [CrossRef] [PubMed]
- Palukaitis, P.; Roossinck, M.J.; Dietzgen, R.G.; Francki, R.I.B. Cucumber mosaic virus. Adv. Virus Res. 1992, 41, 281–348. [Google Scholar]
- Briddon, R.W.; Ghabrial, S.; Lin, N.-S.; Palukaitis, P.; Scholthof, K.-B.G.; Vetten, H.-J. Satellite and other virus-dependent nucleic acids. In Virus Taxonomy. Classification and Nomenclatureof Viruses. Ninth Report of the International Committee on Taxonomyof Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier Academic Press: San Diego, CA, USA, 2012; pp. 1211–1219. [Google Scholar]
- Simon, A.E.; Engel, H.; Johnson, R.P.; Howell, S.H. Identification of regions affecting virulence, RNA processing and infectivity in the virulent satellite of turnip crinkle virus. EMBO J. 1988, 7, 2645–2651. [Google Scholar] [CrossRef]
- Pelczyk, M.; Obrępalska-Stęplowska, A.; Pospieszny, H. Subwirusowe cząsteczki RNA związane z wirusami roślinnymi typu ss(+)RNA. Postępy Biochem. 2006, 52, 212–221. [Google Scholar]
- Schierup, M.H.; Hein, J. Consequences of recombination on traditional phylogenetic analysis. Genetics 2000, 156, 879–891. [Google Scholar] [CrossRef]
- Sanjuán, R.; Domingo-Calap, P. Genetic Diversity and Evolution of Viral Populations. In Encyclopedia of Virology, 2nd ed.; Bamford, D., Zuckerman, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; Volume 1, pp. 53–61. [Google Scholar] [CrossRef]
- Xu, P.; Roossinck, M.J. Cucumber Mosaic Virus D Satellite RNA–Induced Programmed Cell Death in Tomato. Plant Cell 2000, 12, 1079–1092. [Google Scholar]
- García-Arenal, F.; Palukaitis, P. Structure and functional relationships of satellite RNAs of cucumber mosaic virus. Curr. Top Microbiol. Immunol. 1999, 239, 37–63. [Google Scholar]
- Betancourt, M.; Fraile, A.; Garcia-Arenal, F. Cucumber mosaic virus satellite RNAs that induce similar symptoms in melon plants show large differences in fitness. J. Gen. Virol. 2011, 92, 1930–1938. [Google Scholar] [CrossRef]
- Collmer, C.W.; Howell, S.H. Role of satellite RNA in the expression of symptoms caused by plant viruses. Annu. Rev. Phytopathol. 1992, 30, 419–442. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Blancaflor, E.B.; Roossinck, M.J. In spite of induced multiple defense responses, tomato plants infected with Cucumber mosaic virus and D satellite RNA succumb to systemic necrosis. Mol. Plant Microbe Interact. 2003, 16, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Lai, L.; Lin, R.; Jin, C.; Chen, J. Differential effects of Cucumber mosaic virus satellite RNAs in the perturbation of microRNA-regulated gene expression in tomato. Mol. Biol. Rep. 2012, 39, 775–784. [Google Scholar] [CrossRef] [PubMed]
- Shimura, H.; Pantaleo, V.; Ishihara, T.; Myojo, N.; Inaba, J.-I.; Sueda, K.; Burgyán, J.; Masuta, C. A viral satellite RNA induces yellow symptoms on tobacco by targeting a gene involved in chlorophyll biosynthesis using the RNA silencing machinery. PLoS Pathog. 2011, 7, e1002021. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Weaver, S.; Shank, S.D.; Spielman, S.J.; Li, M.; Muse, S.V.; Kosakovsky Pond, S.L. Datamonkey 2.0: A Modern Web Application for Characterizing Selective and Other Evolutionary Processes. Mol. Biol. Evol. 2018, 35, 773–777. [Google Scholar] [CrossRef]
- Subramanian, B.; Gao, S.; Lercher, M.J.; Hu, S.; Chen, W.-H. Evolview v3: A webserver for visualization, annotation, and management of phylogenetic trees. Nucleic Acids Res. 2019, 47, W270–W275. [Google Scholar] [CrossRef]
- Ferriol, I.; Turina, M.; Zamora-Macorra, E.J.; Falk, B.W. RNA1-Independent Replication and GFP Expression from Tomato marchitez virus Isolate M Cloned cDNA. Virology 2016, 106, 500–509. [Google Scholar] [CrossRef]
- Zarzyńska-Nowak, A.; Ferriol, I.; Falk, B.W.; Borodynko-Filas, N.; Hasiów-Jaroszewska, B. Construction of Agrobacterium tumefaciens-mediated tomato black ring virus infectious cDNA clones. Virus Res. 2017, 230, 59–62. [Google Scholar] [CrossRef]
- Hasiów-Jaroszewska, B.; Budzyńska, D.; Borodynko, N.; Pospieszny, H. Rapid detection of genetically diverse tomato black ring virus isolates using reverse transcription loop-mediated isothermal amplification. Arch. Virol. 2015, 160, 3075–3078. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| satRNA | Host | Location | Accession | Reference |
|---|---|---|---|---|
| sTBRV-Byd4 | Robinia pseudoacacia | Wielkopolska, Poland | ON953201 | this study |
| sTBRV-Byd7 | R. pseudoacacia | Wielkopolska, Poland | ON953202 | this study |
| sTBRV-Byd8 | R. pseudoacacia | Wielkopolska, Poland | ON953203 | this study |
| sTBRV-CH | Armoracia rusticana | Mazowieckie, Poland | ON953204 | this study |
| sTBRV-Goc2 | R. pseudoacacia | Kujawsko-pomorskie, Poland | ON953205 | this study |
| sTBRV-L1 | R. pseudoacacia | Mazowieckie, Poland | ON953206 | this study |
| sTBRV-Nar1 | R. pseudoacacia | Wielkopolska, Poland | ON953207 | this study |
| sTBRV-Nar6 | R. pseudoacacia | Wielkopolska, Poland | ON953208 | this study |
| sTBRV-NR3 | R. pseudoacacia | Wielkopolska, Poland | ON953209 | this study |
| sTBRV-O1 | Cucumis sativus | Wielkopolska, Poland | ON953210 | this study |
| sTBRV-P11 | R. pseudoacacia | Wielkopolska, Poland | ON953211 | this study |
| sTBRV-Sier1 | R. pseudoacacia | Wielkopolska, Poland | ON953212 | this study |
| sTBRV-Szp6 | R. pseudoacacia | Wielkopolska, Poland | ON953213 | this study |
| sTBRV-Wloc3 | R. pseudoacacia | Kujawsko-pomorskie, Poland | ON953214 | this study |
| sTBRV-Wloc4 | R. pseudoacacia | Kujawsko-pomorskie, Poland | ON953215 | this study |
| sTBRV-WM1 | R. pseudoacacia | Wielkopolska, Poland | ON953216 | this study |
| sTBRV-WM2 | R. pseudoacacia | Wielkopolska, Poland | ON953217 | this study |
| sTBRV-WM3 | R. pseudoacacia | Wielkopolska, Poland | ON953218 | this study |
| sTBRV-K8 | Cucurbita pepo | Wielkopolskie, Poland | MN699709 | [15] |
| sTBRV-M1 | R. pseudoacacia | Wielkopolska, Poland | MN699710 | [15] |
| sTBRV-W | Verbena officinalis | Mazowieckie, Poland | MN718462 | [15] |
| sTBRV-S1 | Lactuca sativa | Wielkopolska, Poland | MN699711 | [15] |
| sTBRV-AG | Tagetes patula | Wielkopolska, Poland | MN699708 | [15] |
| sTBRV-SM2 | R. pseudoacacia | Wielkopolska, Poland | MN699711 | [15] |
| sTBRV-E | L. sativa | Norfolk, UK | X05688.1 | [22] |
| sTBRV-C | Apium sp. | Suffolk, UK | X056 89.1 | [22] |
| FUBAR | FEL | MEME | SLAC | Codon Position | |||
|---|---|---|---|---|---|---|---|
| ω > 1 | ω < 1 | ω > 1 | ω < 1 | ω > 1 | ω > 1 | ω < 1 | 19, 52, 74, 99, 114, 125, 162, 169, 174, 208, 224, 227, 244, 245, 300, 313, 351 |
| 16 | 44 | 5 | 41 | 7 | - | 14 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Minicka, J.; Taberska, A.; Zarzyńska-Nowak, A.; Kubska, K.; Budzyńska, D.; Elena, S.F.; Hasiów-Jaroszewska, B. Genetic Diversity of Tomato Black Ring Virus Satellite RNAs and Their Impact on Virus Replication. Int. J. Mol. Sci. 2022, 23, 9393. https://doi.org/10.3390/ijms23169393
Minicka J, Taberska A, Zarzyńska-Nowak A, Kubska K, Budzyńska D, Elena SF, Hasiów-Jaroszewska B. Genetic Diversity of Tomato Black Ring Virus Satellite RNAs and Their Impact on Virus Replication. International Journal of Molecular Sciences. 2022; 23(16):9393. https://doi.org/10.3390/ijms23169393
Chicago/Turabian StyleMinicka, Julia, Agnieszka Taberska, Aleksandra Zarzyńska-Nowak, Katarzyna Kubska, Daria Budzyńska, Santiago F. Elena, and Beata Hasiów-Jaroszewska. 2022. "Genetic Diversity of Tomato Black Ring Virus Satellite RNAs and Their Impact on Virus Replication" International Journal of Molecular Sciences 23, no. 16: 9393. https://doi.org/10.3390/ijms23169393
APA StyleMinicka, J., Taberska, A., Zarzyńska-Nowak, A., Kubska, K., Budzyńska, D., Elena, S. F., & Hasiów-Jaroszewska, B. (2022). Genetic Diversity of Tomato Black Ring Virus Satellite RNAs and Their Impact on Virus Replication. International Journal of Molecular Sciences, 23(16), 9393. https://doi.org/10.3390/ijms23169393