

Immobilization of Wnt Fragment Peptides on Magnetic Nanoparticles or Synthetic Surfaces Regulate Wnt Signaling Kinetics
 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
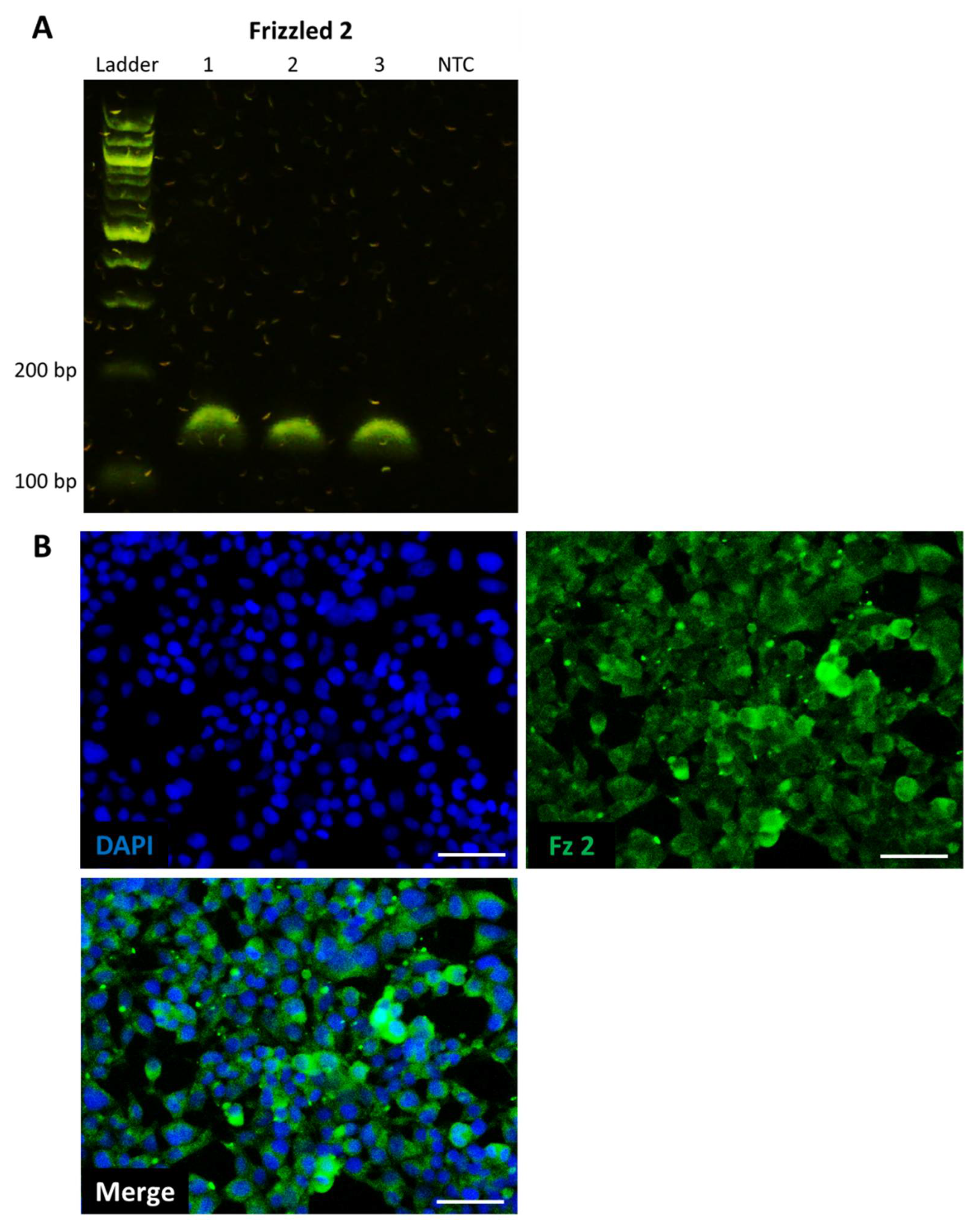
2.1. HEK293 Reporter Cell Line Expresses Target Frizzled Receptor
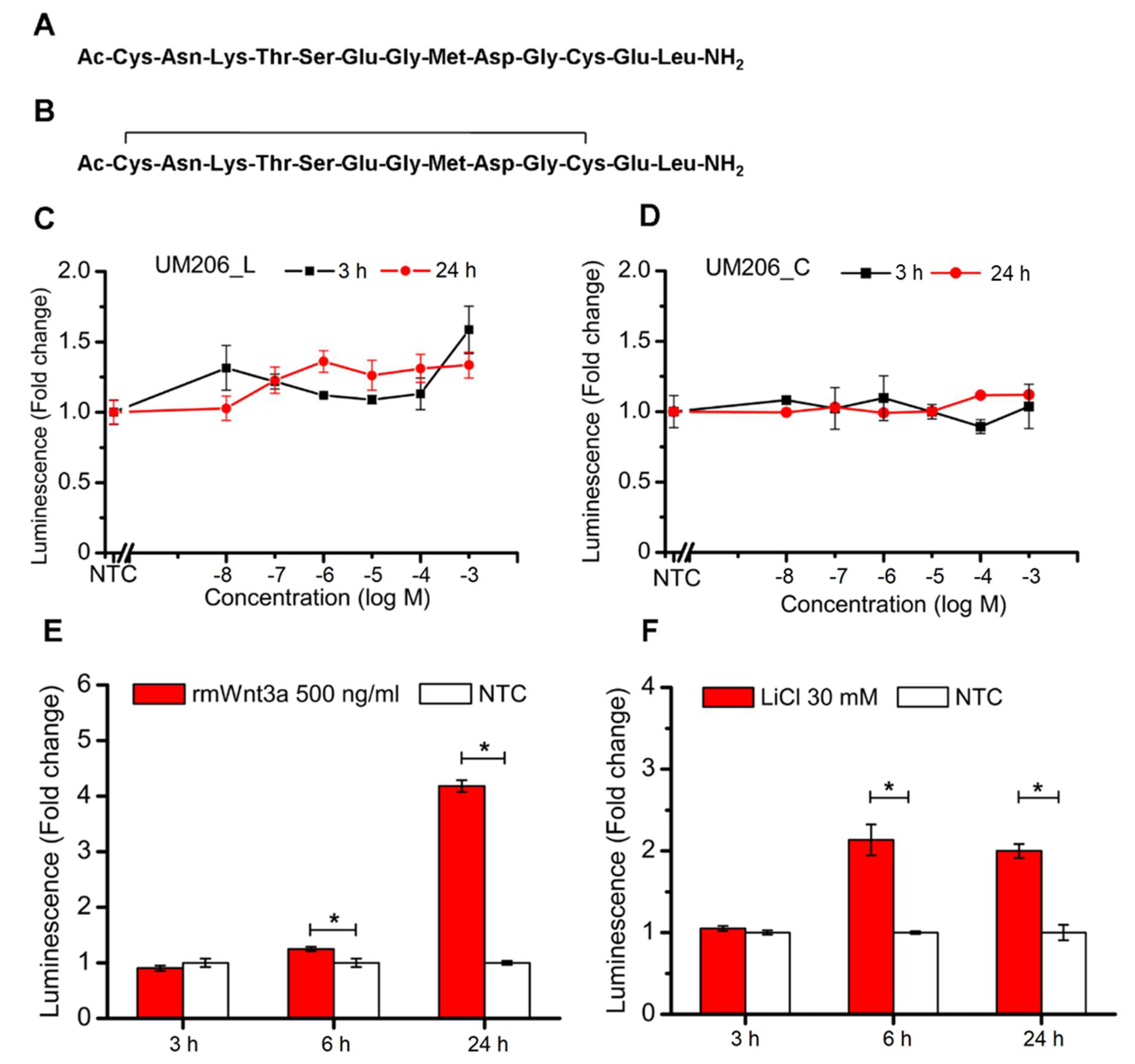
2.2. Soluble Peptides Have No Overall Effect on TCF/LEF Signal Transduction in Reporter Cell Line
2.3. Characterization of Magnetic Nanoparticle (MNP)
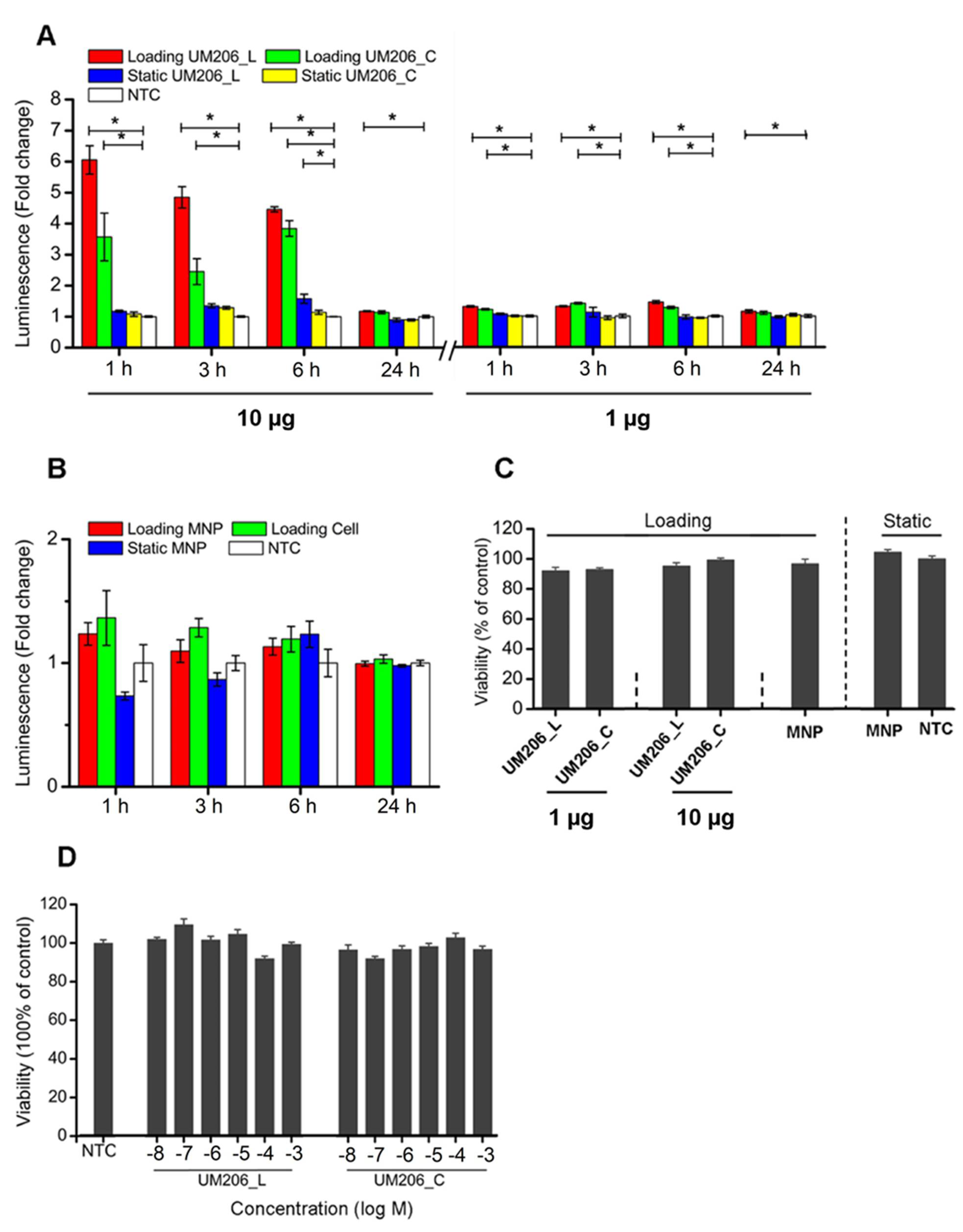
2.4. Nanomechanical Activation of Functionalized MNPs Stimulates β-Catenin Mediated TCF/LEF Signal Transduction
2.5. Magnetic Stimulation Has No Influence on Cell Viability
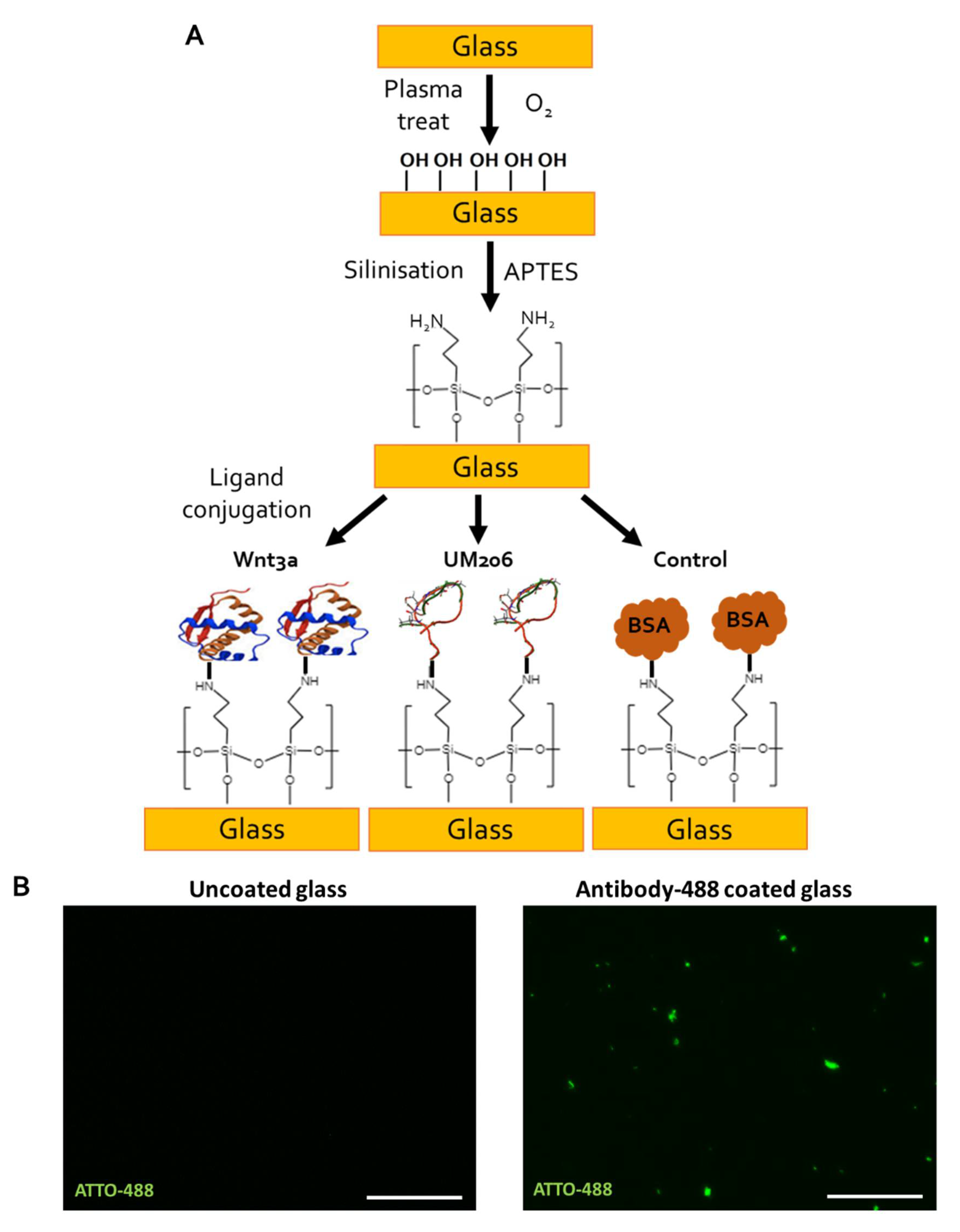
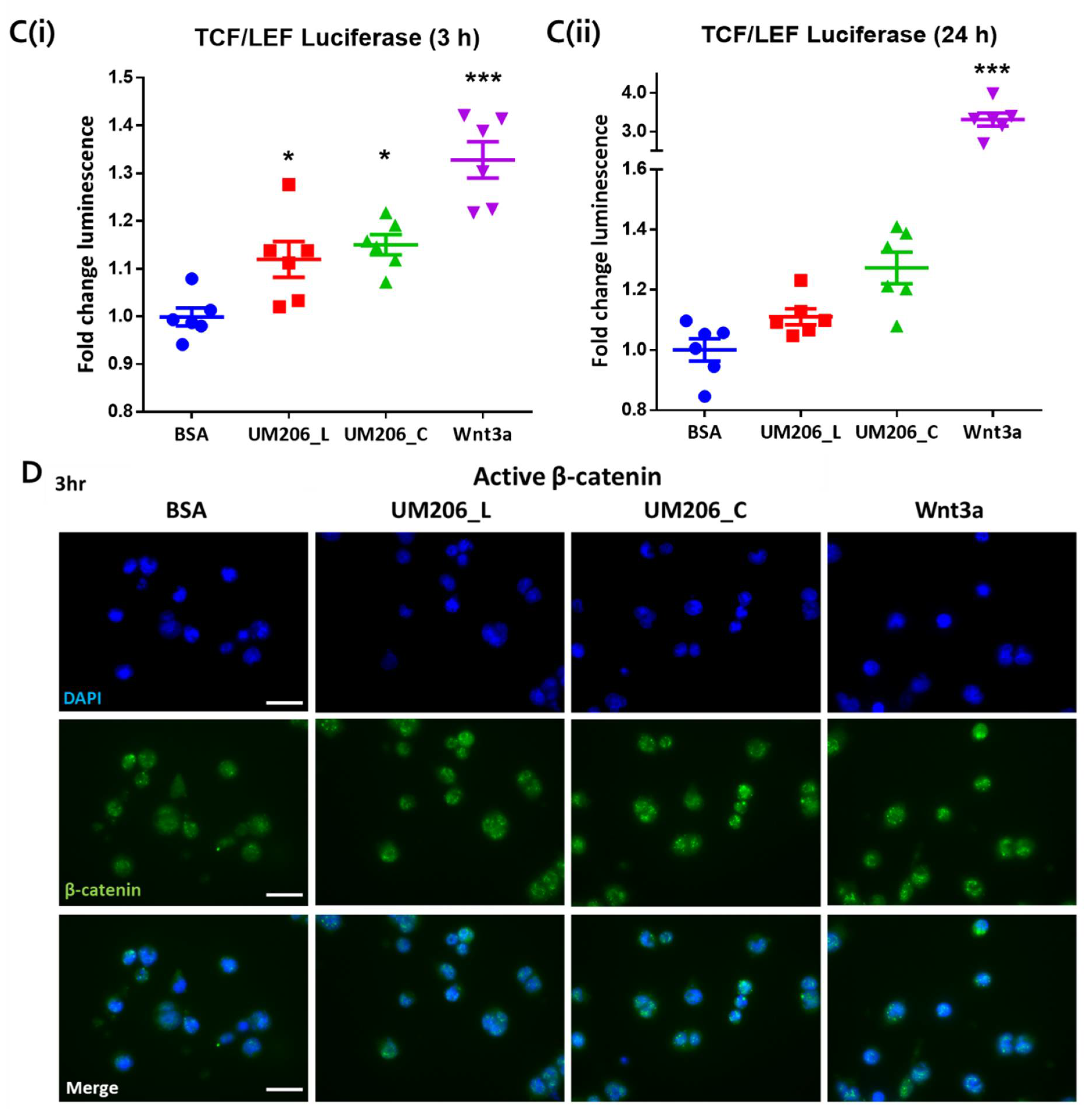
2.6. Immobilised UM206 Retains Activity on Glass Substrate
3. Discussion
4. Materials and Methods
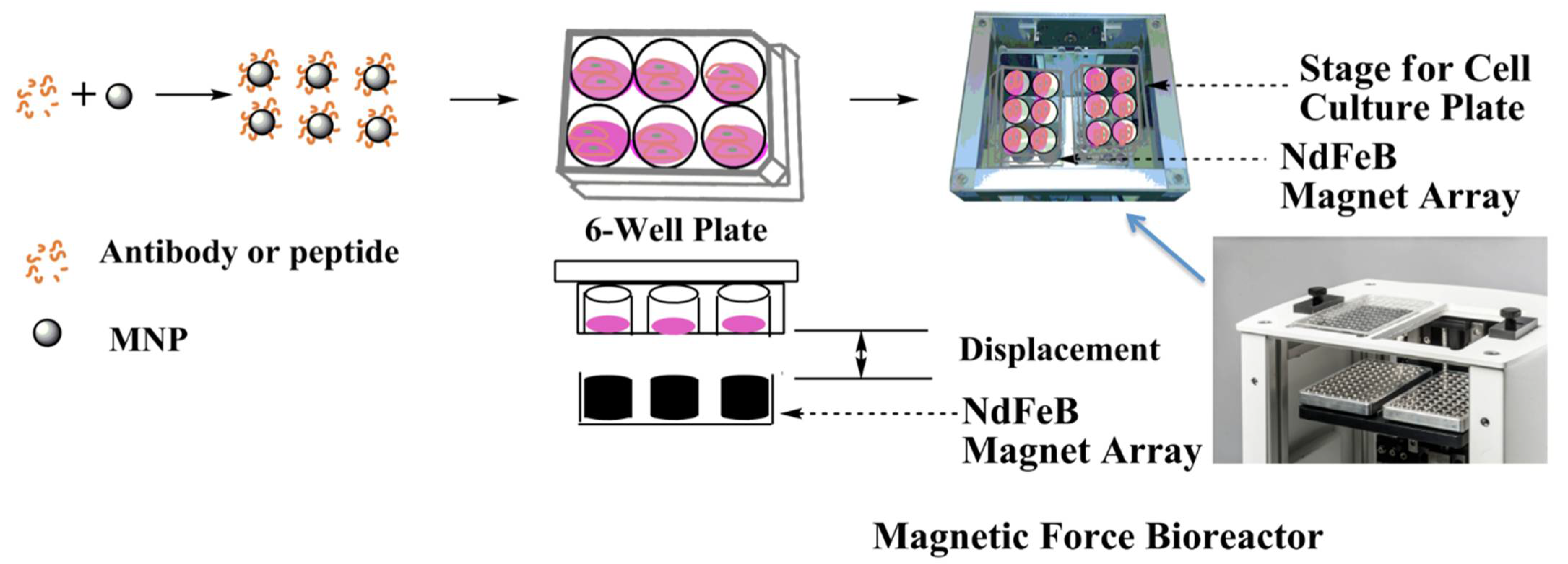
4.1. Magnetic Force Bioreactor
4.2. Construction of a Stably Transfected Reporter Cell Line
4.3. Polymerase Chain Reaction
4.4. Immunocytochemistry
4.5. Effect of Synthetic Peptides on TCF/LEF Signaling Transduction
4.6. Preparation of Peptide-Conjugated Magnetic Nanoparticles
4.7. Characterisation of Magnetic Nanoparticles
4.7.1. Zetasizing and Zeta Potential
4.7.2. Total Protein Assay
4.7.3. Transmission Electron Microscopy
4.7.4. Fourier-Transform Infrared Spectroscopy (FTIR)
4.8. Magneto-Mechanical Stimulation of HEK293 Luc-LEF/TCF Cells
4.9. Cell Viability
4.10. Functionalization of Glass Coverslips
4.11. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clevers, H.; Nusse, R. Wnt/β-catenin signaling and disease. Cell 2012, 149, 1192–1205. [Google Scholar] [CrossRef] [PubMed]
- Li, V.S.; Ng, S.S.; Boersema, P.J.; Low, T.Y.; Karthaus, W.R.; Gerlach, J.; Mohammed, S.; Heck, A.J.R.; Maurice, M.M.; Mahmoudi, T.; et al. Wnt signaling through inhibition of β-catenin degradation in an intact Axin1 complex. Cell 2012, 149, 1245–1256. [Google Scholar] [CrossRef]
- Dann, C.E.; Hsieh, J.-C.; Rattner, A.; Sharma, D.; Nathans, J.; Leahy, D.J. Insights into Wnt binding and signalling from the structures of two Frizzled cysteine-rich domains. Nature 2001, 412, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Dai, X. Winning WNT: Race to Wnt signaling inhibitors. Proc. Natl. Acad. Sci. USA 2011, 108, 5929–5930. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, B.D.; Grashoff, C.; Schwartz, M.A. Dynamic molecular processes mediate cellular mechanotransduction. Nature 2011, 475, 316–323. [Google Scholar] [CrossRef]
- Janmey, P.A.; Miller, R.T. Mechanisms of mechanical signaling in development and disease. J. Cell Sci. 2011, 124, 9–18. [Google Scholar] [CrossRef]
- Heisenberg, C.P.; Bellaïche, Y. Forces in tissue morphogenesis and patterning. Cell 2013, 153, 948–962. [Google Scholar] [CrossRef]
- Lin, C.; Jiang, X.; Dai, Z.; Guo, X.; Weng, T.; Wang, J.; Li, Y.; Feng, G.; Gao, X.; He, L. Sclerostin mediates bone response to mechanical unloading through antagonizing Wnt/beta-catenin signaling. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2009, 24, 1651–1661. [Google Scholar] [CrossRef]
- Robinson, J.A.; Chatterjee-Kishore, M.; Yaworsky, P.J.; Cullen, D.M.; Zhao, W.; Li, C.; Kharode, Y.; Sauter, L.; Babij, P.; Brown, E.L.; et al. Wnt/beta-catenin signaling is a normal physiological response to mechanical loading in bone. J. Biol. Chem. 2006, 281, 31720–31728. [Google Scholar] [CrossRef]
- Sawakami, K.; Robling, A.G.; Ai, M.; Pitner, N.D.; Liu, D.; Warden, S.J.; Li, J.; Maye, P.; Rowe, D.W.; Duncan, R.L.; et al. The Wnt co-receptor LRP5 is essential for skeletal mechanotransduction but not for the anabolic bone response to parathyroid hormone treatment. J. Biol. Chem. 2006, 281, 23698–23711. [Google Scholar] [CrossRef] [Green Version]
- del Rio, A.; Perez-Jimenez, R.; Liu, R.; Roca-Cusachs, P.; Fernandez, J.M.; Sheetz, M.P. Stretching single talin rod molecules activates vinculin binding. Science 2009, 323, 638–641. [Google Scholar] [CrossRef] [PubMed]
- Nikukar, H.; Reid, S.; Tsimbouri, P.M.; Riehle, M.O.; Curtis, A.S.G.; Dalby, M.J. Osteogenesis of Mesenchymal Stem Cells by Nanoscale Mechanotransduction. ACS Nano 2013, 7, 2758–2767. [Google Scholar] [CrossRef] [PubMed]
- Dobson, J.; Cartmell, S.H.; Keramane, A.; El Haj, A.J. Principles and design of a novel magnetic force mechanical conditioning bioreactor for tissue engineering, stem cell conditioning, and dynamic in vitro screening. IEEE Trans. Nanobiosci. 2006, 5, 173–177. [Google Scholar] [CrossRef]
- Dobson, J. Remote control of cellular behaviour with magnetic nanoparticles. Nat. Nanotechnol. 2008, 3, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Mannix, R.J.; Kumar, S.; Cassiola, F.; Montoya-Zavala, M.; Feinstein, E.; Prentiss, M.; Ingber, D.E. Nanomagnetic actuation of receptor-mediated signal transduction. Nat. Nanotechnol. 2008, 3, 36–40. [Google Scholar] [CrossRef]
- Gonçalves, A.I.; Rotherham, M.; Markides, H.; Rodrigues, M.T.; Reis, R.L.; Gomes, M.E.; El Haj, A.J. Triggering the activation of Activin A type II receptor in human adipose stem cells towards tenogenic commitment using mechanomagnetic stimulation. Nanomed. Nanotechnol. Biol. Med. 2018, 14, 1149–1159. [Google Scholar] [CrossRef]
- Henstock, J.R.; Rotherham, M.; El Haj, A.J. Magnetic ion channel activation of TREK1 in human mesenchymal stem cells using nanoparticles promotes osteogenesis in surrounding cells. J. Tissue Eng. 2018, 9, 2041731418808695. [Google Scholar] [CrossRef]
- Hu, B.; El Haj, A.J.; Dobson, J. Receptor-targeted, magneto-mechanical stimulation of osteogenic differentiation of human bone marrow-derived mesenchymal stem cells. Int. J. Mol. Sci. 2013, 14, 19276–19293. [Google Scholar] [CrossRef] [PubMed]
- Rotherham, M.; Nahar, T.; Broomhall, T.J.; Telling, N.D.; El Haj, A.J. Remote magnetic actuation of cell signalling for tissue engineering. Curr. Opin. Biomed. Eng. 2022, 100410. [Google Scholar] [CrossRef]
- Mills, K.M.; Szczerkowski, J.L.A.; Habib, S.J. Wnt ligand presentation and reception: From the stem cell niche to tissue engineering. Open Biol. 2017, 7, 170140. [Google Scholar] [CrossRef] [Green Version]
- Walker, M.; Will, I.; Pratt, A.; Chechik, V.; Genever, P.; Ungar, D. Magnetically Triggered Release of Entrapped Bioactive Proteins from Thermally Responsive Polymer-Coated Iron Oxide Nanoparticles for Stem-Cell Proliferation. ACS Appl. Nano Mater. 2020, 3, 5008–5013. [Google Scholar] [CrossRef] [PubMed]
- Habib, S.J.; Chen, B.C.; Tsai, F.C.; Anastassiadis, K.; Meyer, T.; Betzig, E.; Nusse, R. A localized Wnt signal orients asymmetric stem cell division in vitro. Science 2013, 339, 1445–1448. [Google Scholar] [CrossRef]
- Lowndes, M.; Rotherham, M.; Price, J.C.; El Haj, A.J.; Habib, S.J. Immobilized WNT Proteins Act as a Stem Cell Niche for Tissue Engineering. Stem Cell Rep. 2016, 7, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Lowndes, M.; Junyent, S.; Habib, S.J. Constructing cellular niche properties by localized presentation of Wnt proteins on synthetic surfaces. Nat. Protoc. 2017, 12, 1498–1512. [Google Scholar] [CrossRef] [PubMed]
- Okuchi, Y.; Reeves, J.; Ng, S.S.; Doro, D.H.; Junyent, S.; Liu, K.J.; El Haj, A.J.; Habib, S.J. Wnt-modified materials mediate asymmetric stem cell division to direct human osteogenic tissue formation for bone repair. Nat. Mater. 2021, 20, 108–118. [Google Scholar] [CrossRef]
- Blankesteijn, W.M.; Laeremans, H.; Hackeng, T.M. Antagonistic Peptides for Frizzled-1 and Frizzled-2. U.S. Patent No. 8,598,122, 18 February 2010. [Google Scholar]
- Rotherham, M.; Henstock, J.R.; Qutachi, O.; El Haj, A.J. Remote regulation of magnetic particle targeted Wnt signaling for bone tissue engineering. Nanomed. Nanotechnol. Biol. Med. 2018, 14, 173–184. [Google Scholar]
- Rotherham, M.; Nahar, T.; Goodman, T.; Telling, N.; Gates, M.; El Haj, A. Magnetic Mechanoactivation of Wnt Signaling Augments Dopaminergic Differentiation of Neuronal Cells. Adv. Biosyst. 2019, 3, 1900091. [Google Scholar] [CrossRef]
- Rotherham, M.; El Haj, A.J. Remote Activation of the Wnt/beta-Catenin Signalling Pathway Using Functionalised Magnetic Particles. PLoS ONE 2015, 10, e0121761. [Google Scholar] [CrossRef]
- Markides, H.; McLaren, J.S.; Telling, N.D.; Alom, N.; Al-Mutheffer, E.a.A.; Oreffo, R.O.C.; Zannettino, A.; Scammell, B.E.; White, L.J.; El Haj, A.J. Translation of remote control regenerative technologies for bone repair. NPJ Regen. Med. 2018, 3, 9. [Google Scholar] [CrossRef]
- Tao, Y.; Mis, M.; Blazer, L.; Ustav, M.J.; Steinhart, Z.; Chidiac, R.; Kubarakos, E.; O’Brien, S.; Wang, X.; Jarvik, N.; et al. Tailored tetravalent antibodies potently and specifically activate Wnt/Frizzled pathways in cells, organoids and mice. Elife 2019, 8, e46134. [Google Scholar]
- Kilinc, D.; Dennis, C.L.; Lee, G.U. Bio-Nano-Magnetic Materials for Localized Mechanochemical Stimulation of Cell Growth and Death. Adv. Mater. 2016, 28, 5672–5680. [Google Scholar] [CrossRef]
- Na, S.; Collin, O.; Chowdhury, F.; Tay, B.; Ouyang, M.; Wang, Y.; Wang, N. Rapid signal transduction in living cells is a unique feature of mechanotransduction. Proc. Natl. Acad. Sci. USA 2008, 105, 6626–6631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, E.A.; Calderwood, D.A. Forces and Bond Dynamics in Cell Adhesion. Science 2007, 316, 1148–1153. [Google Scholar] [CrossRef] [PubMed]
- Schihada, H.; Kowalski-Jahn, M.; Turku, A.; Schulte, G. Deco nvolution of WNT-induced Frizzled conformational dynamics with fluorescent biosensors. Biosens. Bioelectron. 2021, 177, 112948. [Google Scholar] [CrossRef] [PubMed]
- Apodaca, G. Modulation of membrane traffic by mechanical stimuli. Am. J. Physiol. Ren. Physiol. 2002, 282, F179–F190. [Google Scholar] [CrossRef]
- Hu, B.; Dobson, J.; El Haj, A.J. Control of smooth muscle alpha-actin (SMA) up-regulation in HBMSCs using remote magnetic particle mechano-activation. Nanomedicine 2014, 10, 45–55. [Google Scholar] [CrossRef]
- Ring, L.; Perobner, I.; Karow, M.; Jochum, M.; Neth, P.; Faussner, A. Reporter gene HEK 293 cells and WNT/Frizzled fusion proteins as tools to study WNT signaling pathways. Biol. Chem. 2011, 392, 1011–1020. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Size (nm) | Zeta (mV) | ||
|---|---|---|---|
| Control | MNP-Uncoated | 314.7 ± 3.3 | −30.8 ± 0.4 |
| 10 μg peptide per 1 mg particle | MNP-UM206_L | 319.0 ± 0.7 | −16.8 ± 0.7 |
| MNP-UM206_C | 329.3 ± 4.7 | −14.9 ± 0.2 | |
| 1 μg peptide per 1 mg particle | MNP-UM206_L | 323.0 ± 2.1 | −15.2 ± 0.3 |
| MNP-UM206_C | 317.9 ± 2.9 | −14.2 ± 0.1 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, B.; Rotherham, M.; Farrow, N.; Roach, P.; Dobson, J.; El Haj, A.J. Immobilization of Wnt Fragment Peptides on Magnetic Nanoparticles or Synthetic Surfaces Regulate Wnt Signaling Kinetics. Int. J. Mol. Sci. 2022, 23, 10164. https://doi.org/10.3390/ijms231710164
Hu B, Rotherham M, Farrow N, Roach P, Dobson J, El Haj AJ. Immobilization of Wnt Fragment Peptides on Magnetic Nanoparticles or Synthetic Surfaces Regulate Wnt Signaling Kinetics. International Journal of Molecular Sciences. 2022; 23(17):10164. https://doi.org/10.3390/ijms231710164
Chicago/Turabian StyleHu, Bin, Michael Rotherham, Neil Farrow, Paul Roach, Jon Dobson, and Alicia J. El Haj. 2022. "Immobilization of Wnt Fragment Peptides on Magnetic Nanoparticles or Synthetic Surfaces Regulate Wnt Signaling Kinetics" International Journal of Molecular Sciences 23, no. 17: 10164. https://doi.org/10.3390/ijms231710164
APA StyleHu, B., Rotherham, M., Farrow, N., Roach, P., Dobson, J., & El Haj, A. J. (2022). Immobilization of Wnt Fragment Peptides on Magnetic Nanoparticles or Synthetic Surfaces Regulate Wnt Signaling Kinetics. International Journal of Molecular Sciences, 23(17), 10164. https://doi.org/10.3390/ijms231710164