
Morphological and Molecular Responses of Lateolabrax maculatus Skeletal Muscle Cells to Different Temperatures

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
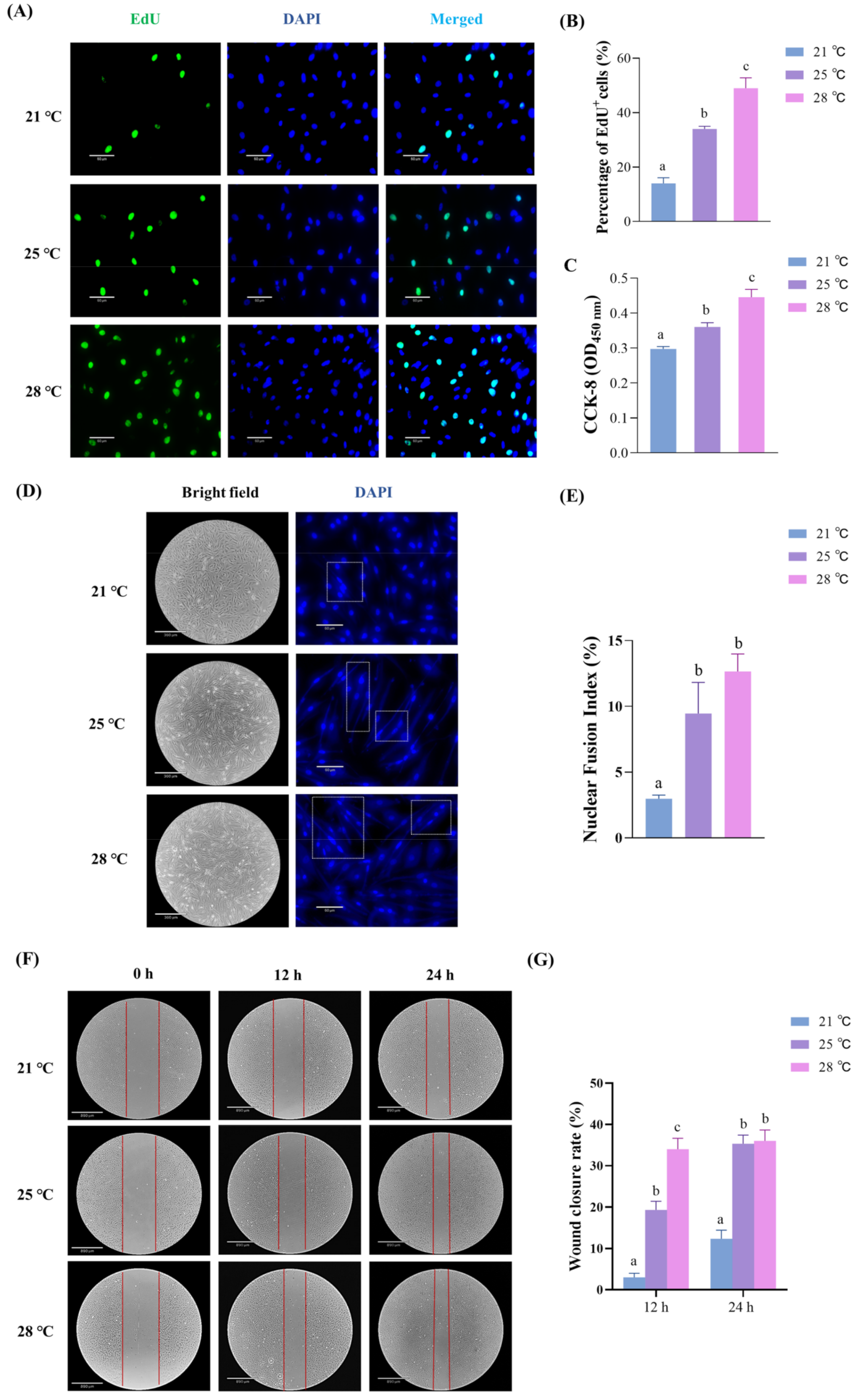
2.1. The Effects of Temperature on the Proliferation, Differentiation and Migration of Skeletal Muscle Cells
2.2. Statistics of RNA-Seq Data
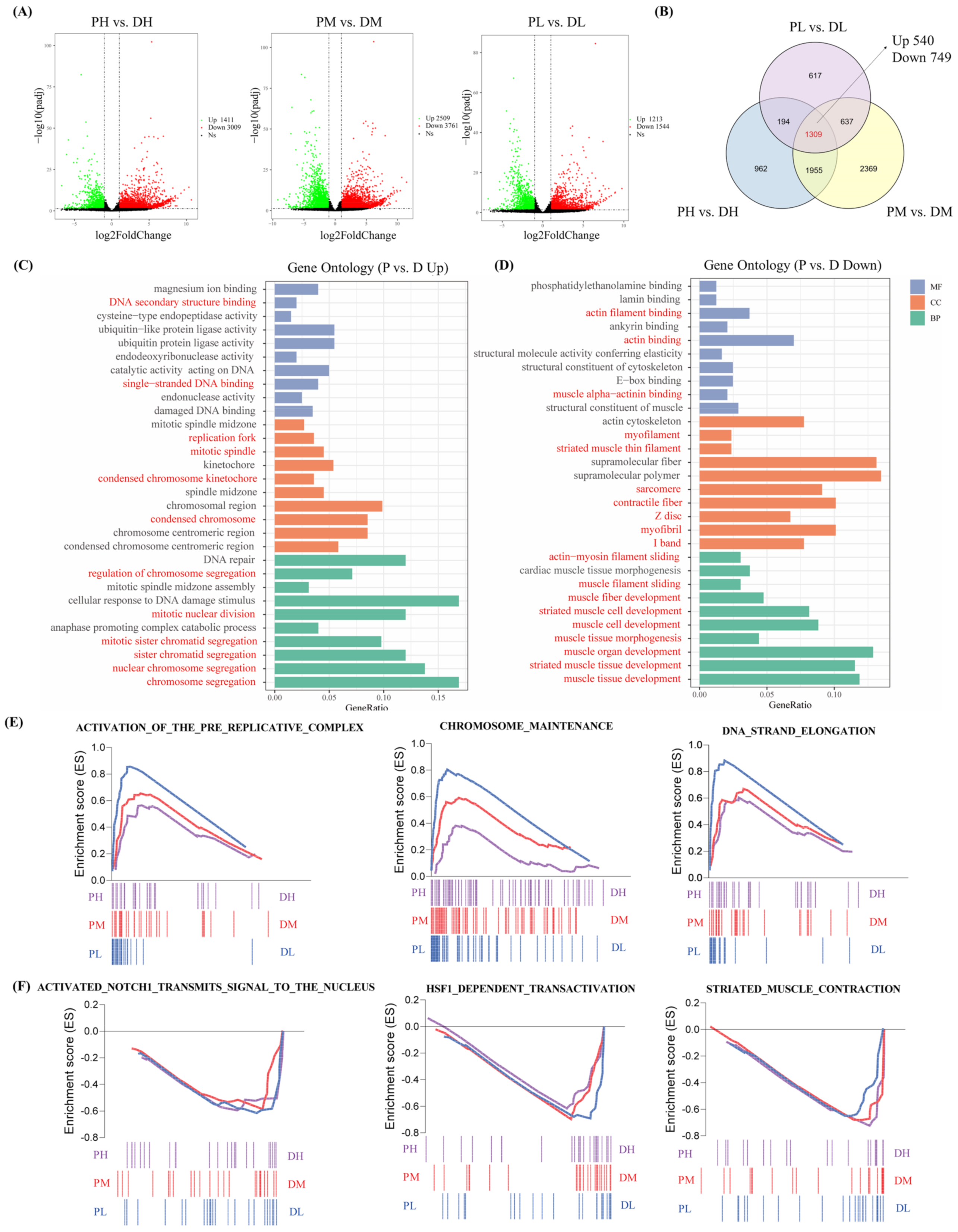
2.3. Comparative Transcriptome Analysis between Cells in the Proliferation and Differentiation Stages
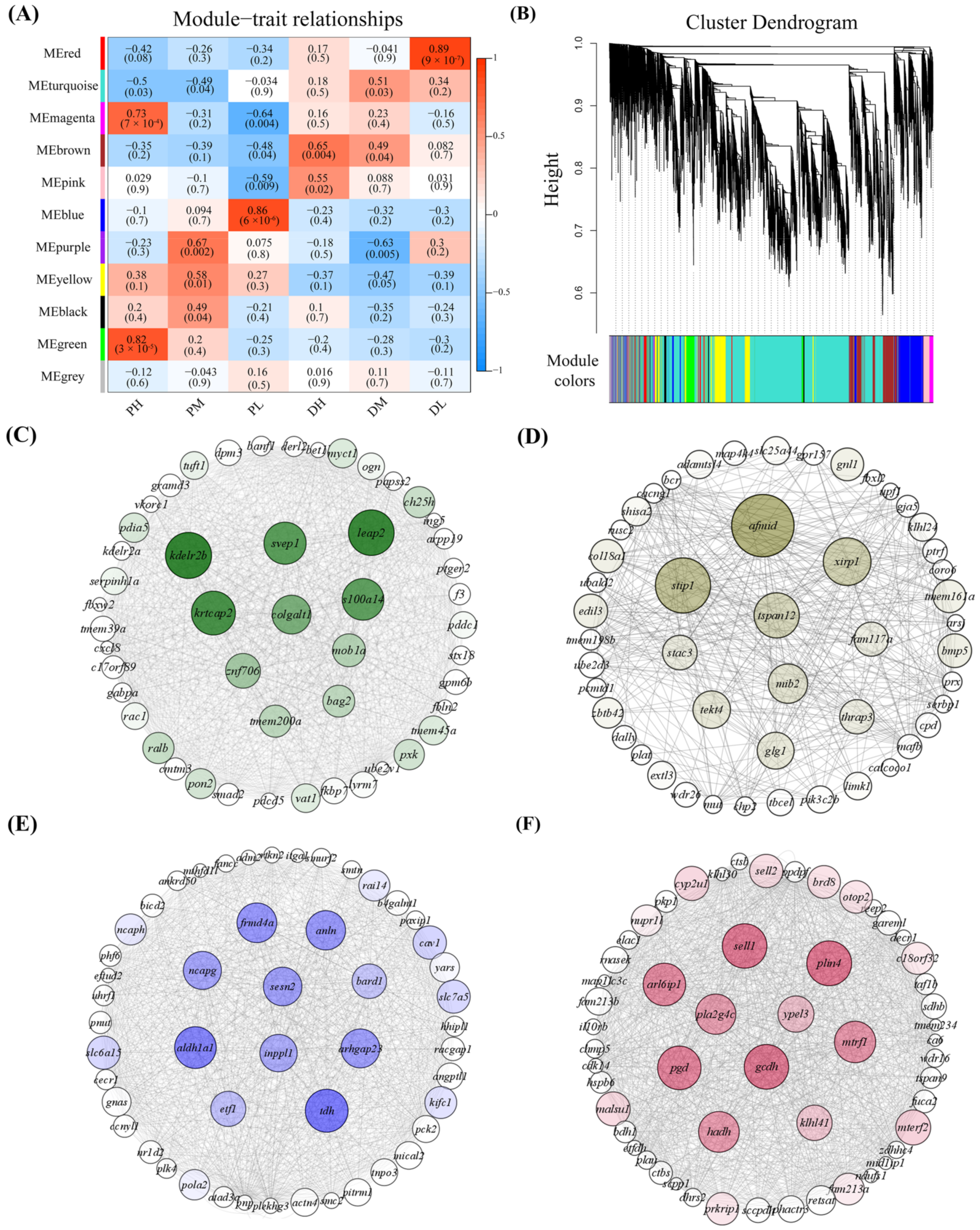
2.4. Identification of Significant Modules and Genes Associated with Temperature by WGCNA
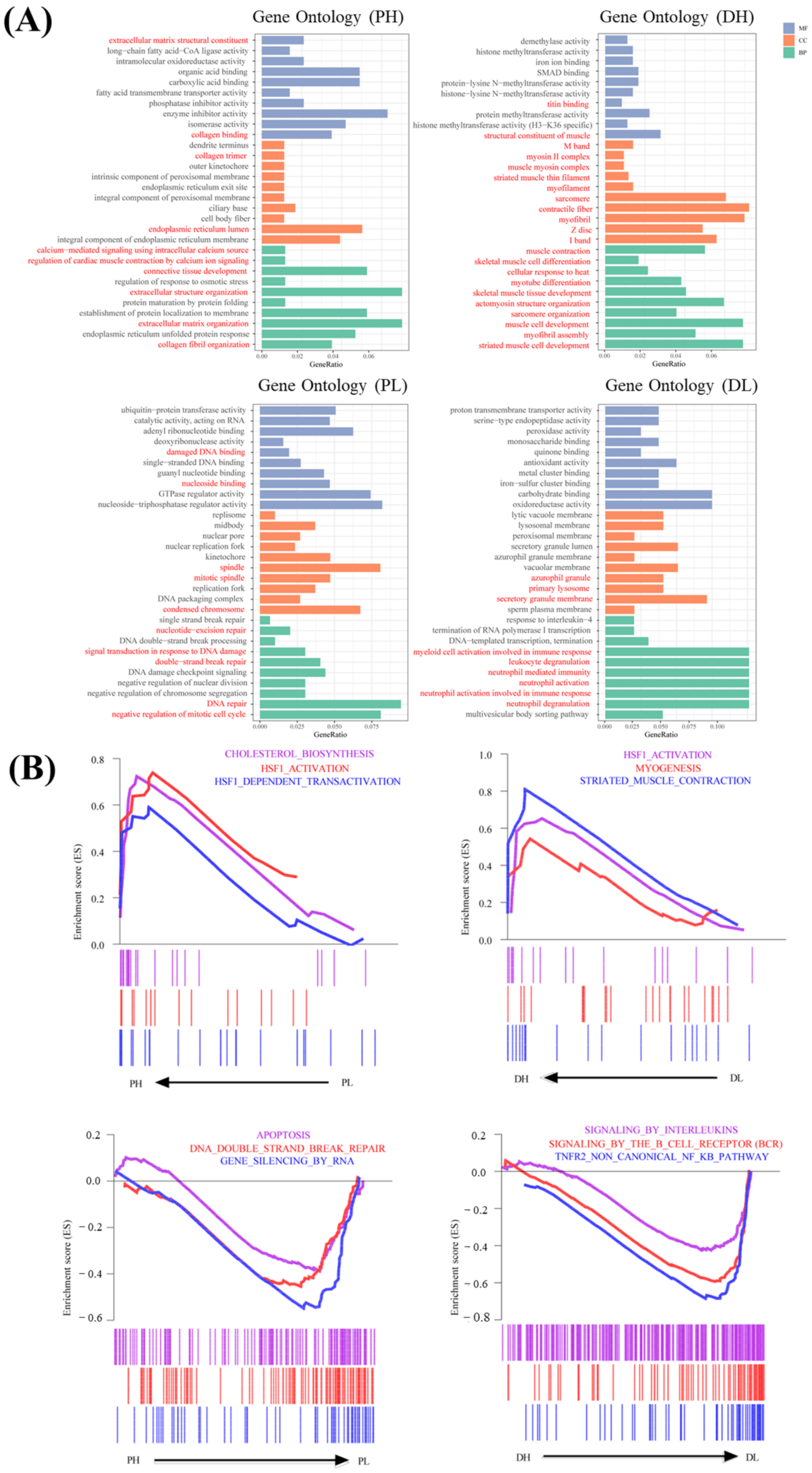
2.5. Functional Enrichment Analysis for Significant Modules Affected by Temperature
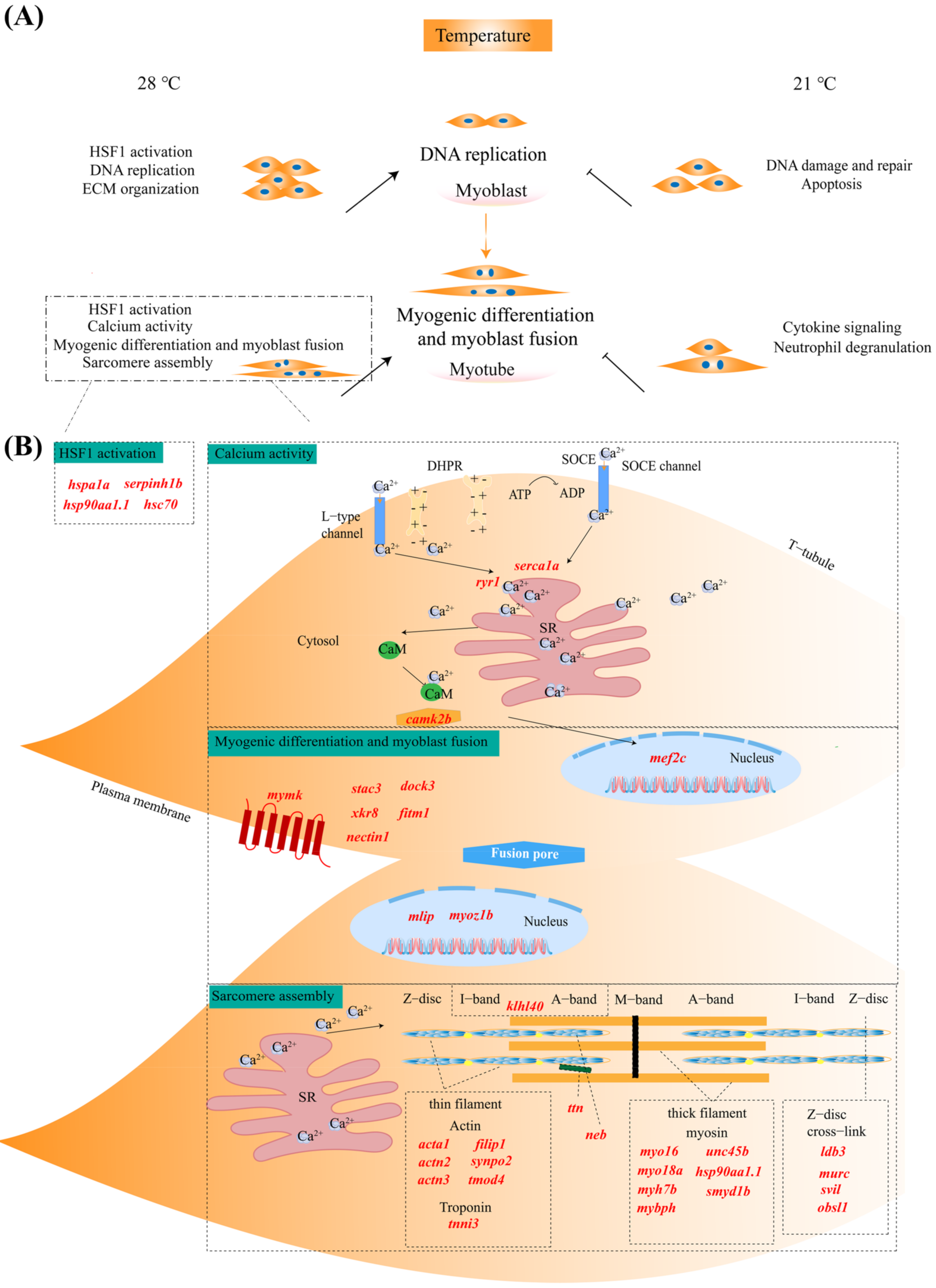
2.6. Putative Functional Roles of Genes in Skeletal Muscle Cells Affected by Temperature
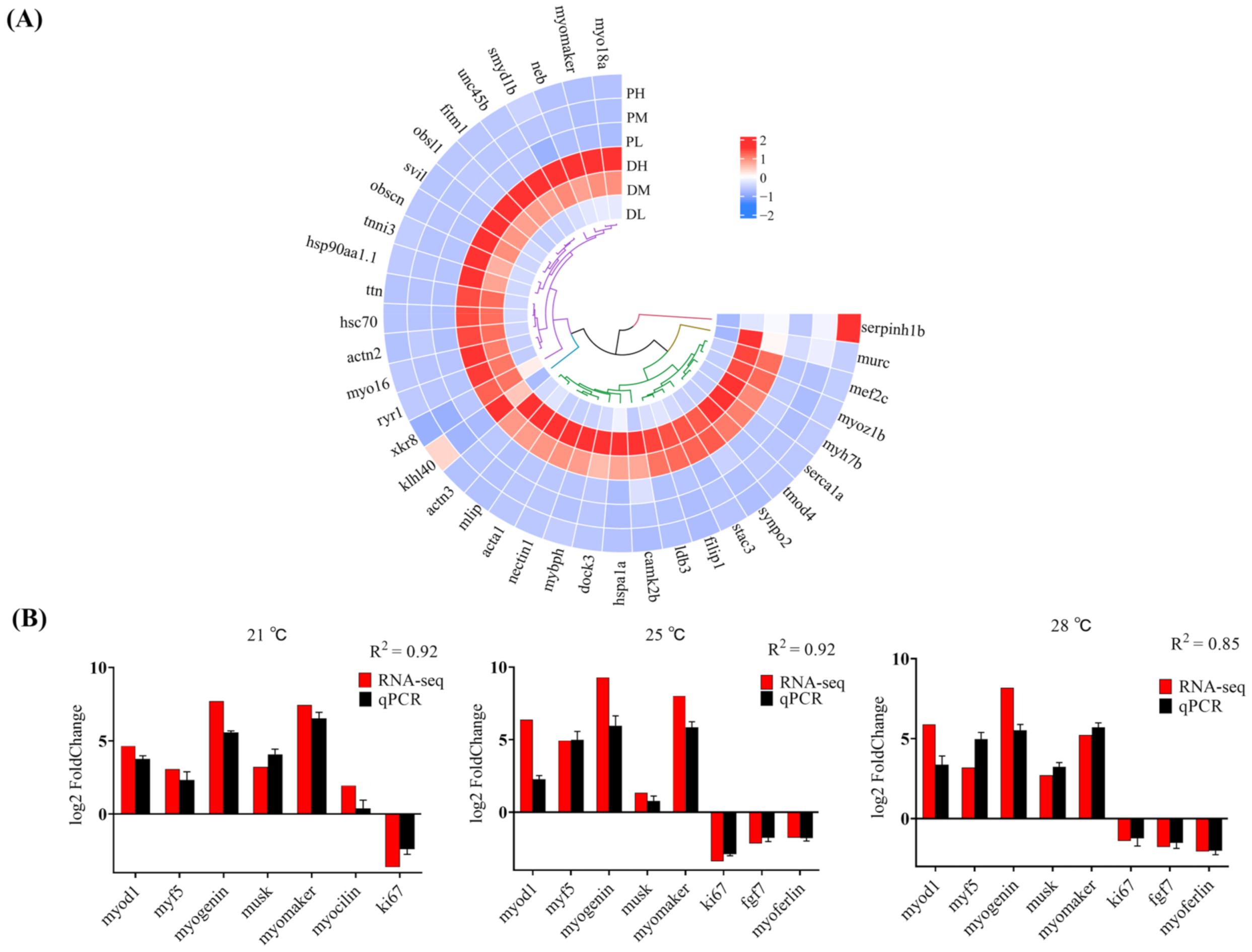
2.7. Validation of RNA-Seq Data via qPCR
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Tissue Isolation and Cell Culture
- Cell proliferation at different temperatures. The isolated cells were plated at a density of 15,000 cells per well in culture flasks and incubated in a 25 °C incubator with GM for 24 h. A subset of cells was divided into three parts of equal amounts and cultured at experimental temperatures of 21 °C, 25 °C, and 28 °C. The GM was changed every 24 h. After 72 h of proliferation, the cell medium was removed, and the cells were rinsed with cold PBS and collected for RNA extraction (Figure S2A).
- Cell differentiation at different temperatures. The other part of the isolated cells was divided into three equal parts and cultured with GM for 72 h of proliferation at 25 °C, reaching approximately 90% confluency. Then, the GM was replaced with differentiation medium (DM) (L-15, 2% horse serum (G-CLONE, Beijing, China), 1% 100 U mL−1 penicillin, and 100 µg mL−1 streptomycin) to induce differentiation, and the cells were cultured in incubators at 21 °C, 25 °C, and 28 °C. The DM was changed every 24 h. After 48 h of differentiation, the cell medium was removed, and the cells were rinsed with cold PBS and collected for RNA extraction (Figure S2B).
4.3. Cell Proliferation Assay
4.4. Cell Differentiation Assay
4.5. In Vitro Wound Scratch Assay
4.6. RNA Extraction and Transcriptome Sequencing
4.7. Identification of Differential Expressed Genes (DEGs)
4.8. Functional Enrichment Analysis
4.9. Weighted Gene Co-Expression Network Analysis (WGCNA) and Hub Genes Identification
4.10. Validation of Gene Expression by Real-Time PCR
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vélez, E.J.; Lutfi, E.; Azizi, S.; Perelló, M.; Salmerón, C.; Riera-Codina, M.; Ibarz, A.; Fernández-Borràs, J.; Blasco, J.; Capilla, E.; et al. Understanding fish muscle growth regulation to optimize aquaculture production. Aquaculture 2017, 467, 28–40. [Google Scholar] [CrossRef]
- Mommsen, T.P. Paradigms of growth in fish. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2001, 129, 207–219. [Google Scholar] [CrossRef]
- Biga, P.R.; Goetz, F.W. Zebrafish and giant danio as models for muscle growth: Determinate vs. indeterminate growth as determined by morphometric analysis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 291, 1327–1337. [Google Scholar] [CrossRef] [PubMed]
- Chal, J.; Pourquié, O. Making muscle: Skeletal myogenesis in vivo and in vitro. Development 2017, 144, 2104–2122. [Google Scholar] [CrossRef]
- Buckingham, M.; Rigby, P.W.J. Gene Regulatory Networks and Transcriptional Mechanisms that Control Myogenesis. Dev. Cell 2014, 28, 225–238. [Google Scholar] [CrossRef]
- Bryson-Richardson, R.J.; Currie, P.D. The genetics of vertebrate myogenesis. Nat. Rev. Genet. 2008, 9, 632–646. [Google Scholar] [CrossRef]
- Zammit, P.S. Function of the myogenic regulatory factors Myf5, MyoD, Myogenin and MRF4 in skeletal muscle, satellite cells and regenerative myogenesis. Semin. Cell Dev. Biol. 2017, 72, 19–32. [Google Scholar] [CrossRef]
- Shan, T.; Xu, Z.; Wu, W.; Liu, J.; Wang, Y. Roles of Notch1 Signaling in Regulating Satellite Cell Fates Choices and Postnatal Skeletal Myogenesis. J. Cell. Physiol. 2017, 232, 2964–2967. [Google Scholar] [CrossRef]
- Johnston, I.A.; Bower, N.I.; Macqueen, D.J. Growth and the regulation of myotomal muscle mass in teleost fish. J. Exp. Biol. 2011, 214, 1617–1628. [Google Scholar] [CrossRef]
- Steinbacher, P.; Marschallinger, J.; Obermayer, A.; Neuhofer, A.; Sänger, A.M.; Stoiber, W. Temperature-dependent modification of muscle precursor cell behaviour is an underlying reason for lasting effects on muscle cellularity and body growth of teleost fish. J. Exp. Biol. 2011, 214, 1791–1801. [Google Scholar] [CrossRef] [Green Version]
- Garcia de la serrana, D.; Devlin, R.H.; Johnston, I.A. RNAseq analysis of fast skeletal muscle in restriction-fed transgenic coho salmon (Oncorhynchus kisutch): An experimental model uncoupling the growth hormone and nutritional signals regulating growth. BMC Genom. 2015, 16, 564. [Google Scholar] [CrossRef] [PubMed]
- Melo, L.H.; Martins, Y.S.; Melo, R.M.C.; Prado, P.S.; Luz, R.K.; Bazzoli, N.; Rizzo, E. Low salinity negatively affects early larval development of Nile tilapia, Oreochromis niloticus: Insights from skeletal muscle and molecular biomarkers. Zygote 2019. [Google Scholar] [CrossRef]
- Rossi, G.S.; Turko, A.J.; Wright, P.A. Oxygen drives skeletal muscle remodeling in an amphibious fish out of water. J. Exp. Biol. 2018, 221, jeb180257. [Google Scholar] [CrossRef] [PubMed]
- Garcia de la serrana, D.; Vieira, V.L.A.; Andree, K.B.; Darias, M.; Estévez, A.; Gisbert, E.; Johnston, I.A. Development Temperature Has Persistent Effects on Muscle Growth Responses in Gilthead Sea Bream. PLoS ONE 2012, 7, e51884. [Google Scholar] [CrossRef] [PubMed]
- Johnston, I.A.; Lee, H.T.; Macqueen, D.J.; Paranthaman, K.; Kawashima, C.; Anwar, A.; Kinghorn, J.R.; Dalmay, T. Embryonic temperature affects muscle fibre recruitment in adult zebrafish: Genome-wide changes in gene and microRNA expression associated with the transition from hyperplastic to hypertrophic growth phenotypes. J. Exp. Biol. 2009, 212, 1781–1793. [Google Scholar] [CrossRef]
- Anastasiadi, D.; Díaz, N.; Piferrer, F. Small ocean temperature increases elicit stage-dependent changes in DNA methylation and gene expression in a fish, the European sea bass. Sci. Rep. 2017, 7, 12401. [Google Scholar] [CrossRef]
- Furuichi, Y.; Kawabata, Y.; Aoki, M.; Mita, Y.; Fujii, N.L.; Manabe, Y. Excess Glucose Impedes the Proliferation of Skeletal Muscle Satellite Cells Under Adherent Culture Conditions. Front. Cell Dev. Biol. 2021, 9, 640399. [Google Scholar] [CrossRef]
- Kelc, R.; Trapecar, M.; Vogrin, M.; Cencic, A. Skeletal muscle-derived cell cultures as potent models in regenerative medicine research. Muscle Nerve 2013, 47, 477–482. [Google Scholar] [CrossRef]
- Millay, D.P.; O’Rourke, J.R.; Sutherland, L.B.; Bezprozvannaya, S.; Shelton, J.M.; Bassel-Duby, R.; Olson, E.N. Myomaker is a membrane activator of myoblast fusion and muscle formation. Nature 2013, 499, 301–305. [Google Scholar] [CrossRef]
- Bi, P.; Ramirez-Martinez, A.; Li, H.; Cannavino, J.; McAnally, J.R.; Shelton, J.M.; Sánchez-Ortiz, E.; Bassel-Duby, R.; Olson, E.N. Control of muscle formation by the fusogenic micropeptide myomixer. Science 2017, 356, 323–327. [Google Scholar] [CrossRef] [Green Version]
- Ciecierska, A.; Motyl, T.; Sadkowski, T. Transcriptomic profile of primary culture of skeletal muscle cells isolated from semitendinosus muscle of beef and dairy bulls. Int. J. Mol. Sci. 2020, 21, 4794. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Wang, X.; Li, M.; Song, W.; Huang, K.; Zhang, F.; Zhang, Q.; Qi, J.; He, Y. Establishment of myoblast cell line and identification of key genes regulating myoblast differentiation in a marine teleost, Sebastes schlegelii. Gene 2021, 802, 145869. [Google Scholar] [CrossRef] [PubMed]
- Metzger, K.; Dannenberger, D.; Tuchscherer, A.; Ponsuksili, S.; Kalbe, C. Effects of temperature on proliferation of myoblasts from donor piglets with different thermoregulatory maturities. BMC Mol. Cell Biol. 2021, 22, 36. [Google Scholar] [CrossRef]
- Clark, D.L.; Coy, C.S.; Strasburg, G.M.; Reed, K.M.; Velleman, S.G. Temperature effect on proliferation and differentiation of satellite cells from Turkeys with different growth rates. Poult. Sci. 2016, 95, 934–947. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, N.; Goswami, M.; Srivastava, J.K.; Mishra, A.K.; Lakra, W.S. Establishment and characterization of a new muscle cell line of zebrafish (Danio rerio) as an in vitro model for gene expression studies. Anim. Biotechnol. 2016, 27, 166–173. [Google Scholar] [CrossRef]
- Landemaine, A.; Ramirez-Martinez, A.; Monestier, O.; Sabin, N.; Rescan, P.Y.; Olson, E.N.; Gabillard, J.C. Trout myomaker contains 14 minisatellites and two sequence extensions but retains fusogenic function. J. Biol. Chem. 2019, 294, 6364–6374. [Google Scholar] [CrossRef]
- Lu, K.L.; Cai, L.S.; Wang, L.; Song, K.; Zhang, C.X.; Rahimnejad, S. Effects of dietary protein/energy ratio and water temperature on growth performance, digestive enzymes activity and non-specific immune response of spotted seabass (Lateolabrax maculatus). Aquac. Nutr. 2020, 26, 2023–2031. [Google Scholar] [CrossRef]
- Shin, M.K.; Park, H.R.; Yeo, W.J.; Han, K.N. Effects of Thermal Stress on the mRNA Expression of SOD, HSP90, and HSP70 in the Spotted Sea Bass (Lateolabrax maculatus). Ocean Sci. J. 2018, 53, 43–52. [Google Scholar] [CrossRef]
- Cheng, Y.; Li, X.; Wang, L.; Lu, K.; Song, K.; Ai, Q.; Mai, K.; Zhang, C. Effects of dietary arginine levels on growth, immune function of physical barriers and serum parameters of spotted seabass (Lateolabrax maculatus) reared at different water temperatures. Aquaculture 2021, 541, 736812. [Google Scholar] [CrossRef]
- Person-Le Ruyet, J.; Mahé, K.; Le Bayon, N.; Le Delliou, H. Effects of temperature on growth and metabolism in a Mediterranean population of European sea bass, Dicentrarchus labrax. Aquaculture 2004, 237, 269–280. [Google Scholar] [CrossRef] [Green Version]
- Guderley, H. Metabolic responses to low temperature in fish muscle. Biol. Rev. Camb. Philos. Soc. 2004, 79, 409–427. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y. Studies on the Mechanism and Physiological Response to Temperature Tolerance of Juvenile Spotted Sea Bass (Lateolabrax maculatus); Ocean University of China: Qingdao, China, 2019. [Google Scholar]
- Harding, R.L.; Halevy, O.; Yahav, S.; Velleman, S.G. The effect of temperature on proliferation and differentiation of chicken skeletal muscle satellite cells isolated from different muscle types. Physiol. Rep. 2016, 4, e12770. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, S.H.; Subramaniyan, S.A.; Kang, D.; Park, J.; Khan, M.; Shim, K. Modulatory effect of heat stress on viability of primary cultured chicken satellite cells and expression of heat shock proteins ex vivo. Anim. Biotechnol. 2021, 32, 774–785. [Google Scholar] [CrossRef]
- Schöffl, F.; Prandl, R.; Reindl, A. Update on Signal Transduction Regulation of the Heat-Shock Response. Plant Physiol. 1998, 117, 1135–1141. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Tu, K.; Fu, Q.; Schmitt, D.C.; Zhou, L.; Lu, N.; Zhao, Y. Multifaceted roles of HSF1 in cancer. Tumor Biol. 2015, 36, 4923–4931. [Google Scholar] [CrossRef]
- Baldelli, S.; Limongi, D.; Coni, C.; Ciccarone, F.; Ciotti, M.; Checconi, P.; Palamara, A.T.; Ciriolo, M.R. BK Polyomavirus Activates HSF1 Stimulating Human Kidney Hek293 Cell Proliferation. Oxid. Med. Cell. Longev. 2021, 2021, 917699. [Google Scholar] [CrossRef] [PubMed]
- Bhatti, M.; Dinn, S.; Miskiewicz, E.I.; MacPhee, D.J. Expression of heat shock factor 1, heat shock protein 90 and associated signaling proteins in pregnant rat myometrium: Implications for myometrial proliferation. Reprod. Biol. 2019, 19, 374–385. [Google Scholar] [CrossRef]
- Zamkova, M.; Khromova, N.; Kopnin, B.P.; Kopnin, P. Ras-induced ROS upregulation affecting cell proliferation is connected with cell type-specific alterations of HSF1/SESN3/p21Cip1/WAF1 pathways. Cell Cycle 2013, 12, 826–836. [Google Scholar] [CrossRef]
- Shi, X.; Deng, Z.; Wang, S.; Zhao, S.; Xiao, L.; Zou, J.; Li, T.; Tan, S.; Tan, S.; Xiao, X. Increased hsf1 promotes infiltration and metastasis in cervical cancer via enhancing mtdh-vegf-c expression. OncoTargets Ther. 2021, 14, 1305–1315. [Google Scholar] [CrossRef]
- Ohno, Y.; Egawa, T.; Yokoyama, S.; Nakai, A.; Sugiura, T.; Ohira, Y.; Yoshioka, T.; Goto, K. Deficiency of heat shock transcription factor 1 suppresses heat stress-associated increase in slow soleus muscle mass of mice. Acta Physiol. 2015, 215, 191–203. [Google Scholar] [CrossRef]
- Cloutier, G.; Sallenbach-Morrissette, A.; Beaulieu, J.F. Non-integrin laminin receptors in epithelia. Tissue Cell 2019, 56, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liu, Y.; Zhang, H. Extracellular matrix: An important regulator of cell functions and skeletal muscle development. Cell Biosci. 2021, 11, 65. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-X.; Wu, B.-B.; Gong, L.; An, C.-R.; Lin, J.-X.; Li, Q.-K.; Jiang, D.-M.; Jin, K.-X.; Mechakra, A.; Bunpetch, V.; et al. Dissecting cell diversity and connectivity in skeletal muscle for myogenesis. Cell Death Dis. 2019, 10, 427. [Google Scholar] [CrossRef] [PubMed]
- Stern, M.M.; Myers, R.L.; Hammam, N.; Stern, K.A.; Eberli, D.; Kritchevsky, S.B.; Soker, S.; Van Dyke, M. The influence of extracellular matrix derived from skeletal muscle tissue on the proliferation and differentiation of myogenic progenitor cells ex vivo. Biomaterials 2009, 30, 2393–2399. [Google Scholar] [CrossRef]
- Koohestani, F.; Braundmeier, A.G.; Mahdian, A.; Seo, J.; Bi, J.J.; Nowak, R.A. Extracellular matrix collagen alters cell proliferation and cell cycle progression of human uterine leiomyoma smooth muscle cells. PLoS ONE 2013, 8, e75844. [Google Scholar] [CrossRef]
- Dumont, N.A.; Bentzinger, C.F.; Sincennes, M.C.; Rudnicki, M.A. Satellite cells and skeletal muscle regeneration. Compr. Physiol. 2015, 5, 1027–1059. [Google Scholar] [CrossRef]
- Du, S.J.; Li, H.; Bian, Y.; Zhong, Y. Heat-shock protein 90α1 is required for organized myofibril assembly in skeletal muscles of zebrafish embryos. Proc. Natl. Acad. Sci. USA 2008, 105, 554–559. [Google Scholar] [CrossRef]
- Gao, C.-Q.; Zhao, Y.-L.; Li, H.; Sui, W.-G.; Yan, H.-C.; Wang, X.-Q. Heat stress inhibits proliferation, promotes growth, and induces apoptosis in cultured Lantang swine skeletal muscle satellite cells. J. Zhejiang Univ. Sci. B 2015, 16, 549–559. [Google Scholar] [CrossRef]
- Oguro, A.; Sakurai, T.; Fujita, Y.; Lee, S.; Kubota, H.; Nagata, K.; Atomi, Y. The molecular chaperone HSP47 rapidly senses gravitational changes in myoblasts. Genes Cells 2006, 11, 1253–1265. [Google Scholar] [CrossRef]
- Gehlert, S.; Bloch, W.; Suhr, F. Ca2+-dependent regulations and signaling in skeletal muscle: From electro-mechanical coupling to adaptation. Int. J. Mol. Sci. 2015, 16, 1066–1095. [Google Scholar] [CrossRef]
- Friday, B.B.; Mitchell, P.O.; Kegley, K.M.; Pavlath, G.K. Calcineurin initiates skeletal muscle differentiation by activating MEF2 and MyoD. Differentiation 2003, 71, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Meissner, G. The structural basis of ryanodine receptor ion channel function. J. Gen. Physiol. 2017, 149, 1065–1089. [Google Scholar] [CrossRef] [PubMed]
- Rassier, D.E. Sarcomere mechanics in striated muscles: From molecules to sarcomeres to cells. Am. J. Physiol. Cell Physiol. 2017, 313, C134–C145. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.J.; Hyun, C.; Woo, J.S.; Park, C.S.; Kim, D.H.; Lee, E.H. Stromal interaction molecule 1 (STIM1) regulates sarcoplasmic/endoplasmic reticulum Ca2+-ATPase 1a (SERCA1a) in skeletal muscle. Pflugers Arch. Eur. J. Physiol. 2014, 466, 987–1001. [Google Scholar] [CrossRef] [PubMed]
- Stringer, R.N.; Jurkovicova-Tarabova, B.; Huang, S.; Haji-Ghassemi, O.; Idoux, R.; Liashenko, A.; Souza, I.A.; Rzhepetskyy, Y.; Lacinova, L.; Van Petegem, F.; et al. A rare CACNA1H variant associated with amyotrophic lateral sclerosis causes complete loss of Cav3.2 T-type channel activity. Mol. Brain 2020, 13, 33. [Google Scholar] [CrossRef] [PubMed]
- Perestenko, P.; Watanabe, M.; Beusnard-Bee, T.; Guna, P.; McIlhinney, J. The second C2-domain of copine-2, copine-6 and copine-7 is responsible for their calcium-dependent membrane association. FEBS J. 2015, 282, 3722–3736. [Google Scholar] [CrossRef]
- Konig, S.; Béguet, A.; Bader, C.R.; Bernheim, L. The calcineurin pathway links hyperpolarization (Kir2.1)-induced Ca2+ signals to human myoblast differentiation and fusion. Development 2006, 133, 3107–3114. [Google Scholar] [CrossRef]
- Benavides Damm, T.; Egli, M. Calcium’s role in mechanotransduction during muscle development. Cell. Physiol. Biochem. 2014, 33, 249–272. [Google Scholar] [CrossRef]
- Ridgeway, A.G.; Wilton, S.; Skerjanc, I.S. Myocyte enhancer factor 2C and myogenin up-regulate each other’s expression and induce the development of skeletal muscle in P19 cells. J. Biol. Chem. 2000, 275, 41–46. [Google Scholar] [CrossRef]
- Jin, W.; Liu, M.; Peng, J.; Jiang, S. Function analysis of Mef2c promoter in muscle differentiation. Biotechnol. Appl. Biochem. 2017, 64, 647–656. [Google Scholar] [CrossRef]
- Dodou, E.; Xu, S.M.; Black, B.L. Mef2C Is Activated Directly By Myogenic Basic Helix-Loop-Helix Proteins during Skeletal Muscle Development in Vivo. Mech. Dev. 2003, 120, 1021–1032. [Google Scholar] [CrossRef]
- Cong, X.; Doering, J.; Mazala, D.A.G.; Chin, E.R.; Grange, R.W.; Jiang, H. The SH3 and cysteine-rich domain 3 (Stac3) gene is important to growth, fiber composition, and calcium release from the sarcoplasmic reticulum in postnatal skeletal muscle. Skelet. Muscle 2016, 6, 17. [Google Scholar] [CrossRef] [PubMed]
- Polster, A.; Nelson, B.R.; Olson, E.N.; Beam, K.G. Stac3 has a direct role in skeletal muscle-type excitation-contraction coupling that is disrupted by a myopathy-causing mutation. Proc. Natl. Acad. Sci. USA 2016, 113, 10986–10991. [Google Scholar] [CrossRef] [PubMed]
- Reinholt, B.M.; Ge, X.; Cong, X.; Gerrard, D.E.; Jiang, H. Stac3 Is a Novel Regulator of Skeletal Muscle Development in Mice. PLoS ONE 2013, 8, e62760. [Google Scholar] [CrossRef]
- Bower, N.I.; Garcia De La Serrana, D.; Cole, N.J.; Hollway, G.E.; Lee, H.T.; Assinder, S.; Johnston, I.A. Stac3 is required for myotube formation and myogenic differentiation in vertebrate skeletal muscle. J. Biol. Chem. 2012, 287, 43936–43949. [Google Scholar] [CrossRef]
- Reid, A.L.; Wang, Y.; Samani, A.; Hightower, R.M.; Lopez, M.A.; Gilbert, S.R.; Ianov, L.; Crossman, D.K.; Dell’Italia, L.J.; Millay, D.P.; et al. DOCK3 is a dosage-sensitive regulator of skeletal muscle and Duchenne muscular dystrophy-associated pathologies. Hum. Mol. Genet. 2020, 29, 2855–2871. [Google Scholar] [CrossRef]
- Ren, R.M.; Liu, H.; Zhao, S.H.; Cao, J.H. Targeting of miR-432 to myozenin1 to regulate myoblast proliferation and differentiation. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef]
- Kim, G.W.; Nam, G.H.; Kim, I.S.; Park, S.Y. Xk-related protein 8 regulates myoblast differentiation and survival. FEBS J. 2017, 284, 3575–3588. [Google Scholar] [CrossRef]
- Henderson, C.A.; Gomez, C.G.; Novak, S.M.; Mi-Mi, L.; Gregorio, C.C. Overview of the muscle cytoskeleton. Compr. Physiol. 2017, 7, 891–944. [Google Scholar] [CrossRef]
- Ebashi, S.; Ebashi, F. Alpha-actinin, a new structural protein from striated muscle. I. Preparation and action on actomyosin-ATP interaction. J. Biochem. 1965, 58, 7–12. [Google Scholar] [CrossRef]
- Militello, G.; Hosen, M.R.; Ponomareva, Y.; Gellert, P.; Weirick, T.; John, D.; Hindi, S.M.; Mamchaoui, K.; Mouly, V.; Döring, C.; et al. A novel long non-coding RNA Myolinc regulates myogenesis through TDP-43 and Filip1. J. Mol. Cell Biol. 2018, 10, 102–117. [Google Scholar] [CrossRef]
- Kai, F.B.; Fawcett, J.P.; Duncan, R. Synaptopodin-2 induces assembly of peripheral actin bundles and immature focal adhesions to promote lamellipodia formation and prostate cancer cell migration. Oncotarget 2015, 6, 11162–11174. [Google Scholar] [CrossRef] [PubMed]
- Sheng, J.-J.; Jin, J.-P. TNNI1, TNNI2 and TNNI3: Evolution, regulation, and protein structure–function relationships. Gene 2015, 576, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Gokhin, D.S.; Lewis, R.A.; McKeown, C.R.; Nowak, R.B.; Kim, N.E.; Littlefield, R.S.; Lieber, R.L.; Fowler, V.M. Tropomodulin isoforms regulate thin filament pointed-end capping and skeletal muscle physiology. J. Cell Biol. 2010, 189, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Garg, A.; O’Rourke, J.; Long, C.; Doering, J.; Ravenscroft, G.; Bezprozvannaya, S.; Nelson, B.R.; Beetz, N.; Li, L.; Chen, S.; et al. KLHL40 deficiency destabilizes thin filament proteins and promotes Nemaline myopathy. J. Clin. Investig. 2014, 124, 3529–3539. [Google Scholar] [CrossRef] [PubMed]
- Mouton, J.; Loos, B.; Moolman-Smook, J.C.; Kinnear, C.J. Ascribing novel functions to the sarcomeric protein, myosin binding protein H (MyBPH) in cardiac sarcomere contraction. Exp. Cell Res. 2015, 331, 338–351. [Google Scholar] [CrossRef]
- Etard, C.; Armant, O.; Roostalu, U.; Gourain, V.; Ferg, M.; Strähle, U. Loss of function of myosin chaperones triggers Hsf1-mediated transcriptional response in skeletal muscle cells. Genome Biol. 2015, 16, 267. [Google Scholar] [CrossRef]
- Armant, O.; Gourain, V.; Etard, C.; Strähle, U. Whole transcriptome data analysis of zebrafish mutants affecting muscle development. Data Br. 2016, 8, 61–68. [Google Scholar] [CrossRef]
- Tagawa, M.; Ueyama, T.; Ogata, T.; Takehara, N.; Nakajima, N.; Isodono, K.; Asada, S.; Takahashi, T.; Matsubara, H.; Oh, H. MURC, a muscle-restricted coiled-coil protein, is involved in the regulation of skeletal myogenesis. Am. J. Physiol. Cell Physiol. 2008, 295, 490–498. [Google Scholar] [CrossRef]
- Hedberg-Oldfors, C.; Meyer, R.; Nolte, K.; Rahim, Y.A.; Lindberg, C.; Karason, K.; Thuestad, I.J.; Visuttijai, K.; Geijer, M.; Begemann, M.; et al. Loss of supervillin causes myopathy with myofibrillar disorganization and autophagic vacuoles. Brain 2020, 143, 2406–2420. [Google Scholar] [CrossRef]
- Geister, K.A.; Lopez-Jimenez, A.J.; Houghtaling, S.; Ho, T.H.; Vanacore, R.; Beier, D.R. Loss of function of Colgalt1 disrupts collagen post-translational modification and causes musculoskeletal defects. DMM Dis. Model. Mech. 2019, 12, dmm037176. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhu, J.; Mao, Z.; Zhang, G.; Hu, X.; Chen, F. Tuft1 promotes thyroid carcinoma cell invasion and proliferation and suppresses apoptosis through the Akt-mTOR/GSK3β signaling pathway. Am. J. Transl. Res. 2018, 10, 4376–4384. [Google Scholar] [PubMed]
- Lin, H.; Zeng, W.; Lei, Y.; Chen, D.; Nie, Z. Tuftelin 1 (TUFT1) Promotes the Proliferation and Migration of Renal Cell Carcinoma via PI3K/AKT Signaling Pathway. Pathol. Oncol. Res. 2021, 27, 640936. [Google Scholar] [CrossRef]
- Sun, L.; Chen, G.; Sun, A.; Wang, Z.; Huang, H.; Gao, Z.; Liang, W.; Liu, C.; Li, K. BAG2 Promotes Proliferation and Metastasis of Gastric Cancer via ERK1/2 Signaling and Partially Regulated by miR186. Front. Oncol. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Ma, W.; Lu, X.; Liu, H.; Lin, K.; Wang, Y.; Ye, Z.; Sun, L.; Huang, Z.; Pan, T.; et al. KDELR2 promotes breast cancer proliferation via HDAC3-mediated cell cycle progression. Cancer Commun. 2021, 41, 904–920. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Gao, W.; Li, X.; Yu, L.; Luo, D.; Liu, Y.; Yu, X. S100A14 promotes progression and gemcitabine resistance in pancreatic cancer. Pancreatology 2021, 21, 589–598. [Google Scholar] [CrossRef]
- Wang, F.; Xue, Q.; Xu, D.; Jiang, Y.; Tang, C.; Liu, X. Identifying the hub gene in gastric cancer by bioinformatics analysis and in vitro experiments. Cell Cycle 2020, 19, 1326–1337. [Google Scholar] [CrossRef]
- Wang, C.; Wang, Z.; Zhang, L.; Lin, X. MiR-29c inhibits the metastasis of oral squamous cell carcinoma and promotes its cell cycle arrest by targeting SERPINH1. Ann. Transl. Med. 2021, 9, 1423. [Google Scholar] [CrossRef] [PubMed]
- Carrasco-Rando, M.; Ruiz-Gómez, M. Mind bomb 2, a founder myoblast-specific protein, regulates myoblast fusion and muscle stability. Development 2008, 135, 849–857. [Google Scholar] [CrossRef]
- Beckley, S.J.; Hunter, M.C.; Kituyi, S.N.; Wingate, I.; Chakraborty, A.; Schwarz, K.; Makhubu, M.P.; Rousseau, R.P.; Ruck, D.K.; de la Mare, J.A.; et al. STIP1/HOP regulates the actin cytoskeleton through interactions with actin and changes in actin-binding proteins cofilin and profilin. Int. J. Mol. Sci. 2020, 21, 3152. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Voza, F.; Ezzeddine, N.; Frasch, M. Drosophila mind bomb2 is required for maintaining muscle integrity and survival. J. Cell Biol. 2007, 179, 219–227. [Google Scholar] [CrossRef]
- Noë, S.; Corvelyn, M.; Willems, S.; Costamagna, D.; Aerts, J.-M.; Van Campenhout, A.; Desloovere, K. The Myotube Analyzer: How to assess myogenic features in muscle stem cells. Skelet. Muscle 2022, 12, 12. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. feature Counts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef]
- Reimand, J.; Isserlin, R.; Voisin, V.; Kucera, M.; Tannus-Lopes, C.; Rostamianfar, A.; Wadi, L.; Meyer, M.; Wong, J.; Xu, C.; et al. Pathway enrichment analysis and visualization of omics data using g: Profiler, GSEA, Cytoscape and EnrichmentMap. Nat. Protoc. 2019, 14, 482–517. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Wen, H.; Qi, X.; Zhang, Y.; Dong, X.; Zhang, K.; Zhang, M.; Li, J.; Li, Y. Morphological and Molecular Responses of Lateolabrax maculatus Skeletal Muscle Cells to Different Temperatures. Int. J. Mol. Sci. 2022, 23, 9812. https://doi.org/10.3390/ijms23179812
Zhang J, Wen H, Qi X, Zhang Y, Dong X, Zhang K, Zhang M, Li J, Li Y. Morphological and Molecular Responses of Lateolabrax maculatus Skeletal Muscle Cells to Different Temperatures. International Journal of Molecular Sciences. 2022; 23(17):9812. https://doi.org/10.3390/ijms23179812
Chicago/Turabian StyleZhang, Jingru, Haishen Wen, Xin Qi, Yonghang Zhang, Ximeng Dong, Kaiqiang Zhang, Meizhao Zhang, Jifang Li, and Yun Li. 2022. "Morphological and Molecular Responses of Lateolabrax maculatus Skeletal Muscle Cells to Different Temperatures" International Journal of Molecular Sciences 23, no. 17: 9812. https://doi.org/10.3390/ijms23179812
APA StyleZhang, J., Wen, H., Qi, X., Zhang, Y., Dong, X., Zhang, K., Zhang, M., Li, J., & Li, Y. (2022). Morphological and Molecular Responses of Lateolabrax maculatus Skeletal Muscle Cells to Different Temperatures. International Journal of Molecular Sciences, 23(17), 9812. https://doi.org/10.3390/ijms23179812