

Insights into Muscle Contraction Derived from the Effects of Small-Molecular Actomyosin-Modulating Compounds


Abstract
:1. Introduction
2. Results
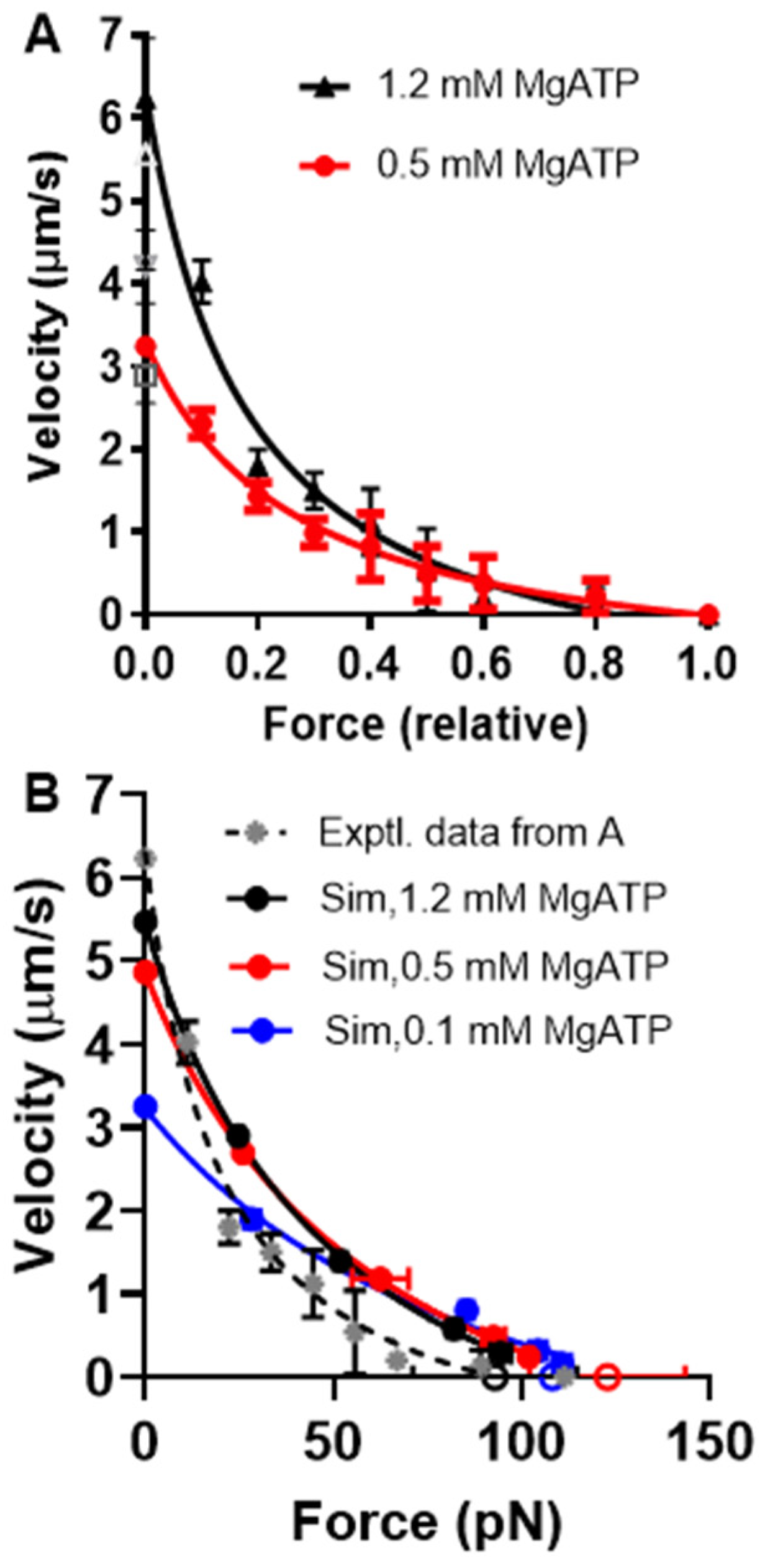
2.1. Simulation of Force–Velocity relationships under Physiological Conditions
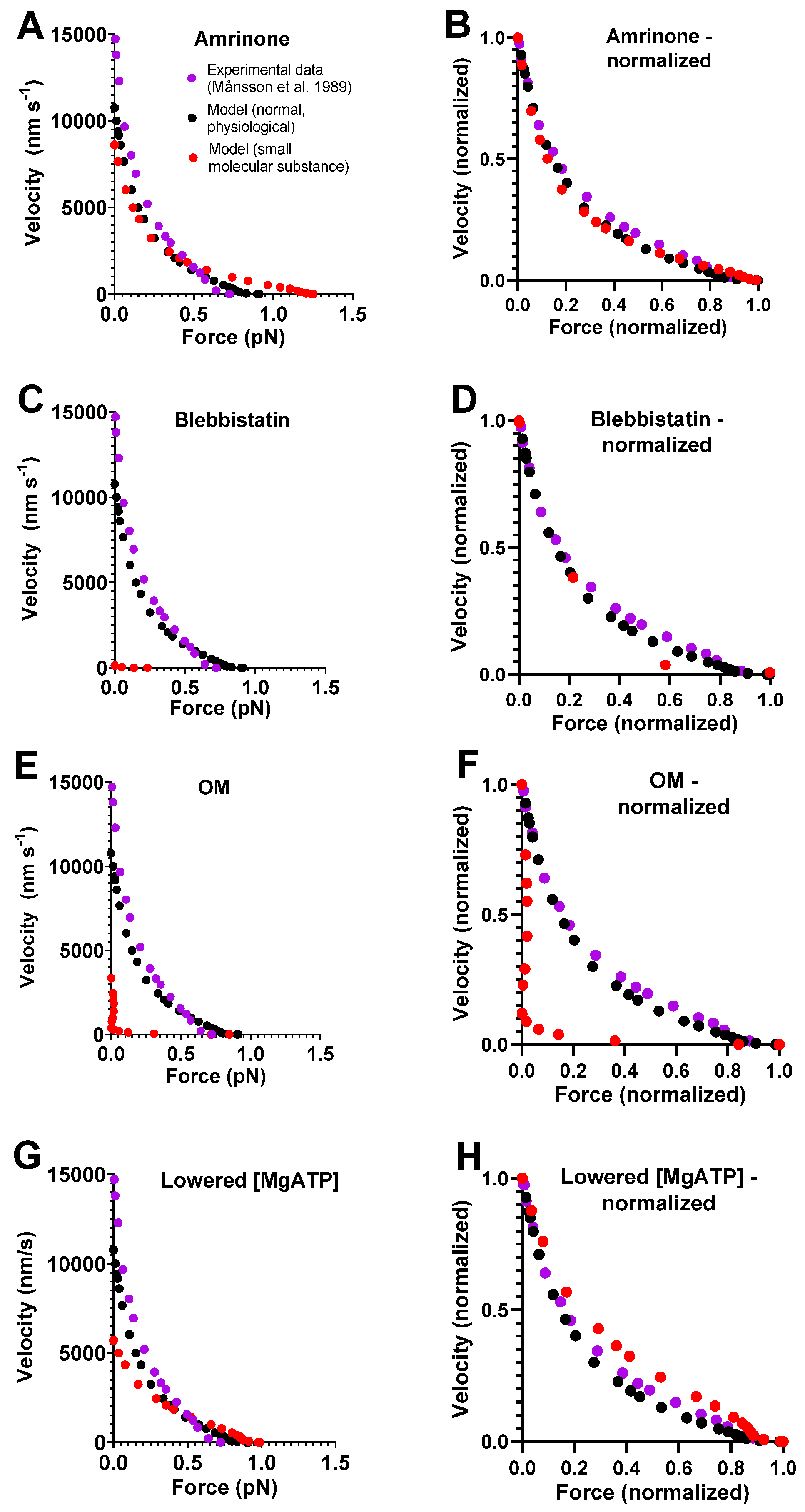
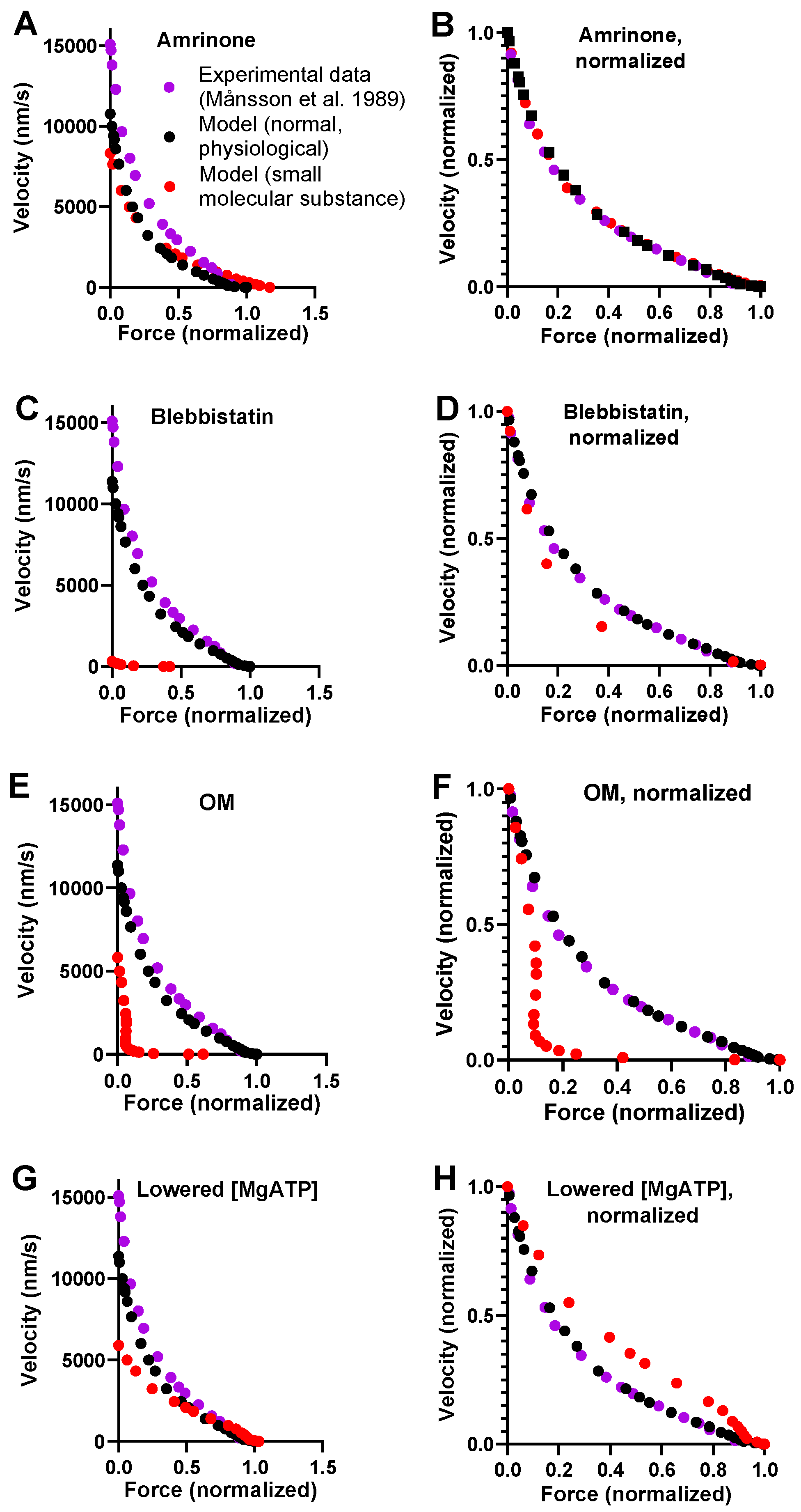
2.2. Effects of Small Molecular Compounds on the Force–Velocity Relationship
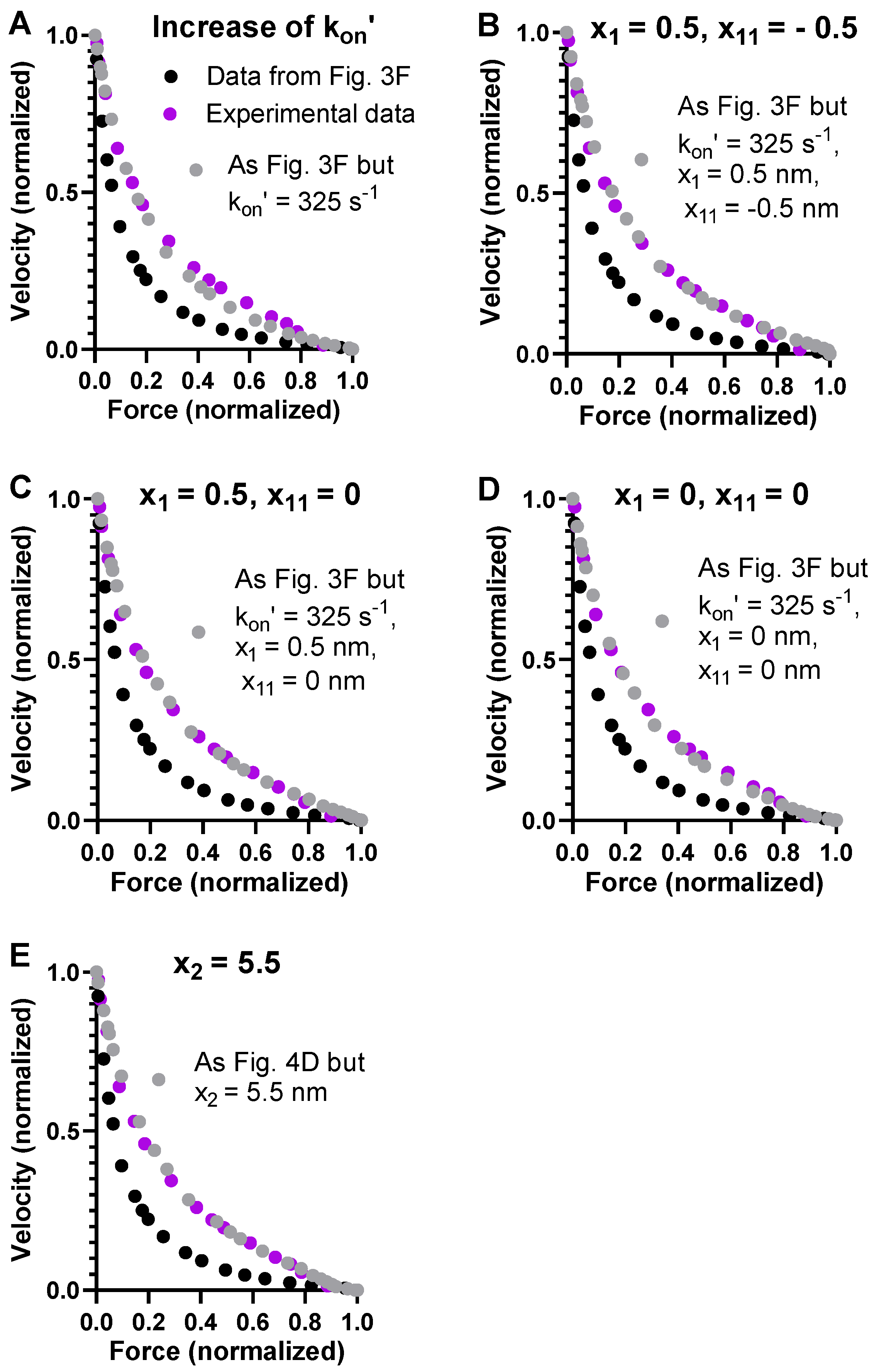
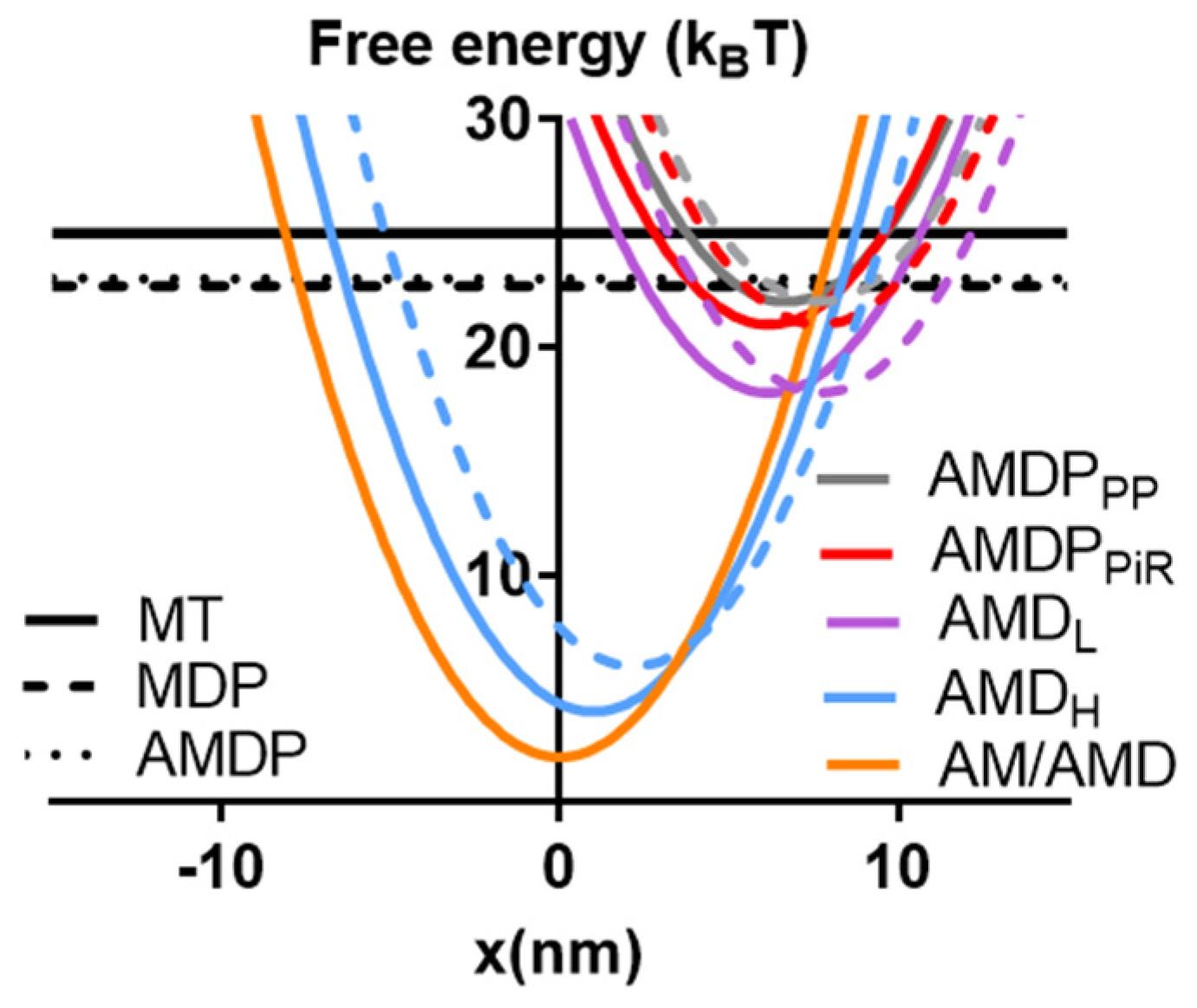
2.3. Towards an Optimized Model
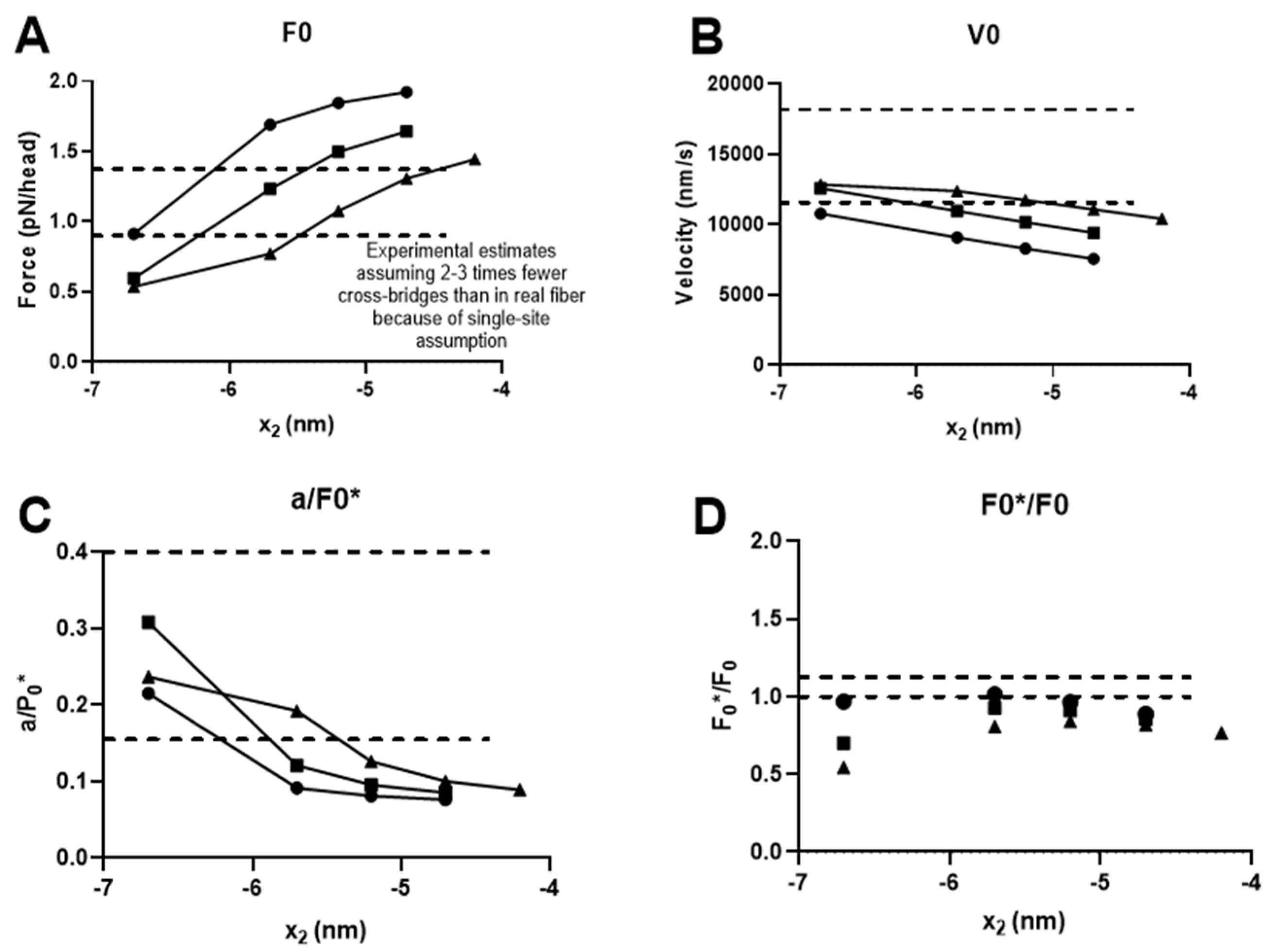
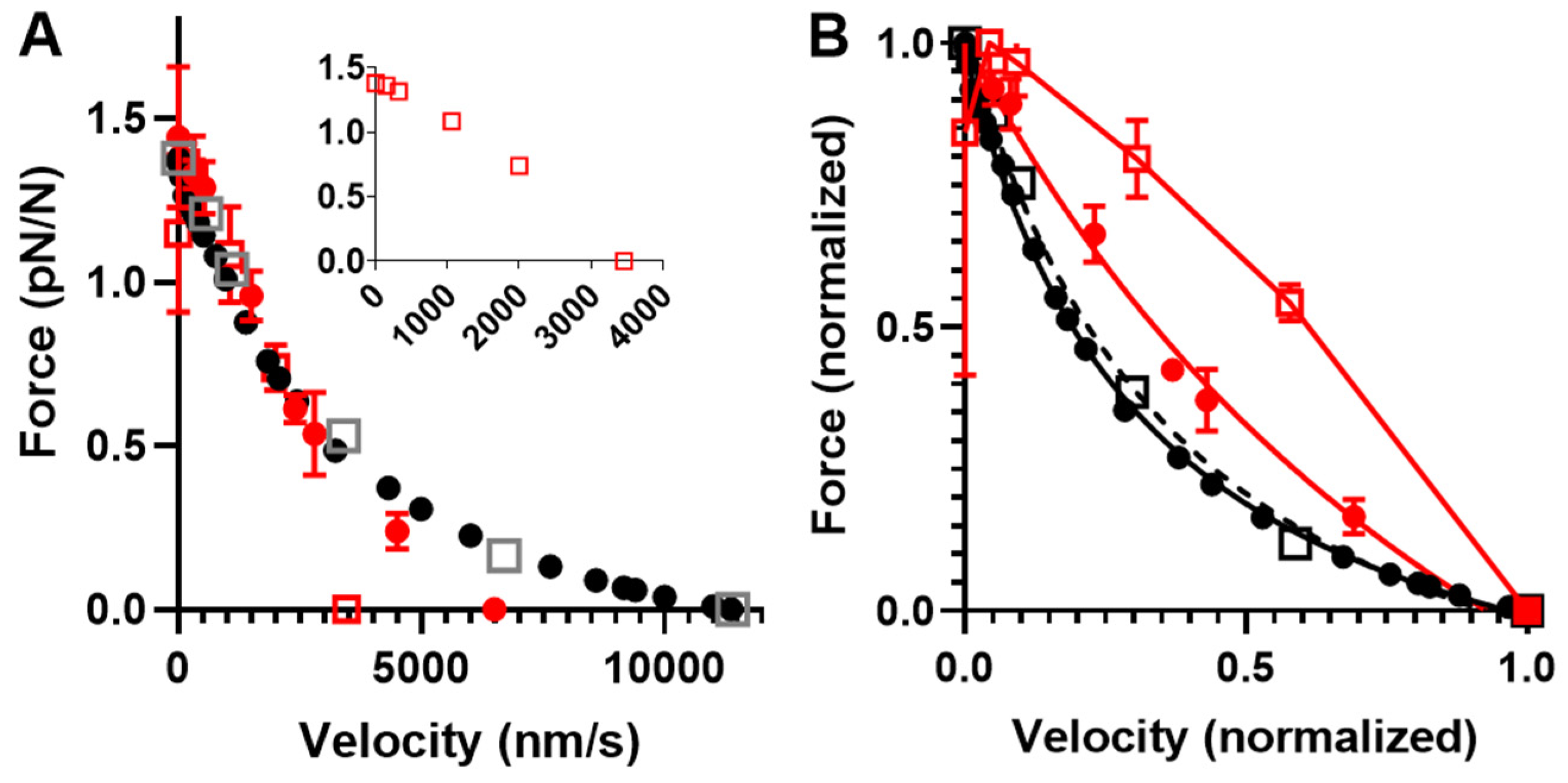
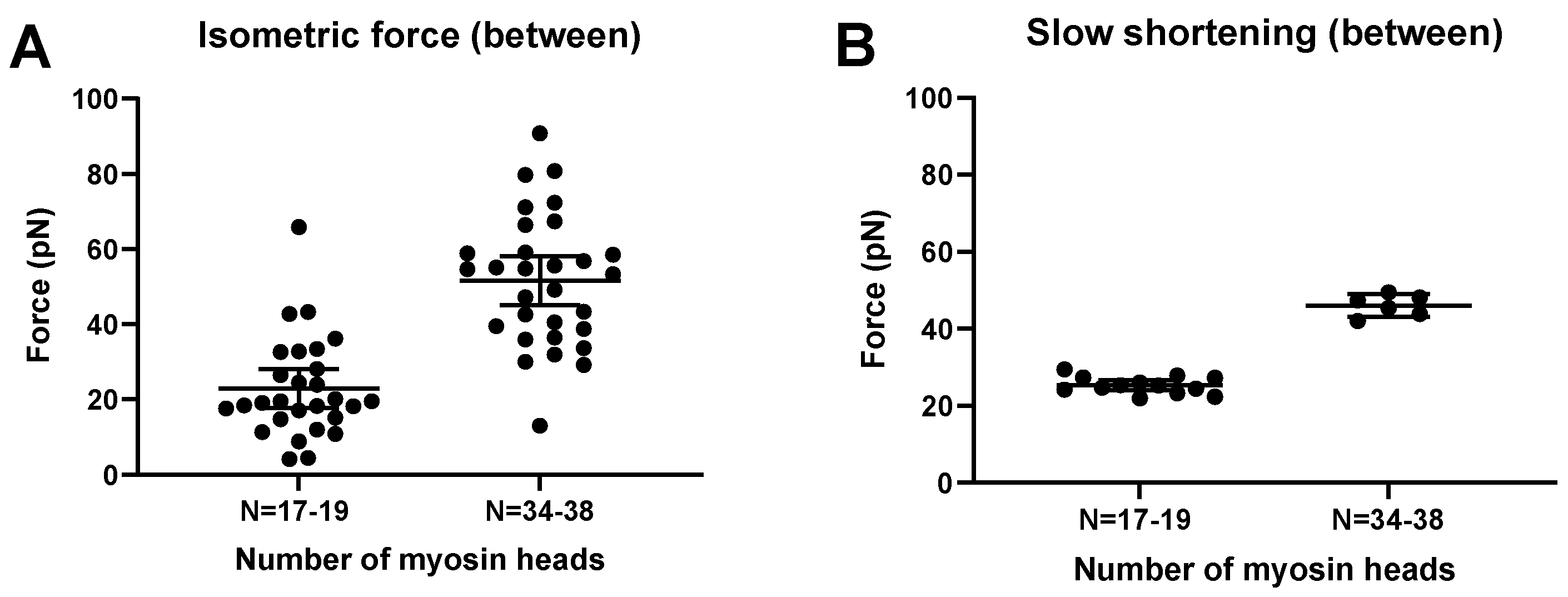
2.4. Prediction of Ensemble Contractile Function with and without Small Molecular Compounds Using Optimized Model
3. Discussion
3.1. Summary and Main Implications
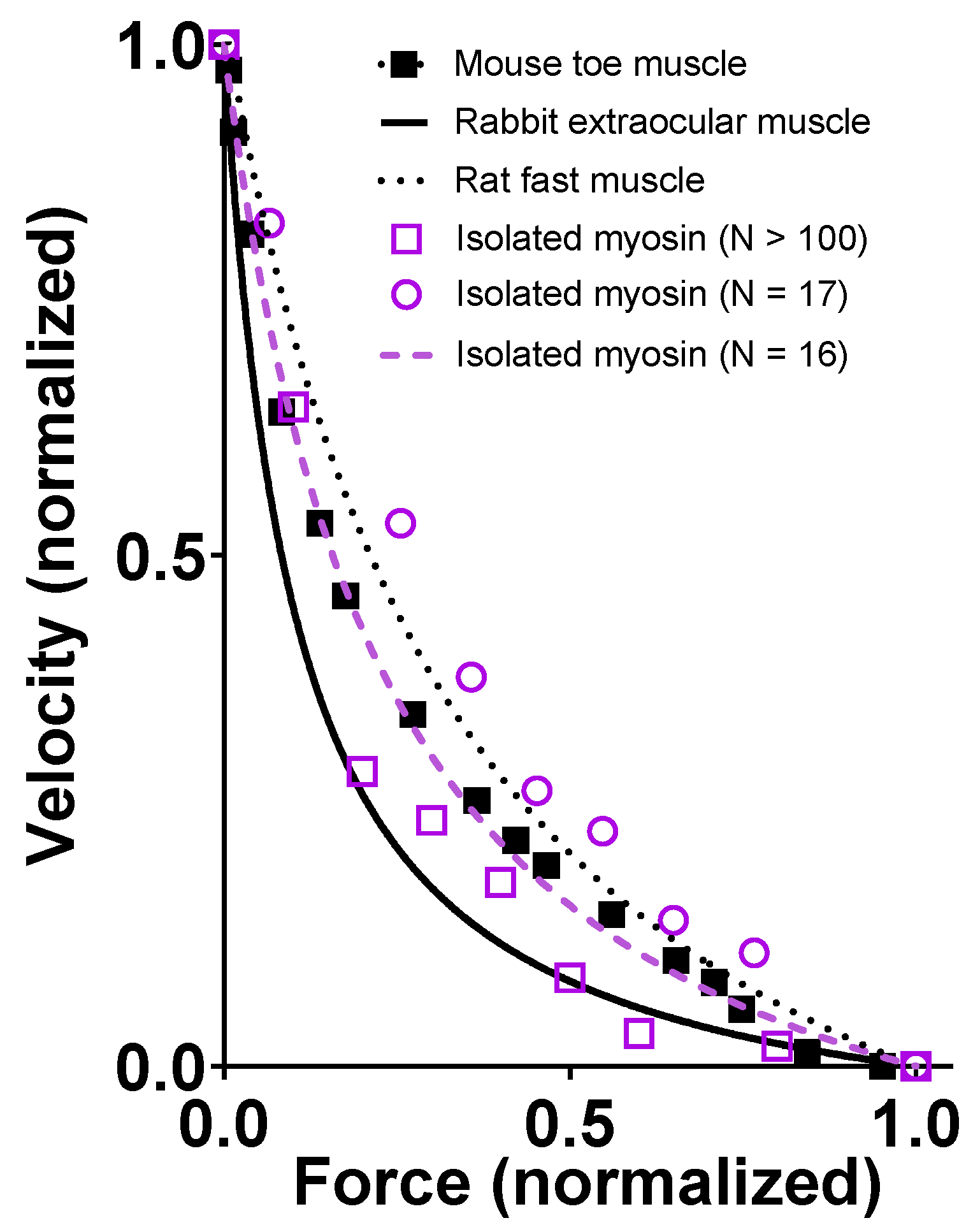
3.2. Comparison with Previous Studies
3.3. The Need to Assume Higher Cross-Bridge Attachment Rate during Shortening
3.4. Monte Carlo Simulations
3.5. Limitations
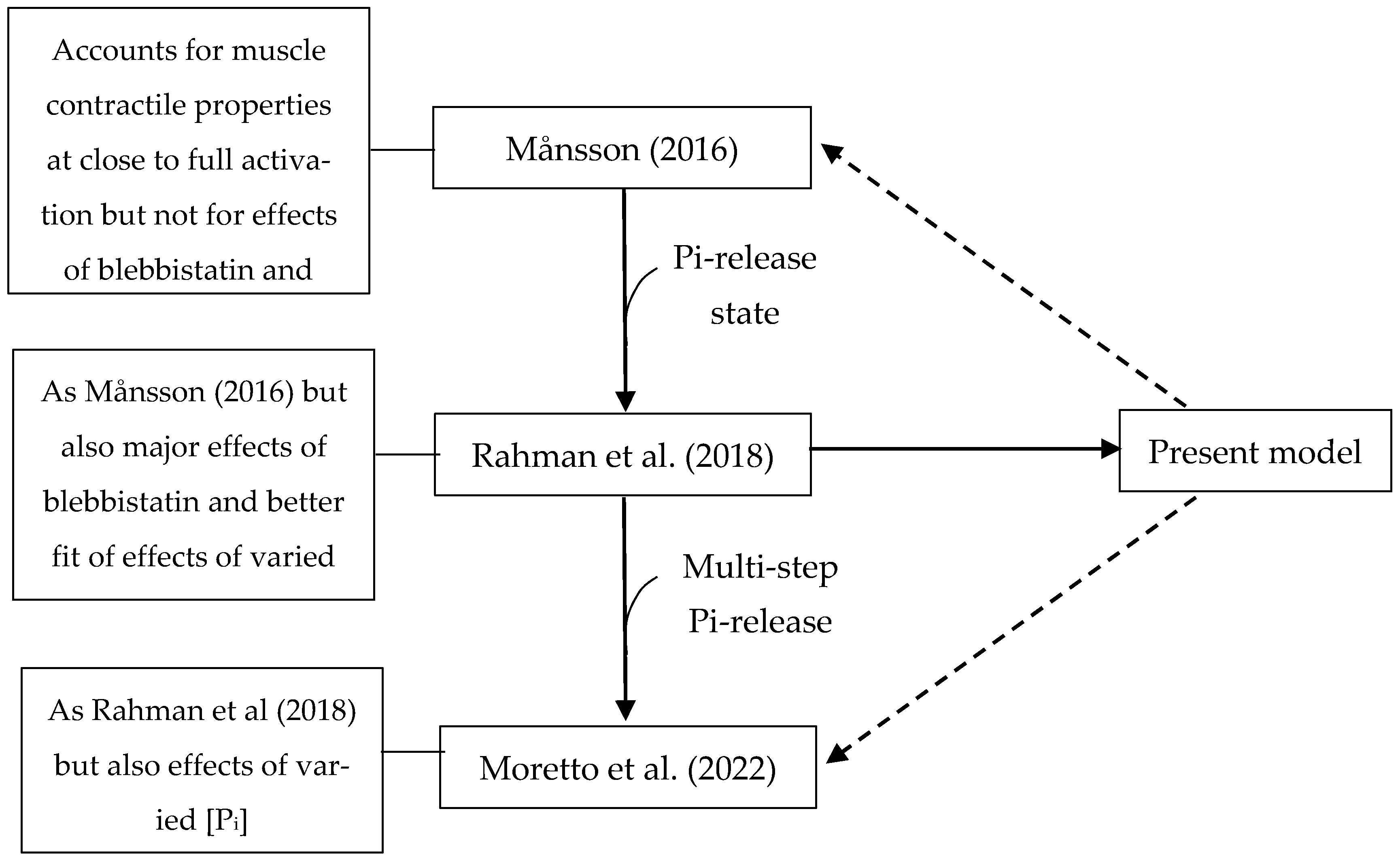
3.6. Relation to Other Similar Models
4. Materials and Methods
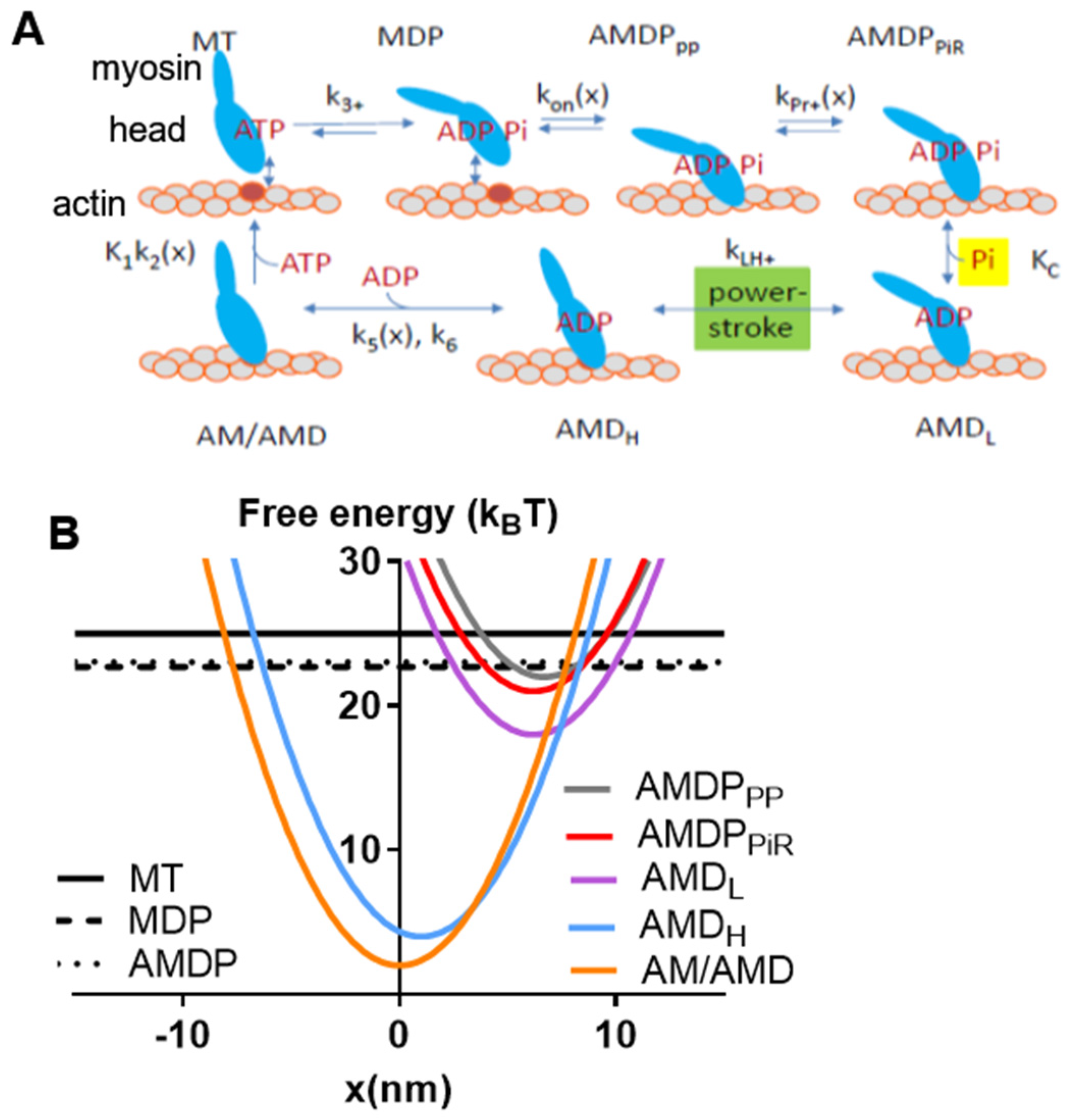
4.1. Modelling—General
4.2. Solutions of Ordinary Differential Equations and Derivation of Simulated Muscle Properties from State Probabilities
4.3. Monte Carlo Simulations
4.4. Fit of Simulated and Experimental Data to Hill (1938) Hyperbolic Equation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huxley, A.F. Muscle structure and theories of contraction. Prog. Biophys. Biophys. Chem. 1957, 7, 255–318. [Google Scholar] [CrossRef]
- Hill, T.L. Theoretical formalism for the sliding filament model of contraction of striated muscle. Part I. Prog. Biophys. Mol. Biol. 1974, 28, 267–340. [Google Scholar] [CrossRef]
- Hill, T.L. Theoretical formalism for the sliding filament model of contraction of striated muscle. Part II. Prog. Biophys. Mol. Biol. 1975, 29, 105–159. [Google Scholar] [CrossRef]
- Eisenberg, E.; Hill, T.L. A cross-bridge model of muscle contraction. Prog. Biophys. Mol. Biol. 1978, 33, 55–82. [Google Scholar] [CrossRef]
- Mansson, A. Actomyosin based contraction: One mechanokinetic model from single molecules to muscle? J. Muscle Res. Cell Motil. 2016, 37, 181–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.A.; Usaj, M.; Rassier, D.E.; Mansson, A. Blebbistatin Effects Expose Hidden Secrets in the Force-Generating Cycle of Actin and Myosin. Biophys. J. 2018, 115, 386–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansson, A. Comparing models with one versus multiple myosin-binding sites per actin target zone: The power of simplicity. J. Gen. Physiol. 2019, 151, 578–592. [Google Scholar] [CrossRef] [Green Version]
- Mansson, A.; Persson, M.; Shalabi, N.; Rassier, D.E. Non-linear actomyosin elasticity in muscle? Biophys. J. 2019, 116, 330–346. [Google Scholar] [CrossRef] [Green Version]
- Mansson, A. Hypothesis: Single Actomyosin Properties Account for Ensemble Behavior in Active Muscle Shortening and Isometric Contraction. Int. J. Mol. Sci. 2020, 21, 8399. [Google Scholar] [CrossRef]
- Mansson, A.; Usaj, M.; Moretto, L.; Rassier, D.E. Do Actomyosin Single-Molecule Mechanics Data Predict Mechanics of Contracting Muscle? Int. J. Mol. Sci. 2018, 19, 1863. [Google Scholar] [CrossRef]
- Alousi, A.A.; Farah, A.E.; Lesher, G.Y.; Opalka, C.J., Jr. Cardiotonic activity of amrinone—Win 40680 [5-amino-3,4’-bipyridine-6(1H)-one]. Circ. Res. 1979, 45, 666–677. [Google Scholar] [CrossRef] [Green Version]
- Mansson, A.; Edman, K.A. Effects of amrinone on the contractile behaviour of frog striated muscle fibres. Acta Physiol. Scand. 1985, 125, 481–493. [Google Scholar] [CrossRef]
- Mansson, A.; Morner, J.; Edman, K.A. Effects of amrinone on twitch, tetanus and shortening kinetics in mammalian skeletal muscle. Acta Physiol. Scand. 1989, 136, 37–45. [Google Scholar] [CrossRef]
- Morner, S.E.; Mansson, A. Effects of amrinone on the electromechanical coupling in frog skeletal muscle fibres. Acta Physiol. Scand. 1990, 139, 289–295. [Google Scholar] [CrossRef]
- Klinth, J.; Arner, A.; Mansson, A. Cardiotonic bipyridine amrinone slows myosin-induced actin filament sliding at saturating [MgATP]. J. Muscle Res. Cell Motil. 2003, 24, 15–32. [Google Scholar] [CrossRef]
- Albet-Torres, N.; Bloemink, M.J.; Barman, T.; Candau, R.; Frölander, K.; Geeves, M.A.; Golker, K.; Herrmann, C.; Lionne, C.; Piperio, C.; et al. Drug effect unveils inter-head cooperativity and strain-dependent ADP release in fast skeletal actomyosin. J. Biol. Chem. 2009, 284, 22926–22937. [Google Scholar] [CrossRef] [Green Version]
- Persson, M.; Bengtsson, E.; ten Siethoff, L.; Mansson, A. Nonlinear cross-bridge elasticity and post-power-stroke events in fast skeletal muscle actomyosin. Biophys. J. 2013, 105, 1871–1881. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Ajtai, K.; Burghardt, T.P. Analytical comparison of natural and pharmaceutical ventricular myosin activators. Biochemistry 2014, 53, 5298–5306. [Google Scholar] [CrossRef] [Green Version]
- Aksel, T.; Yu, E.C.; Sutton, S.; Ruppel, K.M.; Spudich, J.A. Ensemble force changes that result from human cardiac myosin mutations and a small-molecule effector. Cell Rep. 2015, 11, 910–920. [Google Scholar] [CrossRef] [Green Version]
- Winkelmann, D.A.; Forgacs, E.; Miller, M.T.; Stock, A.M. Structural basis for drug-induced allosteric changes to human beta-cardiac myosin motor activity. Nat. Commun. 2015, 6, 7974. [Google Scholar] [CrossRef]
- Planelles-Herrero, V.J.; Hartman, J.J.; Robert-Paganin, J.; Malik, F.I.; Houdusse, A. Mechanistic and structural basis for activation of cardiac myosin force production by omecamtiv mecarbil. Nat. Commun. 2017, 8, 190. [Google Scholar] [CrossRef] [Green Version]
- Rohde, J.A.; Thomas, D.D.; Muretta, J.M. Heart failure drug changes the mechanoenzymology of the cardiac myosin powerstroke. Proc. Natl. Acad. Sci. USA 2017, 114, E1796–E1804. [Google Scholar] [CrossRef] [Green Version]
- Swenson, A.M.; Tang, W.; Blair, C.A.; Fetrow, C.M.; Unrath, W.C.; Previs, M.J.; Campbell, K.S.; Yengo, C.M. Omecamtiv Mecarbil Enhances the Duty Ratio of Human beta-Cardiac Myosin Resulting in Increased Calcium Sensitivity and Slowed Force Development in Cardiac Muscle. J. Biol. Chem. 2017, 292, 3768–3778. [Google Scholar] [CrossRef] [Green Version]
- Woody, M.S.; Greenberg, M.J.; Barua, B.; Winkelmann, D.A.; Goldman, Y.E.; Ostap, E.M. Positive cardiac inotrope omecamtiv mecarbil activates muscle despite suppressing the myosin working stroke. Nat. Commun. 2018, 9, 3838. [Google Scholar] [CrossRef] [Green Version]
- Governali, S.; Caremani, M.; Gallart, C.; Pertici, I.; Stienen, G.; Piazzesi, G.; Ottenheijm, C.; Lombardi, V.; Linari, M. Orthophosphate increases the efficiency of slow muscle-myosin isoform in the presence of omecamtiv mecarbil. Nat. Commun. 2020, 11, 3405. [Google Scholar] [CrossRef]
- Cleland, J.G.; Teerlink, J.R.; Senior, R.; Nifontov, E.M.; Mc Murray, J.J.; Lang, C.C.; Tsyrlin, V.A.; Greenberg, B.H.; Mayet, J.; Francis, D.P.; et al. The effects of the cardiac myosin activator, omecamtiv mecarbil, on cardiac function in systolic heart failure: A double-blind, placebo-controlled, crossover, dose-ranging phase 2 trial. Lancet 2011, 378, 676–683. [Google Scholar] [CrossRef]
- Malik, F.I.; Hartman, J.J.; Elias, K.A.; Morgan, B.P.; Rodriguez, H.; Brejc, K.; Anderson, R.L.; Sueoka, S.H.; Lee, K.H.; Finer, J.T.; et al. Cardiac myosin activation: A potential therapeutic approach for systolic heart failure. Science 2011, 331, 1439–1443. [Google Scholar] [CrossRef] [Green Version]
- Straight, A.F.; Cheung, A.; Limouze, J.; Chen, I.; Westwood, N.J.; Sellers, J.R.; Mitchison, T.J. Dissecting temporal and spatial control of cytokinesis with a myosin II Inhibitor. Science 2003, 299, 1743–1747. [Google Scholar] [CrossRef] [Green Version]
- Ramamurthy, B.; Yengo, C.M.; Straight, A.F.; Mitchison, T.J.; Sweeney, H.L. Kinetic mechanism of blebbistatin inhibition of nonmuscle myosin IIb. Biochemistry 2004, 43, 14832–14839. [Google Scholar] [CrossRef]
- Minozzo, F.C.; Rassier, D.E. Effects of blebbistatin and Ca2+ concentration on force produced during stretch of skeletal muscle fibers. Am. J. Physiol. Cell Physiol. 2010, 299, C1127–C1135. [Google Scholar] [CrossRef]
- Kovacs, M.; Toth, J.; Hetenyi, C.; Malnasi-Csizmadia, A.; Sellers, J.R. Mechanism of blebbistatin inhibition of myosin II. J. Biol. Chem. 2004, 279, 35557–35563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allingham, J.S.; Smith, R.; Rayment, I. The structural basis of blebbistatin inhibition and specificity for myosin II. Nat. Struct. Mol. Biol. 2005, 12, 378–379. [Google Scholar] [CrossRef] [PubMed]
- Takacs, B.; Billington, N.; Gyimesi, M.; Kintses, B.; Malnasi-Csizmadia, A.; Knight, P.J.; Kovacs, M. Myosin complexed with ADP and blebbistatin reversibly adopts a conformation resembling the start point of the working stroke. Proc. Natl. Acad. Sci. USA 2010, 107, 6799–6804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornachione, A.S.; Rassier, D.E. A non-cross-bridge, static tension is present in permeabilized skeletal muscle fibers after active force inhibition or actin extraction. Am. J. Physiol. Cell Physiol. 2012, 302, C566–C574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauscher, A.A.; Gyimesi, M.; Kovacs, M.; Malnasi-Csizmadia, A. Targeting Myosin by Blebbistatin Derivatives: Optimization and Pharmacological Potential. Trends Biochem. Sci. 2018, 43, 700–713. [Google Scholar] [CrossRef] [PubMed]
- Gyimesi, M.; Horvath, A.I.; Turos, D.; Suthar, S.K.; Penzes, M.; Kurdi, C.; Canon, L.; Kikuti, C.; Ruppel, K.M.; Trivedi, D.V.; et al. Single Residue Variation in Skeletal Muscle Myosin Enables Direct and Selective Drug Targeting for Spasticity and Muscle Stiffness. Cell 2020, 183, 335–346.e13. [Google Scholar] [CrossRef] [PubMed]
- Stewart, M.; Franks-Skiba, K.; Cooke, R. Myosin regulatory light chain phosphorylation inhibits shortening velocities of skeletal muscle fibers in the presence of the myosin inhibitor blebbistatin. J. Muscle Res. Cell Motil. 2009, 30, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Kampourakis, T.; Zhang, X.; Sun, Y.B.; Irving, M. Omecamtiv mercabil and blebbistatin modulate cardiac contractility by perturbing the regulatory state of the myosin filament. J. Physiol. 2018, 596, 31–46. [Google Scholar] [CrossRef]
- Edman, K.A.; Mansson, A.; Caputo, C. The biphasic force-velocity relationship in frog muscle fibres and its evaluation in terms of cross-bridge function. J. Physiol. 1997, 503, 141–156. [Google Scholar] [CrossRef]
- Mansson, A. Actomyosin-ADP states, inter-head cooperativity and the force-velocity relation of skeletal muscle. Biophys. J. 2010, 98, 1237–1246. [Google Scholar] [CrossRef]
- Piazzesi, G.; Francini, F.; Linari, M.; Lombardi, V. Tension transients during steady lengthening of tetanized muscle fibres of the frog. J. Physiol. (Lond). 1992, 445, 659–711. [Google Scholar] [CrossRef]
- Adamovic, I.; Mijailovich, S.M.; Karplus, M. The elastic properties of the structurally characterized myosin II S2 subdomain: A molecular dynamics and normal mode analysis. Biophys. J. 2008, 94, 3779–3789. [Google Scholar] [CrossRef] [Green Version]
- Kaya, M.; Higuchi, H. Nonlinear elasticity and an 8-nm working stroke of single myosin molecules in myofilaments. Science 2010, 329, 686–689. [Google Scholar] [CrossRef] [Green Version]
- Kaya, M.; Tani, Y.; Washio, T.; Hisada, T.; Higuchi, H. Coordinated force generation of skeletal myosins in myofilaments through motor coupling. Nat. Commun. 2017, 8, 16036. [Google Scholar] [CrossRef]
- Hwang, Y.; Washio, T.; Hisada, T.; Higuchi, H.; Kaya, M. A reverse stroke characterizes the force generation of cardiac myofilaments, leading to an understanding of heart function. Proc. Natl. Acad. Sci. USA 2021, 118, e2011659118. [Google Scholar] [CrossRef]
- Llinas, P.; Isabet, T.; Song, L.; Ropars, V.; Zong, B.; Benisty, H.; Sirigu, S.; Morris, C.; Kikuti, C.; Safer, D.; et al. How actin initiates the motor activity of Myosin. Dev. Cell 2015, 33, 401–412. [Google Scholar] [CrossRef] [Green Version]
- Mansson, A. Changes in force and stiffness during stretch of skeletal muscle fibers, effects of hypertonicity. Biophys. J. 1989, 56, 429–433. [Google Scholar] [CrossRef] [Green Version]
- Ranatunga, K.W. The force-velocity relation of rat fast- and slow-twitch muscles examined at different temperatures. J. Physiol. 1984, 351, 517–529. [Google Scholar] [CrossRef]
- Asmussen, G.; Beckers-Bleukx, G.; Marechal, G. The force-velocity relation of the rabbit inferior oblique muscle; influence of temperature. Pflug. Arch. Eur. J. Physiol. 1994, 426, 542–547. [Google Scholar] [CrossRef]
- Pellegrino, M.A.; Canepari, M.; Rossi, R.; D’Antona, G.; Reggiani, C.; Bottinelli, R. Orthologous myosin isoforms and scaling of shortening velocity with body size in mouse, rat, rabbit and human muscles. J. Physiol. 2003, 546, 677–689. [Google Scholar] [CrossRef]
- Nyitrai, M.; Rossi, R.; Adamek, N.; Pellegrino, M.A.; Bottinelli, R.; Geeves, M.A. What limits the velocity of fast-skeletal muscle contraction in mammals? J. Mol. Biol. 2006, 355, 432–442. [Google Scholar] [CrossRef]
- Van der Heide, U.; Ketelaars, M.; Treijtel, B.W.; de Beer, E.L.; Blange, T. Strain dependence of the elastic properties of force-producing cross- bridges in rigor skeletal muscle. Biophys. J. 1997, 72, 814–821. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; White, H.D.; Belknap, B.; Winkelmann, D.A.; Forgacs, E. Omecamtiv Mecarbil modulates the kinetic and motile properties of porcine beta-cardiac myosin. Biochemistry 2015, 54, 1963–1975. [Google Scholar] [CrossRef]
- Ferenczi, M.A.; Goldman, Y.E.; Simmons, R.M. The dependence of force and shortening velocity on substrate concentration in skinned muscle fibres from Rana temporaria. J. Physiol. 1984, 350, 519–543. [Google Scholar] [CrossRef]
- Cooke, R.; Bialek, W. Contraction of glycerinated muscle fibers as a function of the ATP concentration. Biophys. J. 1979, 28, 241–258. [Google Scholar] [CrossRef] [Green Version]
- Capitanio, M.; Canepari, M.; Cacciafesta, P.; Lombardi, V.; Cicchi, R.; Maffei, M.; Pavone, F.S.; Bottinelli, R. Two independent mechanical events in the interaction cycle of skeletal muscle myosin with actin. Proc. Natl. Acad. Sci. USA 2006, 103, 87–92. [Google Scholar] [CrossRef] [Green Version]
- Piazzesi, G.; Reconditi, M.; Linari, M.; Lucii, L.; Bianco, P.; Brunello, E.; Decostre, V.; Stewart, A.; Gore, D.B.; Irving, T.C.; et al. Skeletal muscle performance determined by modulation of number of Myosin motors rather than motor force or stroke size. Cell 2007, 131, 784–795. [Google Scholar] [CrossRef] [Green Version]
- Edman, K.A. Contractile properties of mouse single muscle fibers, a comparison with amphibian muscle fibers. J. Exp. Biol. 2005, 208, 1905–1913. [Google Scholar] [CrossRef] [Green Version]
- Westerblad, H.; Bruton, J.D.; Lannergren, J. The effect of intracellular pH on contractile function of intact, single fibres of mouse muscle declines with increasing temperature. J. Physiol. 1997, 500, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Duke, T.A. Molecular model of muscle contraction. Proc. Natl. Acad. Sci. USA 1999, 96, 2770–2775. [Google Scholar] [CrossRef]
- Bottinelli, R.; Cappelli, V.; Morner, S.E.; Reggiani, C. Effects of amrinone on shortening velocity and force development in skinned skeletal muscle fibres. J. Muscle Res. Cell Motil. 1993, 14, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Walcott, S.; Warshaw, D.M.; Debold, E.P. Mechanical coupling between myosin molecules causes differences between ensemble and single-molecule measurements. Biophys. J. 2012, 103, 501–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pate, E.; Cooke, R. Simulation of stochastic processes in motile crossbridge systems. J. Muscle Res. Cell Motil. 1991, 12, 376–393. [Google Scholar] [CrossRef] [PubMed]
- Debold, E.P.; Patlak, J.B.; Warshaw, D.M. Slip sliding away: Load-dependence of velocity generated by skeletal muscle myosin molecules in the laser trap. Biophys. J. 2005, 89, L34–L36. [Google Scholar] [CrossRef] [Green Version]
- Debold, E.P.; Walcott, S.; Woodward, M.; Turner, M.A. Direct observation of phosphate inhibiting the force-generating capacity of a miniensemble of Myosin molecules. Biophys. J. 2013, 105, 2374–2384. [Google Scholar] [CrossRef] [Green Version]
- Pertici, I.; Bongini, L.; Melli, L.; Bianchi, G.; Salvi, L.; Falorsi, G.; Squarci, C.; Bozo, T.; Cojoc, D.; Kellermayer, M.S.Z.; et al. A myosin II nanomachine mimicking the striated muscle. Nat. Commun. 2018, 9, 3532. [Google Scholar] [CrossRef] [Green Version]
- Pertici, I.; Bianchi, G.; Bongini, L.; Lombardi, V.; Bianco, P. A Myosin II-Based Nanomachine Devised for the Study of Ca(2+)-Dependent Mechanisms of Muscle Regulation. Int. J. Mol. Sci. 2020, 21, 7372. [Google Scholar] [CrossRef]
- Matusovsky, O.S.; Kodera, N.; MacEachen, C.; Ando, T.; Cheng, Y.S.; Rassier, D.E. Millisecond Conformational Dynamics of Skeletal Myosin II Power Stroke Studied by High-Speed Atomic Force Microscopy. ACS Nano 2021, 15, 2229–2239. [Google Scholar] [CrossRef]
- Hill, A.V. The heat of shortening and the dynamic constants of muscle. Proc. R. Soc. London. Ser. B-Biol. Sci. 1938, 126, 136–195. [Google Scholar]
- Cheng, Y.S.; de Souza Leite, F.; Rassier, D.E. The load dependence and the force-velocity relation in intact myosin filaments from skeletal and smooth muscles. Am. J. Physiol. Cell Physiol. 2020, 318, C103–C110. [Google Scholar] [CrossRef]
- Mansson, A. The effects of inorganic phosphate on muscle force development and energetics: Challenges in modelling related to experimental uncertainties. J. Muscle Res. Cell Motil. 2021, 44, 33–46. [Google Scholar] [CrossRef]
- Piazzesi, G.; Lombardi, V. A cross-bridge model that is able to explain mechanical and energetic properties of shortening muscle. Biophys. J. 1995, 68, 1966–1979. [Google Scholar] [CrossRef]
- Caremani, M.; Melli, L.; Dolfi, M.; Lombardi, V.; Linari, M. The working stroke of the myosin II motor in muscle is not tightly coupled to release of orthophosphate from its active site. J. Physiol. 2013, 591, 5187–5205. [Google Scholar] [CrossRef]
- Lombardi, V.; Piazzesi, G.; Linari, M. Rapid regeneration of the actin-myosin power stroke in contracting muscle. Nature 1992, 355, 638–641. [Google Scholar] [CrossRef]
- Huxley, A.F.; Tideswell, S. Rapid regeneration of power stroke in contracting muscle by attachment of second myosin head. J. Muscle Res. Cell Motil. 1997, 18, 111–114. [Google Scholar] [CrossRef]
- Chen, Y.D.; Brenner, B. On the regeneration of the actin-myosin power stroke in contracting muscle. Proc. Natl. Acad. Sci. USA 1993, 90, 5148–5152. [Google Scholar] [CrossRef] [Green Version]
- Linari, M.; Brunello, E.; Reconditi, M.; Fusi, L.; Caremani, M.; Narayanan, T.; Piazzesi, G.; Lombardi, V.; Irving, M. Force generation by skeletal muscle is controlled by mechanosensing in myosin filaments. Nature 2015, 528, 276–279. [Google Scholar] [CrossRef] [Green Version]
- Marcucci, L.; Reggiani, C. Mechanosensing in Myosin Filament Solves a 60 Years Old Conflict in Skeletal Muscle Modeling between High Power Output and Slow Rise in Tension. Front. Physiol. 2016, 7, 427. [Google Scholar] [CrossRef] [Green Version]
- Spudich, J.A. Hypertrophic and dilated cardiomyopathy: Four decades of basic research on muscle lead to potential therapeutic approaches to these devastating genetic diseases. Biophys. J. 2014, 106, 1236–1249. [Google Scholar] [CrossRef] [Green Version]
- Trivedi, D.V.; Adhikari, A.S.; Sarkar, S.S.; Ruppel, K.M.; Spudich, J.A. Hypertrophic cardiomyopathy and the myosin mesa: Viewing an old disease in a new light. Biophys. Rev. 2018, 10, 27–48. [Google Scholar] [CrossRef] [Green Version]
- Uyeda, T.Q.; Kron, S.J.; Spudich, J.A. Myosin step size. Estimation from slow sliding movement of actin over low densities of heavy meromyosin. J. Mol. Biol. 1990, 214, 699–710. [Google Scholar] [CrossRef]
- Brizendine, R.K.; Alcala, D.B.; Carter, M.S.; Haldeman, B.D.; Facemyer, K.C.; Baker, J.E.; Cremo, C.R. Velocities of unloaded muscle filaments are not limited by drag forces imposed by myosin cross-bridges. Proc. Natl. Acad. Sci. USA 2015, 112, 11235–11240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yengo, C.M.; Takagi, Y.; Sellers, J.R. Temperature dependent measurements reveal similarities between muscle and non-muscle myosin motility. J. Muscle Res. Cell Motil. 2012, 33, 385–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riveline, D.; Ott, A.; Julicher, F.; Winkelmann, D.A.; Cardoso, O.; Lacapere, J.J.; Magnusdottir, S.; Viovy, J.L.; Gorre-Talini, L.; Prost, J. Acting on actin: The electric motility assay. Eur. Biophys. J. 1998, 27, 403–408. [Google Scholar] [CrossRef]
- Moretto, L.; Usaj, M.; Matusovsky, O.; Rassier, D.E.; Friedman, R.; Mansson, A. Multistep orthophosphate release tunes actomyosin energy transduction. Nat. Commun. 2022, 13, 4575. [Google Scholar] [CrossRef]
- Linari, M.; Piazzesi, G.; Pertici, I.; Dantzig, J.A.; Goldman, Y.E.; Lombardi, V. Straightening Out the Elasticity of Myosin Cross-Bridges. Biophys. J. 2020, 118, 994–1002. [Google Scholar]
- Huxley, A.F.; Tideswell, S. Filament compliance and tension transients in muscle. J. Muscle Res. Cell Motil. 1996, 17, 507–511. [Google Scholar] [CrossRef]
- Eisenberg, E.; Hill, T.L.; Chen, Y. Cross-bridge model of muscle contraction. Quantitative analysis. Biophys. J. 1980, 29, 195–227. [Google Scholar] [CrossRef] [Green Version]
- Huxley, A.F.; Simmons, R.M. Proposed mechanism of force generation in striated muscle. Nature 1971, 233, 533–538. [Google Scholar] [CrossRef]
- Gillespie, D.T. A general method for numerically simulating the stochastic time evolution of coupled chemical reactions. J. Comput. Phys. 1976, 22, 403–434. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Numerical Value of Parameter |
|---|---|
| x1(AMDP, AMDPPP) | 7.2 nm |
| x1 1(AMDPPiR, AMDL) | 6.7 nm |
| x2(AMDH) | 1.0 nm |
| x3 | 0 nm |
| ΔGw (MDP − AMDP) | 0 kBT |
| ΔGAMDP-AMDP-PP ≡ ΔGon (AMDP − AMDPPP) | 0.7 kBT |
| ΔG PiR (AMDPPP − AMDPPiR) | 1 kBT |
| ΔGAMDPPiR-AMDL (AMDPPiR − AMDL) | kBT ln([Pi]/KC) |
| ΔGAMDL-AMDH (AMDL − AMDH) | 14 kBT |
| ΔGAMDH-AMD(AMDH − AMD) | 2 kBT |
| ΔGATP | 13.1 + ln ([MgATP]/ ([MgADP][Pi]) kBT |
| ks | 2.8 pN/nm |
| Parameter | Numerical Value of Parameter |
|---|---|
| k+3+ k−3 | 220 s−1 |
| K3 | 10 |
| k−5 | 2000 s−1 |
| Kc | 10 mM |
| kon’ | 130 s−1 |
| kPr+’ | 3000 s−1 b |
| kP+ | 10000 s−1 |
| kLH+ | 6000 s−1 |
| xcrit | 0.6 nm |
| k6 | 5000 s−1 |
| Physiological [Pi] | 0.5 mM |
| [MgATP] | 5 mM |
| K1 | 1.7 mM−1 |
| k2 | 2000 s−1 |
| Parameter | 30 °C | 22 °C | 15 °C | 5 °C |
|---|---|---|---|---|
| ΔGAMDL-AMDH | 12 kBT | 9.2 kBT | 7.2 kBT | 5.1 kBT |
| ΔGAMDH-AMD | 4 kBT | 4 kBT | 4 kBT | 4 kBT |
| kon’ | 325 s−1 | 168 s−1 | 121 s−1 | 62.5 s−1 |
| x1 | 0 nm | 0 nm | 0 nm | 0 nm |
| x11 | 0 nm | 0 nm | 0 nm | 0 nm |
| x2 | −5.5 nm | −5.5 nm | −5.5 nm | −5.5 nm |
| k+3+ k−3(Recovery stroke+hydrolysis) | 220 s−1 | 87.9 s−1 | 39.4 s−1 | 12.5 s−1 |
| K3 | 10 | 7.5 | 5.8 | 4 |
| k2 | 2000 s−1 | 1207 s−1 | 776 s−1 | 413 s−1 |
| kP+’ | 3000 s−1 | 1925 s−1 | 1305 s−1 | 750 s−1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Månsson, A.; Rassier, D.E. Insights into Muscle Contraction Derived from the Effects of Small-Molecular Actomyosin-Modulating Compounds. Int. J. Mol. Sci. 2022, 23, 12084. https://doi.org/10.3390/ijms232012084
Månsson A, Rassier DE. Insights into Muscle Contraction Derived from the Effects of Small-Molecular Actomyosin-Modulating Compounds. International Journal of Molecular Sciences. 2022; 23(20):12084. https://doi.org/10.3390/ijms232012084
Chicago/Turabian StyleMånsson, Alf, and Dilson E. Rassier. 2022. "Insights into Muscle Contraction Derived from the Effects of Small-Molecular Actomyosin-Modulating Compounds" International Journal of Molecular Sciences 23, no. 20: 12084. https://doi.org/10.3390/ijms232012084
APA StyleMånsson, A., & Rassier, D. E. (2022). Insights into Muscle Contraction Derived from the Effects of Small-Molecular Actomyosin-Modulating Compounds. International Journal of Molecular Sciences, 23(20), 12084. https://doi.org/10.3390/ijms232012084