

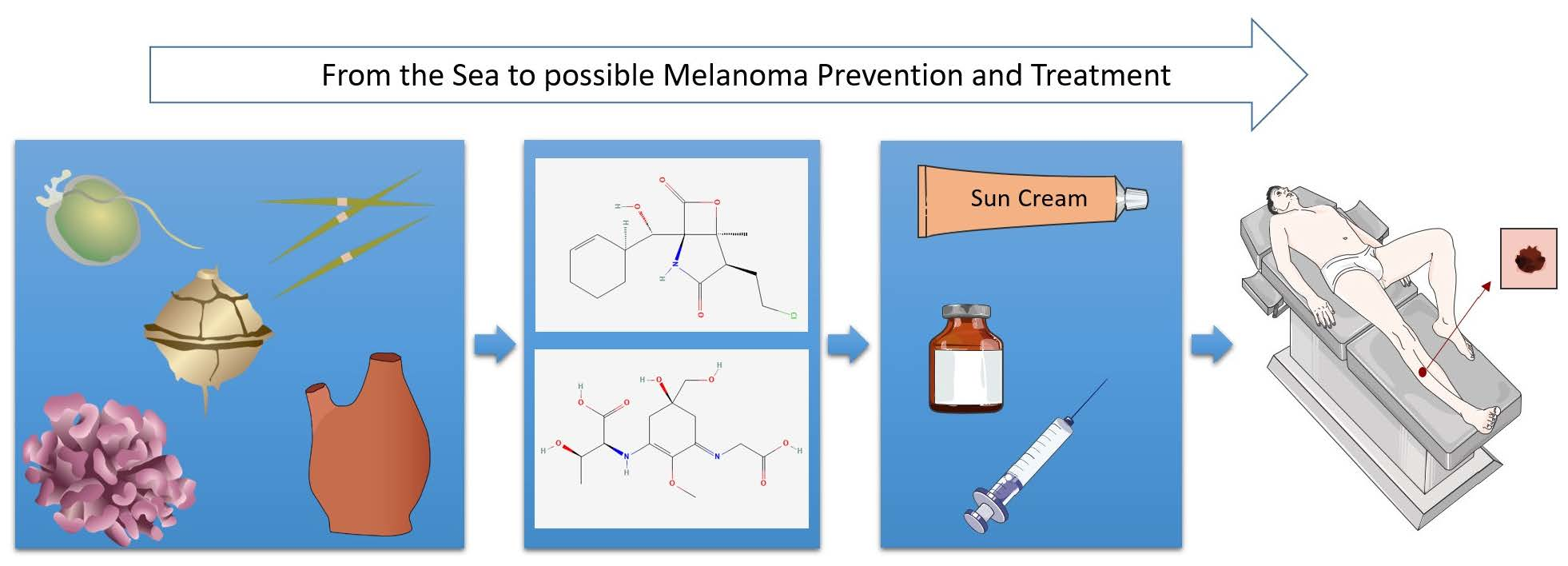
Marine Compounds for Melanoma Treatment and Prevention




Abstract
:
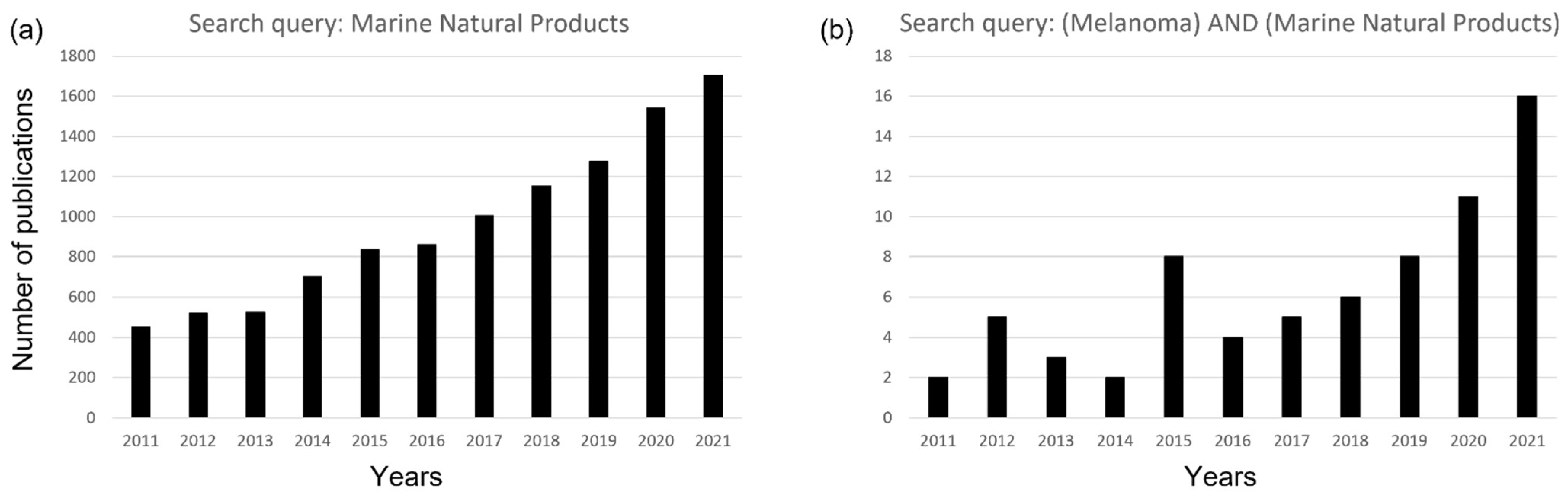
1. Introduction
2. Marine Microorganisms
2.1. Bacteria
2.2. Fungi
2.3. Microalgae
3. Marine Macro-Organisms
3.1. Macroalgae
3.2. Sponges
3.3. Mollusks, Cnidarians and Echinoderms
3.4. Tunicates

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Marine Organism | In Vitro/In Vivo | IC50/GI50/LC50 or Tested Concentration | Administration | Mechanism of Action | Ref. |
|---|---|---|---|---|---|---|
| Macroalgae | ||||||
| Ascophyllan | Ascophyllum Nodosum | In vivo mel animal model B16 | 25 mg/kg | Intraperitoneal Injection | Inhibition of matrix metallo-protease-9 | [69] |
| Spatane diterpinoids | Stoechospermum marginatum | In vitro on melanoma cell lines:B16F10 In vivo animal model C57BL/6 grafted with B16F10 melanoma cell line | IC50 3.95 μM 4, 10, 15 mg/Kg | In cell culture media Intraperitoneal injection | Apoptosis via activation of the caspase-mediated apoptotic pathway and PI3K/Akt pathway | [54] |
| Fucoidan CF | Chordaria flagelliformis | In vivo/ex vivo murine model grafted with B16 melanoma cell line | 0.01 mg/mouse | Intravenous injection | Stimulation of the innate immune system via CD11c integrins | [55] |
| Fucoxanthin containing extracts | Undaria pinnatifida | Melanoma cell line Malme-3M | IC50 (48 h) 27.96 ± 1.36 μM IC50 (72 h) 17.33 ± 2.65 μM | In cell culture media | N/A | [56] |
| Fucoxanthin (FX) | Fucus evanescens | Human melanoma SKMEL-28 cell line | IC50 114 μM | In cell culture media | Inhibition of the growth of human cell melanoma | [57] |
| Fucoxanthin | Ishige okamurae | B16F10 melanoma cell line | 30 μM | In cell culture media | CD44, CXCR4 and MMP9 reduction | [58] |
| L.s.-1.0 fr. (O-sulfated mannoglucuronofucans) L.s.-P fr. (sulfated polysaccharides) | Saccharina latissima | B6 mice inoculated with B16F10 melanoma cell line | 50 mg/kg | Intraperitoneal injection | Anti-angiogenesis | [59] |
| FSAR(fucoidanfr) FVES(fucoidan fr) Crude Fucoidan | Sargassum henslowianum Fucusvesiculosus | B16 melanoma cell line C57BL/6JJCL mice | 0.2–0.8 mg/mL 50 mg/kg body wt | In cell culture media In vivo injection | Apoptosis mediated by activation of caspase-3 | [49,60] |
| Polysaccharide fractions (SPPs) | Sargassum pallidum | B16 melanoma cell line | 25, 100, and 400 μg/mL | In cell culture media | immune stimulation | [61] |
| Sponges | ||||||
| Monanchocidin-A | Monanchora sp. | In vitro on melanoma cell lines: -LOX IMVI -MALME-3M -M14 -MDA-MB435 -SK-MEL-2 -SK-MEL-28 -SK-MEL-5 -UACC257 -UACC-62 | GI50 0.022 μM GI50 0.095 μM GI50 0.018 μM GI50 0.023 μM GI50 0.13 μM GI50 0.063 μM GI50 0.034 μM GI50 0.035 μM GI50 0.024 μM | In cell culture media | N/A | [62] |
| Bengamides | Myxococcus virescens | B16 melanoma murine model | 60 mg/kg | Mice injection | Inhibition of methionine amino peptidases [66] | [16] |
| Jaspine-B | Jaspis sp. | In vitro on melanoma cell lines: Human SK-Mel28; Murine B16 | IC50 0.5 μM | In cell culture media | Cell death via inhibition of sphingomyelin synthase | [67] |
| Halichondrin B | Halicondria okadai Lissodendoryx sp. | In vitro on B-16 melanoma cancer cells | IC50 0.09 ng/mL | In cell culture media | N/A | [70] |
| Arenosclerin-A Arenosclerin-C Haliclonacyclamine E | Arenosclera brasiliensis | In vitro on B16 melanoma cell line | 1.5–7.0 mg/mL | In cell culture media | N/A | [71] |
| Mollusks, Cnidarians and Ehinoderms | ||||||
| Lamellarin D | Lamellaria sp. | HBL skin melanoma cells | 5 μM | In cell culture media | Arresting cells in the G2 phase of the cellular cycle due to its effect on DNA Topoisomerase I | [73] |
| Metanolic, ex Diethyl ether ex n-hexane ex | Holothuria parva | In vitro/Ex vivo | 250, 500, and 1000 μg/mL | In cell culture media | Pro-apoptotic | [74] |
| Sarcophine (+)-7α,8β dihydroxydeepoxysarcophine | Sarcophyton glaucum | B16F10 melanoma cell line | 500 μM | In cell culture media | Inhibit DNA synthesis and PARP activity | [76] |
| Tunicates | ||||||
| Turgencin-A | Synoicum turgens | In vitro on melanoma cell lines: A2058 | IC50 1.4 μM | In cell culture media | N/A | [77] |
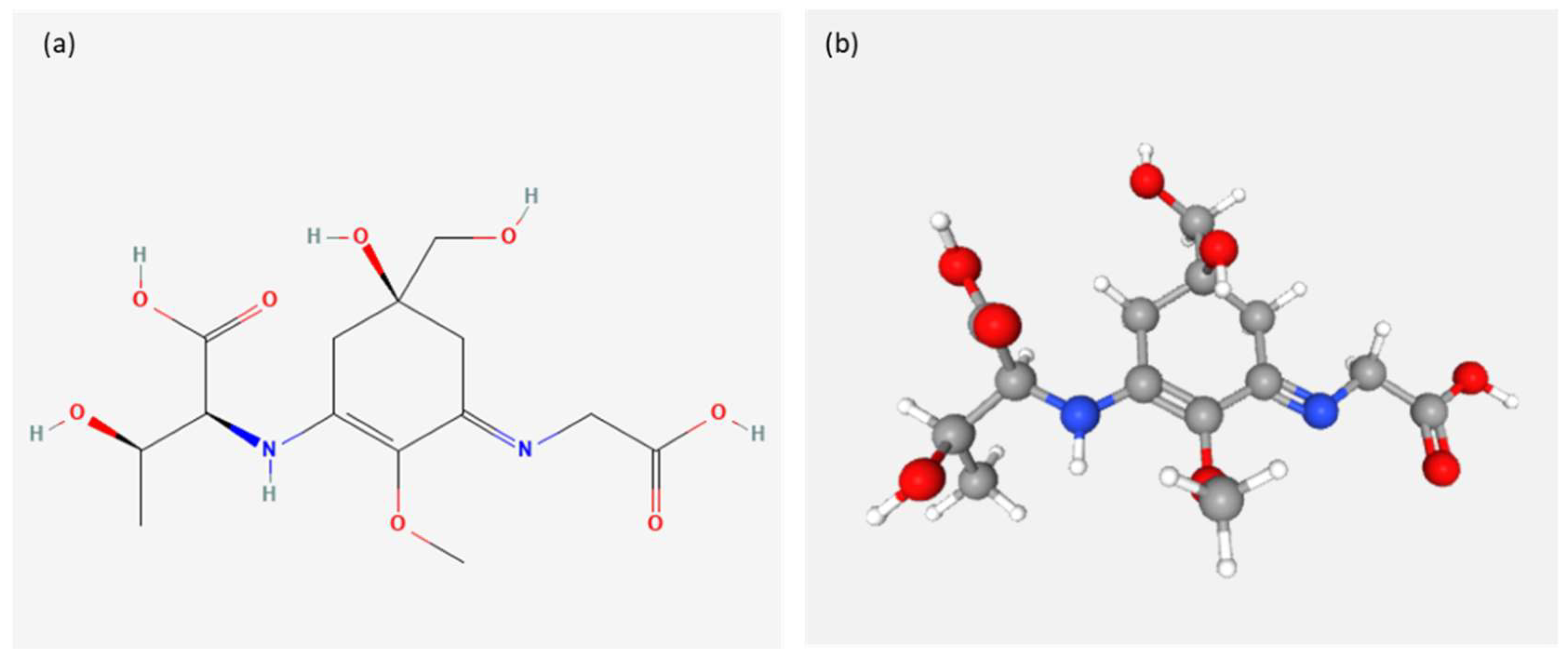
| Ecteinascidin-74 | Ecteinascidia turbinata | Ex vivo | q4d x 3—0.2, 0.1, 0.05 mg/kg | Intravenous | Double-strand breaks (DBSs) [84,85] | [80] |
| Palmerolide-A | Synoicum adareanum | In vitro on melanoma cell line: UACC-66 | LC50 0.018 μM | In cell culture media | Inhibition of V-ATPase | [81] |
| Thiaplidiaquinones A and B | Aplidium conicum | In Vitro on NCI panel | 10 μM | In cell culture media | Pro-apoptosis | [83] |
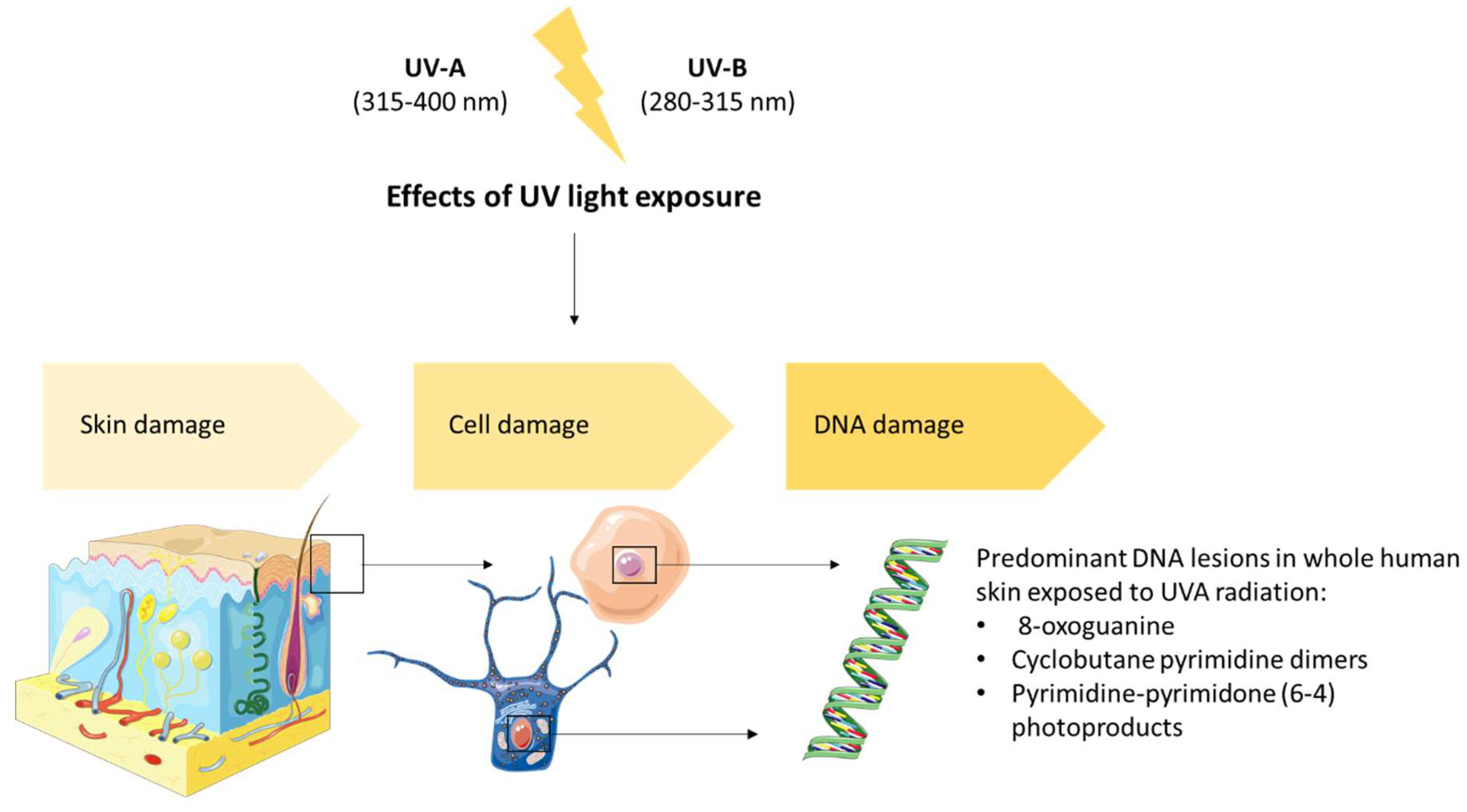
4. Prevention of Damage Induced by UV Solar Radiation
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schneider, S.H.; Mastrandrea, M.D. (Eds.) Encyclopedia of Climate and Weather, 2nd ed.; Oxford University Press: Oxford, UK; New York, NY, USA, 2011; ISBN 978-0-19-976532-4. [Google Scholar]
- Rastrelli, M.; Tropea, S.; Rossi, C.R.; Alaibac, M. Melanoma: Epidemiology, Risk Factors, Pathogenesis, Diagnosis and Classification. In Vivo 2014, 28, 1005–1011. [Google Scholar]
- Islami, F.; Sauer, A.G.; Miller, K.D.; Fedewa, S.A.; Minihan, A.K.; Geller, A.C.; Lichtenfeld, J.L.; Jemal, A. Cutaneous Melanomas Attributable to Ultraviolet Radiation Exposure by State. Int. J. Cancer 2020, 147, 1385–1390. [Google Scholar] [CrossRef]
- Landi, M.T.; Bishop, D.T.; MacGregor, S.; Machiela, M.J.; Stratigos, A.J.; Ghiorzo, P.; Brossard, M.; Calista, D.; Choi, J.; Fargnoli, M.C.; et al. Genome-Wide Association Meta-Analyses Combining Multiple Risk Phenotypes Provide Insights into the Genetic Architecture of Cutaneous Melanoma Susceptibility. Nat. Genet. 2020, 52, 494–504. [Google Scholar] [CrossRef]
- Elder, D.E.; Bastian, B.C.; Cree, I.A.; Massi, D.; Scolyer, R.A. The 2018 World Health Organization Classification of Cutaneous, Mucosal, and Uveal Melanoma: Detailed Analysis of 9 Distinct Subtypes Defined by Their Evolutionary Pathway. Arch. Pathol. Lab. Med. 2020, 144, 500–522. [Google Scholar] [CrossRef]
- Organisation Mondiale de la Santé; Centre International de Recherche sur le Cancer (Eds.) World health organization classification of tumours. In WHO Classification of Skin Tumours, 4th ed.; International Agency for Research on Cancer: Lyon, France, 2018; ISBN 978-92-832-2440-2. [Google Scholar]
- Duncan, L.M. The Classification of Cutaneous Melanoma. Hematol./Oncol. Clin. N. Am. 2009, 23, 501–513. [Google Scholar] [CrossRef] [PubMed]
- The International Agency for Research on Cancer Working Group on Artificial Ultraviolet (UV) Light and Skin Cancer. The Association of Use of Sunbeds with Cutaneous Malignant Melanoma and Other Skin Cancers: A Systematic Review. Int. J. Cancer 2006, 120, 1116–1122. [Google Scholar] [CrossRef]
- Stern, R.S. The Risk of Melanoma in Association with Long-Term Exposure to PUVA. J. Am. Acad. Dermatol. 2001, 44, 755–761. [Google Scholar] [CrossRef]
- Apalla, Z.; Lallas, A.; Sotiriou, E.; Lazaridou, E.; Ioannides, D. Epidemiological Trends in Skin Cancer. Dermatol. Pract. Concept. 2017, 7, 1–6. [Google Scholar] [CrossRef]
- Elwood, J.M.; Jopson, J. Melanoma and Sun Exposure: An Overview of Published Studies. Int. J. Cancer 1997, 73, 198–203. [Google Scholar] [CrossRef]
- Yang, T.; Yamada, K.; Zhou, T.; Harunari, E.; Igarashi, Y.; Terahara, T.; Kobayashi, T.; Imada, C. Akazamicin, a Cytotoxic Aromatic Polyketide from Marine-Derived Nonomuraea sp. J. Antibiot. 2019, 72, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Schneider, Y.K.-H.; Hansen, K.Ø.; Isaksson, J.; Ullsten, S.; Hansen, E.H.; Hammer Andersen, J. Anti-Bacterial Effect and Cytotoxicity Assessment of Lipid 430 Isolated from Algibacter sp. Molecules 2019, 24, 3991. [Google Scholar] [CrossRef] [Green Version]
- Harinantenaina Rakotondraibe, L.; Rasolomampianina, R.; Park, H.-Y.; Li, J.; Slebodnik, C.; Brodie, P.J.; Blasiak, L.C.; Hill, R.; TenDyke, K.; Shen, Y.; et al. Antiproliferative and Antiplasmodial Compounds from Selected Streptomyces Species. Bioorg. Med. Chem. Lett. 2015, 25, 5646–5649. [Google Scholar] [CrossRef]
- Weissman, K.J.; Müller, R. A Brief Tour of Myxobacterial Secondary Metabolism. Bioorg. Med. Chem. 2009, 17, 2121–2136. [Google Scholar] [CrossRef]
- Wenzel, S.C.; Hoffmann, H.; Zhang, J.; Debussche, L.; Haag-Richter, S.; Kurz, M.; Nardi, F.; Lukat, P.; Kochems, I.; Tietgen, H.; et al. Production of the Bengamide Class of Marine Natural Products in Myxobacteria: Biosynthesis and Structure–Activity Relationships. Angew. Chem. Int. Ed. 2015, 54, 15560–15564. [Google Scholar] [CrossRef] [PubMed]
- Dávila-Céspedes, A.; Hufendiek, P.; Crüsemann, M.; Schäberle, T.F.; König, G.M. Marine-Derived Myxobacteria of the Suborder Nannocystineae: An Underexplored Source of Structurally Intriguing and Biologically Active Metabolites. Beilstein J. Org. Chem. 2016, 12, 969–984. [Google Scholar] [CrossRef]
- Tomura, T.; Nagashima, S.; Yamazaki, S.; Iizuka, T.; Fudou, R.; Ojika, M. An Unusual Diterpene—Enhygromic Acid and Deoxyenhygrolides from a Marine Myxobacterium, Enhygromyxa sp. Mar. Drugs 2017, 15, 109. [Google Scholar] [CrossRef]
- Patil, S.; Paradeshi, J.; Chaudhari, B. Anti-Melanoma and UV-B Protective Effect of Microbial Pigment Produced by Marine Pseudomonas aeruginosa GS-33. Nat. Prod. Res. 2016, 30, 2835–2839. [Google Scholar] [CrossRef] [PubMed]
- Tabassum, S.; Khan, R.A.; Arjmand, F.; Sen, S.; Kayal, J.; Juvekar, A.S.; Zingde, S.M. Synthesis and Characterization of Glycoconjugate Tin(IV) Complexes: In Vitro DNA Binding Studies, Cytotoxicity, and Cell Death. J. Organomet. Chem. 2011, 696, 1600–1608. [Google Scholar] [CrossRef]
- Kristoffersen, V.; Jenssen, M.; Jawad, H.R.; Isaksson, J.; Hansen, E.H.; Rämä, T.; Hansen, K.Ø.; Andersen, J.H. Two Novel Lyso-Ornithine Lipids Isolated from an Arctic Marine Lacinutrix sp. Bacterium. Molecules 2021, 26, 5295. [Google Scholar] [CrossRef]
- Zhang, G.; Liu, S.; Liu, Y.; Wang, F.; Ren, J.; Gu, J.; Zhou, K.; Shan, B. A Novel Cyclic Pentapeptide, H-10, Inhibits B16 Cancer Cell Growth and Induces Cell Apoptosis. Oncol. Lett. 2014, 8, 248–252. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.-Y.; Zhou, T.; Zhao, Y.-Y.; Chen, L.; Gong, M.-W.; Xia, Q.-W.; Ying, M.-G.; Zheng, Q.-H.; Zhang, Q.-Q. Antitumor Effects and Related Mechanisms of Penicitrinine A, a Novel Alkaloid with a Unique Spiro Skeleton from the Marine Fungus Penicillium citrinum. Mar. Drugs 2015, 13, 4733–4753. [Google Scholar] [CrossRef]
- Jenssen, M.; Kristoffersen, V.; Motiram-Corral, K.; Isaksson, J.; Rämä, T.; Andersen, J.H.; Hansen, E.H.; Hansen, K.Ø. Chlovalicin B, a Chlorinated Sesquiterpene Isolated from the Marine Mushroom Digitatispora marina. Molecules 2021, 26, 7560. [Google Scholar] [CrossRef]
- Liu, Y.; Sheikh, M.S. Melanoma: Molecular Pathogenesis and Therapeutic Management. Mol. Cell. Pharmacol. 2014, 6, 228. [Google Scholar] [PubMed]
- Jenssen, M.; Rainsford, P.; Juskewitz, E.; Andersen, J.H.; Hansen, E.H.; Isaksson, J.; Rämä, T.; Hansen, K.Ø. Lulworthinone, a New Dimeric Naphthopyrone From a Marine Fungus in the Family Lulworthiaceae with Antibacterial Activity against Clinical Methicillin-Resistant Staphylococcus aureus Isolates. Front. Microbiol. 2021, 12, 730740. [Google Scholar] [CrossRef] [PubMed]
- Fan, B.; Dewapriya, P.; Li, F.; Blümel, M.; Tasdemir, D. Pyrenosetins A–C, New Decalinoylspirotetramic Acid Derivatives Isolated by Bioactivity-Based Molecular Networking from the Seaweed-Derived Fungus Pyrenochaetopsis sp. FVE-001. Mar. Drugs 2020, 18, 47. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Kusari, S.; Spiteller, M. Natural Products Containing ‘Decalin’ Motif in Microorganisms. Nat. Prod. Rep. 2014, 31, 1175–1201. [Google Scholar] [CrossRef] [PubMed]
- Jabeen, A.; Reeder, B.; Hisaindee, S.; Ashraf, S.; Darmaki, N.A.; Battah, S.; Al-Zuhair, S. Effect of Enzymatic Pre-Treatment of Microalgae Extracts on Their Anti-Tumor Activity. Biomed. J. 2017, 40, 339–346. [Google Scholar] [CrossRef]
- Lauritano, C.; Andersen, J.H.; Hansen, E.; Albrigtsen, M.; Escalera, L.; Esposito, F.; Helland, K.; Hanssen, K.Ø.; Romano, G.; Ianora, A. Bioactivity Screening of Microalgae for Antioxidant, Anti-Inflammatory, Anticancer, Anti-Diabetes, and Antibacterial Activities. Front. Mar. Sci. 2016, 3, 68. [Google Scholar] [CrossRef]
- Riccio, G.; Martinez, K.A.; Ianora, A.; Lauritano, C. De Novo Transcriptome of the Flagellate Isochrysis galbana Identifies Genes Involved in the Metabolism of Antiproliferative Metabolites. Biology 2022, 11, 771. [Google Scholar] [CrossRef]
- Martínez, K.A.; Saide, A.; Crespo, G.; Martín, J.; Romano, G.; Reyes, F.; Lauritano, C.; Ianora, A. Promising Antiproliferative Compound from the Green Microalga Dunaliella tertiolecta against Human Cancer Cells. Front. Mar. Sci. 2022, 9, 778108. [Google Scholar] [CrossRef]
- Lauritano, C.; Helland, K.; Riccio, G.; Andersen, J.H.; Ianora, A.; Hansen, E.H. Lysophosphatidylcholines and Chlorophyll-Derived Molecules from the Diatom Cylindrotheca closterium with Anti-Inflammatory Activity. Mar. Drugs 2020, 18, 166. [Google Scholar] [CrossRef] [Green Version]
- Pohl, C.; Kock, J. Oxidized Fatty Acids as Inter-Kingdom Signaling Molecules. Molecules 2014, 19, 1273–1285. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, J.; Kubota, T. Bioactive Metabolites from Marine Dinoflagellates. In Comprehensive Natural Products II; Elsevier: Amsterdam, The Netherlands, 2010; pp. 263–325. ISBN 978-0-08-045382-8. [Google Scholar]
- Martínez, K.A.; Lauritano, C.; Druka, D.; Romano, G.; Grohmann, T.; Jaspars, M.; Martín, J.; Díaz, C.; Cautain, B.; de la Cruz, M.; et al. Amphidinol 22, a New Cytotoxic and Antifungal Amphidinol from the Dinoflagellate Amphidinium carterae. Mar. Drugs 2019, 17, 385. [Google Scholar] [CrossRef]
- Kobayashi, J.; Shigemori, H.; Ishibashi, M.; Yamasu, T.; Hirota, H.; Sasaki, T. Amphidinolides G and H: New Potent Cytotoxic Macrolides from the Cultured Symbiotic Dinoflagellate Amphidinium sp. J. Org. Chem. 1991, 56, 5221–5224. [Google Scholar] [CrossRef]
- Nuzzo, G.; Gomes, B.; Gallo, C.; Amodeo, P.; Sansone, C.; Pessoa, O.; Manzo, E.; Vitale, R.; Ianora, A.; Santos, E.; et al. Potent Cytotoxic Analogs of Amphidinolides from the Atlantic Octocoral Stragulum bicolor. Mar. Drugs 2019, 17, 58. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, J.; Ishibashi, M. Bioactive Metabolites of Symbiotic Marine Microorganisms. Chem. Rev. 1993, 93, 1753–1769. [Google Scholar] [CrossRef]
- Ávila-Román, J.; García-Gil, S.; Rodríguez-Luna, A.; Motilva, V.; Talero, E. Anti-Inflammatory and Anticancer Effects of Microalgal Carotenoids. Mar. Drugs 2021, 19, 531. [Google Scholar] [CrossRef]
- Kobayashi, J.; Tsuda, M. Amphidinolides, Bioactive Macrolides from Symbiotic Marine Dinoflagellates. Nat. Prod. Rep. 2004, 21, 77. [Google Scholar] [CrossRef] [PubMed]
- Sushytskyi, L.; Lukáč, P.; Synytsya, A.; Bleha, R.; Rajsiglová, L.; Capek, P.; Pohl, R.; Vannucci, L.; Čopíková, J.; Kaštánek, P. Immunoactive Polysaccharides Produced by Heterotrophic Mutant of Green Microalga Parachlorella kessleri HY1 (Chlorellaceae). Carbohydr. Polym. 2020, 246, 116588. [Google Scholar] [CrossRef]
- Bergé, J.P.; Debiton, E.; Dumay, J.; Durand, P.; Barthomeuf, C. In Vitro Anti-Inflammatory and Anti-Proliferative Activity of Sulfolipids from the Red Alga Porphyridium cruentum. J. Agric. Food Chem. 2002, 50, 6227–6232. [Google Scholar] [CrossRef] [PubMed]
- Carpi, S.; Polini, B.; Poli, G.; Alcantara Barata, G.; Fogli, S.; Romanini, A.; Tuccinardi, T.; Guella, G.; Frontini, F.; Nieri, P.; et al. Anticancer Activity of Euplotin C, Isolated from the Marine Ciliate Euplotes crassus, against Human Melanoma Cells. Mar. Drugs 2018, 16, 166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steelman, L.S.; Chappell, W.H.; Abrams, S.L.; Kempf, C.R.; Long, J.; Laidler, P.; Mijatovic, S.; Maksimovic-Ivanic, D.; Stivala, F.; Mazzarino, M.C.; et al. Roles of the Raf/MEK/ERK and PI3K/PTEN/Akt/MTOR Pathways in Controlling Growth and Sensitivity to Therapy-Implications for Cancer and Aging. Aging 2011, 3, 192–222. [Google Scholar] [CrossRef] [PubMed]
- Yajima, I.; Kumasaka, M.Y.; Thang, N.D.; Goto, Y.; Takeda, K.; Yamanoshita, O.; Iida, M.; Ohgami, N.; Tamura, H.; Kawamoto, Y.; et al. RAS/RAF/MEK/ERK and PI3K/PTEN/AKT Signaling in Malignant Melanoma Progression and Therapy. Dermatol. Res. Pract. 2012, 2012, 354191. [Google Scholar] [CrossRef]
- Carpi, S.; Kawahigashi, Y.; Longo, R.; Weiner, M. From Vertex Operator Algebras to Conformal Nets and Back. Mem. AMS 2018, 254, 1213. [Google Scholar] [CrossRef]
- Cervia, D.; Martini, D.; Garcia-Gil, M.; Di Giuseppe, G.; Guella, G.; Dini, F.; Bagnoli, P. Cytotoxic Effects and Apoptotic Signalling Mechanisms of the Sesquiterpenoid Euplotin C, a Secondary Metabolite of the Marine Ciliate Euplotes crassus, in Tumour Cells. Apoptosis 2006, 11, 829–843. [Google Scholar] [CrossRef]
- Ale, M.T.; Maruyama, H.; Tamauchi, H.; Mikkelsen, J.D.; Meyer, A.S. Fucose-Containing Sulfated Polysaccharides from Brown Seaweeds Inhibit Proliferation of Melanoma Cells and Induce Apoptosis by Activation of Caspase-3 In Vitro. Mar. Drugs 2011, 9, 2605–2621. [Google Scholar] [CrossRef]
- Grether-Beck, S.; Mühlberg, K.; Brenden, H.; Felsner, I.; Brynjólfsdóttir, Á.; Einarsson, S.; Krutmann, J. Bioactive Molecules from the Blue Lagoon: In Vitro and In Vivo Assessment of Silica Mud and Microalgae Extracts for Their Effects on Skin Barrier Function and Prevention of Skin Ageing. Exp. Dermatol. 2008, 17, 771–779. [Google Scholar] [CrossRef]
- Kim, K.-N.; Ahn, G.; Heo, S.-J.; Kang, S.-M.; Kang, M.-C.; Yang, H.-M.; Kim, D.; Roh, S.W.; Kim, S.-K.; Jeon, B.-T.; et al. Inhibition of Tumor Growth In Vitro and In Vivo by Fucoxanthin against Melanoma B16F10 Cells. Environ. Toxicol. Pharmacol. 2013, 35, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Ávila-Román, J.; Talero, E.; de Los Reyes, C.; Zubía, E.; Motilva, V.; García-Mauriño, S. Cytotoxic Activity of Microalgal-Derived Oxylipins against Human Cancer Cell Lines and Their Impact on ATP Levels. Nat. Prod. Commun. 2016, 11, 1871–1875. [Google Scholar] [CrossRef] [PubMed]
- Chinnababu, B.; Purushotham Reddy, S.; Sankara Rao, P.; Loka Reddy, V.; Sudheer Kumar, B.; Rao, J.V.; Prakasham, R.S.; Suresh Babu, K. Isolation, Semi-Synthesis and Bio-Evaluation of Spatane Derivatives from the Brown Algae Stoechospermum marginatum. Bioorg. Med. Chem. Lett. 2015, 25, 2479–2483. [Google Scholar] [CrossRef]
- Velatooru, L.R.; Baggu, C.B.; Janapala, V.R. Spatane Diterpinoid from the Brown Algae, Stoechospermum marginatum Induces Apoptosis via ROS Induced Mitochondrial Mediated Caspase Dependent Pathway in Murine B16F10 Melanoma Cells: Spatane diterpinoid induces apoptosis. Mol. Carcinog. 2016, 55, 2222–2235. [Google Scholar] [CrossRef] [PubMed]
- Anisimova, N.Y.; Ustyuzhanina, N.E.; Donenko, F.V.; Bilan, M.I.; Ushakova, N.A.; Usov, A.I.; Nifantiev, N.E.; Kiselevskiy, M.V. Influence of Fucoidans and Their Derivatives on Antitumor and Phagocytic Activity of Human Blood Leucocytes. Biochem. Mosc. 2015, 80, 925–933. [Google Scholar] [CrossRef]
- Wang, S.; Li, Y.; White, W.; Lu, J. Extracts from New Zealand Undaria pinnatifida Containing Fucoxanthin as Potential Functional Biomaterials against Cancer In Vitro. J. Funct. Biomater. 2014, 5, 29–42. [Google Scholar] [CrossRef]
- Imbs, T.I.; Ermakova, S.P.; Fedoreyev, S.A.; Anastyuk, S.D.; Zvyagintseva, T.N. Isolation of Fucoxanthin and Highly Unsaturated Monogalactosyldiacylglycerol from Brown Alga Fucus evanescens C Agardh and In Vitro Investigation of Their Antitumor Activity. Mar. Biotechnol. 2013, 15, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Chung, T.-W.; Choi, H.-J.; Lee, J.-Y.; Jeong, H.-S.; Kim, C.-H.; Joo, M.; Choi, J.-Y.; Han, C.-W.; Kim, S.-Y.; Choi, J.-S.; et al. Marine Algal Fucoxanthin Inhibits the Metastatic Potential of Cancer Cells. Biochem. Biophys. Res. Commun. 2013, 439, 580–585. [Google Scholar] [CrossRef] [PubMed]
- Croci, D.O.; Cumashi, A.; Ushakova, N.A.; Preobrazhenskaya, M.E.; Piccoli, A.; Totani, L.; Ustyuzhanina, N.E.; Bilan, M.I.; Usov, A.I.; Grachev, A.A.; et al. Fucans, but Not Fucomannoglucuronans, Determine the Biological Activities of Sulfated Polysaccharides from Laminaria saccharina Brown Seaweed. PLoS ONE 2011, 6, e17283. [Google Scholar] [CrossRef]
- Ale, M.T.; Maruyama, H.; Tamauchi, H.; Mikkelsen, J.D.; Meyer, A.S. Fucoidan from Sargassum sp. and Fucus vesiculosus Reduces Cell Viability of Lung Carcinoma and Melanoma Cells In Vitro and Activates Natural Killer Cells in Mice In Vivo. Int. J. Biol. Macromol. 2011, 49, 331–336. [Google Scholar] [CrossRef]
- Gao, Y.; Li, Y.; Niu, Y.; Ju, H.; Chen, R.; Li, B.; Song, X.; Song, L. Chemical Characterization, Antitumor, and Immune-Enhancing Activities of Polysaccharide from Sargassum pallidum. Molecules 2021, 26, 7559. [Google Scholar] [CrossRef] [PubMed]
- Gogineni, V.; Oh, J.; Waters, A.L.; Kelly, M.; Stone, R.; Hamann, M.T. Monanchocidin A from Subarctic Sponges of the Genus Monanchora and Their Promising Selectivity against Melanoma in Vitro. Front. Mar. Sci. 2020, 7, 58. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the 30 Years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef] [PubMed]
- Bauer, A.; Brönstrup, M. Industrial Natural Product Chemistry for Drug Discovery and Development. Nat. Prod. Rep. 2014, 31, 35–60. [Google Scholar] [CrossRef]
- Butler, M.S.; Robertson, A.A.B.; Cooper, M.A. Natural Product and Natural Product Derived Drugs in Clinical Trials. Nat. Prod. Rep. 2014, 31, 1612–1661. [Google Scholar] [CrossRef] [PubMed]
- Towbin, H.; Bair, K.W.; DeCaprio, J.A.; Eck, M.J.; Kim, S.; Kinder, F.R.; Morollo, A.; Mueller, D.R.; Schindler, P.; Song, H.K.; et al. Proteomics-Based Target Identification. J. Biol. Chem. 2003, 278, 52964–52971. [Google Scholar] [CrossRef]
- Salma, Y.; Lafont, E.; Therville, N.; Carpentier, S.; Bonnafé, M.-J.; Levade, T.; Génisson, Y.; Andrieu-Abadie, N. The Natural Marine Anhydrophytosphingosine, Jaspine B, Induces Apoptosis in Melanoma Cells by Interfering with Ceramide Metabolism. Biochem. Pharmacol. 2009, 78, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Okimura, T.; Yamaguchi, K.; Oda, T. The Potent Activity of Sulfated Polysaccharide, Ascophyllan, Isolated from Ascophyllum nodosum to Induce Nitric Oxide and Cytokine Production from Mouse Macrophage RAW264.7 Cells: Comparison between Ascophyllan and Fucoidan. Nitric Oxide 2011, 25, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Abu, R.; Jiang, Z.; Ueno, M.; Isaka, S.; Nakazono, S.; Okimura, T.; Cho, K.; Yamaguchi, K.; Kim, D.; Oda, T. Anti-Metastatic Effects of the Sulfated Polysaccharide Ascophyllan Isolated from Ascophyllum nodosum on B16 Melanoma. Biochem. Biophys. Res. Commun. 2015, 458, 727–732. [Google Scholar] [CrossRef]
- Hirata, Y.; Uemura, D. Halichondrins—Antitumor Polyether Macrolides from a Marine Sponge. Pure Appl. Chem. 1986, 58, 701–710. [Google Scholar] [CrossRef]
- Torres, Y.R.; Berlinck, R.G.S.; Nascimento, G.G.F.; Fortier, S.C.; Pessoa, C.; de Moraes, M.O. Antibacterial Activity against Resistant Bacteria and Cytotoxicity of Four Alkaloid Toxins Isolated from the Marine Sponge Arenosclera brasiliensis. Toxicon 2002, 40, 885–891. [Google Scholar] [CrossRef]
- Andersen, R.J.; Faulkner, D.J.; He, C.H.; Van Duyne, G.D.; Clardy, J. Metabolites of the Marine Prosobranch Mollusk Lamellaria sp. J. Am. Chem. Soc. 1985, 107, 5492–5495. [Google Scholar] [CrossRef]
- Ballot, C.; Martoriati, A.; Jendoubi, M.; Buche, S.; Formstecher, P.; Mortier, L.; Kluza, J.; Marchetti, P. Another Facet to the Anticancer Response to Lamellarin D: Induction of Cellular Senescence through Inhibition of Topoisomerase I and Intracellular Ros Production. Mar. Drugs 2014, 12, 779–798. [Google Scholar] [CrossRef]
- Arast, Y.; Seyed Razi, N.; Nazemi, M.; Seydi, E.; Pourahmad, J. Non-Polar Compounds of Persian Gulf Sea Cucumber Holothuria parva Selectively Induce Toxicity on Skin Mitochondria Isolated from Animal Model of Melanoma. Cutan. Ocul. Toxicol. 2018, 37, 218–227. [Google Scholar] [CrossRef]
- Grossman, D.; Altieri, D.C. Drug Resistance in Melanoma: Mechanisms, Apoptosis, and New Potential Therapeutic Targets. Cancer Metastasis Rev. 2001, 20, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Szymanski, P.T.; Ahmed, S.A.; Radwan, M.M.; Khalifa, S.I.; Fahmy, H. Evaluation of the Anti-Melanoma Activities of Sarcophine, (+)-7α,8β-Dihydroxydeepoxysarcophine and Sarcophytolide from the Red Sea Soft Coral Sarcophyton glaucum. Nat. Prod. Commun. 2014, 9, 1934578X1400900. [Google Scholar] [CrossRef]
- Hansen, I.K.Ø.; Isaksson, J.; Poth, A.G.; Hansen, K.Ø.; Andersen, A.J.C.; Richard, C.S.M.; Blencke, H.-M.; Stensvåg, K.; Craik, D.J.; Haug, T. Isolation and Characterization of Antimicrobial Peptides with Unusual Disulfide Connectivity from the Colonial Ascidian Synoicum turgens. Mar. Drugs 2020, 18, 51. [Google Scholar] [CrossRef] [PubMed]
- Rinehart, K.L.; Holt, T.G.; Fregeau, N.L.; Stroh, J.G.; Keifer, P.A.; Sun, F.; Li, L.H.; Martin, D.G. Ecteinascidins 729, 743, 745, 759A, 759B, and 770: Potent Antitumor Agents from the Caribbean Tunicate Ecteinascidia turbinata. J. Org. Chem. 1990, 55, 4512–4515. [Google Scholar] [CrossRef]
- Jimeno, J.; Faircloth, G.; Sousa-Faro, J.M.F.; Scheuer, P.; Rinehart, K. New Marine Derived Anticancer Therapeutics—A Journey from the Sea to Clinical Trials. Mar. Drugs 2004, 2, 14–29. [Google Scholar] [CrossRef]
- Hendriks, H.R.; Fiebig, H.H.; Giavazzi, R.; Langdon, S.P.; Jimeno, J.M.; Faircloth, G.T. High Antitumour Activity of ET743 against Human Tumour Xenografts from Melanoma, Non-Small-Cell Lung and Ovarian Cancer. Ann. Oncol. 1999, 10, 1233–1240. [Google Scholar] [CrossRef]
- Diyabalanage, T.; Amsler, C.D.; McClintock, J.B.; Baker, B.J. Palmerolide A, a Cytotoxic Macrolide from the Antarctic Tunicate Synoicum adareanum. J. Am. Chem. Soc. 2006, 128, 5630–5631. [Google Scholar] [CrossRef]
- Murray, A.E.; Avalon, N.E.; Bishop, L.; Davenport, K.W.; Delage, E.; Dichosa, A.E.K.; Eveillard, D.; Higham, M.L.; Kokkaliari, S.; Lo, C.-C.; et al. Uncovering the Core Microbiome and Distribution of Palmerolide in Synoicum adareanum Across the Anvers Island Archipelago, Antarctica. Mar. Drugs 2020, 18, 298. [Google Scholar] [CrossRef] [PubMed]
- Harper, J.; Khalil, I.; Shaw, L.; Bourguet-Kondracki, M.-L.; Dubois, J.; Valentin, A.; Barker, D.; Copp, B. Structure-Activity Relationships of the Bioactive Thiazinoquinone Marine Natural Products Thiaplidiaquinones A and B. Mar. Drugs 2015, 13, 5102–5110. [Google Scholar] [CrossRef]
- Pommier, Y.; Kohlhagen, G.; Bailly, C.; Waring, M.; Mazumder, A.; Kohn, K.W. DNA Sequence- and Structure-Selective Alkylation of Guanine N2 in the DNA Minor Groove by Ecteinascidin 743, a Potent Antitumor Compound from the Caribbean Tunicate Ecteinascidia turbinata. Biochemistry 1996, 35, 13303–13309. [Google Scholar] [CrossRef] [PubMed]
- Simoens, C.; Korst, A.E.C.; De Pooter, C.M.J.; Lambrechts, H.A.J.; Pattyn, G.G.O.; Faircloth, G.T.; Lardon, F.; Vermorken, J.B. In Vitro Interaction between Ecteinascidin 743 (ET-743) and Radiation, in Relation to Its Cell Cycle Effects. Br. J. Cancer 2003, 89, 2305–2311. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Kim, A.L.; Feith, D.J.; Pegg, A.E.; Russo, J.; Zhang, H.; Aszterbaum, M.; Kopelovich, L.; Epstein, E.H.; Bickers, D.R.; et al. Ornithine Decarboxylase Is a Target for Chemoprevention of Basal and Squamous Cell Carcinomas in Ptch1+/− Mice. J. Clin. Investig. 2004, 113, 867–875. [Google Scholar] [CrossRef]
- Catanzaro, E.; Bishayee, A.; Fimognari, C. On a Beam of Light: Photoprotective Activities of the Marine Carotenoids Astaxanthin and Fucoxanthin in Suppression of Inflammation and Cancer. Mar. Drugs 2020, 18, 544. [Google Scholar] [CrossRef] [PubMed]
- Starcevic, A.; Akthar, S.; Dunlap, W.C.; Shick, J.M.; Hranueli, D.; Cullum, J.; Long, P.F. Enzymes of the Shikimic Acid Pathway Encoded in the Genome of a Basal Metazoan, Nematostella vectensis, Have Microbial Origins. Proc. Natl. Acad. Sci. USA 2008, 105, 2533–2537. [Google Scholar] [CrossRef] [PubMed]
- Banaszak, A.T.; Barba Santos, M.G.; LaJeunesse, T.C.; Lesser, M.P. The Distribution of Mycosporine-like Amino Acids (MAAs) and the Phylogenetic Identity of Symbiotic Dinoflagellates in Cnidarian Hosts from the Mexican Caribbean. J. Exp. Mar. Biol. Ecol. 2006, 337, 131–146. [Google Scholar] [CrossRef]
- Bandaranayake, W.M. Mycosporines: Are They Nature’s Sunscreens? Nat. Prod. Rep. 1998, 15, 159. [Google Scholar] [CrossRef]
- Shibata, K. Pigments and a UV-Absorbing Substance in Corals and a Blue-Green Alga Living in the Great Barrier Reef. Plant Cell Physiol. 1969, 10, 325–335. [Google Scholar] [CrossRef]
- Wada, N.; Sakamoto, T.; Matsugo, S. Mycosporine-Like Amino Acids and Their Derivatives as Natural Antioxidants. Antioxidants 2015, 4, 603–646. [Google Scholar] [CrossRef]
- Bernillon, J.; Bouillant, M.-L.; Pittet, J.-L.; Favre-Bonvin, J.; Arpin, N. Mycosporine Glutamine and Related Mycosporines in the Fungus Pyronema omphalodes. Phytochemistry 1984, 23, 1083–1087. [Google Scholar] [CrossRef]
- Llewellyn, C.A.; Airs, R.L. Distribution and Abundance of MAAs in 33 Species of Microalgae across 13 Classes. Mar. Drugs 2010, 8, 1273–1291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michalek-Wagner, K. Seasonal and Sex-Specific Variations in Levels of Photo-Protecting Mycosporine-like Amino Acids (MAAs) in Soft Corals. Mar. Biol. 2001, 139, 651–660. [Google Scholar] [CrossRef]
- Chrapusta, E.; Kaminski, A.; Duchnik, K.; Bober, B.; Adamski, M.; Bialczyk, J. Mycosporine-Like Amino Acids: Potential Health and Beauty Ingredients. Mar. Drugs 2017, 15, 326. [Google Scholar] [CrossRef]
- Cardozo, K.H.M.; Guaratini, T.; Barros, M.P.; Falcão, V.R.; Tonon, A.P.; Lopes, N.P.; Campos, S.; Torres, M.A.; Souza, A.O.; Colepicolo, P.; et al. Metabolites from Algae with Economical Impact. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2007, 146, 60–78. [Google Scholar] [CrossRef]
- Stoyneva-Gärtner, M.; Uzunov, B.; Gärtner, G. Enigmatic Microalgae from Aeroterrestrial and Extreme Habitats in Cosmetics: The Potential of the Untapped Natural Sources. Cosmetics 2020, 7, 27. [Google Scholar] [CrossRef]
- Singh, A.; Čížková, M.; Bišová, K.; Vítová, M. Exploring Mycosporine-Like Amino Acids (MAAs) as Safe and Natural Protective Agents against UV-Induced Skin Damage. Antioxidants 2021, 10, 683. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. PubChem Compound Summary for CID 6857486, Porphyra-334. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/porphyra-334 (accessed on 13 July 2022).
- Rastogi, R.P.; Sinha, R.P. Biotechnological and Industrial Significance of Cyanobacterial Secondary Metabolites. Biotechnol. Adv. 2009, 27, 521–539. [Google Scholar] [CrossRef]
- Siezen, R.J. Microbial Sunscreens: Genomics Update. Microb. Biotechnol. 2011, 4, 1–7. [Google Scholar] [CrossRef]
- Mourelle, M.; Gómez, C.; Legido, J. The Potential Use of Marine Microalgae and Cyanobacteria in Cosmetics and Thalassotherapy. Cosmetics 2017, 4, 46. [Google Scholar] [CrossRef]
- Matsui, K.; Nazifi, E.; Hirai, Y.; Wada, N.; Matsugo, S.; Sakamoto, T. The Cyanobacterial UV-Absorbing Pigment Scytonemin Displays Radical-Scavenging Activity. J. Gen. Appl. Microbiol. 2012, 58, 137–144. [Google Scholar] [CrossRef]
- De Freitas Coêlho, D.; Tundisi, L.L.; Cerqueira, K.S.; da Silva Rodrigues, J.R.; Mazzola, P.G.; Tambourgi, E.B.; de Souza, R.R. Microalgae: Cultivation Aspects and Bioactive Compounds. Braz. Arch. Biol. Technol. 2019, 62, e19180343. [Google Scholar] [CrossRef]
- Stevenson, C.S.; Capper, E.A.; Roshak, A.K.; Marquez, B.; Eichman, C.; Jackson, J.R.; Mattern, M.; Gerwick, W.H.; Jacobs, R.S.; Marshall, L.A. The Identification and Characterization of the Marine Natural Product Scytonemin as a Novel Antiproliferative Pharmacophore. J. Pharmacol. Exp. Ther. 2002, 303, 858–866. [Google Scholar] [CrossRef]
- Jahan, A.; Ahmad, I.Z.; Fatima, N.; Ansari, V.A.; Akhtar, J. Algal Bioactive Compounds in the Cosmeceutical Industry: A Review. Phycologia 2017, 56, 410–422. [Google Scholar] [CrossRef]
- Alparslan, L.; Şekeroğlu, N.; Kijjoa, A. The Potential of Marine Resources in Cosmetics. Curr. Perspect. Med. Aromat. Plants (CUPMAP) 2018, 1, 53–66. [Google Scholar] [CrossRef]
- Stoyneva-Gärtner, M.; Stoykova, P.; Uzunov, B.; Dincheva, I.; Atanassov, I.; Draganova, P.; Borisova, C.; Gärtner, G. Carotenoids in Five Aeroterrestrial Strains from Vischeria/Eustigmatos Group: Updating the Pigment Pattern of Eustigmatophyceae. Biotechnol. Biotechnol. Equip. 2019, 33, 250–267. [Google Scholar] [CrossRef]
- Galasso, C.; Corinaldesi, C.; Sansone, C. Carotenoids from Marine Organisms: Biological Functions and Industrial Applications. Antioxidants 2017, 6, 96. [Google Scholar] [CrossRef] [PubMed]
- Sathasivam, R.; Ki, J.-S. A Review of the Biological Activities of Microalgal Carotenoids and Their Potential Use in Healthcare and Cosmetic Industries. Mar. Drugs 2018, 16, 26. [Google Scholar] [CrossRef] [PubMed]
- Thomas, N.; Kim, S.-K. Beneficial Effects of Marine Algal Compounds in Cosmeceuticals. Mar. Drugs 2013, 11, 146–164. [Google Scholar] [CrossRef] [PubMed]
- Wijesinghe, W.A.J.P.; Jeon, Y.-J. Biological Activities and Potential Cosmeceutical Applications of Bioactive Components from Brown Seaweeds: A Review. Phytochem. Rev. 2011, 10, 431–443. [Google Scholar] [CrossRef]
- Bastonini, E.; Kovacs, D.; Picardo, M. Skin Pigmentation and Pigmentary Disorders: Focus on Epidermal/Dermal Cross-Talk. Ann. Dermatol. 2016, 28, 279. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, H.; Tanaka, J.; Shan, S.-J.; Maoka, T. Anti-Pigmentary Activity of Fucoxanthin and Its Influence on Skin MRNA Expression of Melanogenic Molecules. J. Pharm. Pharmacol. 2010, 62, 1137–1145. [Google Scholar] [CrossRef] [PubMed]
- Matsui, M.; Tanaka, K.; Higashiguchi, N.; Okawa, H.; Yamada, Y.; Tanaka, K.; Taira, S.; Aoyama, T.; Takanishi, M.; Natsume, C.; et al. Protective and Therapeutic Effects of Fucoxanthin against Sunburn Caused by UV Irradiation. J. Pharmacol. Sci. 2016, 132, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Tavares, R.S.N.; Kawakami, C.M.; de Castro Pereira, K.; do Amaral, G.T.; Benevenuto, C.G.; Maria-Engler, S.S.; Colepicolo, P.; Debonsi, H.M.; Gaspar, L.R. Fucoxanthin for Topical Administration, a Phototoxic vs. Photoprotective Potential in a Tiered Strategy Assessed by In Vitro Methods. Antioxidants 2020, 9, 328. [Google Scholar] [CrossRef]
- Fryer, M.J. Evidence for the photoprotective effects of vitamin E. Photochem. Photobiol. 1993, 58, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Barthelman, M.; Martinez, J.; Alberts, D.; Gensler, H.L. Inhibition of Cyclobutane Pyrimidine Dimer Formation in Epidermal P53 Gene of UV-irradiated Mice by A-tocopherol. Nutr. Cancer 1997, 29, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.D.; Meeran, S.M.; Katiyar, S.K. Dietary Grape Seed Proanthocyanidins Inhibit UVB-Induced Oxidative Stress and Activation of Mitogen-Activated Protein Kinases and Nuclear Factor-ΚB Signaling in in Vivo SKH-1 Hairless Mice. Mol. Cancer Ther. 2007, 6, 995–1005. [Google Scholar] [CrossRef]
- Xing, Y.-X.; Li, P.; Miao, Y.-X.; Du, W.; Wang, C.-B. Involvement of ROS/ASMase/JNK Signalling Pathway in Inhibiting UVA-Induced Apoptosis of HaCaT Cells by Polypeptide from Chlamys farreri. Free. Radic. Res. 2008, 42, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Mantena, S.K.; Katiyar, S.K. Grape Seed Proanthocyanidins Inhibit UV-Radiation-Induced Oxidative Stress and Activation of MAPK and NF-ΚB Signaling in Human Epidermal Keratinocytes. Free. Radic. Biol. Med. 2006, 40, 1603–1614. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Wan, S.; Jiang, Q.; Amaral, A.; Lu, S.; Hu, G.; Bi, Z.; Kouttab, N.; Chu, W.; Wan, Y. All-Trans Retinoic Acid Attenuates Ultraviolet Radiation-Induced down-Regulation of Aquaporin-3 and Water Permeability in Human Keratinocytes. J. Cell. Physiol. 2008, 215, 506–516. [Google Scholar] [CrossRef] [PubMed]
- Sandgruber, F.; Gielsdorf, A.; Baur, A.C.; Schenz, B.; Müller, S.M.; Schwerdtle, T.; Stangl, G.I.; Griehl, C.; Lorkowski, S.; Dawczynski, C. Variability in Macro- and Micronutrients of 15 Commercially Available Microalgae Powders. Mar. Drugs 2021, 19, 310. [Google Scholar] [CrossRef]
- Susanto, E.; Fahmi, A.S.; Hosokawa, M.; Miyashita, K. Variation in Lipid Components from 15 Species of Tropical and Temperate Seaweeds. Mar. Drugs 2019, 17, 630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muthusamy, V.; Hodges, L.D.; Macrides, T.A.; Boyle, G.M.; Piva, T.J. Effect of Novel Marine Nutraceuticals on IL-1α-Mediated TNF-α Release from UVB-Irradiated Human Melanocyte-Derived Cells. Oxid. Med. Cell. Longev. 2011, 2011, 728645. [Google Scholar] [CrossRef]
- Gray-Schopfer, V.C.; Karasarides, M.; Hayward, R.; Marais, R. Tumor Necrosis Factor-α Blocks Apoptosis in Melanoma Cells When BRAF Signaling Is Inhibited. Cancer Res. 2007, 67, 122–129. [Google Scholar] [CrossRef]
- Ivanov, V.N.; Ronai, Z. Down-Regulation of Tumor Necrosis Factor α Expression by Activating Transcription Factor 2 Increases UVC-Induced Apoptosis of Late-Stage Melanoma Cells. J. Biol. Chem. 1999, 274, 14079–14089. [Google Scholar] [CrossRef]
- Cadet, J.; Douki, T.; Ravanat, J.-L. Oxidatively Generated Damage to Cellular DNA by UVB and UVA Radiation. Photochem. Photobiol. 2015, 91, 140–155. [Google Scholar] [CrossRef]
- Brenner, M.; Hearing, V.J. The Protective Role of Melanin against UV Damage in Human Skin. Photochem. Photobiol. 2008, 84, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Huynh, T.T.; Chan, K.S.; Piva, T.J. Effect of Ultraviolet Radiation on the Expression of Pp38MAPK and Furin in Human Keratinocyte-Derived Cell Lines. Photodermatol. Photoimmunol. Photomed. 2009, 25, 20–29. [Google Scholar] [CrossRef]
- Leng, H.; Luo, X.; Ma, L.; Kang, K.; Zheng, Z. Reversal of Ultraviolet B-Induced Immunosuppression by Inhibition of the Extracellular Signal-Regulated Mitogen-Activated Protein Kinase. Photodermatol. Photoimmunol. Photomed. 2009, 25, 264–269. [Google Scholar] [CrossRef]
- Bivik, C.; Öllinger, K. JNK Mediates UVB-Induced Apoptosis Upstream Lysosomal Membrane Permeabilization and Bcl-2 Family Proteins. Apoptosis 2008, 13, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Sample, A.; He, Y.-Y. Mechanisms and Prevention of UV-Induced Melanoma. Photodermatol. Photoimmunol. Photomed. 2018, 34, 13–24. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. PubChem Compound Summary for CID 11347535, Marizomib. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/11347535 (accessed on 13 July 2022).
- Waters, A.L.; Hill, R.T.; Place, A.R.; Hamann, M.T. The Expanding Role of Marine Microbes in Pharmaceutical Development. Curr. Opin. Biotechnol. 2010, 21, 780–786. [Google Scholar] [CrossRef] [Green Version]
- Testa, U.; Castelli, G.; Pelosi, E. Melanoma: Genetic Abnormalities, Tumor Progression, Clonal Evolution and Tumor Initiating Cells. Med. Sci. 2017, 5, 28. [Google Scholar] [CrossRef] [PubMed]
- Liu-Smith, F.; Jia, J.; Zheng, Y. UV-Induced Molecular Signaling Differences in Melanoma and Non-Melanoma Skin Cancer. In Ultraviolet Light in Human Health, Diseases and Environment; Ahmad, S.I., Ed.; Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2017; Volume 996, pp. 27–40. ISBN 978-3-319-56016-8. [Google Scholar]
- Bobos, M. Histopathologic Classification and Prognostic Factors of Melanoma: A 2021 Update. Ital. J. Dermatol. Venereol. 2021, 156, 300–321. [Google Scholar] [CrossRef]
- Vidwans, S.J.; Flaherty, K.T.; Fisher, D.E.; Tenenbaum, J.M.; Travers, M.D.; Shrager, J. A Melanoma Molecular Disease Model. PLoS ONE 2011, 6, e18257. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Wakamatsu, K.; Sarna, T. Photodegradation of Eumelanin and Pheomelanin and Its Pathophysiological Implications. Photochem. Photobiol. 2018, 94, 409–420. [Google Scholar] [CrossRef]
- Raimondi, S.; Suppa, M.; Gandini, S. Melanoma Epidemiology and Sun Exposure. Acta Derm. Venereol. 2020, 100, adv00136. [Google Scholar] [CrossRef] [PubMed]
- Sweet, M.; Kirkham, N.; Bendall, M.; Currey, L.; Bythell, J.; Heupel, M. Evidence of Melanoma in Wild Marine Fish Populations. PLoS ONE 2012, 7, e41989. [Google Scholar] [CrossRef]
- Potts, B.C.; Albitar, M.X.; Anderson, K.C.; Baritaki, S.; Berkers, C.; Bonavida, B.; Chandra, J.; Chauhan, D.; Cusack, J.C.; Fenical, W.; et al. Marizomib, a Proteasome Inhibitor for All Seasons: Preclinical Profile and a Framework for Clinical Trials. Curr. Cancer Drug Targets 2011, 11, 254–284. [Google Scholar] [CrossRef]
- Millward, M.; Price, T.; Townsend, A.; Sweeney, C.; Spencer, A.; Sukumaran, S.; Longenecker, A.; Lee, L.; Lay, A.; Sharma, G.; et al. Phase 1 Clinical Trial of the Novel Proteasome Inhibitor Marizomib with the Histone Deacetylase Inhibitor Vorinostat in Patients with Melanoma, Pancreatic and Lung Cancer Based on in Vitro Assessments of the Combination. Investig. New Drugs 2012, 30, 2303–2317. [Google Scholar] [CrossRef] [PubMed]
- Kong, D.-X.; Jiang, Y.-Y.; Zhang, H.-Y. Marine Natural Products as Sources of Novel Scaffolds: Achievement and Concern. Drug Discov. Today 2010, 15, 884–886. [Google Scholar] [CrossRef]
- Spainhour, C.B. Natural Products. In Drug Discovery Handbook; Gad, S.C., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2005; pp. 11–72. ISBN 978-0-471-72878-8. [Google Scholar]
- Saide, A.; Lauritano, C.; Ianora, A. A Treasure of Bioactive Compounds from the Deep Sea. Biomedicines 2021, 9, 1556. [Google Scholar] [CrossRef]
- Saide, A.; Martínez, K.A.; Ianora, A.; Lauritano, C. Unlocking the Health Potential of Microalgae as Sustainable Sources of Bioactive Compounds. Int. J. Mol. Sci. 2021, 22, 4383. [Google Scholar] [CrossRef] [PubMed]
- Fenical, W. Marine Microbial Natural Products: The Evolution of a New Field of Science. J. Antibiot. 2020, 73, 481–487. [Google Scholar] [CrossRef]
- Rangel, M.; de Barcellos Falkenberg, M. An Overview of the Marine Natural Products in Clinical Trials and on the Market. J. Coast. Life Med. 2015, 3, 421–428. [Google Scholar] [CrossRef]
- Blunt, J.W.; Copp, B.R.; Munro, M.H.G.; Northcote, P.T.; Prinsep, M.R. Marine Natural Products. Nat. Prod. Rep. 2006, 23, 26. [Google Scholar] [CrossRef]
- Bode, H.B.; Bethe, B.; Höfs, R.; Zeeck, A. Big Effects from Small Changes: Possible Ways to Explore Nature’s Chemical Diversity. ChemBioChem 2002, 3, 619. [Google Scholar] [CrossRef]
- Sayed, A.M.; Hassan, M.H.A.; Alhadrami, H.A.; Hassan, H.M.; Goodfellow, M.; Rateb, M.E. Extreme Environments: Microbiology Leading to Specialized Metabolites. J. Appl. Microbiol. 2020, 128, 630–657. [Google Scholar] [CrossRef]
- Wilson, Z.E.; Brimble, M.A. Molecules Derived from the Extremes of Life. Nat. Prod. Rep. 2009, 26, 44–71. [Google Scholar] [CrossRef] [PubMed]
- Wilson, Z.E.; Brimble, M.A. Molecules Derived from the Extremes of Life: A Decade Later. Nat. Prod. Rep. 2021, 38, 24–82. [Google Scholar] [CrossRef] [PubMed]




| Compound | Marine Organism | In Vitro/In Vivo | IC50/GI50/LC50 or Tested Concentration | Administration | Mechanism of Action | Ref. |
|---|---|---|---|---|---|---|
| Bacteria | ||||||
| Aromatic polychete akazamicin Actinofuranone C N-formilantranilic acid | AKA32 strain of actinomycetes Nonomuraea sp. | In vitro on melanoma cell B16 | IC50 1.7 μM IC50 1.2 μM IC50 25 μM, | In cell-culture media | N/A | [12] |
| Lipid 430 | Genus Algibacter | In vitro on melanoma cell A2058 | IC50 175 μM | In cell-culture media | Inhibition of cell proliferation | [13] |
| Enigromic acid Deoxyenigrolides A Deoxyenigrolides B | Mixobacteria Enhygromyxa sp. | In vitro on melanoma cell B16 | IC50 46 μM | In cell-culture media | N/A | [18] |
| Phenazine-1-carboxylic acid (PCA) | Pseudomonas aeruginosa GS-33. | In vitro SK-MEL-2 melanoma cells | GI50 of 2.30 μg/mL since GI50 value of 10 μg/mL | In cell-culture media | Reduced cell density Induction of apoptosis | [19] |
| Lyso-ornithine lipids | Genus Lacinutrix | In vitro on melanoma cells A2058 | 50 µM, 100 µM, 150 µM | In cell-culture media | N/A | [21] |
| Fungi | ||||||
| H-10 | Genus Fusarum | In vitro in melanoma model H10 | 50 µM | In cell-culture media | Induction of the apoptosis of cells via a mitochondrial pathway. Increased activity of caspases 3. Inhibition of cell growth. | [22] |
| Penicitrinine A | Penicilium citrinum | In vitro on melanoma cells A735 | IC50 20.12 µM | In cell-culture media | Induction of apoptosis by decreasing of the expression of Bcl-2 and increasing of the expression of Bax. Anti-metastatic effects. Inhibition of proliferation | [23] |
| Chlovalicin B | Digiratispora marina | In vitro on melanoma cells A2058 | IC50 37 µM | In cell-culture media | N/A | [24] |
| Lulworthinone | Lulworthiaceae family | In vitro on melanoma cells A2058 | From 6.25 µg/mL to 100 µg/mL | In cell-culture media | Inhibition of cell proliferation. | [25] |
| Pyrenosetin A Pyrenosetin B Pyrenosetin C Phomasetin | crude extract of Pyrenochaetopsis sp. FVE-001 | In vitro on melanoma cells A375 | IC50 2.8 µM IC50 6.3 µM IC50 140.3 µM IC50 37.3 µM. | In cell-culture media | N/A | [27] |
| Microalgae | ||||||
| Oxylipin 13-HOTE | Chlamydomonas debaryana | In vitro on melanoma cancer cell line UACC-62 | IC50 71.9 ± 3.6 μM | In cell-culture media | Decreased the level of ATP in UACC-62 in dose-dependent manner | [52] |
| Oxylipin 15-HEPE | Nannochloropsis gaditana | In vitro on mela-noma cancer cell line UACC-62 | IC50 53.9 ± 6.4 μM | In cell-culture media | Decreased the level of ATP in UACC-62 in dose-dependent manner | [52] |
| Raw extracts | Skeletonema marinoi (clone FE60) | In vitro on melanoma A2058 cells | 25-100 μg/mL | In cell-culture media | N/A | [30] |
| Raw extracts and fractions | Isochrysis galbana | In vitro on melanoma A2058 cells | 100 μg/mL | In cell-culture media | N/A | [31] |
| Amphidinol 22 | Amphidinium carterae | In vitro on melanoma cells A2058 | IC50 16.4 μM | In cell-culture media | N/A | [36] |
| Linear polyketide stragulin A | genus Amphidinium/Stragulum bicolor | In vitro on melanoma cell A2058 derived from metastatic site. | IC50 0.18 µM | In cell-culture media | N/A | [37] |
| Euplotin C | Euplotes crassus | In vitro on melanoma cells A2058 | N/A | In the cell-culture media | Down-regulation of the levels of B-Raf, ERK1/2 and p-Akt, promotion of the apoptosis by activation of the RyR | [44] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montuori, E.; Capalbo, A.; Lauritano, C. Marine Compounds for Melanoma Treatment and Prevention. Int. J. Mol. Sci. 2022, 23, 10284. https://doi.org/10.3390/ijms231810284
Montuori E, Capalbo A, Lauritano C. Marine Compounds for Melanoma Treatment and Prevention. International Journal of Molecular Sciences. 2022; 23(18):10284. https://doi.org/10.3390/ijms231810284
Chicago/Turabian StyleMontuori, Eleonora, Anita Capalbo, and Chiara Lauritano. 2022. "Marine Compounds for Melanoma Treatment and Prevention" International Journal of Molecular Sciences 23, no. 18: 10284. https://doi.org/10.3390/ijms231810284
APA StyleMontuori, E., Capalbo, A., & Lauritano, C. (2022). Marine Compounds for Melanoma Treatment and Prevention. International Journal of Molecular Sciences, 23(18), 10284. https://doi.org/10.3390/ijms231810284