
Early Life Stress Affects Bdnf Regulation: A Role for Exercise Interventions

 ,
,
Abstract
:1. Introduction
2. Modeling Early Life Stress and Exercise in Rodents
3. Behavioral Outcomes in Stress and Exercise Models
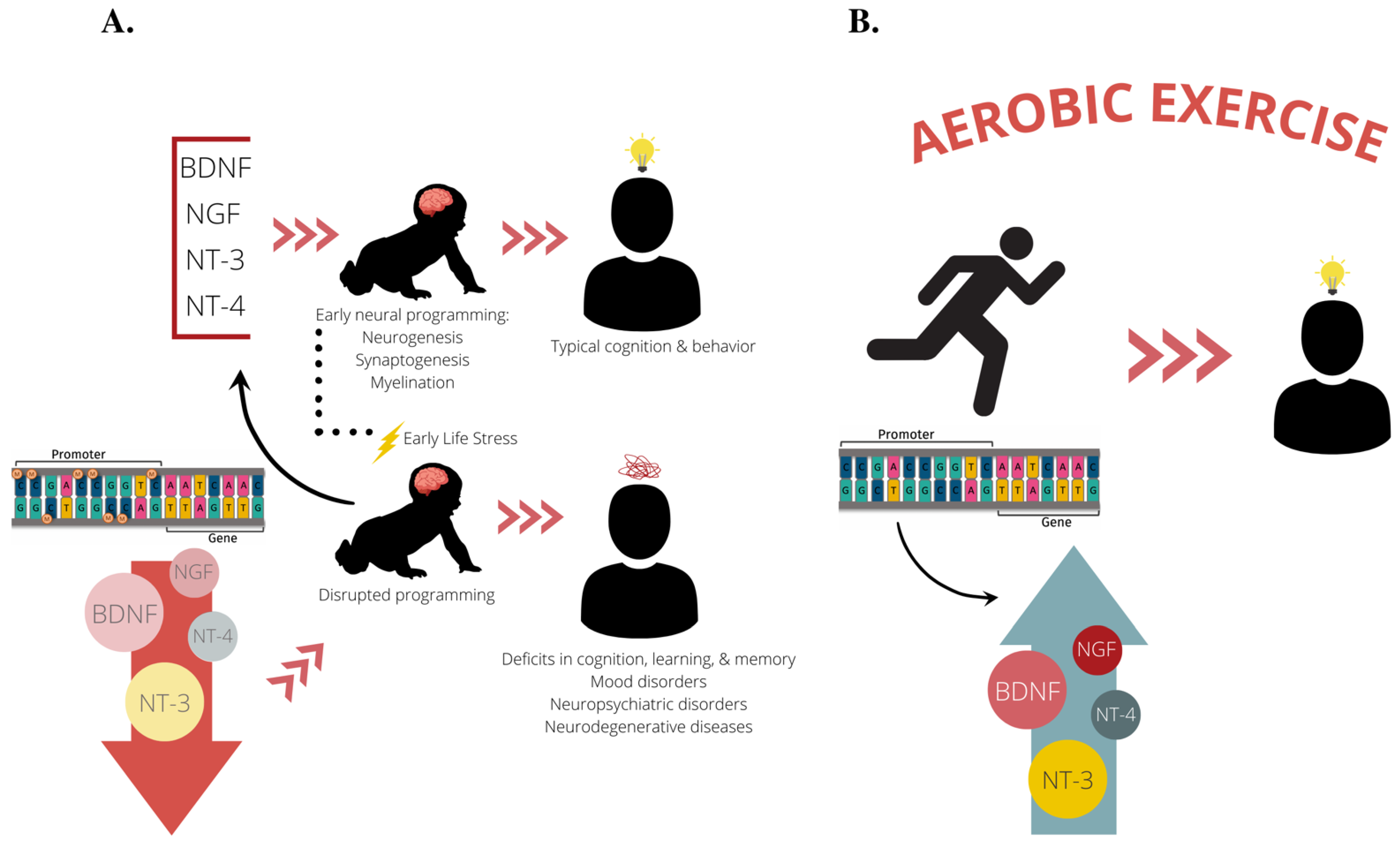
4. Epigenetics and Neurotrophins
5. Stress, Exercise and Neurotrophins
6. Call to Action: Further Exploring ELS & Exercise
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Hoeijmakers, L.; Lucassen, P.J.; Korosi, A. The Interplay of Early-Life Stress, Nutrition, and Immune Activation Programs Adult Hippocampal Structure and Function. Front. Mol. Neurosci. 2015, 7, 103. [Google Scholar] [CrossRef] [PubMed]
- Andersen, S.L. Trajectories of Brain Development: Point of Vulnerability or Window of Opportunity? Neurosci. Biobehav. Rev. 2003, 27, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Burns, S.B.; Szyszkowicz, J.K.; Luheshi, G.N.; Lutz, P.E.; Turecki, G. Plasticity of the Epigenome during Early-Life Stress. Semin. Cell Dev. Biol. 2018, 77, 115–132. [Google Scholar] [CrossRef] [PubMed]
- Milbocker, K.A.; Campbell, T.S.; Collins, N.; Kim, S.H.; Smith, I.F.; Roth, T.L.; Klintsova, A.Y. Glia-Driven Brain Circuit Refinement is Altered by Early-Life Adversity: Behavioral Outcomes. Front. Behav. Neurosci. 2021, 15. [Google Scholar] [CrossRef] [PubMed]
- Roth, T.L.; Lubin, F.D.; Funk, A.J.; Sweatt, J.D. Lasting Epigenetic Influence of Early-Life Adversity on the BDNF Gene. Biol. Psychiatry 2009, 65, 760–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, E.J.; Reichardt, L.F. Neurotrophins: Roles in Neuronal Development and Function. Annu. Rev. Neurosci. 2001, 24, 677. [Google Scholar] [CrossRef] [Green Version]
- Duman, R.S. Role of Neurotrophic Factors in the Etiology and Treatment of Mood Disorders. NeuroMolecular Med. 2004, 5, 11–25. [Google Scholar] [CrossRef]
- Mitchelmore, C.; Gede, L. Brain Derived Neurotrophic Factor: Epigenetic Regulation in Psychiatric Disorders. Brain Res. 2014, 1586, 162–172. [Google Scholar] [CrossRef] [Green Version]
- Woo, N.H.; Lu, B. Regulation of Cortical Interneurons by Neurotrophins: From Development to Cognitive Disorders. Neuroscientist 2006, 12, 43–56. [Google Scholar] [CrossRef]
- Tessarollo, L. Pleiotropic Functions of Neurotrophins in Development. Cytokine Growth Factor Rev. 1998, 9, 125–137. [Google Scholar] [CrossRef]
- Oliveira, S.L.B.; Pillat, M.M.; Cheffer, A.; Lameu, C.; Schwindt, T.T.; Ulrich, H. Functions of Neurotrophins and Growth Factors in Neurogenesis and Brain Repair. Cytom. Part A 2013, 83, 76–89. [Google Scholar] [CrossRef] [Green Version]
- Blum, R.; Konnerth, A. Neurotrophin-Mediated Rapid Signaling in the Central Nervous System: Mechanisms and Functions. Physiology 2005, 20, 70–78. [Google Scholar] [CrossRef] [Green Version]
- Bath, K.G.; Schilit, A.; Lee, F.S. Stress Effects on BDNF Expression: Effects of Age, Sex, and Form of Stress. Neuroscience 2013, 239, 149–156. [Google Scholar] [CrossRef]
- Miao, Z.; Wang, Y.; Sun, Z. The Relationships between Stress, Mental Disorders, and Epigenetic Regulation of BDNF. Int. J. Mol. Sci. 2020, 21, 1375. [Google Scholar] [CrossRef] [Green Version]
- Alleva, E.; Santucci, D. Psychosocial vs. “Physical” Stress Situations in Rodents and Humans: Role of Neurotrophins. Physiol. Behav. 2001, 73, 313–320. [Google Scholar] [CrossRef]
- Smith, M.A.; Makino, S.; Kvetnansky, R.; Post, R.M. Stress and Glucocorticoids Affect the Expression of Brain-Derived Neurotrophic Factor and Neurotrophin-3 MRNAs in the Hippocampus. J. Neurosci. 1995, 15, 1768–1777. [Google Scholar] [CrossRef] [Green Version]
- Afzalpour, M.E.; Chadorneshin, H.T.; Foadoddini, M.; Eivari, H.A. Comparing Interval and Continuous Exercise Training Regimens on Neurotrophic Factors in Rat Brain. Physiol. Behav. 2015, 147, 78–83. [Google Scholar] [CrossRef]
- Duman, R.S. Neurotrophic Factors and Regulation of Mood: Role of Exercise, Diet and Metabolism. Neurobiol. Aging 2005, 26, 88–93. [Google Scholar] [CrossRef]
- Baek, S.-S. Role of Exercise on the Brain. J. Exerc. Rehabil. 2016, 12, 380. [Google Scholar] [CrossRef] [Green Version]
- Szuhany, K.L.; Bugatti, M.; Otto, M.W. A Meta-Analytic Review of the Effects of Exercise on Brain-Derived Neurotrophic Factor. J. Psychiatr. Res. 2015, 60, 56. [Google Scholar] [CrossRef]
- Swain, R.A.; Berggren, K.L.; Kerr, A.L.; Patel, A.; Peplinski, C.; Sikorski, A.M. On Aerobic Exercise and Behavioral and Neural Plasticity. Brain Sci. 2012, 2, 709–744. [Google Scholar] [CrossRef] [Green Version]
- Grońska-Pęski, M.; Gonçalves, J.T.; Hébert, J.M. Enriched Environment Promotes Adult Hippocampal Neurogenesis through FGFRs. J. Neurosci. 2021, 41, 2899–2910. [Google Scholar] [CrossRef]
- Eriksson, P.S.; Perfilieva, E.; Björk-Eriksson, T.; Alborn, A.M.; Nordborg, C.; Peterson, D.A.; Gage, F.H. Neurogenesis in the Adult Human Hippocampus. Nat. Med. 1998, 4, 1313–1317. [Google Scholar] [CrossRef]
- Manni, L.; Micera, A.; Pistillo, L.; Aloe, L. Neonatal Handling in Eae-Susceptible Rats Alters NGF levels and Mast Cell Distribution in the Brain. Int. J. Dev. Neurosci. 1998, 16, 1–8. [Google Scholar] [CrossRef]
- Fabricius, K.; Wörtwein, G.; Pakkenberg, B. The Impact of Maternal Separation on Adult Mouse Behaviour and on the Total Neuron Number in the Mouse Hippocampus. Brain Struct. Funct. 2008, 212, 403–416. [Google Scholar] [CrossRef] [Green Version]
- Marais, L.; van Rensburg, S.J.; van Zyl, J.M.; Stein, D.J.; Daniels, W.M.U. Maternal Separation of Rat Pups Increases the Risk of Developing Depressive-like Behavior after Subsequent Chronic Stress by Altering Corticosterone and Neurotrophin Levels in the Hippocampus. Neurosci. Res. 2008, 61, 106–112. [Google Scholar] [CrossRef]
- Daniels, W.M.U.; Marais, L.; Stein, D.J.; Russell, V.A. Exercise Normalizes Altered Expression of Proteins in the Ventral Hippocampus of Rats Subjected to Maternal Separation. Exp. Physiol. 2012, 97, 239–247. [Google Scholar] [CrossRef]
- Riem, M.M.E.; Alink, L.R.A.; Out, D.; Van Ijzendoorn, M.H.; Bakermans-Kranenburg, M.J. Beating the Brain about Abuse: Empirical and Meta-Analytic Studies of the Association between Maltreatment and Hippocampal Volume across Childhood and Adolescence. Dev. Psychopathol. 2015, 27, 507–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabrera, C.; Torres, H.; Harcourt, S. The Neurological and Neuropsychological Effects of Child Maltreatment. Aggress. Violent Behav. 2020, 54, 101408. [Google Scholar] [CrossRef]
- Hart, H.; Rubia, K.; Heinze, H.-J.; Holt, D.J. Neuroimaging of Child Abuse: A Critical Review. Front. Hum. Neurosci. 2012, 6, 52. [Google Scholar] [CrossRef]
- Lange, I.; Goossens, L.; Bakker, J.; Michielse, S.; Van Winkel, R.; Lissek, S.; Leibold, N.; Marcelis, M.; Wichers, M.; Van Os, J.; et al. Neurobehavioural Mechanisms of Threat Generalization Moderate the Link between Childhood Maltreatment and Psychopathology in Emerging Adulthood. J. Psychiatry Neurosci. 2019, 44, 185–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bremner, J.D.; Vythilingam, M.; Vermetten, E.; Southwick, S.M.; McGlashan, T.; Nazeer, A.; Khan, S.; Vaccarino, L.V.; Soufer, R.; Garg, P.K.; et al. MRI and PET Study of Deficits in Hippocampal Structure and Function in Women with Childhood Sexual Abuse and Posttraumatic Stress Disorder. Am. J. Psychiatry 2003, 160, 924–932. [Google Scholar] [CrossRef] [Green Version]
- Walker, C.-D.; Bath, K.G.; Joels, M.; Korosi, A.; Larauche, M.; Lucassen, P.J.; Morris, M.J.; Raineki, C.; Roth, T.L.; Sullivan, R.M.; et al. Chronic Early Life Stress Induced by Limited Bedding and Nesting (LBN) Material in Rodents: Critical Considerations of Methodology, Outcomes and Translational Potential. Stress 2017, 20, 421–448. [Google Scholar] [CrossRef] [PubMed]
- Ivy, A.S.; Brunson, K.L.; Sandman, C.; Baram, T.Z. Dysfunctional Nurturing Behavior in Rat Dams with Limited Access to Nesting Material: A Clinically Relevant Model for Early-Life Stress. Neuroscience 2008, 154, 1132–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Levine, J.L.S.; Avila-Quintero, V.; Bloch, M.; Kaffman, A. Systematic Review and Meta-Analysis: Effects of Maternal Separation on Anxiety-like Behavior in Rodents. Transl. Psychiatry 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, M.; Peeri, M.; Hosseini, M.J. Adolescent Voluntary Exercise Attenuated Hippocampal Innate Immunity Responses and Depressive-like Behaviors Following Maternal Separation Stress in Male Rats. Physiol. Behav. 2016, 163, 177–183. [Google Scholar] [CrossRef]
- Ke, Z.; Yip, S.P.; Li, L.; Zheng, X.-X.; Tong, K.-Y. The Effects of Voluntary, Involuntary, and Forced Exercises on Brain-Derived Neurotrophic Factor and Motor Function Recovery: A Rat Brain Ischemia Model. PLoS ONE 2011, 6, e16643. [Google Scholar] [CrossRef] [Green Version]
- Svensson, M.; Rosvall, P.; Boza-Serrano, A.; Andersson, E.; Lexell, J.; Deierborg, T. Forced Treadmill Exercise Can Induce Stress and Increase Neuronal Damage in a Mouse Model of Global Cerebral Ischemia. Neurobiol. Stress 2016, 5, 8–18. [Google Scholar] [CrossRef] [Green Version]
- Uysal, N.; Kiray, M.; Sisman, A.; Camsari, U.; Gencoglu, C.; Baykara, B. Biotechnic & Histochemistry Effects of Voluntary and Involuntary Exercise on Cognitive Functions, and VEGF and BDNF Levels in Adolescent Rats. Biotech. Histochem. 2014, 90, 55–68. [Google Scholar] [CrossRef]
- Greenwood, B.N.; Foley, T.E.; Burhans, D.; Maier, S.F.; Fleshner, M. The Consequences of Uncontrollable Stress Are Sensitive to Duration of Prior Wheel Running. Brain Res. 2005, 1033, 164–178. [Google Scholar] [CrossRef]
- Greenwood, B.N.; Foley, T.E.; Le, T.V.; Strong, P.V.; Loughridge, A.B.; Day, H.E.W.; Fleshner, M. Long-Term Voluntary Wheel Running Is Rewarding and Produces Plasticity in the Mesolimbic Reward Pathway. Behav. Brain Res. 2011, 217, 354–362. [Google Scholar] [CrossRef] [Green Version]
- Sasse, S.K.; Greenwood, B.N.; Masini, C.V.; Nyhuis, T.J.; Fleshner, M.; Day, H.E.W.; Campeau, S.; Masini, C. Chronic Voluntary Wheel Running Facilitates Corticosterone Response Habituation to Repeated Audiogenic Stress Exposure in Male Rats: Original Research Report. Stress 2008, 11, 425–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbink, M.R.; Naninck, E.F.G.; Lucassen, P.J.; Korosi, A. Early-life Stress Diminishes the Increase in Neurogenesis after Exercise in Adult Female Mice. Hippocampus 2017, 27, 839–844. [Google Scholar] [CrossRef]
- Botha, M.; Grace, L.; Bugarith, K.; Russell, V.A.; Kidd, M.; Seedat, S.; Hemmings, S.M. The Impact of Voluntary Exercise on Relative Telomere Length in a Rat Model of Developmental Stress. BMC Res. Notes 2012, 5, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Dimatelis, J.J.; Hendricks, S.; Hsieh, J.; Vlok, N.M.; Bugarith, K.; Daniels, W.M.U.; Russell, V.A. Exercise Partly Reverses the Effect of Maternal Separation on Hippocampal Proteins in 6-hydroxydopamine-lesioned Rat Brain. Wiley Online Libr. 2013, 98, 233–244. [Google Scholar] [CrossRef] [Green Version]
- Eller, O.; Morris, E.; Thyfault, J.; Christianson, J.A. Early Life Stress Reduces Voluntary Exercise and Its Prevention of Diet-Induced Obesity and Metabolic Dysfunction in Mice. Physiol. Behav. 2020, 223, 113000. [Google Scholar]
- Eller, O.C.; Yang, X.; Fuentes, I.M.; Pierce, A.N.; Jones, B.M.; Brake, A.D.; Wang, R.; Dussor, G.; Christianson, J.A. Voluntary Wheel Running Partially Attenuates Early Life Stress-Induced Neuroimmune Measures in the Dura and Evoked Migraine-Like Behaviors in Female Mice. Front. Physiol. 2021, 12, 665732. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, I.M.; Jones, B.M.; Brake, A.D.; Pierce, A.N.; Eller, O.C.; Supple, R.M.; Wright, D.E.; Christianson, J.A. Voluntary Wheel Running Improves Outcomes in an Early Life Stress-Induced Model of Urologic Chronic Pelvic Pain Syndrome in Male Mice. Pain 2021, 162, 1681–1691. [Google Scholar] [CrossRef]
- Grace, L.; Hescham, S.; Kellaway, L.A.; Bugarith, K.; Russell, V.A. Effect of Exercise on Learning and Memory in a Rat Model of Developmental Stress. Metab. Brain Dis. 2009, 24, 643–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hendricks, S.; Ojuka, E.; Kellaway, L.A.; Mabandla, M.V.; Russell, V.A. Effect of Maternal Separation on Mitochondrial Function and Role of Exercise in a Rat Model of Parkinson’s Disease. Metab. Brain Dis. 2012, 27, 387–392. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.Y.; Lin, C.H. A Comparison between Chronic Exercise Training and Desipramine as Treatments for the Depression-like Behavior of Early-Life Maternal Deprivation Rats. Neurosci. Lett. 2010, 480, 201–205. [Google Scholar] [CrossRef] [PubMed]
- James, M.H.; Campbell, E.J.; Walker, F.R.; Smith, D.W.; Richardson, H.N.; Hodgson, D.M.; Dayas, C.V. Exercise Reverses the Effects of Early Life Stress on Orexin Cell Reactivity in Male but Not Female Rats. Front. Behav. Neurosci. 2014, 8, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khorjahani, A.; Peeri, M.; Azarbayjani, M.A. The Therapeutic Effect of Exercise on Anxiety and Bowel Oxidative Stress in the Maternal Separation Animal Model. Basic Clin. Neurosci. 2020, 11, 69–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makena, N.; Bugarith, K.; Russell, V.A. Maternal Separation Enhances Object Location Memory and Prevents Exercise-Induced MAPK/ERK Signalling in Adult Sprague-Dawley Rats. Metab. Brain Dis. 2012, 27, 377–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maniam, J.; Morris, M.J. Voluntary Exercise and Palatable High-Fat Diet Both Improve Behavioural Profile and Stress Responses in Male Rats Exposed to Early Life Stress: Role of Hippocampus. Psychoneuroendocrinology 2010, 35, 1553–1564. [Google Scholar] [CrossRef]
- Marais, L.; Stein, D.J.; Daniels, W.M.U. Exercise Increases BDNF Levels in the Striatum and Decreases Depressive-like Behavior in Chronically Stressed Rats. Metab. Brain Dis. 2009, 24, 587–597. [Google Scholar] [CrossRef]
- Masrour, F.F.; Peeri, M.; Azarbayjani, M.A.; Hosseini, M.J. Voluntary Exercise During Adolescence Mitigated Negative the Effects of Maternal Separation Stress on the Depressive-Like Behaviors of Adult Male Rats: Role of NMDA Receptors. Neurochem. Res. 2018, 43, 1067–1074. [Google Scholar] [CrossRef]
- Park, S.S.; Park, H.S.; Kim, C.J.; Baek, S.S.; Kim, T.W. Exercise Attenuates Maternal Separation-Induced Mood Disorder-like Behaviors by Enhancing Mitochondrial Functions and Neuroplasticity in the Dorsal Raphe. Behav. Brain Res. 2019, 372, 112049. [Google Scholar] [CrossRef]
- Pierce, A.N.; Eller-Smith, O.C.; Christianson, J.A. Voluntary Wheel Running Attenuates Urinary Bladder Hypersensitivity and Dysfunction Following Neonatal Maternal Separation in Female Mice. Neurourol. Urodyn. 2018, 37, 1623–1632. [Google Scholar] [CrossRef] [PubMed]
- Sahafi, E.; Peeri, M.; Hosseini, M.J.; Azarbyjani, M.A. Cardiac Oxidative Stress Following Maternal Separation Stress Was Mitigated Following Adolescent Voluntary Exercise in Adult Male Rat. Physiol. Behav. 2018, 183, 39–45. [Google Scholar] [CrossRef]
- Wearick-Silva, L.E.; Marshall, P.; Viola, T.W.; Centeno-Silva, A.; de Azeredo, L.A.; Orso, R.; Li, X.; Donadio, M.V.; Bredy, T.W.; Grassi-Oliveira, R. Running during Adolescence Rescues a Maternal Separation-Induced Memory Impairment in Female Mice: Potential Role of Differential Exon-Specific BDNF Expression. Dev. Psychobiol. 2017, 59, 268–274. [Google Scholar] [CrossRef]
- Zolfaghari, F.S.; Pirri, F.; Gauvin, E.; Peeri, M.; Amiri, S. Exercise and Fluoxetine Treatment during Adolescence Protect against Early Life Stress-Induced Behavioral Abnormalities in Adult Rats. Pharmacol. Biochem. Behav. 2021, 205, 173190. [Google Scholar] [CrossRef]
- Garza, K.; Jovanovic, T. Impact of Gender on Child and Adolescent PTSD. Curr. Psychiatry Rep. 2017, 19, 1–6. [Google Scholar] [CrossRef]
- Glover, E.M.; Jovanovic, T.; Norrholm, S.D. Estrogen and Extinction of Fear Memories: Implications for Posttraumatic Stress Disorder Treatment. Biol. Psychiatry 2015, 78, 178–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludyga, S.; Gerber, M.; Pühse, U.; Looser, V.N.; Kamijo, K. Systematic Review and Meta-Analysis Investigating Moderators of Long-Term Effects of Exercise on Cognition in Healthy Individuals. Nat. Hum. Behav. 2020, 4, 603–612. [Google Scholar] [CrossRef]
- Roth, T.L. Epigenetic Mechanisms in the Development of Behavior: Advances, Challenges, and Future Promises of a New Field. Dev. Psychopathol. 2013, 25, 1279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dor, Y.; Cedar, H. Principles of DNA Methylation and Their Implications for Biology and Medicine. Lancet 2018, 392, 777–786. [Google Scholar] [CrossRef]
- Martinowich, K.; Hattori, D.; Wu, H.; Fouse, S.; He, F.; Hu, Y.; Fan, G.; Sun, Y.E. DNA Methylation-Related Chromatin Remodeling in Activity-Dependent Bdnf Gene Regulation. Science 2003, 302, 890–893. [Google Scholar] [CrossRef] [Green Version]
- Casey, B.J.; Glatt, C.E.; Tottenham, N.; Soliman, F.; Bath, K.; Amso, D.; Altemus, M.; Pattwell, S.; Jones, R.; Levita, L.; et al. Brain-Derived Neurotrophic Factor as a Model System for Examining Gene by Environment Interactions across Development. Neuroscience 2009, 164, 108–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fumagalli, F.; Molteni, R.; Racagni, G.; Riva, M.A. Stress during Development: Impact on Neuroplasticity and Relevance to Psychopathology. Prog. Neurobiol. 2007, 81, 197–217. [Google Scholar] [CrossRef] [PubMed]
- Conner, J.M.; Lauterborn, J.C.; Yan, Q.; Gall, C.M.; Varon, S. Distribution of Brain-Derived Neurotrophic Factor (BDNF) Protein and MRNA in the Normal Adult Rat CNS: Evidence for Anterograde Axonal Transport. J. Neurosci. 1997, 17, 2295–2313. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, M.E.; Xu, B.; Lu, B.; Hempstead, B.L. New Insights in the Biology of BDNF Synthesis and Release: Implications in CNS Function. J. Neurosci. 2009, 29, 12764–12767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennigan, A.; O’Callaghan, R.M.; Kelly, Á.M. Neurotrophins and Their Receptors: Roles in Plasticity, Neurodegeneration and Neuroprotection. Biochem. Soc. Trans. 2007, 35, 424–427. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Nie, Z.; Shu, H.; Kuang, Y.; Chen, X.; Cheng, J.; Yu, S.; Liu, H. The Role of BDNF on Neural Plasticity in Depression. Front. Cell. Neurosci. 2020, 14, 82. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Nagappan, G.; Lu, Y. BDNF and Synaptic Plasticity, Cognitive Function, and Dysfunction. Handb. Exp. Pharmacol. 2015, 220, 223–250. [Google Scholar] [CrossRef]
- Aid, T.; Kazantseva, A.; Piirsoo, M.; Palm, K.; Timmusk, T. Mouse and Rat BDNF Gene Structure and Expression Revisited. J. Neurosci. Res. 2007, 85, 525–535. [Google Scholar] [CrossRef]
- Blaze, J.; Roth, T.L. Caregiver Maltreatment Causes Altered Neuronal DNA Methylation in Female Rodents. Dev. Psychopathol. 2017, 29, 477–489. [Google Scholar] [CrossRef] [Green Version]
- Doherty, T.S.; Forster, A.; Roth, T.L. Global and Gene-Specific DNA Methylation Alterations in the Adolescent Amygdala and Hippocampus in an Animal Model of Caregiver Maltreatment. Behav. Brain Res. 2016, 298, 55–61. [Google Scholar] [CrossRef] [Green Version]
- Dwivedi, Y.; Rizavi, H.S.; Conley, R.R.; Roberts, R.C.; Tamminga, C.A.; Pandey, G.N. Altered Gene Expression of Brain-Derived Neurotrophic Factor and Receptor Tyrosine Kinase B in Postmortem Brain of Suicide Subjects. Arch. Gen. Psychiatry 2003, 60, 804–815. [Google Scholar] [CrossRef] [Green Version]
- Ginsberg, S.D.; Malek-Ahmadi, M.H.; Alldred, M.J.; Chen, Y.; Chen, K.; Chao, M.V.; Counts, S.E.; Mufson, E.J. Brain-Derived Neurotrophic Factor (BDNF) and TrkB Hippocampal Gene Expression Are Putative Predictors of Neuritic Plaque and Neurofibrillary Tangle Pathology. Neurobiol. Dis. 2019, 132, 104540. [Google Scholar] [CrossRef]
- Hashimoto, T.; Bergen, S.E.; Nguyen, Q.L.; Xu, B.; Monteggia, L.M.; Pierri, J.N.; Sun, Z.; Sampson, A.R.; Lewis, D.A. Relationship of Brain-Derived Neurotrophic Factor and Its Receptor TrkB to Altered Inhibitory Prefrontal Circuitry in Schizophrenia. J. Neurosci. 2005, 25, 372–383. [Google Scholar] [CrossRef] [PubMed]
- Ray, M.T.; Weickert, C.S.; Wyatt, E.; Webster, M.J. Decreased BDNF, TrkB-TK+ and GAD67 MRNA Expression in the Hippocampus of Individuals with Schizophrenia and Mood Disorders. J. Psychiatry Neurosci. 2011, 36, 195–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, D.; Chatterjee-Chakraborty, M.; Rees, S.; Cauchi, J.; de Medeiros, C.B.; Fleming, A.S. Maternal Isolation Alters the Expression of Neural Proteins during Development: ‘Stroking’ Stimulation Reverses These Effects. Brain Res. 2007, 1158, 11–27. [Google Scholar] [CrossRef] [PubMed]
- Choy, K.H.C.; De Visser, Y.; Nichols, N.R.; Van Den Buuse, M. Combined Neonatal Stress and Young-Adult Glucocorticoid Stimulation in Rats Reduce BDNF Expression in Hippocampus: Effects on Learning and Memory. Hippocampus 2008, 18, 655–667. [Google Scholar] [CrossRef]
- Lippmann, M.; Bress, A.; Nemeroff, C.B.; Plotsky, P.M.; Monteggia, L.M. Long-Term Behavioural and Molecular Alterations Associated with Maternal Separation in Rats. Eur. J. Neurosci. 2007, 25, 3091–3098. [Google Scholar] [CrossRef]
- Boersma, G.J.; Lee, R.S.; Cordner, Z.A.; Ewald, E.R.; Purcell, R.H.; Moghadam, A.A.; Tamashiro, K.L. Prenatal Stress Decreases Bdnf Expression and Increases Methylation of Bdnf Exon IV in Rats. Epigenetics 2014, 9, 437–447. [Google Scholar] [CrossRef] [Green Version]
- Fachim, H.A.; Corsi-Zuelli, F.; Loureiro, C.M.; Iamjan, S.A.; Shuhama, R.; Joca, S.; Menezes, P.R.; Heald, A.; Louzada-Junior, P.; Dalton, C.F.; et al. Early-Life Stress Effects on BDNF DNA Methylation in First-Episode Psychosis and in Rats Reared in Isolation. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2021, 108, 110188. [Google Scholar] [CrossRef]
- Seo, M.K.; Ly, N.N.; Lee, C.H.; Cho, H.Y.; Choi, C.M.; Nhu, L.H.; Lee, J.G.; Lee, B.J.; Kim, G.M.; Yoon, B.J.; et al. Early Life Stress Increases Stress Vulnerability through BDNF Gene Epigenetic Changes in the Rat Hippocampus. Neuropharmacology 2016, 105, 388–397. [Google Scholar] [CrossRef]
- Perroud, N.; Salzmann, A.; Prada, P.; Nicastro, R.; Hoeppli, M.E.; Furrer, S.; Ardu, S.; Krejci, I.; Karege, F.; Malafosse, A. Response to Psychotherapy in Borderline Personality Disorder and Methylation Status of the BDNF Gene. Transl. Psychiatry 2013, 3, 207. [Google Scholar] [CrossRef] [Green Version]
- Pilkay, S.R.; Combs-Orme, T.; Tylavsky, F.; Bush, N.; Smith, A.K. Maternal Trauma and Fear History Predict BDNF Methylation and Gene Expression in Newborns. PeerJ 2020, 2020, e8858. [Google Scholar] [CrossRef]
- Cammisuli, D.M.; Bonuccelli, U.; Daniele, S.; Martini, C.; Fusi, J.; Franzoni, F. Aerobic Exercise and Healthy Nutrition as Neuroprotective Agents for Brain Health in Patients with Parkinson’s Disease: A Critical Review of the Literature. Antioxidants 2020, 9, 380. [Google Scholar] [CrossRef] [PubMed]
- Marley, C.J.; Brugniaux, J.V.; Davis, D.; Calverley, T.A.; Owens, T.S.; Stacey, B.S.; Tsukamoto, H.; Ogoh, S.; Ainslie, P.N.; Bailey, D.M. Long-Term Exercise Confers Equivalent Neuroprotection in Females Despite Lower Cardiorespiratory Fitness. Neuroscience 2020, 427, 58–63. [Google Scholar] [CrossRef]
- Håkansson, K.; Ledreux, A.; Daffner, K.; Terjestam, Y.; Bergman, P.; Carlsson, R.; Kivipelto, M.; Winblad, B.; Granholm, A.C.; Mohammed, A.K.H. BDNF Responses in Healthy Older Persons to 35 Minutes of Physical Exercise, Cognitive Training, and Mindfulness: Associations with Working Memory Function. J. Alzheimers Dis. 2017, 55, 645. [Google Scholar] [CrossRef] [PubMed]
- Boschen, K.E.; McKeown, S.E.; Roth, T.L.; Klintsova, A.Y. Impact of Exercise and a Complex Environment on Hippocampal Dendritic Morphology, Bdnf Gene Expression, and DNA Methylation in Male Rat Pups Neonatally Exposed to Alcohol. Dev. Neurobiol. 2017, 77, 708–725. [Google Scholar] [CrossRef] [PubMed]
- Voisey, J.; Lawford, B.; Bruenig, D.; Harvey, W.; Morris, C.P.; Young, R.M.D.; Mehta, D. Differential BDNF Methylation in Combat Exposed Veterans and the Association with Exercise. Gene 2019, 698, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Piepmeier, A.T.; Etnier, J.L. Brain-Derived Neurotrophic Factor (BDNF) as a Potential Mechanism of the Effects of Acute Exercise on Cognitive Performance. J. Sport Health Sci. 2015, 4, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Murawska-Ciałowicz, E.; Wiatr, M.; Ciałowicz, M.; de Assis, G.G.; Borowicz, W.; Rocha-Rodrigues, S.; Paprocka-Borowicz, M.; Marques, A. BDNF Impact on Biological Markers of Depression—Role of Physical Exercise and Training. Int. J. Environ. Res. Public Health 2021, 18, 7553. [Google Scholar] [CrossRef]
- Teicher, M.H.; Samson, J.A. Annual Research Review: Enduring Neurobiological Effects of Childhood Abuse and Neglect. J. Child Psychol. Psychiatry 2016, 57, 241–266. [Google Scholar] [CrossRef] [Green Version]
- Walsh, N.D.; Dalgleish, T.; Lombardo, M.V.; Dunn, V.J.; Van Harmelen, A.L.; Ban, M.; Goodyer, I.M. General and Specific Effects of Early-Life Psychosocial Adversities on Adolescent Grey Matter Volume. NeuroImage Clin. 2014, 4, 308–318. [Google Scholar] [CrossRef] [Green Version]
- Tan, Z.S.; Spartano, N.L.; Beiser, A.S.; DeCarli, C.; Auerbach, S.H.; Vasan, R.S.; Seshadri, S. Physical Activity, Brain Volume, and Dementia Risk: The Framingham Study. J. Gerontol. A Biol. Sci. Med. Sci. 2017, 72, 789–795. [Google Scholar] [CrossRef] [Green Version]
- Cefis, M.; Prigent-Tessier, A.; Quirié, A.; Pernet, N.; Marie, C.; Garnier, P. The Effect of Exercise on Memory and BDNF Signaling Is Dependent on Intensity. Brain Struct. Funct. 2019, 224, 1975–1985. [Google Scholar] [CrossRef] [PubMed]
- Lippi, G.; Mattiuzzi, C.; Sanchis-Gomar, F. Updated Overview on Interplay between Physical Exercise, Neurotrophins, and Cognitive Function in Humans. J. Sport Health Sci. 2020, 9, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Sanchéz, M.A.; Bustos-Cruz, R.H.; Velasco-Orjuela, G.P.; Quintero, A.P.; Tordecilla-Sanders, A.; Correa-Bautista, J.E.; Triana-Reina, H.R.; García-Hermoso, A.; González-Ruíz, K.; Peña-Guzmán, C.A.; et al. Acute Effects of High Intensity, Resistance, or Combined Protocol on the Increase of Level of Neurotrophic Factors in Physically Inactive Overweight Adults: The BrainFit Study. Front. Physiol. 2018, 9, 741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Han, T.; Zou, X.; Zhang, H.; Feng, W.; Wang, H.; Shen, Y.; Zhang, L.; Fang, G. Long-Term High-Intensity Interval Training Increases Serum Neurotrophic Factors in Elderly Overweight and Obese Chinese Adults. Eur. J. Appl. Physiol. 2021, 121, 2773–2785. [Google Scholar] [CrossRef] [PubMed]
- Milbocker, K.A.; LeBlanc, G.L.; Brengel, E.K.; Hekmatyar, K.S.; Kulkarni, P.; Ferris, C.F.; Klintsova, A.Y. Reduced and Delayed Myelination and Volume of Corpus Callosum in an Animal Model of Fetal Alcohol Spectrum Disorders Partially Benefit from Voluntary Exercise. Sci. Rep. 2022, 12, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Varigonda, A.L.; Edgcomb, J.B.; Zima, B.T. The Impact of Exercise in Improving Executive Function Impairments among Children and Adolescents with ADHD, Autism Spectrum Disorder, and Fetal Alcohol Spectrum Disorder: A Systematic Review and Meta-Analysis. Arch. Clin. Psychiatry 2021, 47, 146–156. [Google Scholar] [CrossRef]
- Motl, R.W. Exercise and Multiple Sclerosis. Adv. Exp. Med. Biol. 2020, 1228, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Bahmani, E.; Hoseini, R.; Amiri, E. The Compensatory Increased BDNF and NGF in Patients with Multiple Sclerosis Following Home-Based Aerobic Training and Vitamin D Supplementation during COVID-19 Outbreak. Res. Sq. 2021. [Google Scholar] [CrossRef]
- Kim, B.; Kang, S. Regular Leisure-Time Physical Activity Is Effective in Boosting Neurotrophic Factors and Alleviating Menopause Symptoms. Int. J. Environ. Res. Public Health 2020, 17, 8624. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
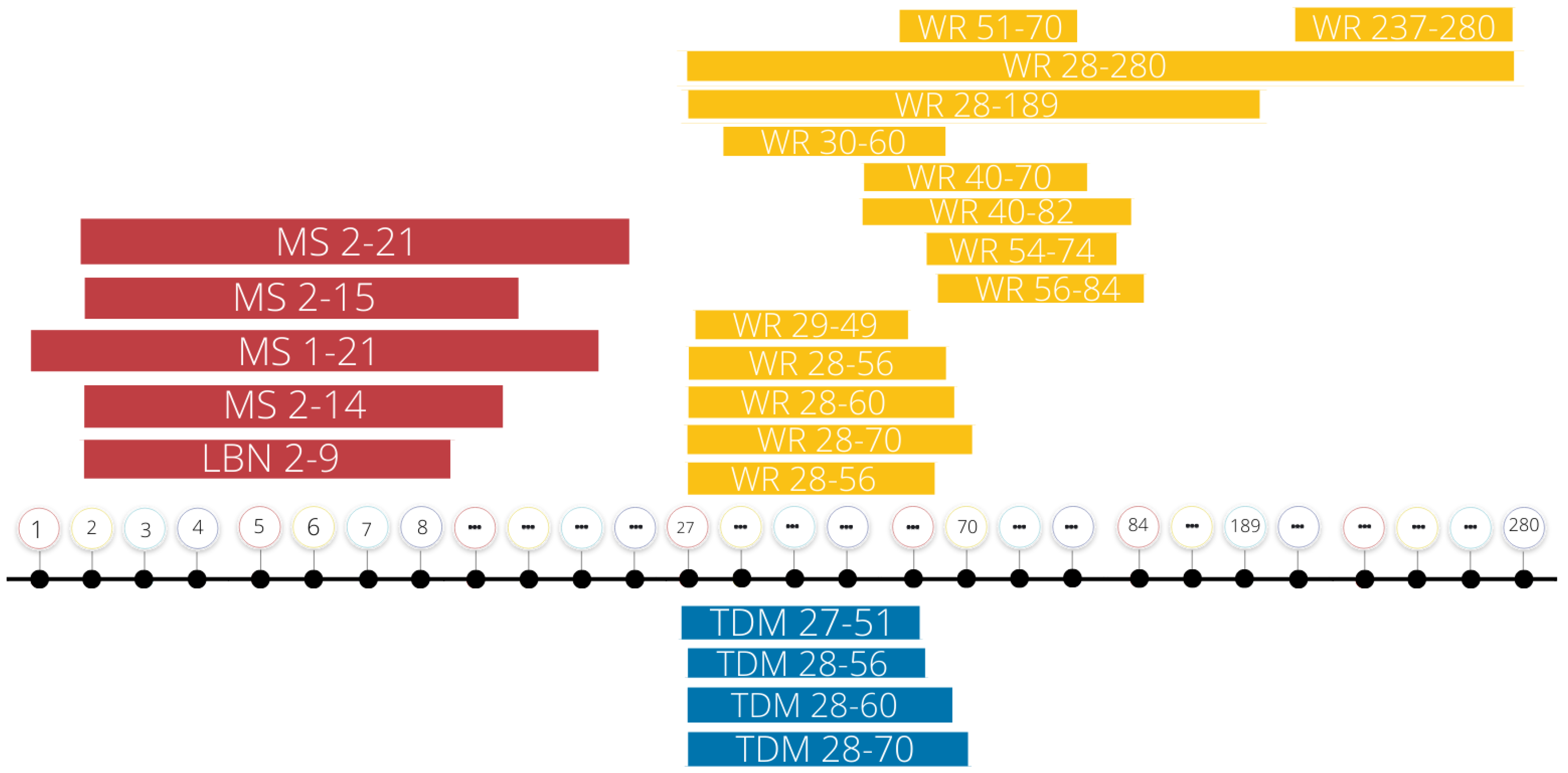
| Subjects | Sex | Form of ELS | Form of Exercise | Reference |
|---|---|---|---|---|
| C57BI/6J Mice | Both | LBN PN2-9 | Access to running wheel (voluntary exercise) PN237-280 | [43] |
| Sprague Dawley Rats | Male | MS PN2-14; 3 h daily | Access to running wheel (voluntary exercise) PN29-49 | [44] |
| Sprague Dawley Rats | Male | MS PN2-14; 3 h daily | Access to running wheel for 5 days a week PN40-82 (voluntary exercise, pair-housed) | [27] |
| Sprague Dawley Rats | Male | MS PN2-14; 3 h daily | Access to running wheel PN54-74 (voluntary exercise) | [45] |
| C57B1/6 Mice | Male | MS PN1-21; 3 h daily | Access to running wheel during 4–27 weeks of age (voluntary exercise, pair-housed) | [46] |
| C57B1/6 Mice | Female | MS PN1-21; 3 h daily | Access to running wheel (voluntary exercise) at either 4 (pair-housed) or 8 (single-housed) weeks of age | [47] |
| C57B1/6 Mice | Male | MS PN1-21; 3 h daily | Access to running wheels 4–8 weeks of age (voluntary exercise, pair housed) | [48] |
| Sprague Dawley Rats | Male | MS PN2-14; 3 h daily | Access to running wheel (voluntary exercise) PN29-49 | [49] |
| Sprague Dawley Rats | Male | MS PN2-14; 3 h daily | Access to running wheels PN54-74 (voluntary exercise) | [50] |
| Sprague Dawley Rats | Male | MD PN2-14; 1 h daily | Running on a treadmill for 6 weeks beginning at PN28 for 10 min/day for the first 5 days and incrementally increased to 60 min/day at a speed of 9 m/min (week 1), 12 m/in (week 3), and 15 m/min (week 6) (involuntary exercise) | [51] |
| Wistar Rats | Both | MS PN2-14; 3 h daily | Access to running wheels for 1 h per day, 5 days a week during PN40-70 (voluntary exercise) | [52] |
| Albino Wistar Rats | Male | MS PN2-14; 3 h daily | Access to running wheels PN28-60 (voluntary exercise, pair housed) | [53] |
| Sprague Dawley Rats | Male | MS PN2-14; 15 or 180 min/day | Access to running wheel (voluntary exercise) PN51-70 | [54] |
| Sprague Dawley Rats | Male | MS PN2-14; 3 h daily | Access to running wheel (voluntary exercise) during weeks 4–10 of age (3–4 animals per cage) | [55] |
| Sprague Dawley Rats | Male | MS PN2-14; 3 h daily | Access to individual running wheels 5 days per week PND40-82 during dark cycle only (voluntary exercise; pair housed with perforated plexiglass divider during WR access). | [56] |
| Albino Wistar Rats | Male | MS PN2-14; 3 h daily | Access to wheel running on PN21 then continuous access to running wheel PN28-60 (voluntary exercise) | [57] |
| Sprague Dawley Rats | Male | MS PN2-21; 3 h daily | Running on a treadmill for 4 weeks, 6 days/week; 3 m/min 5 min warm-up; 10 m/min 30 min exercise (first 2 weeks) or 12 m/min 40 min exercise (last 2 weeks), and 3 m/min 5 min cool-down (involuntary exercise) | [58] |
| C57B1/6 Mice | Female | MS PN1-21; 3 h daily | Access to running wheels at 4–8 weeks of age(voluntary exercise, pair-housed) | [59] |
| Albino Wistar Rats | Both | MS PN2-14; 3 h daily | 2 conditions PN28: Access to running wheel PN28-60 (voluntary exercise) OR treadmill running 5 days/week from PN28-60 for 30 min at 10 m/min (first 2 weeks), 45 min at 15 m/min (week 3), and 60 min at 15 m/min (week 4) (involuntary exercise) | [36] |
| Albino Wistar Rats | Male | MS PN2-14; 3 h daily | Access to running wheels PN28-60 (voluntary exercise) | [60] |
| Balb/c mice | Female | MS PN2-15; 180 min daily | Running on a treadmill 60 min/day at 10 m/min, 5 days per week from PN27-51 (involuntary exercise) | [61] |
| Rats (not specified) | Male | MS PN2-14; 3 h daily | 2 conditions PN28: Access to running wheel for 4 weeks (voluntary exercise) OR treadmill running 5 days/week for 4 weeks for 30 min at 10 m/min (first 2 weeks), 45 min at 15 m/min (week 3), and 60 min at 15 m/min (week 4) (involuntary exercise) | [62] |
| Subjects | Sex | Form of Early Life Stress | Age of Behavioral Test | Behavioral Outcome | Exercise Type | Reference |
|---|---|---|---|---|---|---|
| NMS Mice | Female | MS PN1-31, 3 h daily | PN56 | MS increased light sensitivity in the LDB (migraine mouse model); outcome ameliorated by WR | WR | [47] |
| Sprague-Dawley Rats | Male | MS PN2-14, 3 h daily | PN29 | WR exposure increased anxiety (EMP, OFT) WR exposure improved temporal memory (TO) and spatial learning (MWM) No effect on OIP, NOR, or OLT and no effect of MS | WR | [49] |
| Sprague-Dawley Rats | Male | MD PN12-25, 1 h daily | PN28 | MD increased immobility time on the FST and time spent in the light versus dark (LDB) WR normalized/decreased immobility time (FST) and time spent in the light box (LDB) | TM | [51] |
| Wistar Rats | Both | MS PN2-14, 3 h daily | PN75-79 | MS decreased total OFT locomotion in males and females. WR normalized locomotion in males but further decreased locomotion in females No effect on EPM open arm entries | WR | [52] |
| Albino Wistar Rats | Male | MS PN2-14, 3 h daily | PN21 | MS decreased open arm entries and time spent in open arms on the EMP and increased immobility on the FST; WR normalized behavior | WR | [53] |
| Sprague-Dawley Rats | Male | MS PN2-21, 5 h daily | PN28; 75–81 | MS decreased center duration and entries on the OFT (PN28; pre-WR) No effect of OFT locomotion or center time/entries in adulthood MS decreased open arm entries and time spent in open arms on the EMP; WR normalized behavior No effect of NOR, OIP, or TO | WR | [54] |
| Sprague-Dawley Rats | Male | MS PN2-14, 3 h daily | PN40 | MS increased FST immobility; WR normalized immobility | WR | [56] |
| Albino Wistar Rats | Male | MS PN2-14, 3 h daily | PN60 | MS increased FST immobility and decreased grooming during SPL; WR normalized behavior No effect of OFT | WR | [57] |
| Sprague-Dawley Rats | Male | MS PN2-21, 3 h daily | PN22 | MS decreased open arm entries on the EPM, increased FST immobility, and decreased OFT center locomotion WR increased open arm entries and time spent in open arms (EPM), decreased FST immobility, and increased OFT center locomotion | TM | [58] |
| Albino Wistar Rats | Both | MS PN2-14, 3 h daily | PN21 | MS increased immobility on the FST, decreased sucrose preference (SPT), and decreased grooming during splash test; WR (but not TM) normalized behavioral outcomes No effect on OFT | WR/TM | [36] |
| Albino Wistar Rats | Male | MS PN2-14, 3 h daily | PN60 | MS increased immobility on the FST, decreased sucrose preference (SPT) and decreased grooming during the SPL WR decreased immobility (FST) and normalized grooming time (SPL) No effect on OFT | WR | [60] |
| Balb/c Mice | Female | MS PN2-15, 3 h daily | PN24 | MS impaired NOR performance TM normalized NOR performance | TM | [61] |
| Rats (not specified) | Male | MS PN2-14, 3 h daily | PN61-70 | MS decreased center time on the OFT and arm time and arm entries on the EPM; WR or TM normalized these behaviors MS increased immobility on the FST and decreased sucrose consumption (SPT); WR but not TM normalized these behaviors | WR/TM | [62] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campbell, T.S.; Donoghue, K.M.; Ghosh, U.; Nelson, C.M.; Roth, T.L. Early Life Stress Affects Bdnf Regulation: A Role for Exercise Interventions. Int. J. Mol. Sci. 2022, 23, 11729. https://doi.org/10.3390/ijms231911729
Campbell TS, Donoghue KM, Ghosh U, Nelson CM, Roth TL. Early Life Stress Affects Bdnf Regulation: A Role for Exercise Interventions. International Journal of Molecular Sciences. 2022; 23(19):11729. https://doi.org/10.3390/ijms231911729
Chicago/Turabian StyleCampbell, Taylor S., Katelyn M. Donoghue, Urmi Ghosh, Christina M. Nelson, and Tania L. Roth. 2022. "Early Life Stress Affects Bdnf Regulation: A Role for Exercise Interventions" International Journal of Molecular Sciences 23, no. 19: 11729. https://doi.org/10.3390/ijms231911729
APA StyleCampbell, T. S., Donoghue, K. M., Ghosh, U., Nelson, C. M., & Roth, T. L. (2022). Early Life Stress Affects Bdnf Regulation: A Role for Exercise Interventions. International Journal of Molecular Sciences, 23(19), 11729. https://doi.org/10.3390/ijms231911729