
Characterization of a C-Type Lectin Domain-Containing Protein with Antibacterial Activity from Pacific Abalone (Haliotis discus hannai)

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
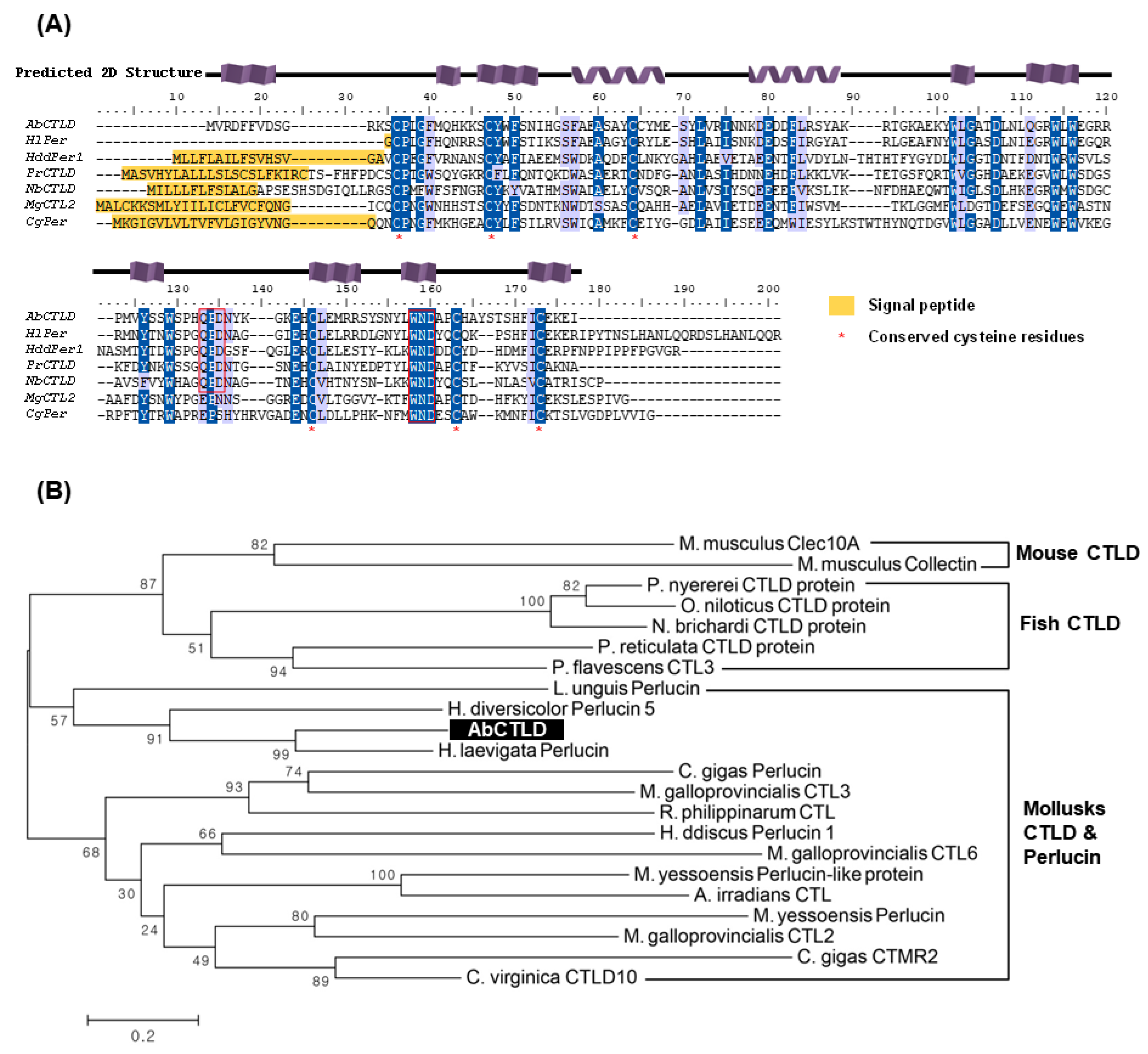
2.1. Sequence Analysis of the Gene Encoding AbCTLD
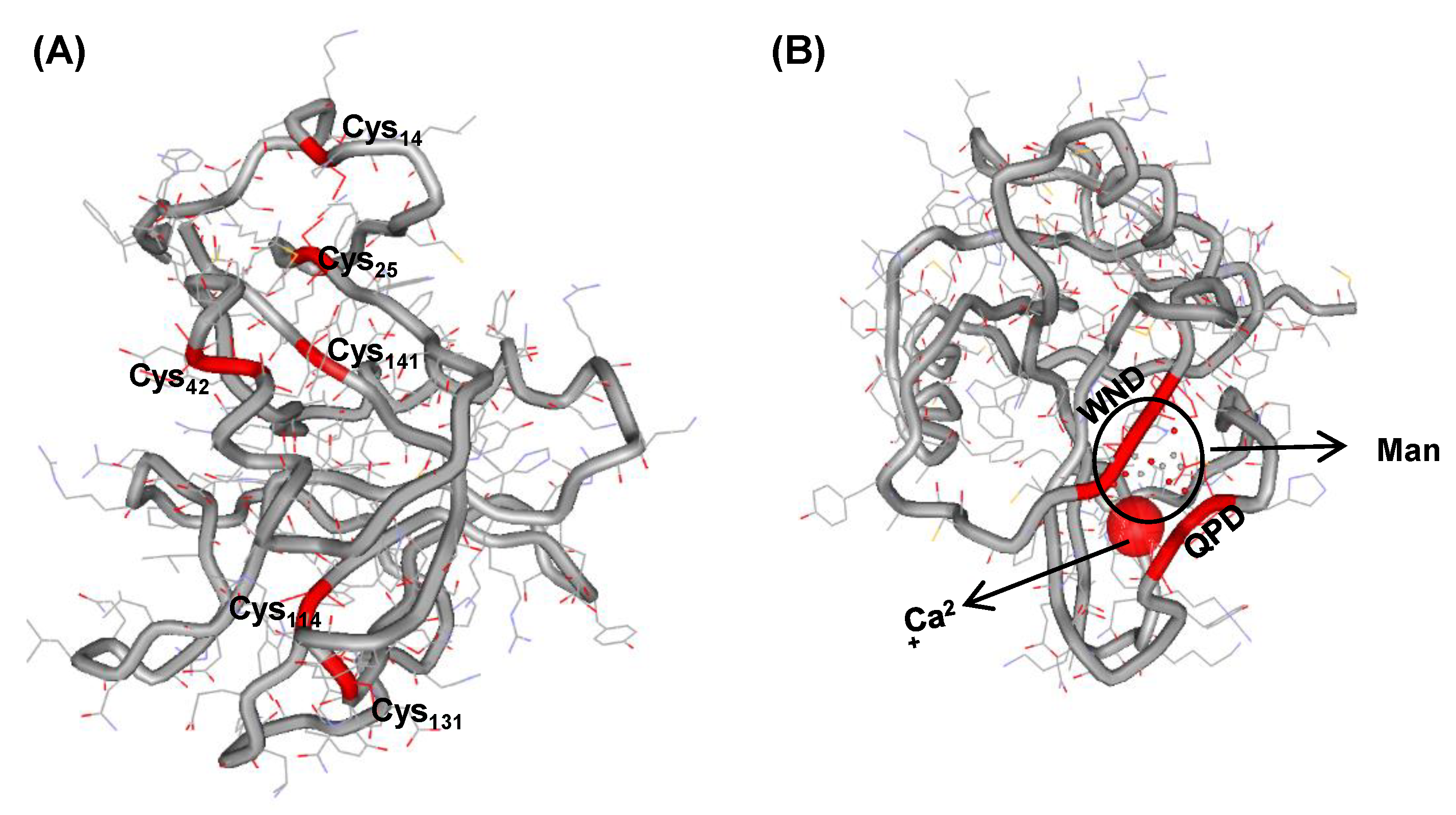
2.2. Structure Modeling and Ligand Prediction
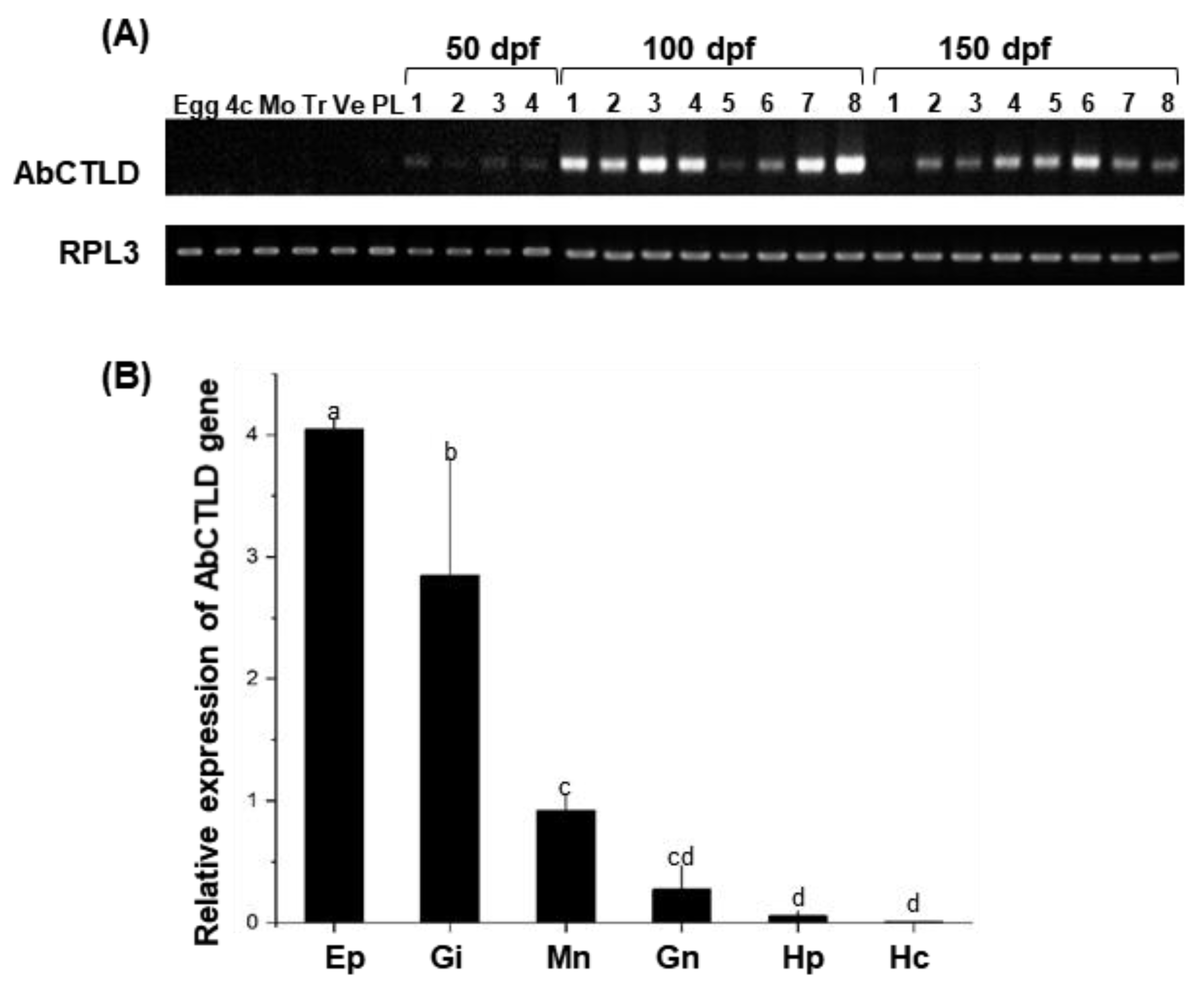
2.3. Developmental and Tissue-Specific Expression Pattern of AbCTLD
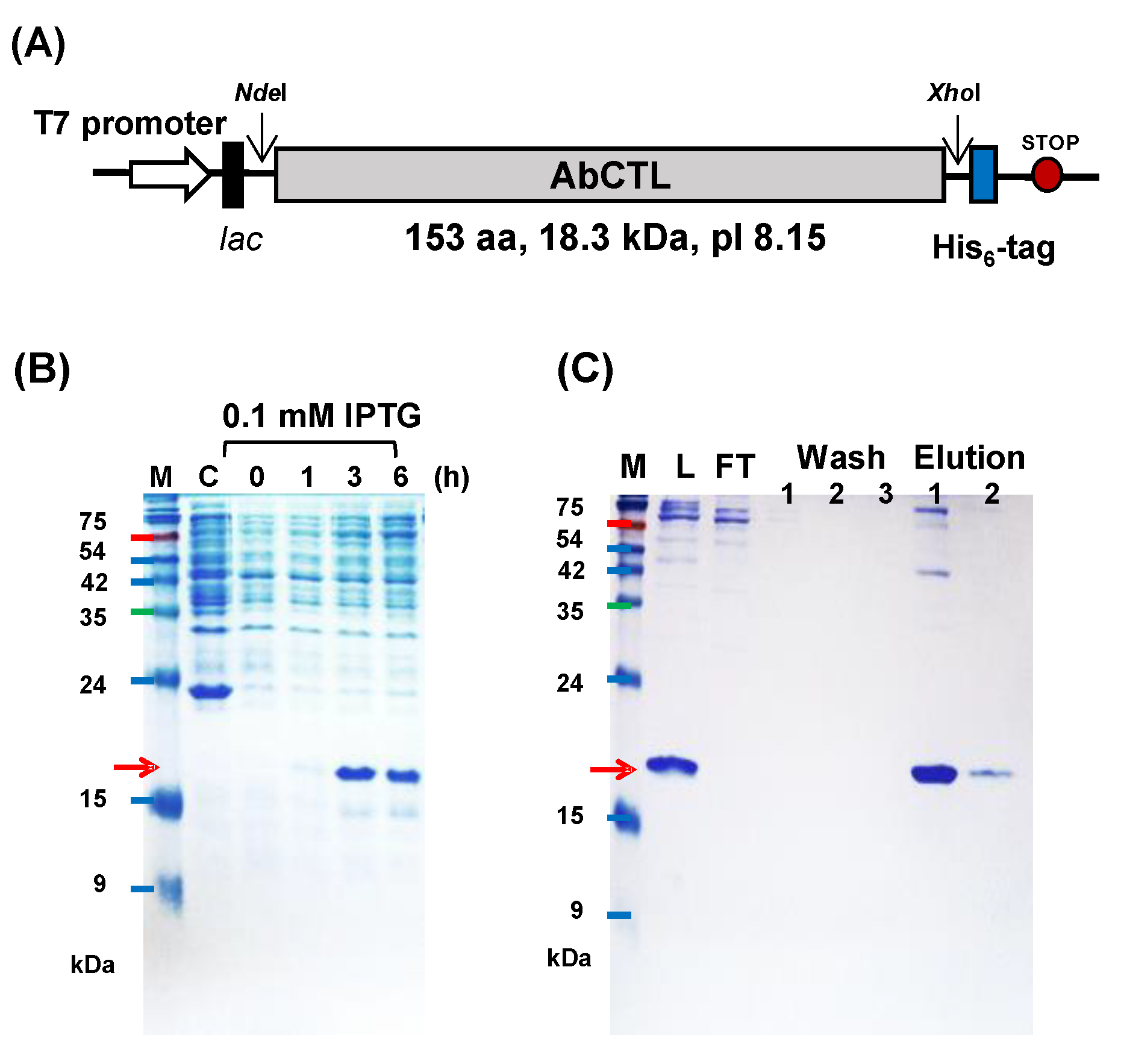
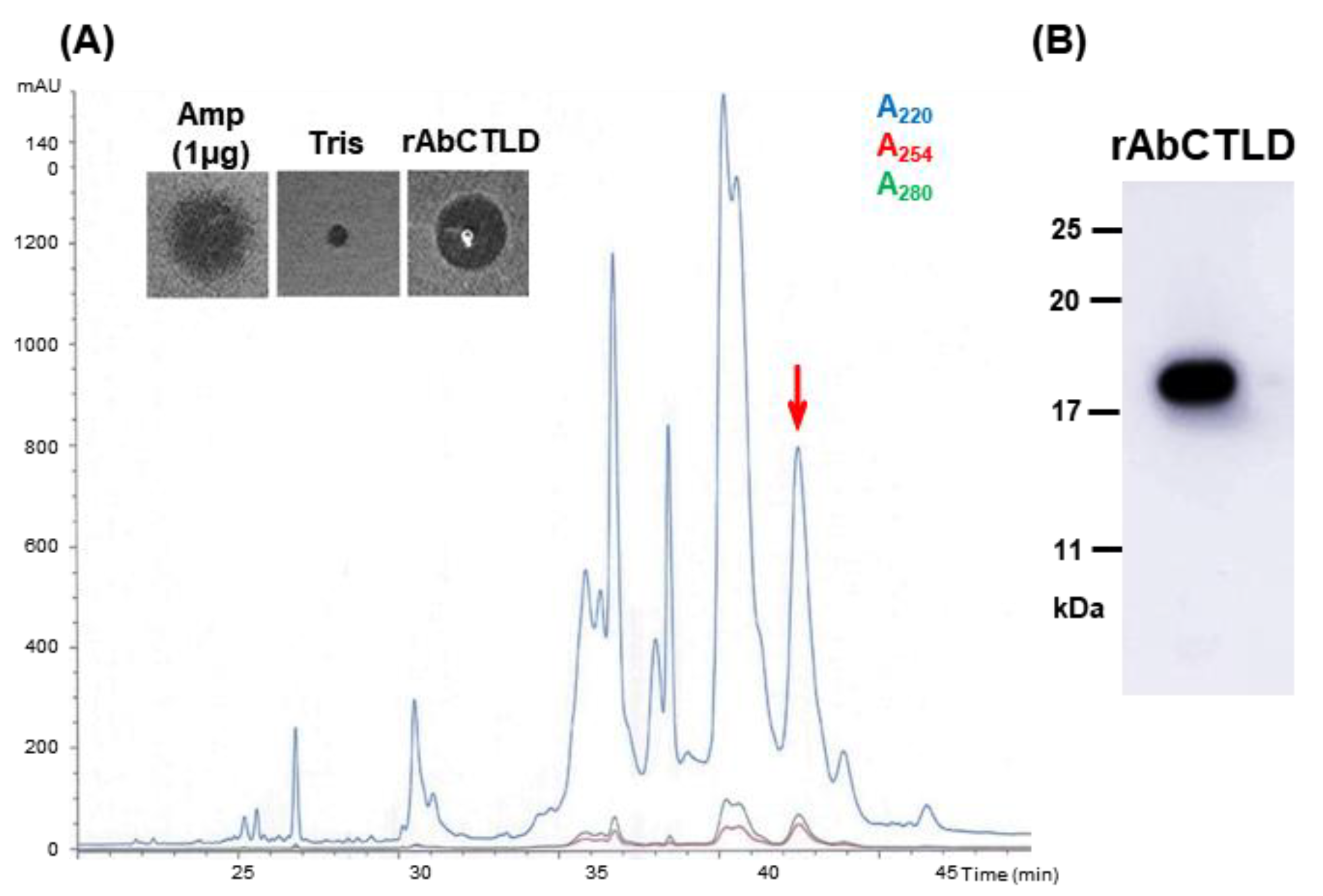
2.4. Purification of His6-Tagged rAbCTLD
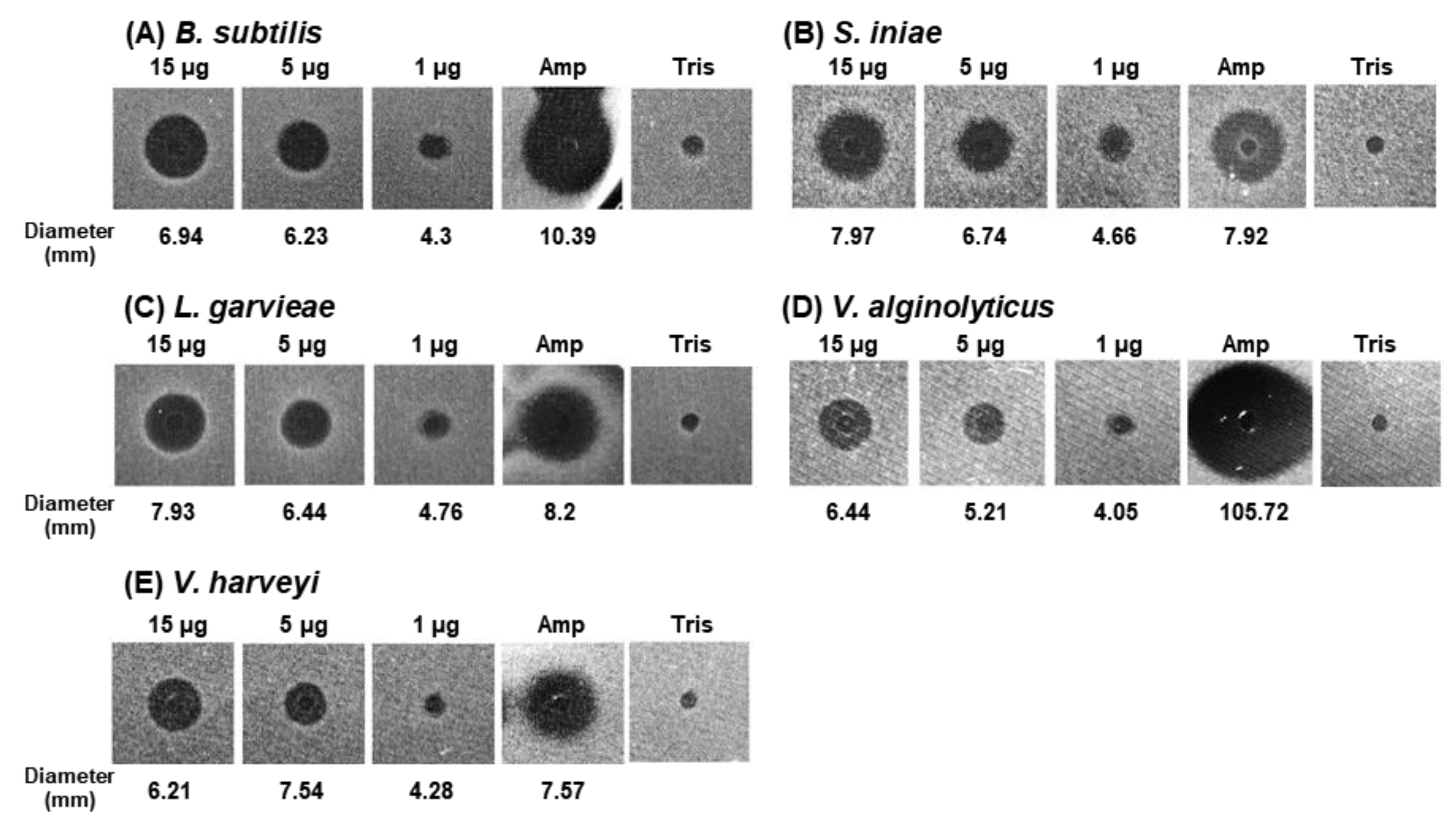
2.5. Antibacterial Activity of AbCTLD
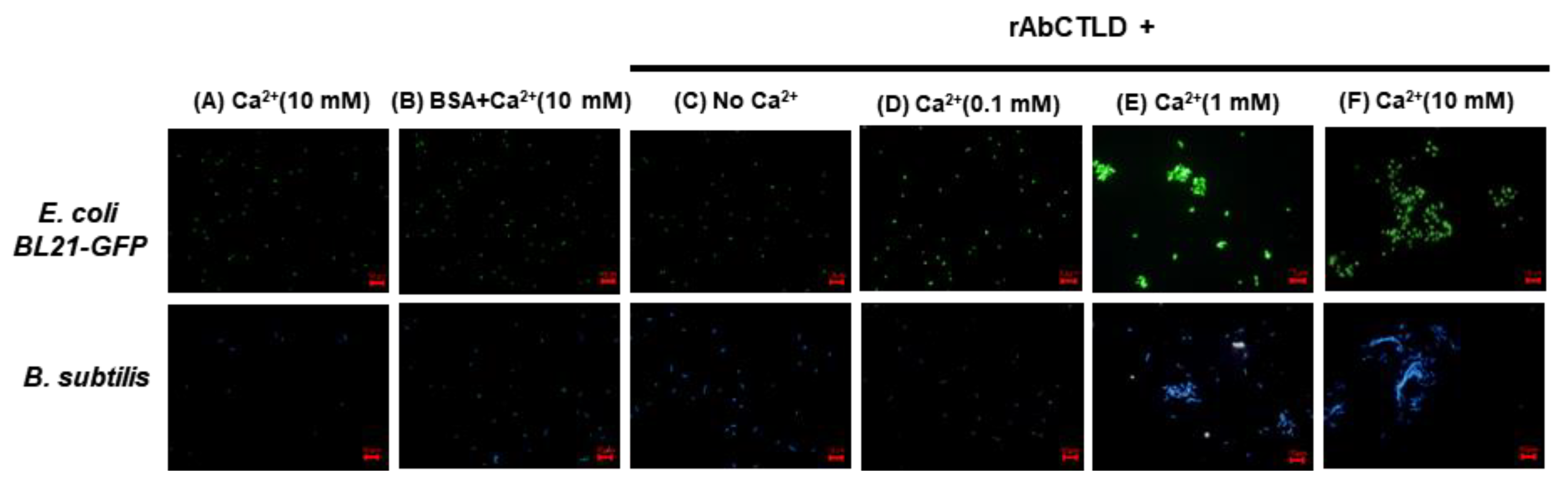
2.6. Bacterial Agglutination Stimulated by rAbCTLD
3. Discussion
4. Materials and Methods
4.1. Cloning the AbCTLD Gene
4.2. Phylogenetic Tree and Bioinformatic Analysis
4.3. Analysis of AbCTLD Gene Expression across Pacific Abalone Development
4.4. Analysis of AbCTLD Gene Expression in Adult Abalone Tissue
4.5. Purification of AbCTLD
4.6. Antibacterial Assays
4.7. Bacterial Agglutination Assays
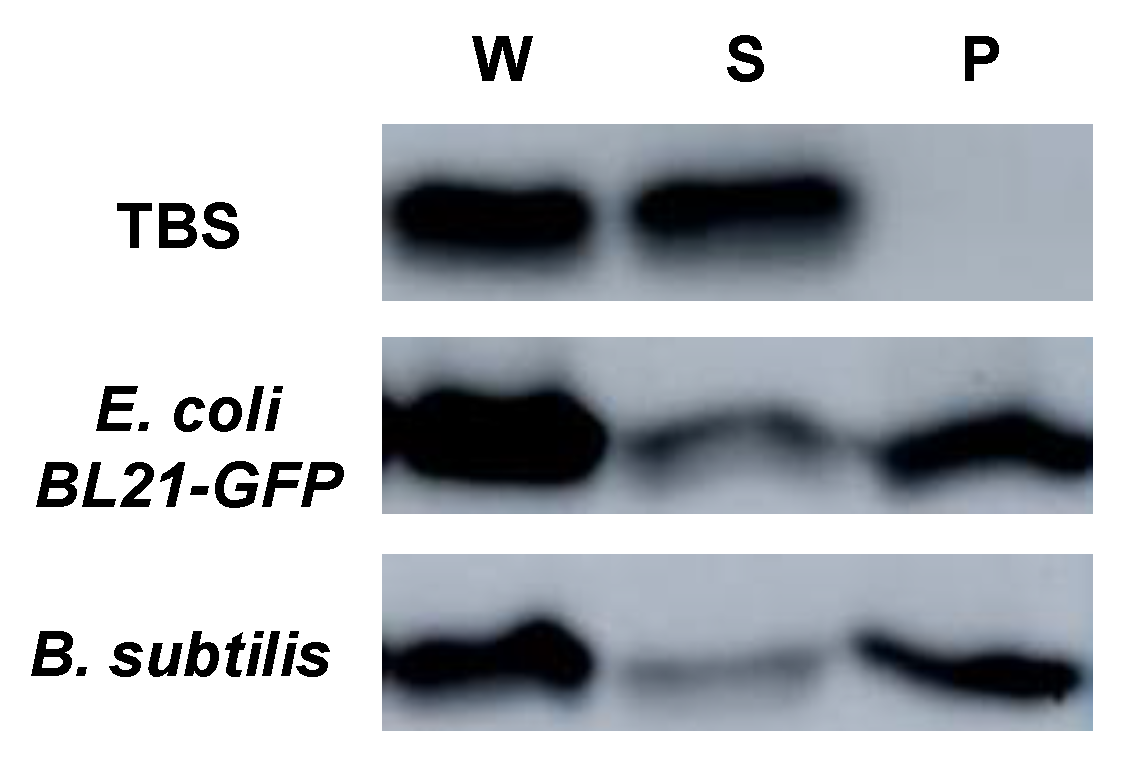
4.8. Bacterial Binding Assays
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cook, P.A. Recent trends in worldwide abalone production. J. Shellfish Res. 2016, 35, 581–583. [Google Scholar] [CrossRef]
- Park, C.J.; Lee, J.H.; Noh, J.K.; Kim, H.C.; Park, J.W.; Hwang, I.J.; Kim, S.Y. Growth of Pacific abalone, Haliotis discus hannai, using selection breeding techniques. Korean J. Malacol. 2012, 28, 343–347. [Google Scholar] [CrossRef] [Green Version]
- Oh, Y.D.; Sun, S.C.; Lee, K.S.; Lim, H.K. Growth and survival of purebred and hybrid according to intraspecific hybridization between Haliotis discus hannai and H. discus discus. Korean J. Malacol. 2018, 34, 31–41. [Google Scholar] [CrossRef]
- Tripp-Valdez, M.A.; Cicala, F.; Galindo-Sánchez, C.E.; Chacón-Ponce, K.D.; López-Landavery, E.; Díaz, D. Re-Araujo, F.; Lafarga-De la Cruz, F. Growth Performance and Transcriptomic Response of Warm-Acclimated Hybrid Abalone Haliotis rufescens (♀) × H. corrugata (♂). Mar. Biotechnol. 2021, 23, 62–76. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.J.; Kim, G.D.; Kim, J.M.; Lim, H.K. Differentially-Expressed Genes Associated with Faster Growth of the Pacific Abalone, Haliotis discus hannai. Int. J. Mol. Sci. 2015, 16, 27520–27534. [Google Scholar] [CrossRef] [Green Version]
- Oh, Y.D.; Choi, M.J.; Cho, Y.S.; Jeong, T.H.; Kim, J.M.; Lim, H.K. Discovery of Neuropeptides in the Ganglia of the Pacific Abalone Haliotis discus hannai Using Long-Read Transcriptome Analysis. J. Shellfish Res. 2020, 39, 99–111. [Google Scholar] [CrossRef]
- Loker, E.S.; Adema, C.M.; Zhang, S.M.; Kepler, T.B. Invertebrate immune systems- not homogeneous, not simple, not well understood. Immunol. Rev. 2004, 198, 10–24. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Song, X.; Wang, L.; Song, L. Pathogen-Derived Carbohydrate Recognition in Molluscs Immune Defense. Int. J. Mol. Sci. 2018, 19, 721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckeridge, M.S. The evolution of the Glycomic Codes of extracellular matrices. Biosystems 2018, 164, 112–120. [Google Scholar] [CrossRef]
- Drickamer, K.; Fadden, A.J. Genomic analysis of C-type lectins. Biochem. Soc. Symp. 2002, 69, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Lan, T.; Lia, Z.; Peng, M.; Niu, D.; Li, Y.; Li, J. A four-CRD C-type lectin from razor clam Sinonovacula constricta mediates agglutination and phagocytosis. Gene 2020, 728, 144287. [Google Scholar] [CrossRef]
- Yang, J.; Wang, L.; Zhang, H.; Qiu, L.; Wang, H.; Song, L. C-type lectin in Chlamys farreri (CfLec-1) mediating immune recognition and opsonization. PLoS ONE 2011, 6, e17089. [Google Scholar] [CrossRef] [Green Version]
- Weis, W.I.; Drickamer, K.; Hendrickson, W.A. Structure of a C-type mannose-binding protein complexed with an oligosaccharide. Nature 1992, 360, 127–134. [Google Scholar] [CrossRef]
- Zelensky, A.N.; Gready, J.E. The C-type lectin-like domain superfamily. FEBS J. 2005, 272, 6179–6217. [Google Scholar] [CrossRef]
- Jin, X.K.; Li, S.; Guo, X.N.; Cheng, L.; Wu, M.H.; Tan, S.J.; Zhu, Y.T.; Yu, A.Q.; Li, W.W.; Wang, Q. Two antibacterial C-type lectins from crustacean, Eriocheir sinensis, stimulated cellular encapsulation in vitro. Dev. Comp. Immunol. 2013, 41, 544–552. [Google Scholar] [CrossRef]
- Xu, W.T.; Wang, X.W.; Zhang, X.W.; Zhao, X.F.; Yu, X.Q.; Wang, J.X. A new C-type lectin (FcLec5) from the Chinese white shrimp Fenneropenaeus chinensis. Amino Acids 2010, 39, 1227–1239. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.W.; Xu, W.T.; Zhang, X.W.; Zhao, X.F.; Yu, X.Q.; Wang, J.X. A C-type lectin is involved in the innate immune response of Chinese white shrimp. Fish Shellfish Immunol. 2009, 27, 556–562. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Song, X.; Zhao, J.; Mu, C.; Wang, L.; Zhang, H.; Zhou, Z.; Liu, X.; Song, L. A C-type lectin (AiCTL-3) from bay scallop Argopecten irradians with mannose/galactose binding ability to bind various bacteria. Gene 2013, 531, 31–38. [Google Scholar] [CrossRef]
- Brown, G.D.; Willment, J.A.; Whitehead, L. C-type lectins in immunity and homeostasis. Nat. Rev. Immunol. 2018, 18, 374–389. [Google Scholar] [CrossRef]
- Drickamer, K.; Taylor, M.E. Recent insights into structures and functions of C-type lectins in the immune system. Curr. Opin. Struct. Biol. 2015, 34, 26–34. [Google Scholar] [CrossRef] [Green Version]
- Day, A.J. The C-type carbohydrate recognition domain (CRD) superfamily. Biochem. Soc. Trans. 1994, 22, 83–88. [Google Scholar] [CrossRef] [Green Version]
- Zelensky, A.N.; Gready, J.E. Comparative analysis of structural properties of the C-type-lectin-like domain (CTLD). Proteins 2003, 52, 466–477. [Google Scholar] [CrossRef]
- Drickamer, K. Engineering galactose-binding activity into a C-type mannose-binding protein. Nature 1992, 360, 183–186. [Google Scholar] [CrossRef]
- Li, H.; Zhang, H.; Jiang, S.; Wang, W.; Xin, L.; Wang, H.; Wang, L.; Song, L. A single-CRD C-type lectin from oyster Crassostrea gigas mediates immune recognition and pathogen elimination with a potential role in the activation of complement system. Fish Shellfish Immunol. 2015, 44, 566–575. [Google Scholar] [CrossRef]
- Kang, Y.S.; Kim, Y.M.; Park, K.I.; Kim Cho, S.; Choi, K.S.; Cho, M. Analysis of EST and lectin expressions in hemocytes of Manila clams (Ruditapes philippinarum) (Bivalvia: Mollusca) infected with Perkinsus olseni. Dev. Comp. Immunol. 2006, 30, 1119–1131. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, L.; Huang, M.; Zhang, H.; Song, L. The immune role of C-type lectins in molluscs. ISJ-Invert. Surviv. J. 2011, 8, 241–246. [Google Scholar]
- Wang, X.W.; Wang, J.X. Diversity and multiple functions of lectins in shrimp immunity. Dev. Comp. Immunol. 2013, 39, 27–38. [Google Scholar] [CrossRef]
- Iwanaga, S.; Lee, B.L. Recent advances in the innate immunity of invertebrate animals. J. Biochem. Mol. Biol. 2005, 38, 128–150. [Google Scholar] [CrossRef] [Green Version]
- Mann, K.; Weiss, I.M.; André, S.; Gabius, H.J.; Fritz, M. The amino-acid sequence of the abalone (Haliotis laevigata) nacre protein perlucin. Detection of a functional C-type lectin domain with galactose/mannose specificity. Eur. J. Biochem. 2000, 267, 5257–5264. [Google Scholar] [CrossRef]
- Kim, B.H.; Park, M.W.; Son, M.H.; Kim, T.I.; Myeong, J.I.; Cho, J.K. A Study on the Optimum Stocking Density of the Juvenile Abalone, Hailotis discus hannai Net Cage Culture or Indoor Tank Culture. Korean J. Malacol. 2013, 29, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Drickamer, K. Ca2+-dependent carbohydrate-recognition domains in animal proteins. Curr. Opin. Struct. Biol. 1993, 3, 393–400. [Google Scholar] [CrossRef]
- Weis, W.I.; Taylor, M.E.; Drickamer, K. The C-type lectin superfamily in the immune system. Immunol. Rev. 1998, 163, 19–34. [Google Scholar] [CrossRef]
- Vasta, G.R.; Quesenberry, M.; Ahmed, H.; O’Leary, N. C-type lectins and galectins mediate innate and adaptive immune functions: Their roles in the complement activation pathway. Dev. Comp. Immunol. 1999, 23, 401–420. [Google Scholar] [CrossRef]
- Li, J.; Chen, Y.; Gu, W.; Xu, F.; Li, H.; Shan, S.; Sun, X.; Yin, M.; Yang, G.; Chen, L. Characterization of a common carp intelectin gene with bacterial binding and agglutination activity. Fish Shellfish Immunol. 2021, 108, 32–41. [Google Scholar] [CrossRef]
- Li, D.; Nie, H.; Dong, S.; Huo, Z.; Yan, X. Molecular cloning and expression analysis of C-type lectin (RpCTL) in Manila clam Ruditapes philippinarum after lipopolysaccharide challenge. Fish Shellfish Immunol. 2019, 86, 981–993. [Google Scholar] [CrossRef]
- Adhya, M.; Choi, K.S.; Yu, Y.; Cho, M. Expression and localization of MCsialec, a sialic acid-specific lectin in the marine bivalve Manila clam, Ruditapes philppinarum. J. Fish Dis. 2010, 33, 889–899. [Google Scholar] [CrossRef] [PubMed]
- Low, D.E.; Liu, E.; Fuller, J.; McGeer, A. Streptococcus iniae: An emerging pathogen in the aquaculture industry. In Emerging Infections Vol. 3; Scheld, W.M., Craig, W.A., Armstrong, D., Hughes, J.M., Eds.; ASM Press: Washington, DC, USA, 1999; p. 53. [Google Scholar]
- Meyburgh, C.M.; Bragg, R.R.; Boucher, C.E. Lactococcus garvieae: An emerging bacterial pathogen of fish. Dis. Aquat. Organ. 2017, 123, 67–79. [Google Scholar] [CrossRef]
- George, M.R.; John, K.R.; Iyappan, T.; Jeyaseelan, M.J. Genetic heterogeneity among Vibrio alginolyticus isolated from shrimp farms by PCR fingerprinting. Lett. Appl. Microbiol. 2005, 40, 369–372. [Google Scholar] [CrossRef] [PubMed]
- Lavilla-Pitogo, C.R.; Baticados, M.C.L.; Cruz-Lacierda, E.R.; De La Pena, L.D. Occurrence of luminous bacterial disease of Penaeus monodon larvae in the Philippines. Aquaculture. 1990, 91, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Nam, B.H.; Jung, M.; Subramaniyam, S.; Yoo, S.I.; Markkandan, K.; Moon, J.Y.; Kim, Y.O.; Kim, D.G.; An, C.M.; Shin, Y.; et al. Transcriptome analysis revealed changes of multiple genes involved in Haliotis discus hannai innate immunity during Vibrio parahemolyticus infection. PLoS ONE 2016, 11, e0153474. [Google Scholar] [CrossRef]
- Zhang, J.; Qiu, R.; Hu, Y.H. HdhCTL1 is a novel C-type lectin of abalone Haliotis discus hannai that agglutinates Gram-negative bacterial pathogens. Fish Shellfish Immunol. 2014, 41, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Whang, I.; Lee, J. A novel C-type lectin from abalone, Haliotis discus discus, agglutinates Vibrio alginolyticus. Dev. Comp. Immunol. 2008, 32, 1034–1040. [Google Scholar] [CrossRef]
- Dambuza, I.M.; Brown, G.D. C-type lectins in immunity: Recent developments. Curr. Opin. Immunol. 2015, 32, 21–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lepenies, B.; Lee, J.; Sonkaria, S. Targeting C-type lectin receptors with multivalent carbohydrate ligands. Adv. Drug Deliv. Rev. 2013, 65, 1271–1281. [Google Scholar] [CrossRef] [PubMed]
- Weis, W.I.; Kahn, R.; Fourme, R.; Drickamer, K.; Hendrickson, W.A. Structure of the calcium-dependent lectin domain from a rat mannose-binding protein determined by MAD phasing. Science 1991, 254, 1608–1615. [Google Scholar] [CrossRef]
- Shi, Y.; Zhao, X.; Wang, Z.; Shao, Y.; Zhang, W.; Bao, Y.; Li, C. Novel Ca2+-independent C-type lectin involved in immune defense of the razor clam Sinonovacula constricta. Fish Shellfish Immunol. 2019, 84, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Asahida, T.; Kobayashi, T.; Saitoh, K.; Nakayama, I. Tissue preservation and total DNA extraction from fish stored at ambient temperature using buffers containing high concentration of urea. Fish. Sci. 1996, 62, 727–730. [Google Scholar] [CrossRef] [Green Version]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Roy, A.; Zhang, Y. Protein-ligand binding site recognition using complementary binding-specific substructure comparison and sequence profile alignment. Bioinformatics 2013, 29, 2588–2595. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.J.; Rho, D.Y.; Jeong, T.H.; Lim, H.K.; Kim, J.M. An Efficient Method for Purifying High-Quality RNA from the Hepatopancreas of the Pacific Abalone Haliotis discus hannai. J. Shellfish Res. 2018, 37, 199–205. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Duncan, D.B. Multiple range and multiple F test. Biometrics. 1955, 11, 1–42. [Google Scholar] [CrossRef]
- Lehrer, R.I.; Rosenman, M.; Harwig, S.S.; Jackson, R.; Eisenhauer, P. Ultrasensitive assays for endogenous antimicrobial polypeptides. J. Immunol. Methods 1991, 137, 167–173. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purpose | Primer | Sequence (5′-3′) |
|---|---|---|
| Identification of AbCTLD gene structure in genomic DNA | 5UTR-F | GGC TTA CAA TGT TGT GAA CTT CTG |
| Par-R2 | GTA CGC GTG ACA CGG AGC AT | |
| Par-F2 | AGC GCT TAC TGT TGC TAC ATG G | |
| 3UTR-R | CAA CAC TAA ACC ATG TTT CAG TCG G | |
| Cloning of full-length AbCTLD into pET expression vector | FullF-NdeI | CAT ATG GTG CGT GAT TTT TTT GTG GAT TC |
| FullR-XhoI | CTC GAG GAT TTC CTT TTC GCA AAT GAA GTG | |
| RT-PCR of AbCTLD | AbCTLD-F | CCT CTT GGG TTT ATG CAG CAC |
| AbCTLD-R | CGG ACT GTC TCA TTT CCA GAC | |
| RT-PCR of RPL3 (Housekeeping gene) | RPL3-F | TGT CAC CAT CCT TGA GGC AC |
| RPL3-R | CAG GAA CAG GCT TCT CCA GG | |
| qRT-PCR of AbCTLD | qAbCTLD-F | GGT GCC ACT GAT CTG AAC CT |
| qAbCTLD-R | AGG ACC GTC TCA TTT CCA GA | |
| qRT-PCR of RPL3 (Housekeeping gene) | qRPL3-F | AGT CCT TCC CTA AGG ATG ACA AG |
| qRPL3-R | GCC TCC ACA ACT TCC TTC TTA TT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, M.-J.; Kim, Y.R.; Park, N.G.; Kim, C.-H.; Oh, Y.D.; Lim, H.K.; Kim, J.-M. Characterization of a C-Type Lectin Domain-Containing Protein with Antibacterial Activity from Pacific Abalone (Haliotis discus hannai). Int. J. Mol. Sci. 2022, 23, 698. https://doi.org/10.3390/ijms23020698
Choi M-J, Kim YR, Park NG, Kim C-H, Oh YD, Lim HK, Kim J-M. Characterization of a C-Type Lectin Domain-Containing Protein with Antibacterial Activity from Pacific Abalone (Haliotis discus hannai). International Journal of Molecular Sciences. 2022; 23(2):698. https://doi.org/10.3390/ijms23020698
Chicago/Turabian StyleChoi, Mi-Jin, Yeo Reum Kim, Nam Gyu Park, Cheorl-Ho Kim, Young Dae Oh, Han Kyu Lim, and Jong-Myoung Kim. 2022. "Characterization of a C-Type Lectin Domain-Containing Protein with Antibacterial Activity from Pacific Abalone (Haliotis discus hannai)" International Journal of Molecular Sciences 23, no. 2: 698. https://doi.org/10.3390/ijms23020698
APA StyleChoi, M. -J., Kim, Y. R., Park, N. G., Kim, C. -H., Oh, Y. D., Lim, H. K., & Kim, J. -M. (2022). Characterization of a C-Type Lectin Domain-Containing Protein with Antibacterial Activity from Pacific Abalone (Haliotis discus hannai). International Journal of Molecular Sciences, 23(2), 698. https://doi.org/10.3390/ijms23020698