
Induction of Isochromanones by Co-Cultivation of the Marine Fungus Cosmospora sp. and the Phytopathogen Magnaporthe oryzae

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
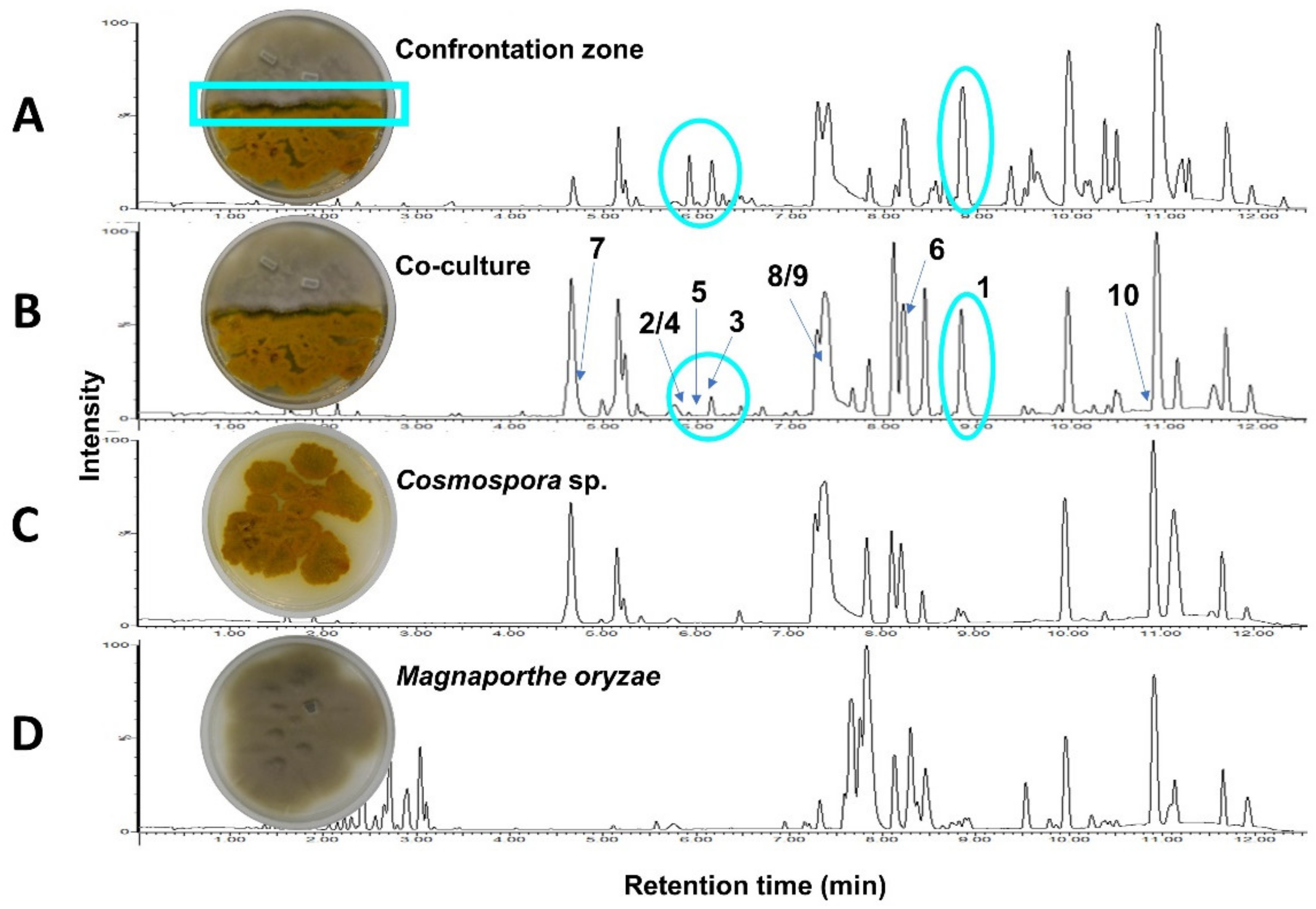
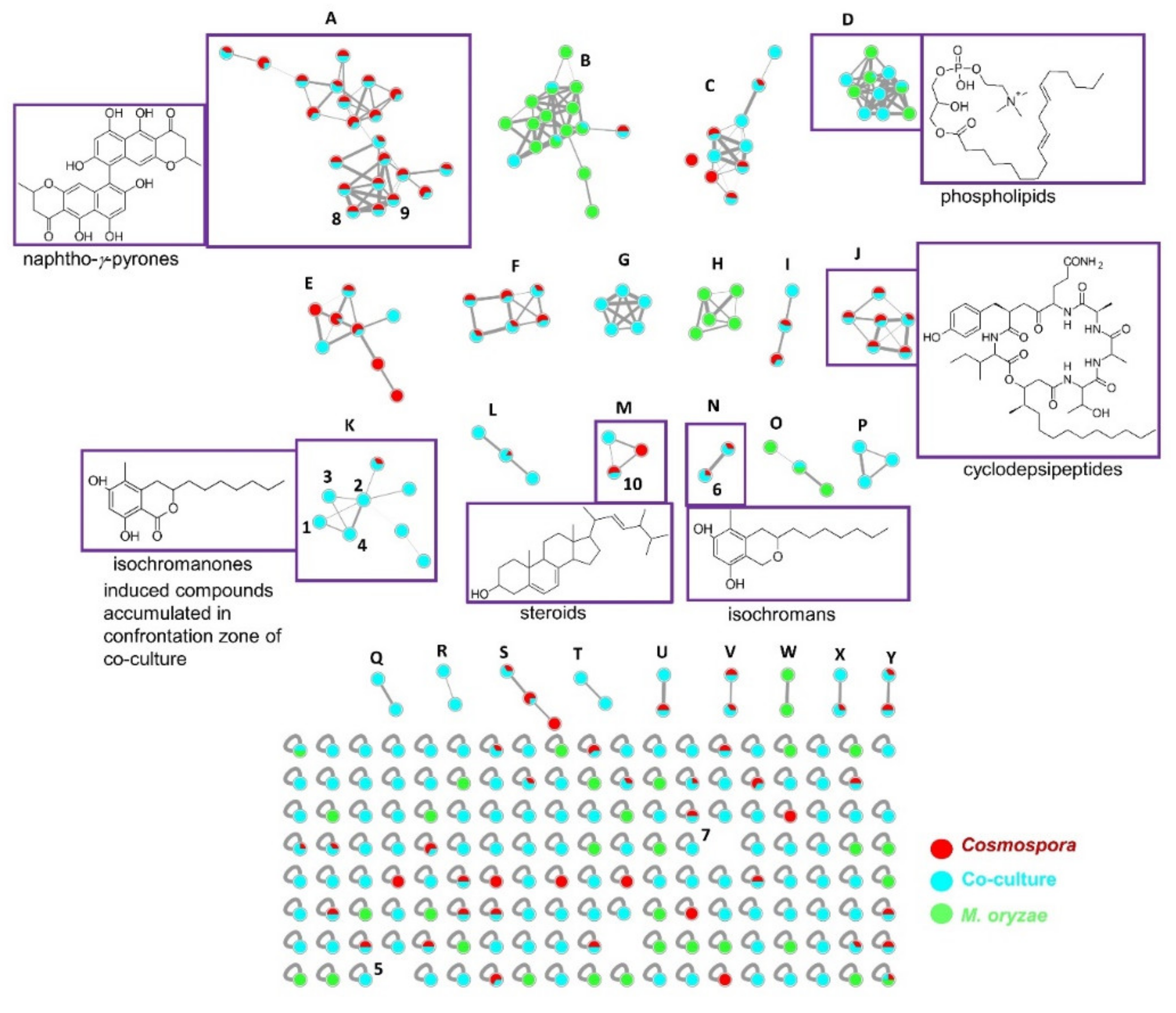
2.1. Co-Culture and Description of MN and Annotations
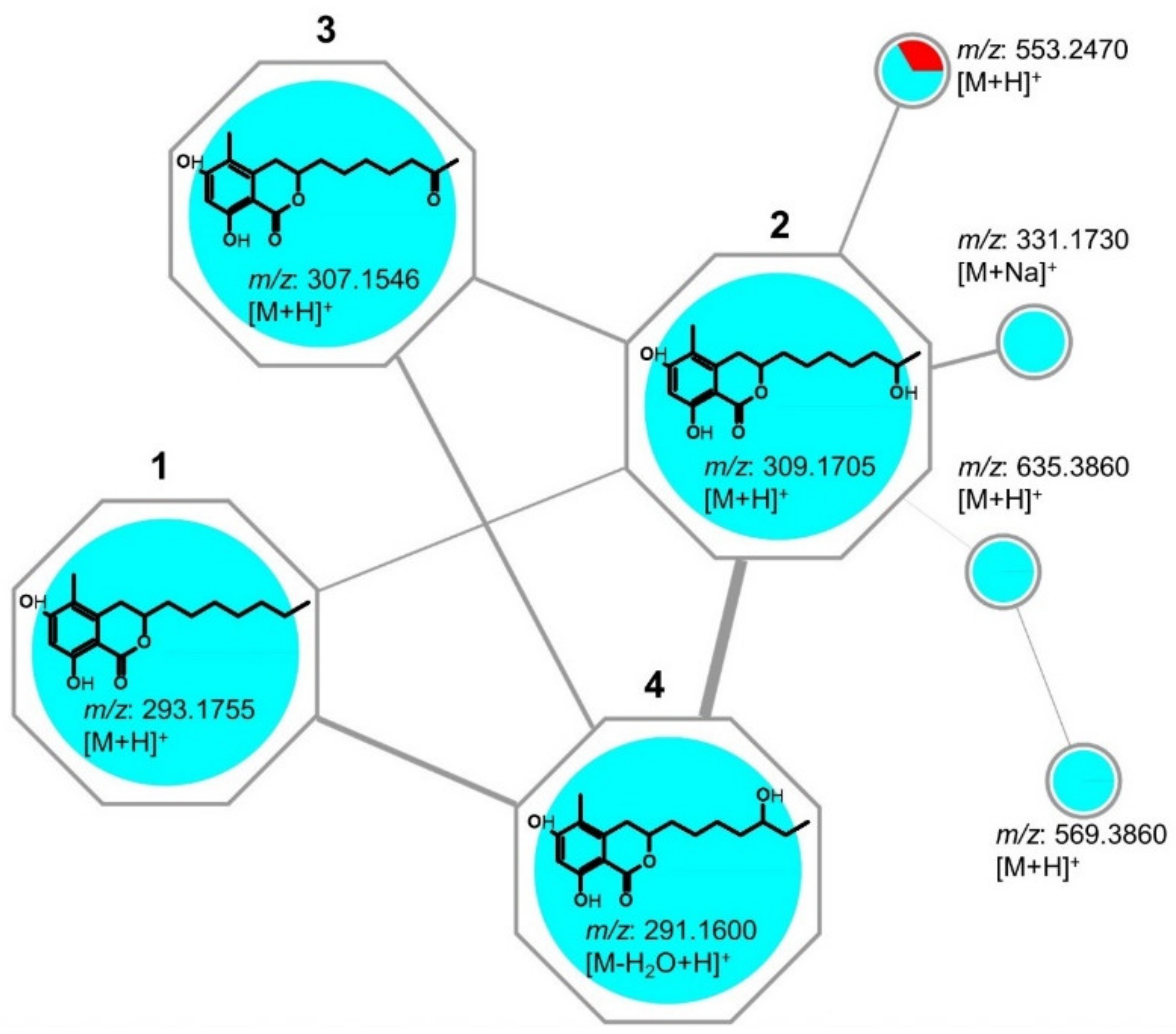
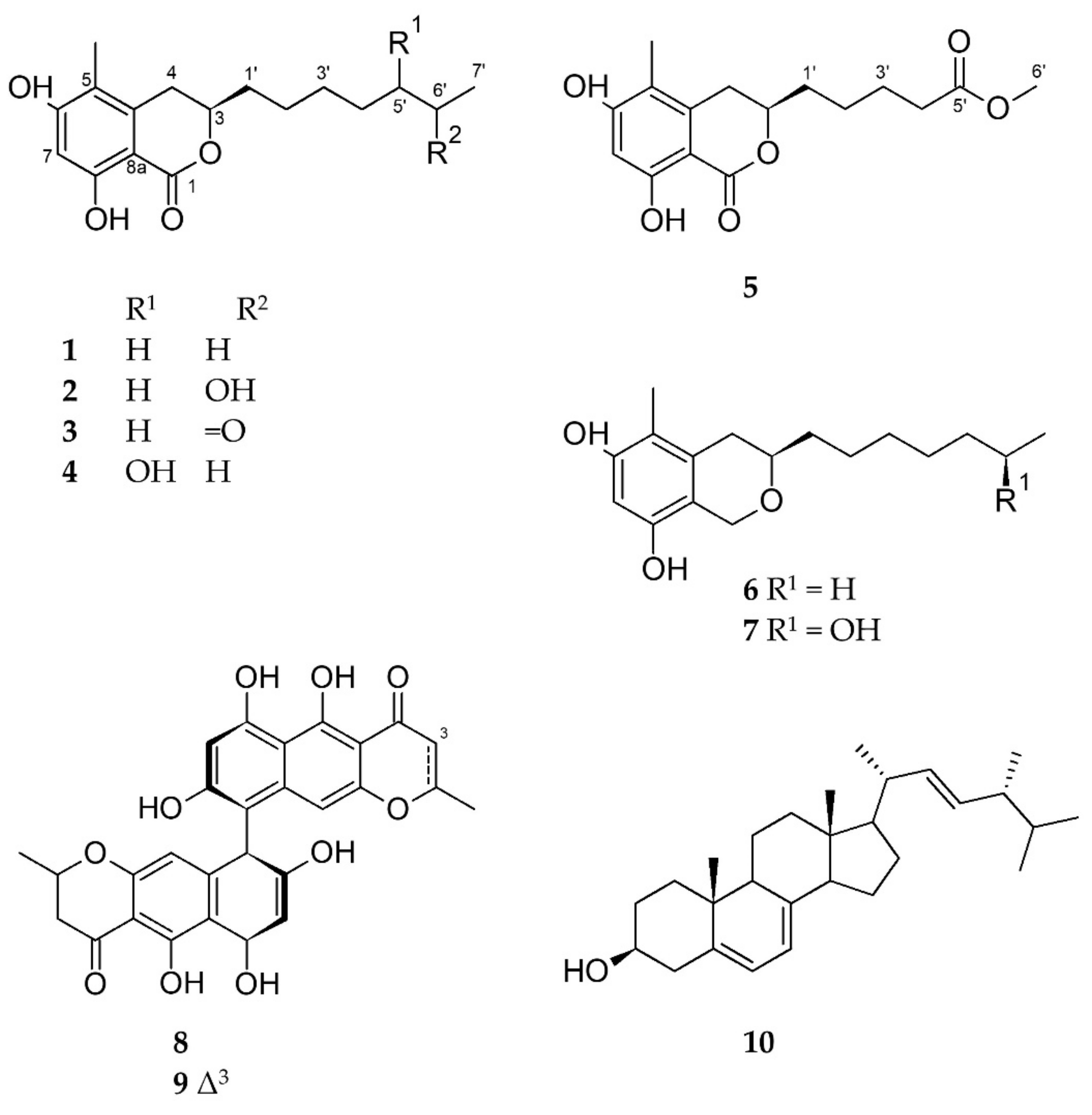
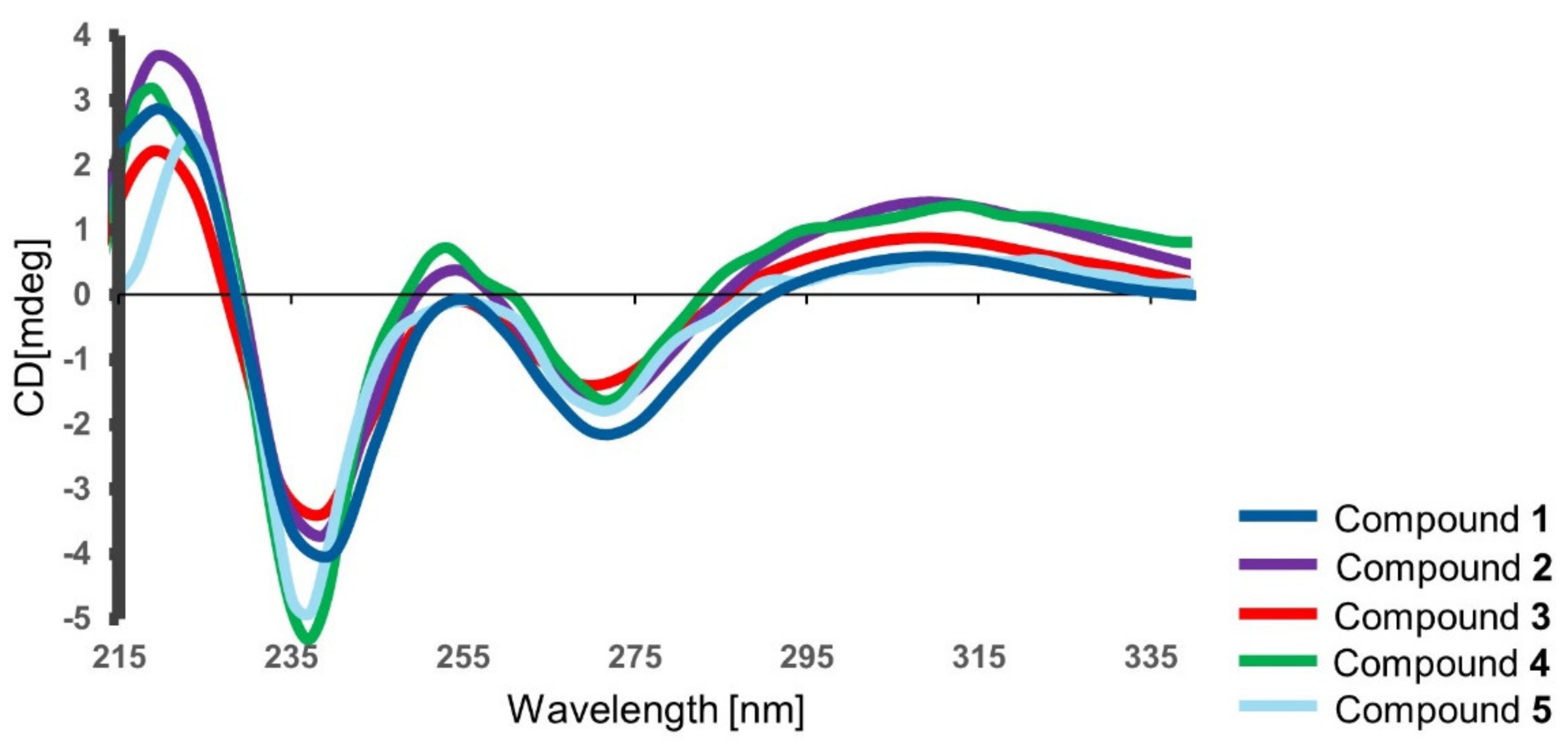
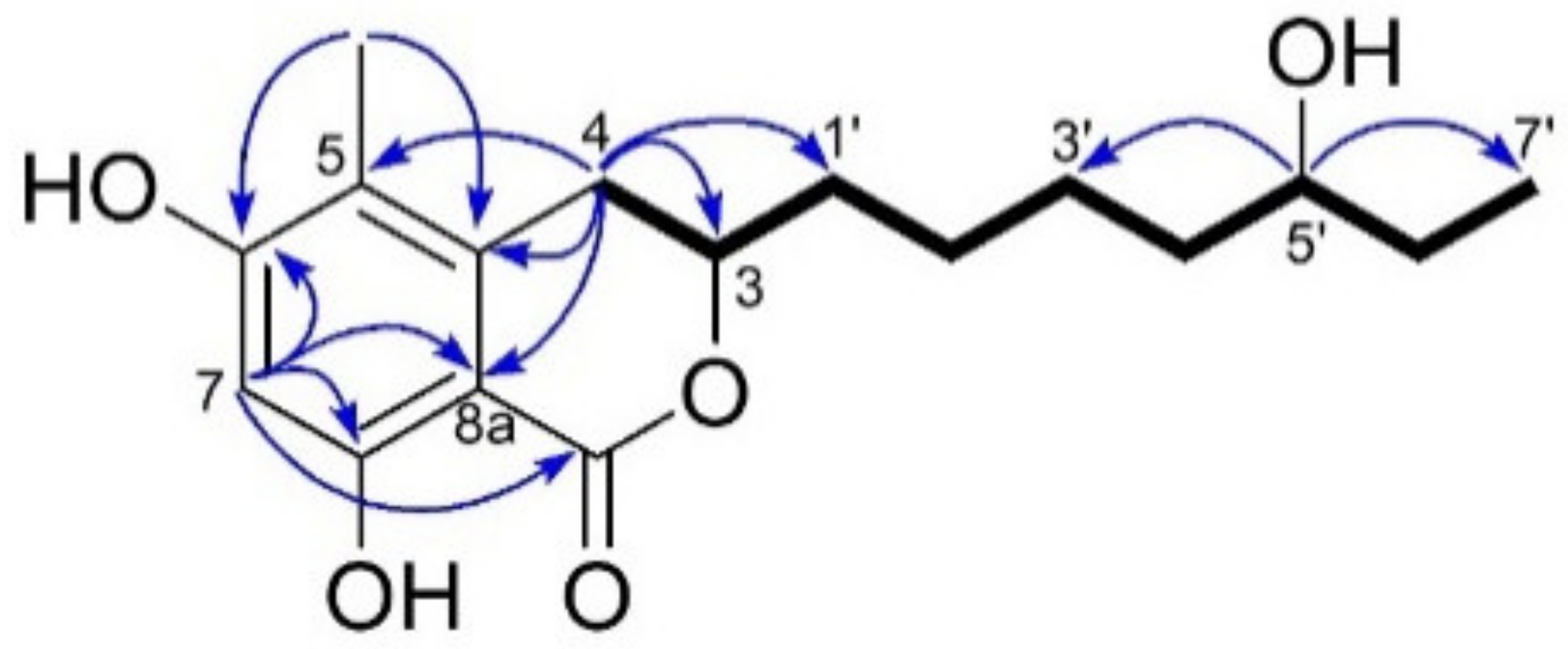
2.2. Isolation & Structure Elucidation
2.3. Bioactivity of Pure Compounds
2.4. Optimization Study to Enhance Production of the Soudanones
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Fungal Collection and Taxonomy
4.3. UPLC-QtoF-MS Analysis
4.4. Molecular Networking and Annotations
4.5. Large Scale Co-Cultivation and Isolation of Compounds
4.6. Co-Cultivation by Overlaid Inoculation on PDA and Extraction
4.7. Co-Cultivation in Liquid Broth (PDB)
4.8. Biological Assays
4.9. Mosher’s Esterification
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nofiani, R.; de Mattos-Shipley, K.; Lebe, K.E.; Han, L.-C.; Iqbal, Z.; Bailey, A.M.; Willis, C.L.; Simpson, T.J.; Cox, R.J. Strobilurin biosynthesis in Basidiomycete fungi. Nat. Commun. 2018, 9, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Sørensen, J.L.; Hansen, F.T.; Arvas, M.; Syed, M.F.; Hassan, L.; Benz, J.P.; Record, E.; Henrissat, B.; Pöggeler, S.; et al. Genome sequencing and analyses of two marine fungi from the North Sea unraveled a plethora of novel biosynthetic gene clusters. Sci. Rep. 2018, 8, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harwani, D.; Begani, J.; Lakhani, J. Co-cultivation strategies to induce de novo synthesis of novel chemical scaffolds from cryptic secondary metabolite gene clusters. In Fungi and their Role in Sustainable Development: Current Perspectives; Gehlot, P., Singh, J., Eds.; Springer: Singapore, 2018; pp. 617–631. [Google Scholar]
- Oppong-Danquah, E.; Parrot, D.; Blümel, M.; Labes, A.; Tasdemir, D. Molecular networking-based metabolome and bioactivity analyses of marine-adapted fungi co-cultivated with phytopathogens. Front. Microbiol. 2018, 9, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Serrano, R.; González-Menéndez, V.; Rodríguez, L.; Martín, J.; Tormo, J.R.; Genilloud, O. Co-culturing of fungal strains against Botrytis cinerea as a model for the induction of chemical diversity and therapeutic agents. Front. Microbiol. 2017, 8, 1–15. [Google Scholar] [CrossRef]
- Chaudhry, V.; Runge, P.; Sengupta, P.; Doehlemann, G.; Parker, J.E.; Kemen, E. Shaping the leaf microbiota: Plant–microbe–microbe interactions. J. Exp. Bot. 2020, 72, 36–56. [Google Scholar] [CrossRef]
- De Souza, E.M.; Granada, C.E.; Sperotto, R.A. Plant pathogens affecting the establishment of plant-symbiont Interaction. Front. Plant Sci. 2016, 7, 1–5. [Google Scholar] [CrossRef]
- Koyama, K.; Natori, S. Further characterization of seven bis (naphtho-γ-pyrone) congeners of ustilaginoidins, coloring matters of Claviceps virens (Ustilaginoidea virens). Chem. Pharm. Bull. 1988, 36, 146–152. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.; Wang, A.; Xu, D.; Wang, W.; Meng, J.; Dai, J.; Liu, Y.; Lai, D.; Zhou, L. New ustilaginoidins from rice false smut balls caused by Villosiclava virens and their phytotoxic and cytotoxic activities. J. Agric. Food Chem. 2017, 65, 5151–5160. [Google Scholar] [CrossRef]
- Kock, I.; Draeger, S.; Schulz, B.; Elsässer, B.; Kurtán, T.; Kenéz, Á.; Rheinheimer, J. Pseudoanguillosporin A and B: Two new isochromans isolated from the endophytic fungus Pseudoanguillospora sp. Eur. J. Org. Chem. 2009, 2009, 1427–1434. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, L.; Ren, W.; Zhao, D.; Zhu, Y.; Wu, X. Bioactive metabolites from Chaetomium globosum L18, an endophytic fungus in the medicinal plant Curcuma wenyujin. Phytomedicine 2012, 19, 364–368. [Google Scholar] [CrossRef]
- Heald, S.L.; Jeffs, P.W.; Wheat, R.W. The identification of ergosterol and Δ9(11)-dehydroergosterol from mycelia of Coccidioides immitis by reverse-phase high-performance liquid and gas chromatography and ultraviolet and mass spectrometry. Exp. Mycol. 1981, 5, 162–166. [Google Scholar] [CrossRef]
- El-Elimat, T.; Figueroa, M.; Ehrmann, B.M.; Cech, N.B.; Pearce, C.J.; Oberlies, N.H. High-resolution MS, MS/MS, and UV database of fungal secondary metabolites as a dereplication protocol for bioactive natural products. J. Nat. Prod. 2013, 76, 1709–1716. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Zhao, X.; Guo, M.; Liu, H.; Zheng, Z. Growth and metabolism of Beauveria bassiana spores and mycelia. BMC Microbiol. 2015, 15, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Kupchan, S.M.; Tsou, G.; Sigel, C.W. Datiscacin, a novel cytotoxic cucurbitacin 20-acetate from Datisca glomerata. J. Org. Chem. 1973, 38, 1420–1421. [Google Scholar] [CrossRef]
- Rusman, Y.; Held, B.W.; Blanchette, R.A.; Wittlin, S.; Salomon, C.E. Soudanones A-G: Antifungal isochromanones from the Ascomycetous fungus Cadophora sp. isolated from an iron mine. J. Nat. Prod. 2015, 78, 1456–1460. [Google Scholar] [CrossRef]
- Seco, J.M.; Quiñoá, E.; Riguera, R. The assignment of absolute aonfiguration by NMR. Chem. Rev. 2004, 104, 17–117. [Google Scholar] [CrossRef]
- Seki, M.; Mori, K. Synthesis of a prenylated and immunosuppressive marine galactosphingolipid with cyclopropane-containing alkyl chains: (2S,3R,11S,12R,2′′′R,5′′′Z,11′′′S,12′′′R)-plakoside A and its (2S,3R,11R,12S,2′′′R,5′′′Z,11′′′R,12′′′S) isomer. Eur. J. Org. Chem. 2001, 2001, 3797–3809. [Google Scholar] [CrossRef]
- Matsumoto, M.; Minato, H.; Kondo, E.; Mitsugi, T.; Katagiri, K. Cephalochromin, dihydroisoustilaginoidin A, and iso-ustilaginoidin A from Verticillium sp. K-113. J. Antibiot. 1975, 28, 602–604. [Google Scholar] [CrossRef] [Green Version]
- Sung, A.A.; Gromek, S.M.; Balunas, M.J. Upregulation and identification of antibiotic activity of a marine-derived Streptomyces sp. via co-cultures with human pathogens. Mar. Drugs 2017, 15, 250. [Google Scholar] [CrossRef] [Green Version]
- Pal, S.; Chatare, V.; Pal, M. Isocoumarin and its derivatives: An overview on their synthesis and applications. Curr. Org. Chem. 2011, 15, 782–800. [Google Scholar] [CrossRef]
- Noor, A.O.; Almasri, D.M.; Bagalagel, A.A.; Abdallah, H.M.; Mohamed, S.G.A.; Mohamed, G.A.; Ibrahim, S.R.M. Naturally occurring isocoumarins derivatives from endophytic fungi: Sources, isolation, structural characterization, biosynthesis, and biological activities. Molecules 2020, 25, 395. [Google Scholar] [CrossRef] [Green Version]
- Qadeer, G.; Rama, N.; Fan, Z.-J.; Liu, B.; Liu, X.-F. Synthesis, herbicidal, fungicidal and insecticidal evaluation of 3-(dichlorophenyl)- isocoumarins and (±)-3-(dichlorophenyl)-3,4-dihydroisocoumarins. J. Braz. Chem. Soc. 2007, 18, 1176–1182. [Google Scholar] [CrossRef] [Green Version]
- Shabir, G.; Saeed, A.; El-Seedi, H.R. Natural isocoumarins: Structural styles and biological activities, the revelations carry on. Phytochemistry 2021, 181, 1–23. [Google Scholar] [CrossRef]
- Netzker, T.; Flak, M.; Krespach, M.K.C.; Stroe, M.C.; Weber, J.; Schroeckh, V.; Brakhage, A.A. Microbial interactions trigger the production of antibiotics. Curr. Opin. Microbiol. 2018, 45, 117–123. [Google Scholar] [CrossRef]
- Zheng, C.J.; Sohn, M.J.; Lee, S.; Hong, Y.S.; Kwak, J.H.; Kim, W.G. Cephalochromin, a FabI-directed antibacterial of microbial origin. Biochem. Biophys. Res. Commun. 2007, 362, 1107–1112. [Google Scholar] [CrossRef]
- Lu, S.; Sun, W.; Meng, J.; Wang, A.; Wang, X.; Tian, J.; Fu, X.; Dai, J.; Liu, Y.; Lai, D.; et al. Bioactive bis-naphtho-γ-pyrones from rice false smut pathogen Ustilaginoidea virens. J. Agric. Food Chem. 2015, 63, 3501–3508. [Google Scholar] [CrossRef]
- Hsiao, C.-J.; Hsiao, G.; Chen, W.-L.; Wang, S.-W.; Chiang, C.-P.; Liu, L.-Y.; Guh, J.-H.; Lee, T.-H.; Chung, C.-L. Cephalochromin induces G0/G1 cell cycle arrest and apoptosis in A549 human non-small-cell lung cancer cells by inflicting mitochondrial disruption. J. Nat. Prod. 2014, 77, 758–765. [Google Scholar] [CrossRef]
- Oppong-Danquah, E.; Budnicka, P.; Blümel, M.; Tasdemir, D. Design of fungal co-cultivation based on comparative metabolomics and bioactivity for discovery of marine fungal agrochemicals. Mar. Drugs 2020, 18, 73. [Google Scholar] [CrossRef] [Green Version]
- Keller, N.P. Translating biosynthetic gene clusters into fungal armor and weaponry. Nat. Chem. Biol. 2015, 11, 671–677. [Google Scholar] [CrossRef] [Green Version]
- Klaubauf, S.; Tharreau, D.; Fournier, E.; Groenewald, J.Z.; Crous, P.W.; de Vries, R.P.; Lebrun, M.H. Resolving the polyphyletic nature of Pyricularia (Pyriculariaceae). Stud. Mycol. 2014, 79, 85–120. [Google Scholar] [CrossRef]
- Masi, M.; Santoro, E.; Clement, S.; Meyer, S.; Scafato, P.; Superchi, S.; Evidente, A. Further secondary metabolites produced by the fungus Pyricularia grisea isolated from buffelgrass (Cenchrus ciliaris). Chirality 2020, 32, 1234–1242. [Google Scholar] [CrossRef] [PubMed]
- Herrera, C.S.; Rossman, A.Y.; Samuels, G.J.; Pereira, O.L.; Chaverri, P. Systematics of the Cosmospora viliuscula species complex. Mycologia 2015, 107, 532–557. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Lai, D.; Zhou, L. Secondary metabolites from Acremonium fungi: Diverse structures and bioactivities. Mini-Rev. Med. Chem. 2017, 17, 603–632. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.H.; Lu, C.K.; Wang, G.J.; Chang, Y.C.; Yang, W.B.; Ju, Y.M. Sesquiterpene glycosides from Cosmospora joca. J. Nat. Prod. 2011, 74, 1561–1567. [Google Scholar] [CrossRef] [PubMed]
- Overy, D.; Calati, K.; Kahn, J.N.; Hsu, M.-J.; Martín, J.; Collado, J.; Roemer, T.; Harris, G.; Parish, C.A. Isolation and structure elucidation of parnafungins C and D, isoxazolidinone-containing antifungal natural products. Bioorg. Med. Chem. Lett. 2009, 19, 1224–1227. [Google Scholar] [CrossRef]
- Parish, C.A.; Smith, S.K.; Calati, K.; Zink, D.; Wilson, K.; Roemer, T.; Jiang, B.; Xu, D.; Bills, G.; Platas, G.; et al. Isolation and structure elucidation of parnafungins, antifungal natural products that Inhibit mRNA polyadenylation. J. Am. Chem. Soc. 2008, 130, 7060–7066. [Google Scholar] [CrossRef]
- Seo, C.; Sohn, J.H.; Oh, H.; Kim, B.Y.; Ahn, J.S. Isolation of the protein tyrosine phosphatase 1B inhibitory metabolite from the marine-derived fungus Cosmospora sp. SF-5060. Bioorg. Med. Chem. Lett. 2009, 19, 6095–6097. [Google Scholar] [CrossRef]
- Shiono, Y.; Miyazaki, N.; Murayama, T.; Koseki, T.; Harizon; Katja, D.G.; Supratman, U.; Nakata, J.; Kakihara, Y.; Saeki, M.; et al. GSK-3β inhibitory activities of novel dichroloresorcinol derivatives from Cosmospora vilior isolated from a mangrove plant. Phytochem. Lett. 2016, 18, 122–127. [Google Scholar] [CrossRef]
- Schroeckh, V.; Scherlach, K.; Nutzmann, H.W.; Shelest, E.; Schmidt-Heck, W.; Schuemann, J.; Martin, K.; Hertweck, C.; Brakhage, A.A. Intimate bacterial-fungal interaction triggers biosynthesis of archetypal polyketides in Aspergillus nidulans. Proc. Natl. Acad. Sci. USA 2009, 106, 14558–14563. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, D.D.; Wu, C.-H.; Moree, W.J.; Lamsa, A.; Medema, M.H.; Zhao, X.; Gavilan, R.G.; Aparicio, M.; Atencio, L.; Jackson, C.; et al. MS/MS networking guided analysis of molecule and gene cluster families. Proc. Natl. Acad. Sci. USA 2013, 110, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Chambers, M.C.; Maclean, B.; Burke, R.; Amodei, D.; Ruderman, D.L.; Neumann, S.; Gatto, L.; Fischer, B.; Pratt, B.; Egertson, J. A cross-platform toolkit for mass spectrometry and proteomics. Nat. Biotechnol. 2012, 30, 918–920. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Van Santen, J.A.; Jacob, G.; Singh, A.L.; Aniebok, V.; Balunas, M.J.; Bunsko, D.; Neto, F.C.; Castaño-Espriu, L.; Chang, C.; Clark, T.N.; et al. The Natural Products Atlas: An open access knowledge base for microbial natural products discovery. ACS Cent. Sci. 2019, 5, 1824–1833. [Google Scholar] [CrossRef] [Green Version]
- Buckingham, J. Dictionary of Natural Products (On-Line Web Edition). Available online: http://www.chemnetbase.com (accessed on 1 December 2021).
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.-M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis. Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [Green Version]
- Oppong-Danquah, E.; Passaretti, C.; Chianese, O.; Blümel, M.; Tasdemir, D. Mining the metabolome and the agricultural and pharmaceutical potential of sea foam-derived fungi. Mar. Drugs 2020, 18, 128. [Google Scholar] [CrossRef] [Green Version]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Toth, I.A.N. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar] [CrossRef] [Green Version]
- Dean, R.; Kan, J.A.V.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Pietro, A.D.; Spanu, P.D.; Foster, G.D. The top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | 1 a | 1 b | 2 b | 3 b | 4 b | 5 b |
|---|---|---|---|---|---|---|
| 3 | 4.47 m | 4.46 m | 4.45 m | 4.45 m | 4.47 m | 4.46 m |
| 4 | 3.03 dd (16.7, 3.4) 2.69 dd (16.7,11.5) | 2.94 dd (16.5, 3.4) 2.70 dd (16.5, 11.6) | 2.93 dd (16.5, 3.2) 2.69 dd (16.5, 11.8) | 2.93 dd (16.5, 3.3) 2.69 dd (16.5, 11.7) | 2.94 dd (16.5, 3.2) 2.71 dd (16.5, 11.8) | 2.94 dd (16.5, 3.3) 2.70 dd (16.5, 11.7) |
| 7 | 6.24 s | 6.31 s | 6.31 s | 6.30 s | 6.29 s | 6.3 s |
| 1’ | 1.83 m 1.76 m | 1.88 m 1.73 m | 1.87 m 1.73 m | 1.86 m 1.76 m | 1.90 m 1.75 m | 1.90 m 1.76 m |
| 2’ | 1.58 m 1.48 m | 1.56 m 1.45 m | 1.60 m 1.48 m | 1.62 m 1.48 m | 1.64 m 1.52 m | 1.64 m 1.53 m |
| 3’ | 1.35 m | 1.34 m | 1.37 m | 1.36 m | 1.52 m 1.42 m | 1.70 m |
| 4’ | 1.35 m | 1.32 m | 1.36 m 1.44 m | 1.62 m | 1.53 m 1.45 m | 2.36 t (7.3) |
| 5’ | 1.35 m | 1.26 m | 1.45 m | 2.45 t (7.3) | 3.54 m | |
| 6’ | 1.35 m | 1.29 m | 3.81 m | 1.52 m 1.44 m | 3.68 s | |
| 7’ | 0.91 t (6.9) | 0.88 t (6.9) | 1.20 d (6.1) | 2.15 s | 0.95 t (7.5) | |
| 5-Me | 2.04 s | 2.07 s | 2.06 s | 2.06 s | 2.07 s | 2.07 s |
| OH | 11.26 s | 11.26 s | 11.18 s | 11.26 s | 11.26 s |
| Position | 1 | 2 | 3 | 4 | 5 |
|---|---|---|---|---|---|
| 1 | 170.6, C | 170.6, C | 170.5, C | 170.5, C | 170.4, C |
| 3 | 78.7, CH | 78.6, CH | 78.5, CH | 78.5, CH | 78.3, CH |
| 4 | 30.6, CH2 | 30.7, CH2 | 30.7, CH2 | 30.7, CH2 | 30.6, CH2 |
| 4a | 139.5, C | 139.4, C | 139.4, C | 139.5, C | 139.4, C |
| 5 | 113.1, C | 113.2, C | 113.0, C | 112.9, C | 113.0, C |
| 6 | 160.9, C | 161.0, C | 160.7, C | 160.5, C | 160.6, C |
| 7 | 101.6, CH | 101.6, CH | 101.6, CH | 101.6, CH | 101.7, CH |
| 8 | 162.5, C | 162.5, C | 162.5, C | 162.5, C | 162.6, C |
| 8a | 102.1, C | 102.0, C | 102.1, C | 102.2, C | 102.2, C |
| 1’ | 35.1, CH2 | 35.0, CH2 | 34.9, CH2 | 35.1, CH2 | 34.8, CH2 |
| 2’ | 25.1, CH2 | 25.0, CH2 | 24.9, CH2 | 25.6, CH2 | 24.6, CH2 |
| 3’ | 29.5, CH2 | 29.4, CH2 | 29.0, CH2 | 25.2, CH2 | 24.8, CH2 |
| 4’ | 29.3, CH2 | 25.7, CH2 | 23.6, CH2 | 36.8, CH2 | 34.0, CH3 |
| 5’ | 31.9, CH2 | 39.2, CH2 | 43.7, CH2 | 73.3, CH | 174.1, C |
| 6’ | 22.8, CH2 | 68.4, CH | 209.3, C | 30.4, CH2 | 51.7, CH3 |
| 7’ | 14.2, CH3 | 23.7, CH3 | 30.1, CH3 | 10.0, CH3 | |
| 5-Me | 10.6, CH3 | 10.6, CH3 | 10.6, CH3 | 10.6, CH3 | 10.6, CH3 |
| Compound | Bacteria | Oomycete | Fungus | |||
|---|---|---|---|---|---|---|
| Ps | Xc | Ea | Rs | Pi | Mo | |
| 1 | >100 | >100 | >100 | >100 | >100 | >100 |
| 2 | >100 | 71.5 | >100 | >100 | 27.6 | 12.8 |
| 3 | >100 | 15.7 | >100 | >100 | 52.1 | 60.3 |
| 6 | 23.4 | 7.4 | >100 | >100 | 3.2 | 0.8 |
| 7 | >100 | 67.1 | >100 | 42.2 | >100 | >100 |
| 8 | 95.7 | 12.1 | >100 | 27.6 | 2.3 | >100 |
| 9 | >100 | 21.7 | >100 | >100 | 7.2 | >100 |
| 10 | >100 | >100 | >100 | >100 | >100 | >100 |
| Control | 0.4 | 0.5 | 0.2 | 1.0 | 0.6 | 0.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oppong-Danquah, E.; Blümel, M.; Scarpato, S.; Mangoni, A.; Tasdemir, D. Induction of Isochromanones by Co-Cultivation of the Marine Fungus Cosmospora sp. and the Phytopathogen Magnaporthe oryzae. Int. J. Mol. Sci. 2022, 23, 782. https://doi.org/10.3390/ijms23020782
Oppong-Danquah E, Blümel M, Scarpato S, Mangoni A, Tasdemir D. Induction of Isochromanones by Co-Cultivation of the Marine Fungus Cosmospora sp. and the Phytopathogen Magnaporthe oryzae. International Journal of Molecular Sciences. 2022; 23(2):782. https://doi.org/10.3390/ijms23020782
Chicago/Turabian StyleOppong-Danquah, Ernest, Martina Blümel, Silvia Scarpato, Alfonso Mangoni, and Deniz Tasdemir. 2022. "Induction of Isochromanones by Co-Cultivation of the Marine Fungus Cosmospora sp. and the Phytopathogen Magnaporthe oryzae" International Journal of Molecular Sciences 23, no. 2: 782. https://doi.org/10.3390/ijms23020782
APA StyleOppong-Danquah, E., Blümel, M., Scarpato, S., Mangoni, A., & Tasdemir, D. (2022). Induction of Isochromanones by Co-Cultivation of the Marine Fungus Cosmospora sp. and the Phytopathogen Magnaporthe oryzae. International Journal of Molecular Sciences, 23(2), 782. https://doi.org/10.3390/ijms23020782