

Application of FTIR Spectroscopy to Detect Changes in Skeletal Muscle Composition Due to Obesity with Insulin Resistance and STZ-Induced Diabetes
 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Animals
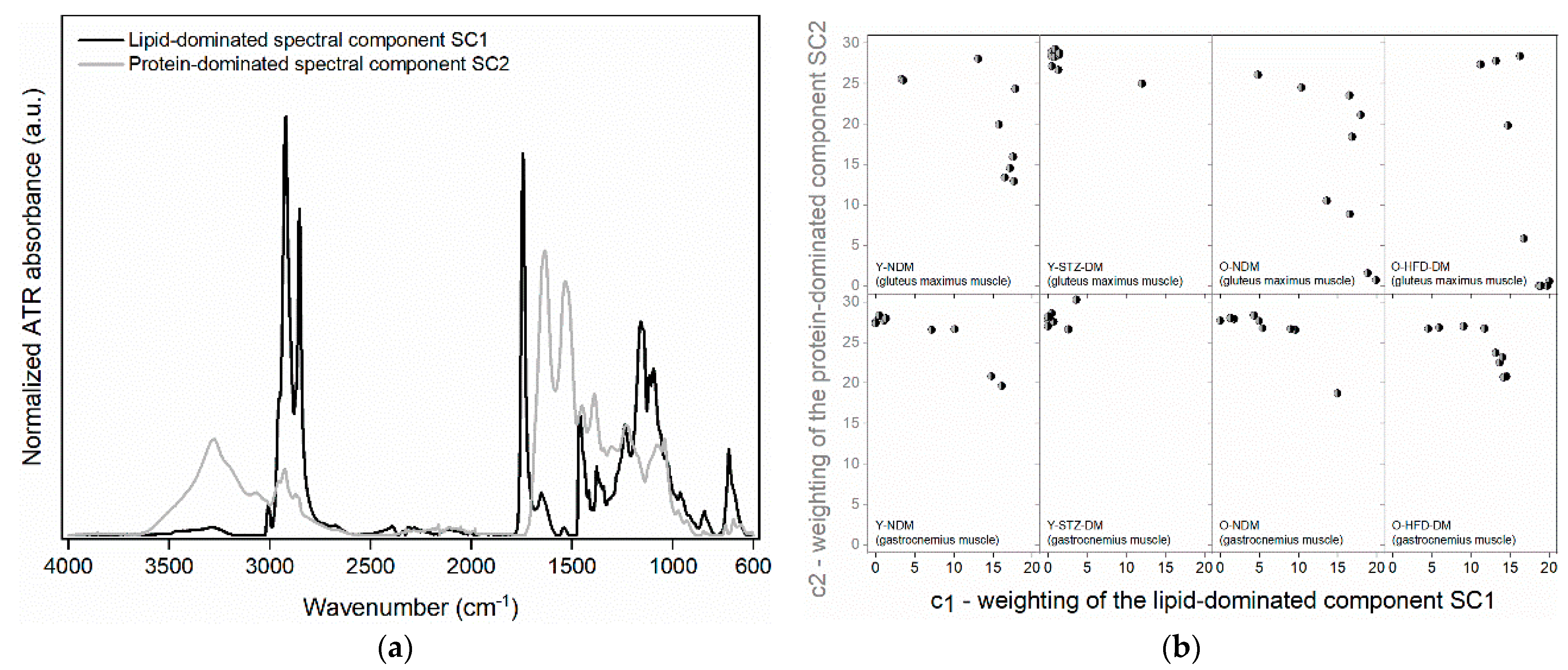
2.2. Muscle Compositional Changes Analysed by FTIR Spectroscopy
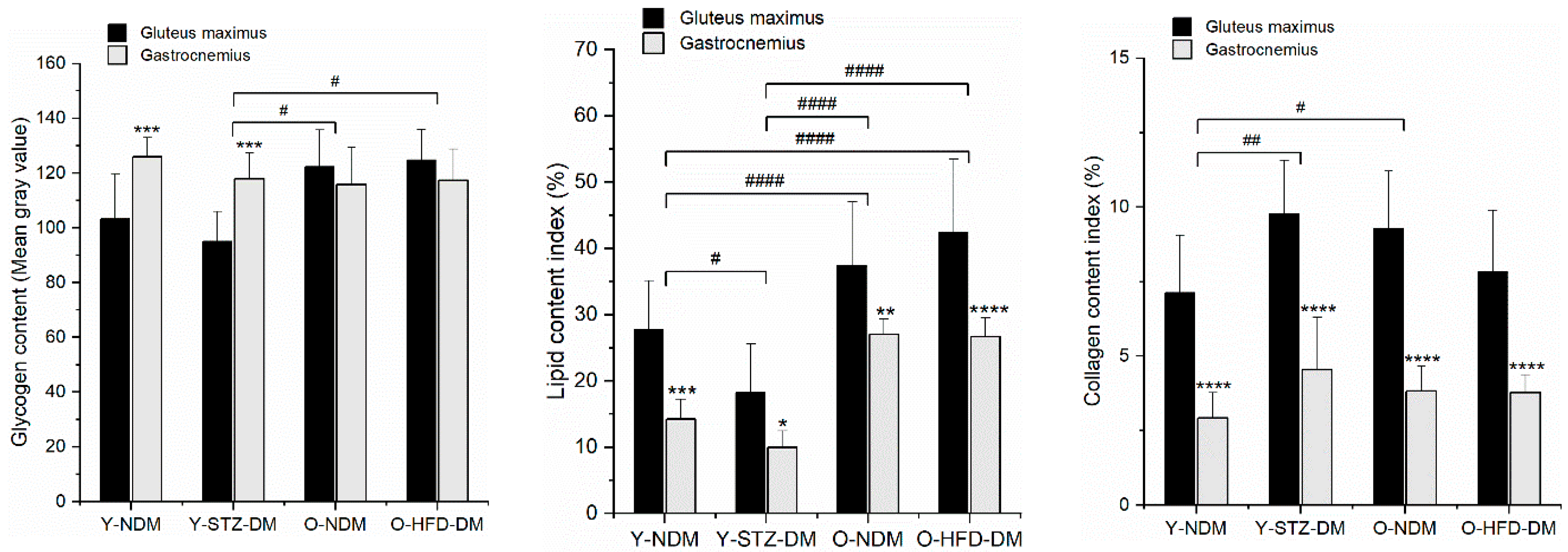
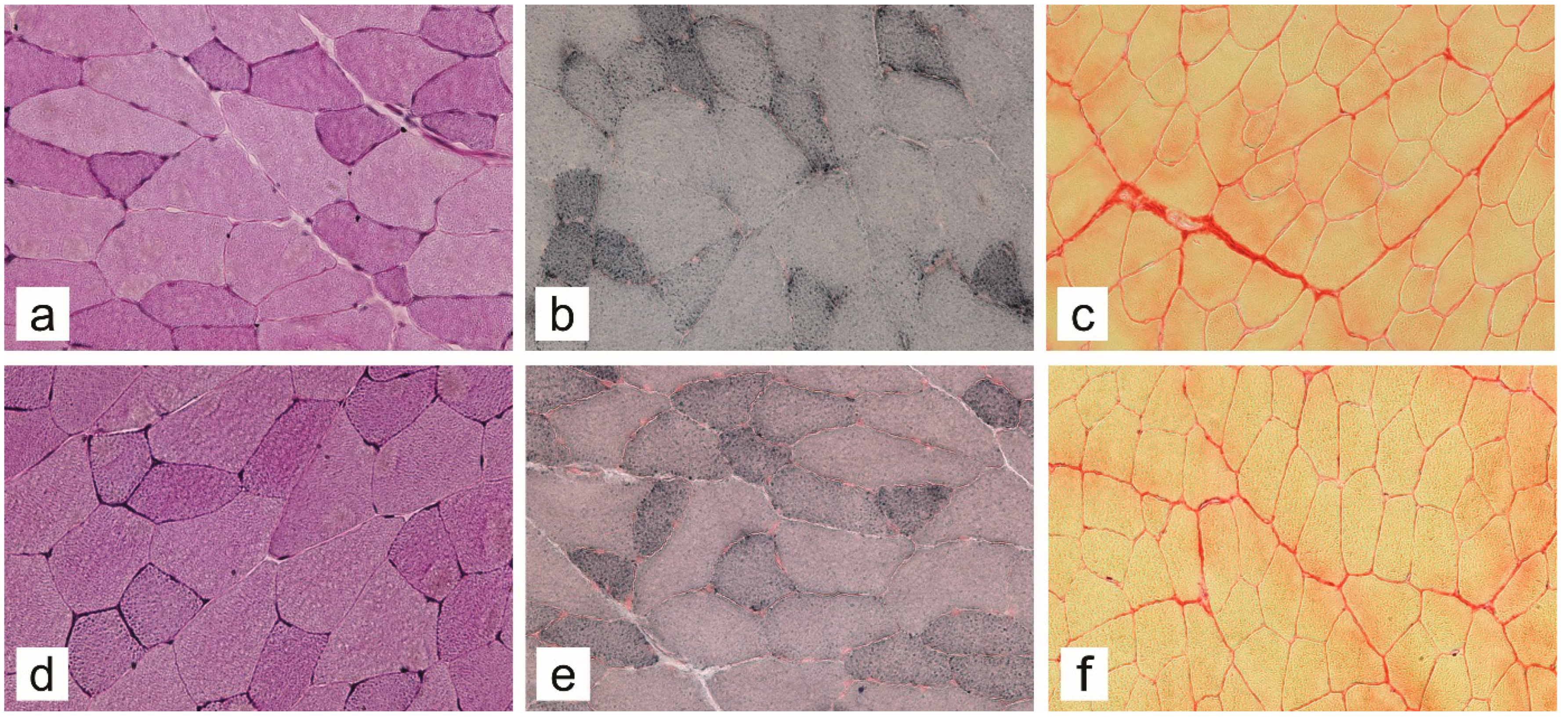
2.3. Muscle Compositional Changes by Histochemical Analysis
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. FTIR Spectroscopic Analysis and Data Processing
4.3. Histochemical Analysis
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Delp, M.D.; Duan, C. Composition and Size of Type I, IIA, IID/X, and IIB Fibers and Citrate Synthase Activity of Rat Muscle. J. Appl. Physiol. 1996, 80, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Meznaric, M.; Cvetko, E. Size and Proportions of Slow-Twitch and Fast-Twitch Muscle Fibers in Human Costal Diaphragm. Biomed Res. Int. 2016, 2016, 5946520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albers, P.H.; Pedersen, A.J.T.; Birk, J.B.; Kristensen, D.E.; Vind, B.F.; Baba, O.; Nøhr, J.; Højlund, K.; Wojtaszewski, J.F.P. Human Muscle Fiber Type-Specific Insulin Signaling: Impact of Obesity and Type 2 Diabetes. Diabetes 2015, 64, 485–497. [Google Scholar] [CrossRef] [Green Version]
- Goodpaster, B.H.; Wolf, D. Skeletal Muscle Lipid Accumulation in Obesity, Insulin Resistance, and Type 2 Diabetes. Pediatr. Diabetes 2004, 5, 219–226. [Google Scholar] [CrossRef]
- Al-Sofiani, M.E.; Ganji, S.S.; Kalyani, R.R. Body Composition Changes in Diabetes and Aging. J. Diabetes Complications 2019, 33, 451–459. [Google Scholar] [CrossRef]
- D’Souza, D.M.; Al-Sajee, D.; Hawke, T.J. Diabetic Myopathy: Impact of Diabetes Mellitus on Skeletal Muscle Progenitor Cells. Front. Physiol. 2013, 4, 379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Watkins, S.; Kelley, D.E. Skeletal Muscle Lipid Content and Oxidative Enzyme Activity in Relation to Muscle Fiber Type in Type 2 Diabetes and Obesity. Diabetes 2001, 50, 817–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umek, N.; Horvat, S.; Cvetko, E. Skeletal Muscle and Fiber Type-Specific Intramyocellular Lipid Accumulation in Obese Mice. Bosn. J. Basic Med. Sci. 2021, 21, 729–737. [Google Scholar] [CrossRef]
- Perseghin, G.; Lattuada, G.; Danna, M.; Sereni, L.P.; Maffi, P.; De Cobelli, F.; Battezzati, A.; Secchi, A.; Del Maschio, A.; Luzi, L. Insulin Resistance, Intramyocellular Lipid Content, and Plasma Adiponectin in Patients with Type 1 Diabetes. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E1174–E1181. [Google Scholar] [CrossRef] [Green Version]
- Bernroider, E.; Brehm, A.; Krssak, M.; Anderwald, C.; Trajanoski, Z.; Cline, G.; Shulman, G.I.; Roden, M. The Role of Intramyocellular Lipids during Hypoglycemia in Patients with Intensively Treated Type 1 Diabetes. J. Clin. Endocrinol. Metab. 2005, 90, 5559–5565. [Google Scholar] [CrossRef]
- Bergman, B.C.; Goodpaster, B.H. Exercise and Muscle Lipid Content, Composition, and Localization: Influence on Muscle Insulin Sensitivity. Diabetes 2020, 69, 848–858. [Google Scholar] [CrossRef] [PubMed]
- Masgrau, A.; Mishellany-Dutour, A.; Murakami, H.; Beaufrère, A.M.; Walrand, S.; Giraudet, C.; Migné, C.; Gerbaix, M.; Metz, L.; Courteix, D.; et al. Time-Course Changes of Muscle Protein Synthesis Associated with Obesity-Induced Lipotoxicity. J. Physiol. 2012, 590, 5199–5210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorcely, B.; Katz, K.; Jagannathan, R.; Chiang, S.S.; Oluwadare, B.; Goldberg, I.J.; Bergman, M. Novel Biomarkers for Prediabetes, Diabetes, and Associated Complications. Diabetes, Metab. Syndr. Obes. Targets Ther. 2017, 10, 345–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.T.; Chang, W.H.; Kuo, T.F.; Shen, M.Y.; Yang, C.W.; Tien, Y.J.; Lai, B.Y.; Chen, Y.R.; Chang, Y.C.; Yang, W.C. Identification of Novel Biomarkers for Pre-Diabetic Diagnosis Using a Combinational Approach. Front. Endocrinol. 2021, 12, 641336. [Google Scholar] [CrossRef]
- Bozkurt, O.; Severcan, M.; Severcan, F. Diabetes Induces Compositional, Structural and Functional Alterations on Rat Skeletal Soleus Muscle Revealed by FTIR Spectroscopy: A Comparative Study with EDL Muscle. Analyst 2010, 135, 3110–3119. [Google Scholar] [CrossRef]
- Petibois, C.; Gouspillou, G.; Wehbe, K.; Delage, J.P.; Déléris, G. Analysis of Type I and IV Collagens by FT-IR Spectroscopy and Imaging for a Molecular Investigation of Skeletal Muscle Connective Tissue. Anal. Bioanal. Chem. 2006, 386, 1961–1966. [Google Scholar] [CrossRef]
- Beć, K.B.; Grabska, J.; Huck, C.W. Biomolecular and Bioanalytical Applications of Infrared Spectroscopy—A Review. Anal. Chim. Acta 2020, 1133, 150–177. [Google Scholar] [CrossRef]
- Dogan, A.; Lasch, P.; Neuschl, C.; Millrose, M.K.; Alberts, R.; Schughart, K.; Naumann, D.; Brockmann, G.A. ATR-FTIR Spectroscopy Reveals Genomic Loci Regulating the Tissue Response in High Fat Diet Fed BXD Recombinant Inbred Mouse Strains. BMC Genom. 2013, 14, 386. [Google Scholar] [CrossRef] [Green Version]
- Toyran, N.; Lasch, P.; Naumann, D.; Turan, B.; Severcan, F. Early Alterations in Myocardia and Vessels of the Diabetic Rat Heart: An FTIR Microspectroscopic Study. Biochem. J. 2006, 397, 427–436. [Google Scholar] [CrossRef]
- Sen, I.; Bozkurt, O.; Aras, E.; Heise, S.; Brockmann, G.A.; Severcan, F. Lipid Profiles of Adipose and Muscle Tissues in Mouse Models of Juvenile Onset of Obesity without High Fat Diet Induction: A Fourier Transform Infrared (FT-IR) Spectroscopic Study. Appl. Spectrosc. 2015, 69, 679–688. [Google Scholar] [CrossRef]
- Wiercigroch, E.; Szafraniec, E.; Czamara, K.; Pacia, M.Z.; Majzner, K.; Kochan, K.; Kaczor, A.; Baranska, M.; Malek, K. Raman and Infrared Spectroscopy of Carbohydrates: A Review. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2017, 185, 317–335. [Google Scholar] [CrossRef]
- Jerônimo, D.P.; De Souza, R.A.; Da Silva, F.F.; Camargo, G.L.; Miranda, H.L.; Xavier, M.; Sakane, K.K.; Ribeiro, W. Detection of Creatine in Rat Muscle by FTIR Spectroscopy. Ann. Biomed. Eng. 2012, 40, 2069–2077. [Google Scholar] [CrossRef]
- Diem, M. Modern Vibrational Spectroscopy and Micro-Spectroscopy: Theory, Instrumentation and Biomedical Applications; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2015; ISBN 9781118824924. [Google Scholar]
- Siebert, F.; Hildebrandt, P. Vibrational Spectroscopy in Life Science; John Wiley & Sons: Hoboken, NJ, USA, 2008; ISBN 9783527405060. [Google Scholar]
- Anastassopoulou, J.; Kyriakidou, M.; Mamareli, V.; Tanis, O.; Rallis, M. The Influence of UV Irradiation on Diabetic Mice Skin. A Vibrational FT-IR and Raman Spectroscopic Study. Chromatogr. Spectrosc. Tech. 2019, 2, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Movasaghi, Z.; Rehman, S.; Rehman, I.U. Fourier Transform Infrared (FTIR) Spectroscopy of Biological Tissues. Appl. Spectrosc. Rev. 2008, 43, 134–179. [Google Scholar] [CrossRef]
- Rohman, A.; Che Man, Y.B. The Optimization of FTIR Spectroscopy Combined with Partial Least Square for Analysis of Animal Fats in Quartenary Mixtures. Spectroscopy 2011, 25, 169–176. [Google Scholar] [CrossRef]
- Wang, Q.; He, H.; Li, B.; Lin, H.; Zhang, Y.; Zhang, J.; Wang, Z. UV–Vis and ATR–FTIR Spectroscopic Investigations of Postmortem Interval Based on the Changes in Rabbit Plasma. PLoS ONE 2017, 12, e0182161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gremlich, H.-U.; Yan, B. Infrared and Raman Spectroscopy of Biological Materials; CRC Press: Boca Raton, FL, USA, 2000; Volume 148, ISBN 0824704096. [Google Scholar]
- Ferreira, I.C.C.; Aguiar, E.M.G.; Silva, A.T.F.; Santos, L.L.D.; Cardoso-Sousa, L.; Araújo, T.G.; Santos, D.W.; Goulart, L.R.; Sabino-Silva, R.; Maia, Y.C.P.; et al. Attenuated Total Reflection-Fourier Transform Infrared (ATR-FTIR) Spectroscopy Analysis of Saliva for Breast Cancer Diagnosis. J. Oncol. 2020, 2020, 4343590. [Google Scholar] [CrossRef] [Green Version]
- Xiao, H.; Cai, G.; Liu, M. Hydroxyl Radical Induced Structural Changes of Collagen. Spectroscopy 2007, 21, 91–103. [Google Scholar] [CrossRef] [Green Version]
- Caine, S.; Heraud, P.; Tobin, M.J.; McNaughton, D.; Bernard, C.C.A. The Application of Fourier Transform Infrared Microspectroscopy for the Study of Diseased Central Nervous System Tissue. Neuroimage 2012, 59, 3624–3640. [Google Scholar] [CrossRef]
- Ami, D.; Neri, T.; Natalello, A.; Mereghetti, P.; Doglia, S.M.; Zanoni, M.; Zuccotti, M.; Garagna, S.; Redi, C.A. Embryonic Stem Cell Differentiation Studied by FT-IR Spectroscopy. Biochim. Biophys. Acta Mol. Cell Res. 2008, 1783, 98–106. [Google Scholar] [CrossRef]
- Alhazmi, H.A. FT-IR Spectroscopy for the Identification of Binding Sites and Measurements of the Binding Interactions of Important Metal Ions with Bovine Serum Albumin. Sci. Pharm. 2019, 87, 5. [Google Scholar] [CrossRef] [Green Version]
- Goormaghtigh, E.; Ruysschaert, J.M.; Raussens, V. Evaluation of the Information Content in Infrared Spectra for Protein Secondary Structure Determination. Biophys. J. 2006, 90, 2946–2957. [Google Scholar] [CrossRef] [Green Version]
- Baker, M.J.; Gazi, E.; Brown, M.D.; Shanks, J.H.; Gardner, P.; Clarke, N.W. FTIR-Based Spectroscopic Analysis in the Identification of Clinically Aggressive Prostate Cancer. Br. J. Cancer 2008, 99, 1859–1866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray-Wijelath, J.; Lyman, D.J.; Wijelath, E.S. Vascular Graft Healing. III. FTIR Analysis of EPTFE Graft Samples from Implanted Bigrafts. J. Biomed. Mater. Res. 2004, 70, 223–232. [Google Scholar] [CrossRef]
- Röhr, D.; Boon, B.D.C.; Schuler, M.; Kremer, K.; Hoozemans, J.J.M.; Bouwman, F.H.; El-Mashtoly, S.F.; Nabers, A.; Großerueschkamp, F.; Rozemuller, A.J.M.; et al. Label-Free Vibrational Imaging of Different Aβ Plaque Types in Alzheimer’s Disease Reveals Sequential Events in Plaque Development. Acta Neuropathol. Commun. 2020, 8, 222. [Google Scholar] [CrossRef]
- Qin, L.; Bi, J.R.; Li, D.M.; Dong, M.; Zhao, Z.Y.; Dong, X.P.; Zhou, D.Y.; Zhu, B.W. Unfolding/Refolding Study on Collagen from Sea Cucumber Based on 2D Fourier Transform Infrared Spectroscopy. Molecules 2016, 21, 1546. [Google Scholar] [CrossRef] [Green Version]
- Scott, D.A.; Renaud, D.E.; Krishnasamy, S.; Meriç, P.; Buduneli, N.; Çetinkalp, Ş.; Liu, K.Z. Diabetes-Related Molecular Signatures in Infrared Spectra of Human Saliva. Diabetol. Metab. Syndr. 2010, 2, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamosaityte, S.; Galli, R.; Uckermann, O.; Sitoci-Ficici, K.H.; Later, R.; Beiermeister, R.; Doberenz, F.; Gelinsky, M.; Leipnitz, E.; Schackert, G.; et al. Biochemical Monitoring of Spinal Cord Injury by FT-IR Spectroscopy-Effects of Therapeutic Alginate Implant in Rat Models. PLoS ONE 2015, 10, e0142660. [Google Scholar] [CrossRef] [PubMed]
- Balan, V.; Mihai, C.T.; Cojocaru, F.D.; Uritu, C.M.; Dodi, G.; Botezat, D.; Gardikiotis, I. Vibrational Spectroscopy Fingerprinting in Medicine: From Molecular to Clinical Practice. Materials 2019, 12, 2884. [Google Scholar] [CrossRef] [Green Version]
- Derenne, A.; Claessens, T.; Conus, C.; Goormaghtigh, E. Infrared Spectroscopy of Membrane Lipids. In Encyclopedia of Biophysics; Roberts, G.C.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1074–1081. ISBN 9783642167126. [Google Scholar] [CrossRef]
- Volpe, M.G.; Costantini, S.; Coccia, E.; Parrillo, L.; Paolucci, M. Evaluation of Metabolic Changes Induced by Polyphenols in the Crayfish Astacus Leptodactylus by Metabolomics Using Fourier Transformed Infrared Spectroscopy. J. Biosci. 2018, 43, 585–596. [Google Scholar] [CrossRef]
- Liang, P.; Wang, H.; Chen, C.; Ge, F.; Liu, D.; Li, S.; Han, B.; Xiong, X.; Zhao, S. The Use of Fourier Transform Infrared Spectroscopy for Quantification of Adulteration in Virgin Walnut Oil. J. Spectrosc. 2013, 2013, 305604. [Google Scholar] [CrossRef] [Green Version]
- Heys, K. Vibrational Spectroscopy as a Tool to Investigate the Effects of Environmental Contaminants in Predatory Birds. Ph.D. Thesis, Lancaster University, Lancaster, UK, 2017. [Google Scholar]
- Ami, D.; Mereghetti, P.; Leri, M.; Giorgetti, S.; Natalello, A.; Doglia, S.M.; Stefani, M.; Bucciantini, M. A FTIR Microspectroscopy Study of the Structural and Biochemical Perturbations Induced by Natively Folded and Aggregated Transthyretin in HL-1 Cardiomyocytes. Sci. Rep. 2018, 8, 12508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belbachir, K.; Noreen, R.; Gouspillou, G.; Petibois, C. Collagen Types Analysis and Differentiation by FTIR Spectroscopy. Anal. Bioanal. Chem. 2009, 395, 829–837. [Google Scholar] [CrossRef]
- Najbjerg, H.; Afseth, N.K.; Young, J.F.; Bertram, H.C.; Pedersen, M.E.; Grimmer, S.; Vogt, G.; Kohler, A. Monitoring Cellular Responses upon Fatty Acid Exposure by Fourier Transform Infrared Spectroscopy and Raman Spectroscopy. Analyst 2011, 136, 1649–1658. [Google Scholar] [CrossRef] [PubMed]
- Böcker, U.; Ofstad, R.; Wu, Z.; Bertram, H.C.; Sockalingum, G.D.; Manfait, M.; Egelandsdal, B.; Kohler, A. Revealing Covariance Structures in Fourier Transform Infrared and Raman Microspectroscopy Spectra: A Study on Pork Muscle Fiber Tissue Subjected to Different Processing Parameters. Appl. Spectrosc. 2007, 61, 1032–1039. [Google Scholar] [CrossRef]
- Lopes, J.S. FTIR, a Potential Tool to Dementia Diagnosis through Analysis of Plasma; Aveiro University: Aveiro, Portugal, 2015. [Google Scholar]
- Simonova, D.; Karamancheva, I. Application of Fourier Transform Infrared Spectroscopy for Tumor Diagnosis. Biotechnol. Biotechnol. Equip. 2013, 27, 4200–4207. [Google Scholar] [CrossRef]
- Sanden, K.W.; Kohler, A.; Afseth, N.K.; Böcker, U.; Rønning, S.B.; Liland, K.H.; Pedersen, M.E. The Use of Fourier-Transform Infrared Spectroscopy to Characterize Connective Tissue Components in Skeletal Muscle of Atlantic Cod (Gadus morhua L.). J. Biophotonics 2019, 12, e201800436. [Google Scholar] [CrossRef] [Green Version]
- Stani, C.; Vaccari, L.; Mitri, E.; Birarda, G. FTIR Investigation of the Secondary Structure of Type I Collagen: New Insight into the Amide III Band. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2020, 229, 118006. [Google Scholar] [CrossRef]
- Rehman, I.U.; Movasaghi, Z.; Rehman, S. Vibrational Spectroscopy for Tissue Analysis; CRC Press: Boca Raton, FL, USA, 2019; ISBN 978-1-4398-3609-5. [Google Scholar]
- Heidari, A. An Analytical and Computational Infrared Spectroscopic Review of Vibrational Modes in Nucleic Acids. Austin J. Anal. Pharm. Chem. 2016, 3, 1058. [Google Scholar]
- Wong, P.T.; Goldstein, S.M.; Grekin, R.C.; Godwin, T.A.; Pivik, C.; Rigas, B. Distinct Infrared Spectroscopic Patterns of Human Basal Cell Carcinoma of the Skin. Cancer Res. 1993, 53, 762–765. [Google Scholar]
- Bellisola, G.; Della Peruta, M.; Vezzalini, M.; Moratti, E.; Vaccari, L.; Birarda, G.; Piccinini, M.; Cinque, G.; Sorio, C. Tracking Infrared Signatures of Drugs in Cancer Cells by Fourier Transform Microspectroscopy. Analyst 2010, 135, 3077–3086. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Yoshida, H. Nondestructive Analyses of Unsaturated Fatty Acid Species in Dietary Oils by Attenuated Total Reflectance with Fourier Transform IR Spectroscopy. Biopolymers 2003, 70, 604–613. [Google Scholar] [CrossRef] [PubMed]
- Andrus, P.G.L.; Strickland, R.D. Cancer Grading by Fourier Transform Infrared Spectroscopy. Biospectroscopy 1998, 4, 37–46. [Google Scholar] [CrossRef]
- Chiriboga, L.; Xie, P.; Yee, H.; Vigorita, V.; Zarou, D.; Zakim, D.; Diem, M. Infrared Spectroscopy of Human Tissue. I. Differentiation and Maturation of Epithelial Cells in the Human Cervix. Biospectroscopy 1998, 4, 47–53. [Google Scholar] [CrossRef]
- Koch, C.; Brandstetter, M.; Wechselberger, P.; Lorantfy, B.; Plata, M.R.; Radel, S.; Herwig, C.; Lendl, B. Ultrasound-Enhanced Attenuated Total Reflection Mid-Infrared Spectroscopy in-Line Probe: Acquisition of Cell Spectra in a Bioreactor. Anal. Chem. 2015, 87, 2314–2320. [Google Scholar] [CrossRef] [PubMed]
- Bridelli, M.G.; Stani, C.; Bedotti, R. Fourier Transform Infrared Conformational Investigation of Type I Collagen Aged by in Vitro Induced Dehydration and Non-Enzymatic Glycation Treatments. J. Biol. Res. Boll. Soc. Ital. Biol. Sper. 2017, 90, 45–50. [Google Scholar] [CrossRef]
- Hampton, C.; Demoin, D.; Glaser, R.E. Vibrational Spectroscopy Tutorial: Sulfur and Phosphorus. Available online: https://pdf4pro.com/cdn/vibrational-spectroscopy-tutorial-sulfur-2df5e9.pdf (accessed on 10 September 2021).
- Banyay, M.; Sarkar, M.; Gräslund, A. A Library of IR Bands of Nucleic Acids in Solution. Biophys. Chem. 2003, 104, 477–488. [Google Scholar] [CrossRef]
- Taillandier, E.; Peticolas, W.L.; Adam, S.; Huynh-Dinh, T.; Igolen, J. Polymorphism of the d(CCCGCGGG)2 Double Helix Studied by FT-Ir Spectroscopy. Spectrochim. Acta Part A Mol. Spectrosc. 1990, 46, 107–112. [Google Scholar] [CrossRef]
- Siamwiza, M.N.; Lord, R.C.; Chen, M.C.; Takamatsu, T.; Harada, I.; Matsuura, H.; Shimanouchi, T. IInterpretation of the Doublet at 850 and 830 Cm-1 in the Raman Spectra of Tyrosyl Residues in Proteins and Certain Model Compounds. Biochemistry 1975, 14, 4870–4876. [Google Scholar] [CrossRef]
- Yadav, R.A.; Dixit, V.; Yogesh, M. Raman and IR Spectral and DFT Based Vibrational and Electronic Characterization of Isolated and Zwitterionic Forms of L-Tyrosin. Pharm. Anal. Acta 2015, 6, 439. [Google Scholar] [CrossRef] [Green Version]
- Yonar, D.; Ocek, L.; Tiftikcioglu, B.I.; Zorlu, Y.; Severcan, F. Relapsing-Remitting Multiple Sclerosis Diagnosis from Cerebrospinal Fluids via Fourier Transform Infrared Spectroscopy Coupled with Multivariate Analysis. Sci. Rep. 2018, 8, 1025. [Google Scholar] [CrossRef] [PubMed]
- Cayrol, B.; Geinguenaud, F.; Lacoste, J.; Busi, F.; Le Dérout, J.; Piétrement, O.; Le Cam, E.; Régnier, P.; Lavelle, C.; Arluison, V. Auto-Assembly of E. Coli DsrA Small Noncoding RNA: Molecular Characteristics and Functional Consequences. RNA Biol. 2009, 6, 434–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geinguenaud, F.; Militello, V.; Arluison, V. Application of FTIR Spectroscopy to Analyze RNA Structure. In RNA Spectroscopy; Humana: New York, NY, 2020; pp. 119–133. ISBN 9781071602782. [Google Scholar]
- Coates, J. Encyclopedia of Analytical Chemistry -Interpretation of Infrared Spectra, A Practical Approach; Wiley: Chichester, UK, 2004. [Google Scholar]
- Aminullah; Mardiah; Sutsuga, H.; Kemala, T. Study of Different Extraction Methods on Finger Print and Fatty Acid of Raw Beef Fat Using Fourier Transform Infrared and Gas Chromatography-Mass Spectrometry. Open Chem. 2018, 16, 1099–1105. [Google Scholar] [CrossRef]
- Coleman, S.K. Skeletal Muscle as a Therapeutic Target for Delaying Type 1 Diabetic Complications. World J. Diabetes 2015, 6, 1323. [Google Scholar] [CrossRef] [PubMed]
- Krause, M.P.; Riddell, M.C.; Hawke, T.J. Effects of Type 1 Diabetes Mellitus on Skeletal Muscle: Clinical Observations and Physiological Mechanisms. Pediatr. Diabetes 2011, 12, 345–364. [Google Scholar] [CrossRef]
- Chang, W.; Hatch, G.M.; Wang, Y.; Yu, F.; Wang, M. The Relationship between Phospholipids and Insulin Resistance: From Clinical to Experimental Studies. J. Cell. Mol. Med. 2019, 23, 702–710. [Google Scholar] [CrossRef] [Green Version]
- Chiu, C.Y.; Yang, R.S.; Sheu, M.L.; Chan, D.C.; Yang, T.H.; Tsai, K.S.; Chiang, C.K.; Liu, S.H. Advanced Glycation End-Products Induce Skeletal Muscle Atrophy and Dysfunction in Diabetic Mice via a RAGE-Mediated, AMPK-down-Regulated, Akt Pathway. J. Pathol. 2016, 238, 470–482. [Google Scholar] [CrossRef]
- Pederson, T.M.; Kramer, D.L.; Rondinone, C.M. Serine/Threonine Phosphorylation of IRS-1 Triggers Its Degradation: Possible Regulation by Tyrosine Phosphorylation. Diabetes 2001, 50, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Bouzakri, K.; Roques, M.; Gual, P.; Espinosa, S.; Guebre-Egziabher, F.; Riou, J.P.; Laville, M.; Le Marchand-Brustel, Y.; Tanti, J.F.; Vidal, H. Reduced Activation of Phosphatidylinositol-3 Kinase and Increased Serine 636 Phosphorylation of Insulin Receptor Substrate-1 in Primary Culture of Skeletal Muscle Cells from Patients with Type 2 Diabetes. Diabetes 2003, 52, 1319–1325. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, M.A.; Forbes, J.M. Glucose and Glycogen in the Diabetic Kidney: Heroes or Villains? EBioMedicine 2019, 47, 590–597. [Google Scholar] [CrossRef] [Green Version]
- Damsbo, P.; Vaag, A.; Hother-Nielsen, O.; Beck-Nielsen, H. Reduced Glycogen Synthase Activity in Skeletal Muscle from Obese Patients with and without Type 2 (Non-Insulin-Dependent) Diabetes Mellitus. Diabetologia 1991, 34, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Berria, R.; Wang, L.; Richardson, D.K.; Finlayson, J.; Belfort, R.; Pratipanawatr, T.; De Filippis, E.A.; Kashyap, S.; Mandarino, L.J. Increased Collagen Content in Insulin-Resistant Skeletal Muscle. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E560–E565. [Google Scholar] [CrossRef] [PubMed]
- Workeneh, B.; Bajaj, M. The Regulation of Muscle Protein Turnover in Diabetes. Int. J. Biochem. Cell Biol. 2013, 45, 2239–2244. [Google Scholar] [CrossRef]
- Binder-Markey, B.I.; Broda, N.M.; Lieber, R.L. Intramuscular Anatomy Drives Collagen Content Variation within and between Muscles. Front. Physiol. 2020, 11, 293. [Google Scholar] [CrossRef] [Green Version]
- Bottinelli, R.; Reggiani, C. Human Skeletal Muscle Fibres: Molecular and Functional Diversity. Prog. Biophys. Mol. Biol. 2000, 73, 195–262. [Google Scholar] [CrossRef]
- Umek, N.; Horvat, S.; Cvetko, E.; Kreft, M.; Janáček, J.; Kubínová, L.; Stopar Pintarič, T.; Eržen, I. 3D Analysis of Capillary Network in Skeletal Muscle of Obese Insulin-Resistant Mice. Histochem. Cell Biol. 2019, 152, 323–331. [Google Scholar] [CrossRef]
- King, A.J.F. The Use of Animal Models in Diabetes Research. Br. J. Pharmacol. 2012, 166, 877–894. [Google Scholar] [CrossRef] [Green Version]
- Wu, K.K.; Huan, Y. Streptozotocin-induced Diabetic Models in Mice and Rats. Curr. Protoc. Pharmacol. 2008, 40, 5.47.1–5.47.14. [Google Scholar] [CrossRef]
- Pan, H.; Ding, Y.; Yan, N.; Nie, Y.; Li, M.; Tong, L. Trehalose Prevents Sciatic Nerve Damage to and Apoptosis of Schwann Cells of Streptozotocin-Induced Diabetic C57BL/6J Mice. Biomed. Pharmacother. 2018, 105, 907–914. [Google Scholar] [CrossRef] [PubMed]
- MCR-ALS GUI V4c. Available online: https://www.umu.se/en/research/infrastructure/visp/downloads/ (accessed on 18 November 2020).
- Gorzsás, A. Vibrational Spectroscopy Core Facility. Available online: https://www.umu.se/en/research/infrastructure/visp/downloads/ (accessed on 21 October 2020).
- Goodpaster, B.H.; Theriault, R.; Watkins, S.C.; Kelley, D.E. Intramuscular Lipid Content Is Increased in Obesity and Decreased by Weight Loss. Metabolism 2000, 49, 467–472. [Google Scholar] [CrossRef]
- Fairchild, T.J.; Fournier, P.A. Glycogen Determination Using Periodic Acid-Schiff: Artifact of Muscle Preparation. Med. Sci. Sports Exerc. 2004, 36, 2053–2058. [Google Scholar] [CrossRef] [PubMed]
- McManus, J.F.A. Histological and Histochemical Uses of Periodic Acid. Biotech. Histochem. 1948, 23, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Vesentini, G.; Marini, G.; Piculo, F.; Damasceno, D.C.; Matheus, S.M.M.; Felisbino, S.L.; Calderon, I.M.P.; Hijaz, A.; Barbosa, A.M.P.; Rudge, M.V.C. Morphological Changes in Rat Rectus Abdominis Muscle Induced by Diabetes and Pregnancy. Brazilian J. Med. Biol. Res. 2018, 51, e7035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Junqueira, L.C.U.; Bignolas, G.; Brentani, R.R. Picrosirius Staining plus Polarization Microscopy, a Specific Method for Collagen Detection in Tissue Sections. Histochem. J. 1979, 11, 447–455. [Google Scholar] [CrossRef]
- Rasband, W.S. ImageJ. Available online: https://imagej.nih.gov/ij/ (accessed on 9 September 2021).
- Ruifrok, A.C.; Johnston, D.A. Quantification of Histochemical Staining by Color Deconvolution. Anal. Quant. Cytol. Histol. 2001, 23, 291–299. [Google Scholar] [PubMed]
- Worsfold, P.; Townshend, A.; Poole, C.F.; Miró, M. Encyclopedia of Analytical Science; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Lawton, W.H.; Sylvestre, E.A. Self Modeling Curve Resolution. Technometrics 1971, 13, 617–633. [Google Scholar] [CrossRef]
- Ben-Amotz, D. Hydration-Shell Vibrational Spectroscopy. J. Am. Chem. Soc. 2019, 141, 10569–10580. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Y-NMD | Y-STZ-DM | O-NDM | O-HFD-DM | |
|---|---|---|---|---|
| Body mass (g) | 26.2 ± 1.3 | 19.9 ± 1.7 * | 26.7 ± 2.5 | 36.9 ± 2.2 * |
| Fasting glucose (mmol L−1) | 6.7 ± 1.0 | 31.3 ± 2.8 # | 6.9 ± 0.9 | 8.7 ± 0.7 Ψ |
| SC1—Peak Wavenumber (cm−1) | SC1—Spectral Assignment | SC2—Peak Wavenumber (cm−1) | SC2—Spectral Assignment |
|---|---|---|---|
| 3467 3290 | O-H stretching of carbohydrates [21] | 3307 3294 3272 3067 | amide A and amide B band originating from a Fermi resonance between the N-H stretching vibration and the first overtone of amide II ([22,23,24,25,26] and references therein) |
| 3007 | Olefinic C=CH stretching vibration of unsaturated fatty acids ([18,26] and references therein, [27]) | ||
| (2959 shoulder) 2922 (2874 shoulder) 2853 | CH3 and CH2 symmetric and antisymmetric stretching mostly of fatty acids and phospholipids [22,27,28,29,30] | 2960 2924 2874 (2853 shoulder) | CH3 and CH2 symmetric and antisymmetric stretching mostly of protein side chains [22,31] |
| 1743 | C=O stretching of lipid esters [20,27,32] | ||
| 1693 | amide I (C=O stretching, C-N stretching, CNN deformation) of β-sheet protein secondary structures [33,34] | ||
| 1679 | amide I (C=O stretching, C-N stretching, CNN deformation) of β-sheet protein secondary structures [34,35] | ||
| 1653 | amide I (C=O stretching, C-N stretching, CNN deformation) of α-helical protein secondary structures [32,34,35,36,37] | 1652 | amide I (C=O stretching, C-N stretching, CNN deformation) of α-helical protein secondary structures [32,34,35,36,37] |
| 1629 | amide I (C=O stretching, C-N stretching, CNN deformation) of β-sheet protein secondary structures [28,32,34,38,39] | 1629 | amide I (C=O stretching, C-N stretching, CNN deformation) of β-sheet protein secondary structures [28,32,34,38,39] |
| 1568 | amide II (C-N stretching coupled with N-H bending) of β-turn protein secondary structures [35] | ||
| 1542 | amide II (C-N stretching coupled with N-H bending) of α-helical and β-sheet protein secondary structures [28,36,37] | 1547 | amide II (C-N stretching coupled with N-H bending) of α-helical and β-sheet protein secondary structures [28,36,37] |
| 1530 | amide II (C-N stretching coupled with N-H bending) of β-turn protein secondary structures [35] | ||
| 1513 | vibration of the tyrosine ring in proteins [28,29,40] | ||
| 1464 1457 1436 1417 | CH3 and CH2 bending vibrations mostly of fatty acids and phospholipids [27,28,41,42,43,44] and cis =C–H bending at 1417 cm−1 of unsaturated fatty acids [45] | 1468 1454 1420 1387 | CH3 and CH2 bending vibrations mostly of protein side chains [28,42,44,46,47,48] |
| 1401 | C=O symmetric stretching of COO− groups of fatty acids [49,50,51] | ||
| 1377 1361 | CH3 symmetric bending mostly of fatty acids and phospholipids [27,43,52] | ||
| 1343 1315 | CH3 wagging mostly of fatty acids and phospholipids [43,51] | 1340 | CH2 side chain vibration in collagen ([16,48,53] and references therein) |
| 1309 | amide III of α-helical protein secondary structures [54] | ||
| 1301 | in-phase CH2 twist mode of fatty acids [55] | ||
| (1279 1263 shoulders) 1239 | PO2− antisymmetric stretching of phospholipids [28,43] | 1241 | amide III vibration from C-N stretching, N-H bending vibration, and wagging vibration of CH2 groups in the glycine backbone and proline side chains of collagen ([16,20,32,53] and references therein) and PO2− antisymmetric stretching of RNA ([56] and references therein) |
| 1231 | PO2− antisymmetric stretching of DNA ([56] and references therein) | ||
| 1172 | vibrations of COH groups of serine, threonine, and tyrosine residues in proteins (1161 cm−1—hydrogen-bonded CO group, and 1173 cm−1—non-hydrogen-bonded CO groups) [57,58] | ||
| 1161 1142 | C-O-C bonds between the glycerol carbon and fatty acid ester carbon of triglycerides [27,59] | ||
| 1122 | symmetric phosphodiester stretching band mainly of RNA ([60] and references therein) | ||
| 1103 | PO2− symmetric stretching and C-O stretching of deoxyribose [52] | ||
| 1117 1096 | –C–O stretching of esters of fatty acids [27,45] and PO2− symmetric stretching of phospholipids [43] | ||
| 1082 | C-C stretching of glycogen [28,61,62] | 1081 | C-O and C-C stretching of glycated collagen [16,63] and PO2− symmetric stretching of nucleic acids ([28,56] and references therein) |
| 1060 | C-O stretching of P-O-C of phospholipids [64] | ||
| 1041 | C-C-O and C-O-H bending of glycated collagen [16,63] and C-O stretching of RNA D-ribose (skeletal motions of nucleic acids) ([56] and references therein) | ||
| 1023 | C-O-H bending of glycogen [28,52,61,62] | ||
| 987 | phospholipids [43] and =C-H bending of monosaccharides and polysaccharides [30] | ||
| 972 (952 shoulder) 929 | C-C stretching of the Z-DNA backbone [65,66] and Ribose phosphate main chain vibration of RNA backbone [65] | ||
| 964 (947 shoulder) | =C-H out-of-plane bending of unsaturated fatty acids [27,49] and possibly with contribution of N+-(CH3)3 vibration of phospholipids [43] | ||
| 913 891 | fatty acids [27] and phospholipids [43] | ||
| 849 830 | tyrosine [67,68] | ||
| 843 (857 shoulder) | fatty acids [27] and phospholipids [43] and possibly with contribution of glycogen [21] | ||
| 828 | P-O antisymmetric stretching of P-O-C of phospholipids [43] | ||
| 792 | guanine C3′-endo/syn conformation in the Z-DNA [69] | ||
| 779 and 772 | guanine–uracil wobble base pair [70,71] | ||
| 767 | CH2 rocking of glycogen [21] | ||
| 722 (692 shoulder) | CH2 rocking of unsaturated aliphatic chains [72] and =C–H out-of-plane bending of unsaturated fatty acids [27,73] | 739 699 661 | amide IV O=C–N deformation and amide V N–H out-of-plane deformation [23] |
| Comparison | Studied Effect/ Purpose of Comparison | Statistically Significant Differences in Macromolecular Composition (Based on Results Shown in Figure 4 Combined with Band Assignments from Table 2) | ||
|---|---|---|---|---|
| c1 | c2 | |||
| Between the groups | STZ-DM relative to Y-NDM | Changes in the muscle composition in STZ-induced type 1 DM relative to the control group/possibility of differentiating the two groups. | Decreased c1 for the STZ-DM group compared to the Y-NDM group means decreased:
| Increased c2 for the STZ-DM group compared to the Y-NDM group means increased:
|
| Changed overall protein composition patterns. | ||||
| O-NDM relative to Y-NDM | Changes in the muscle composition in relation to age/possibility of differentiating the two groups. | - | - | |
| O-HFD-DM relative to Y-NDM | Changes in the muscle composition in relation to age and obesity with insulin resistance. | - | - | |
| O-HFD-DM relative to O-NDM | Changes in the muscle composition in relation to obesity with insulin resistance relative to the control group/possibility of differentiating the two groups. | - | - | |
| O-NDM relative to STZ-DM | Differentiation between muscle composition affected by age and muscle composition affected by STZ-induced type 1 DM/possibility of differentiating the two groups. | Increased c1 for the O-NDM group compared to the STZ-DM group means increased:
| Decreased c2 for the O-NDM group compared to the STZ-DM group means decreased:
| |
| Changed overall protein composition patterns. | ||||
| O-HFD-DM relative to STZ-DM | Differentiation between muscle composition affected by age and obesity with insulin resistance factors, and muscle composition affected by STZ-induced type 1 DM/possibility of differentiating muscle samples based on FTIR spectrum. | Increased c1 for the O-HFD-DM group compared to the STZ-DM group means increased:
| - | |
| Changed overall protein composition patterns. | ||||
| Gluteus maximus(GM) vs. gastrocnemius (GA) muscle composition within the group | Y-NDM (GM relative to GA muscle) | Differentiation between the composition of the gluteus maximus and the gastrocnemius muscle at a younger age/possibility of differentiating the non-weight-bearing and weight-bearing muscles. | Increased c1 for the GM muscle compared to the GA muscle of the Y-NDM group means increased:
| Decreased c1 for the GM muscle compared to the GA muscle of the Y-NDM group means decreased:
|
| Changed overall protein composition patterns. | ||||
| Y-STZ-DM (GM relative to GA muscle) | Differentiation between the composition of the gluteus maximus and the gastrocnemius muscle affected by STZ-induced type 1/possibility of differentiating the non-weight-bearing and weight-bearing muscles. | Increased c1 for the GM muscle compared to the GA muscle of the Y-STZ-DM group means increased:
| ||
| Changed overall protein composition patterns. | ||||
| O-NDM (GM relative to GA muscle) | Differentiation between the composition of the gluteus maximus and the gastrocnemius muscle in older age/possibility of differentiating the non-weight-bearing and weight-bearing muscles. | Increased c1 for the GM muscle compared to the GA muscle of the O-NDM group means increased:
| Decreased c2 for the GM muscle compared to the GA muscle of the O-NDM group means decreased:
| |
| Changed overall protein composition patterns. | ||||
| O-HFD-DM (GM relative to GA muscle) | Differentiation between the composition of the gluteus maximus and the gastrocnemius muscle affected by age and obesity with insulin resistance factors/possibility of differentiating the non-weight-bearing and weight-bearing muscles. | Increased c1 for the GM muscle compared to the GA muscle of the O-HFD-DM group means increased:
| ||
| Changed overall protein composition patterns. | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zupančič, B.; Umek, N.; Ugwoke, C.K.; Cvetko, E.; Horvat, S.; Grdadolnik, J. Application of FTIR Spectroscopy to Detect Changes in Skeletal Muscle Composition Due to Obesity with Insulin Resistance and STZ-Induced Diabetes. Int. J. Mol. Sci. 2022, 23, 12498. https://doi.org/10.3390/ijms232012498
Zupančič B, Umek N, Ugwoke CK, Cvetko E, Horvat S, Grdadolnik J. Application of FTIR Spectroscopy to Detect Changes in Skeletal Muscle Composition Due to Obesity with Insulin Resistance and STZ-Induced Diabetes. International Journal of Molecular Sciences. 2022; 23(20):12498. https://doi.org/10.3390/ijms232012498
Chicago/Turabian StyleZupančič, Barbara, Nejc Umek, Chiedozie Kenneth Ugwoke, Erika Cvetko, Simon Horvat, and Jože Grdadolnik. 2022. "Application of FTIR Spectroscopy to Detect Changes in Skeletal Muscle Composition Due to Obesity with Insulin Resistance and STZ-Induced Diabetes" International Journal of Molecular Sciences 23, no. 20: 12498. https://doi.org/10.3390/ijms232012498
APA StyleZupančič, B., Umek, N., Ugwoke, C. K., Cvetko, E., Horvat, S., & Grdadolnik, J. (2022). Application of FTIR Spectroscopy to Detect Changes in Skeletal Muscle Composition Due to Obesity with Insulin Resistance and STZ-Induced Diabetes. International Journal of Molecular Sciences, 23(20), 12498. https://doi.org/10.3390/ijms232012498