
Aberrant Transferrin and Ferritin Upregulation Elicits Iron Accumulation and Oxidative Inflammaging Causing Ferroptosis and Undermines Estradiol Biosynthesis in Aging Rat Ovaries by Upregulating NF-Κb-Activated Inducible Nitric Oxide Synthase: First Demonstration of an Intricate Mechanism

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
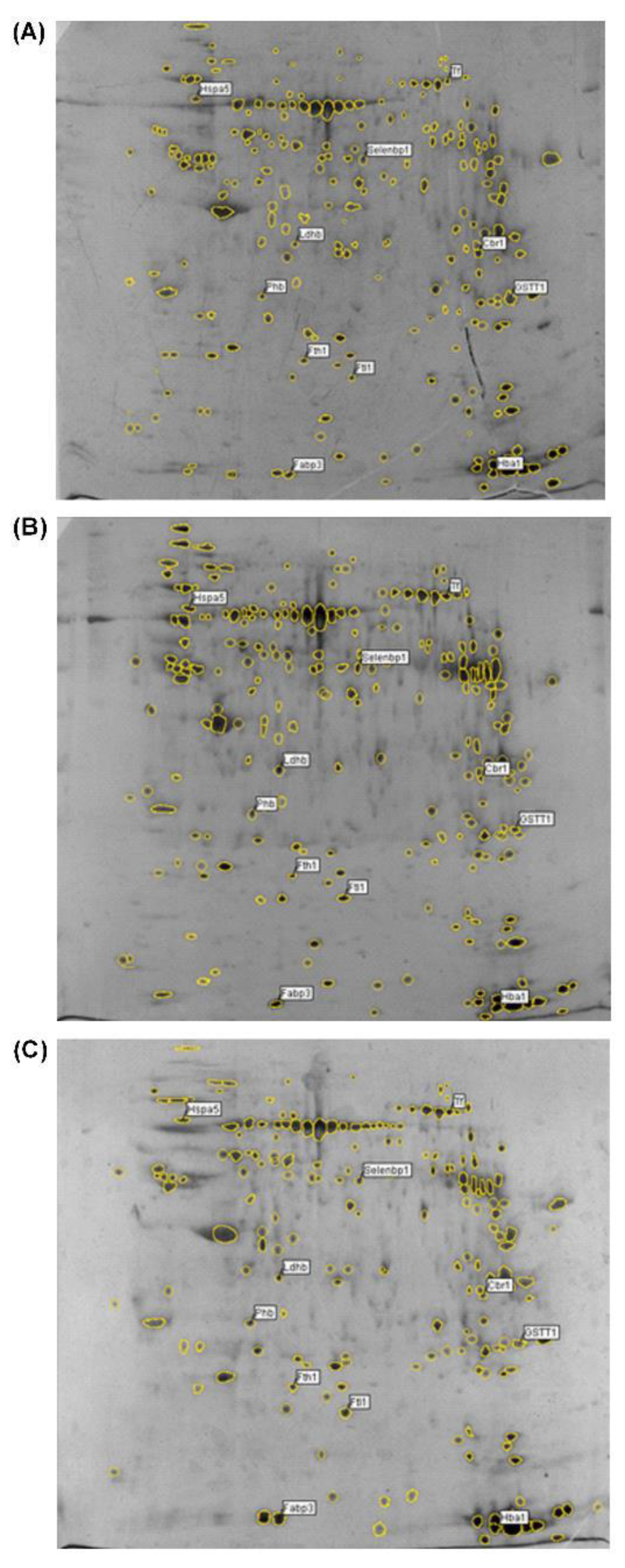
2.1. Age-Associated Changes in the Expression of Proteins in Rat Ovarian Tissue Demonstrated by Two-Dimensional Gel-Based Proteomics

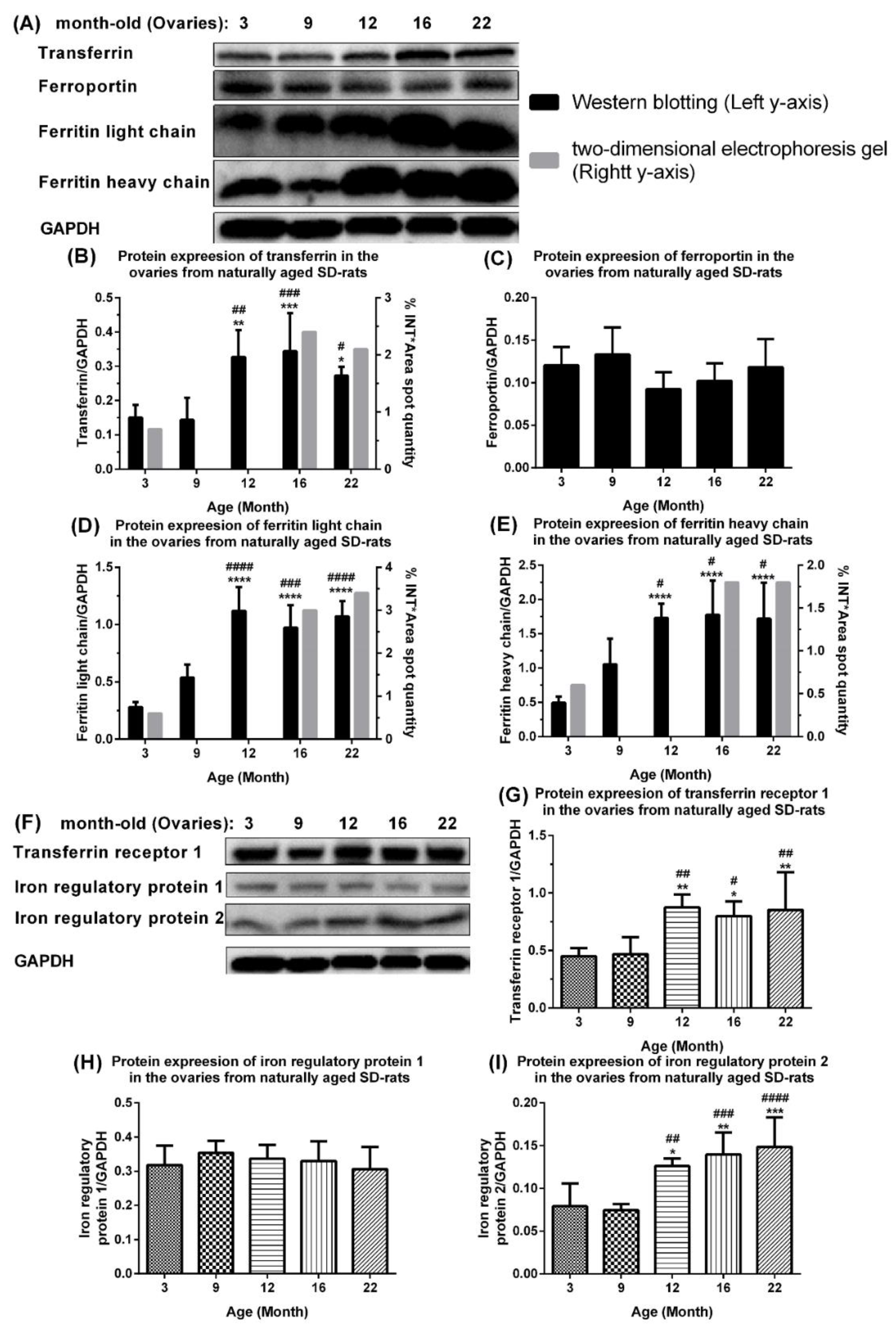
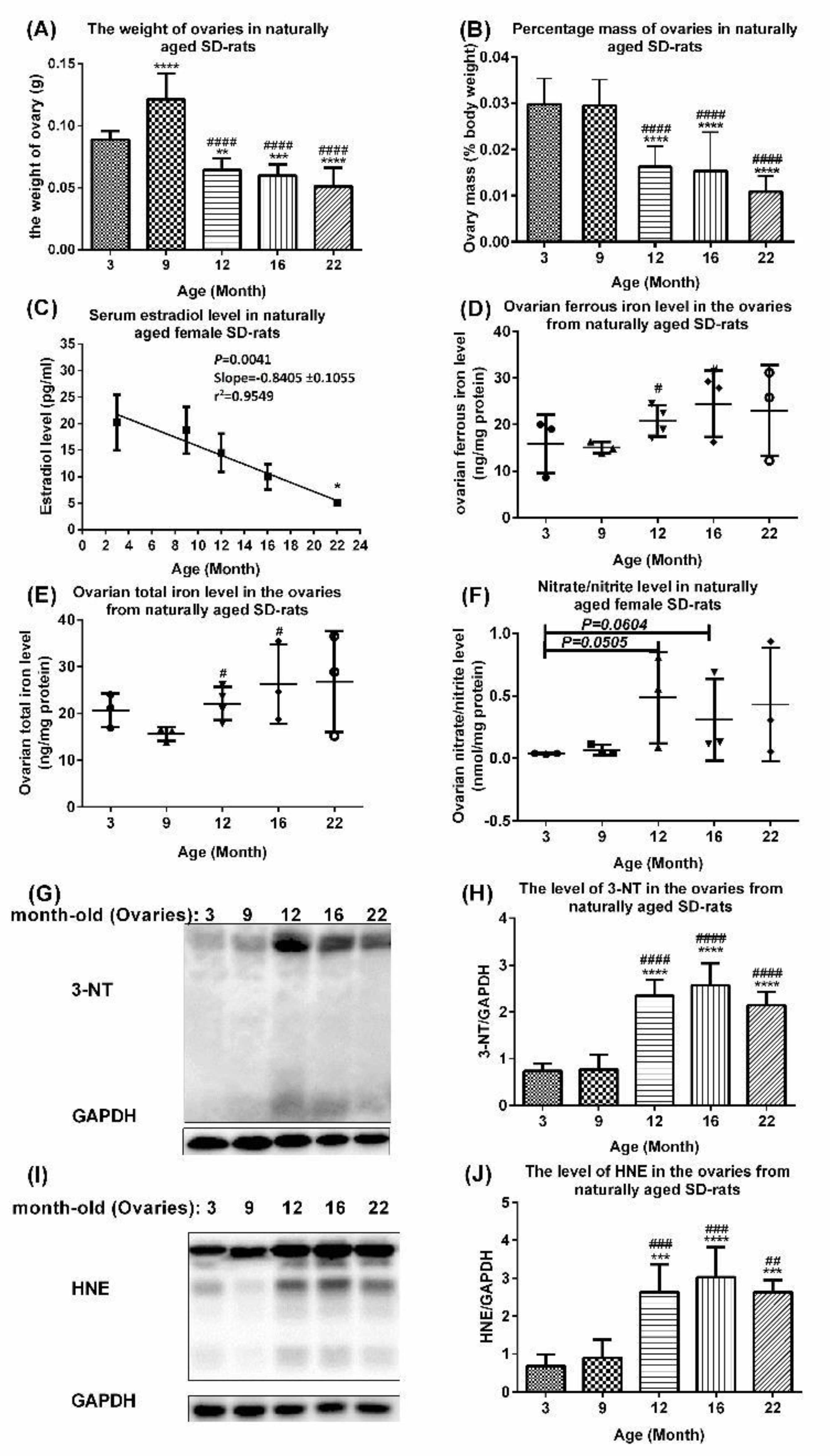
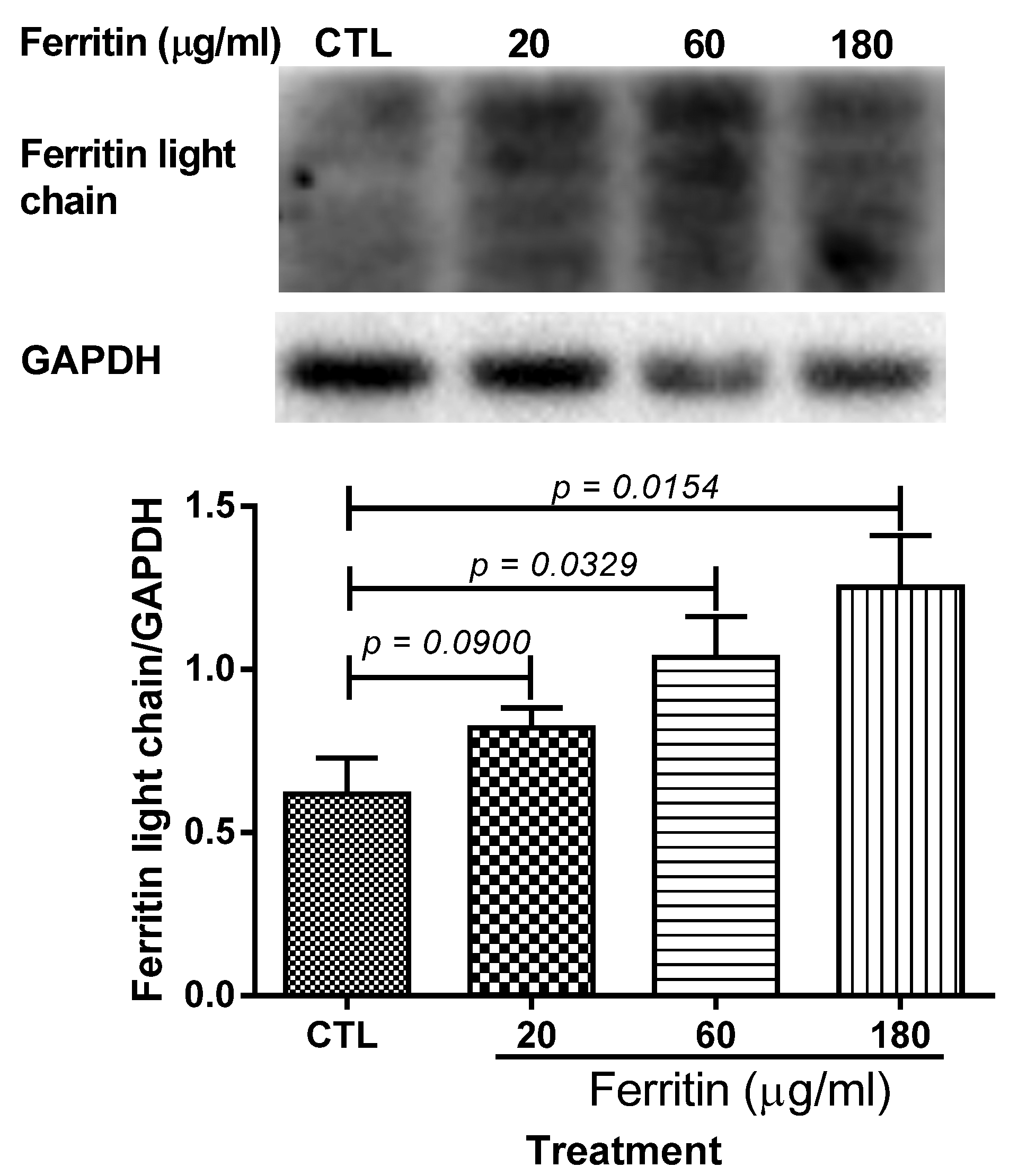
2.2. Aberrant Upregulation of Transferrin, Ferritin, Transferrin Receptor 1, and Iron Regulatory Protein 2 in Ovaries of Naturally Aging Rats



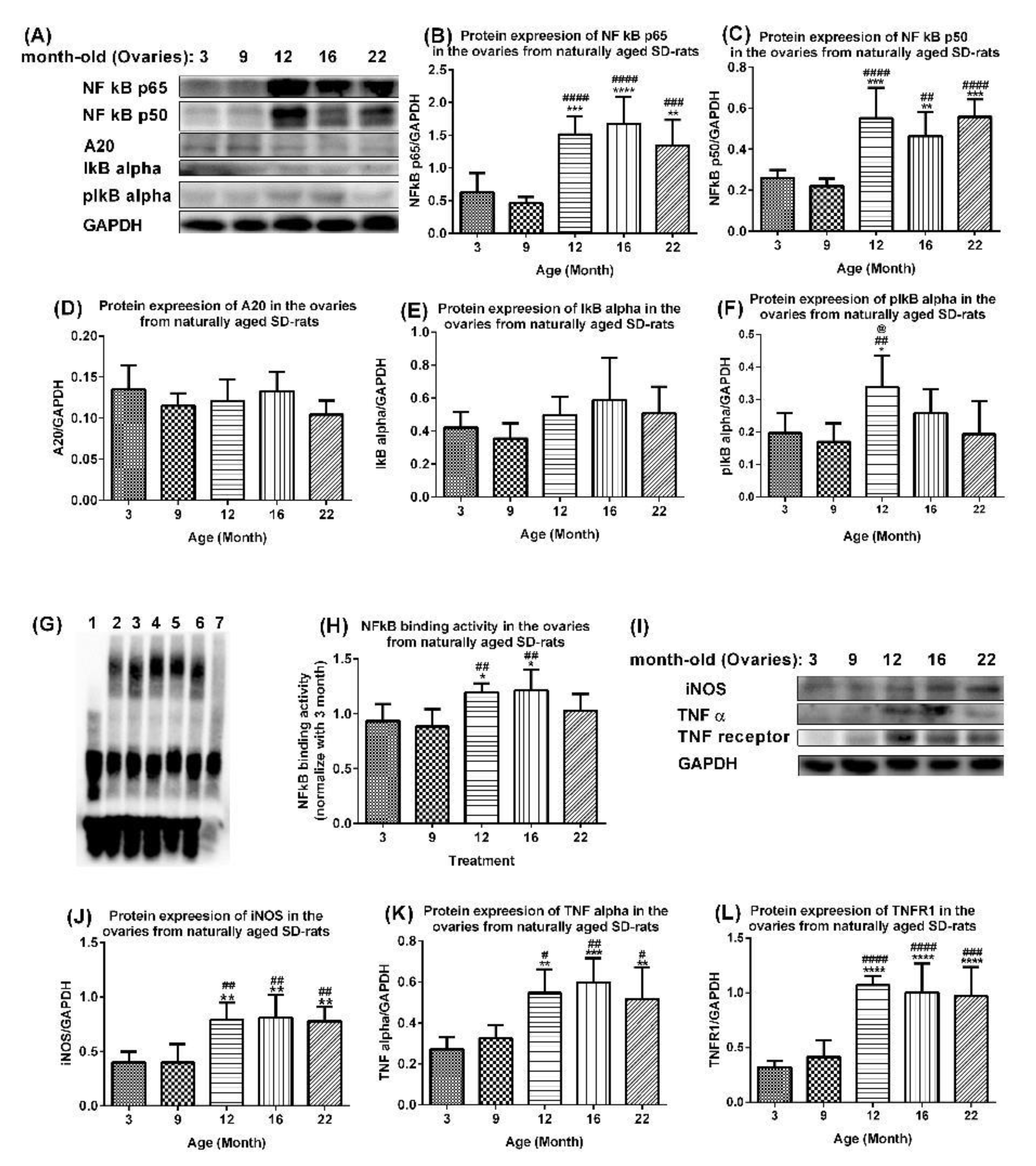
2.3. Age-Associated Aberrant Activation of NF-Κb/Inos and TNF Alpha Pathways in Ovaries of Aging Rats

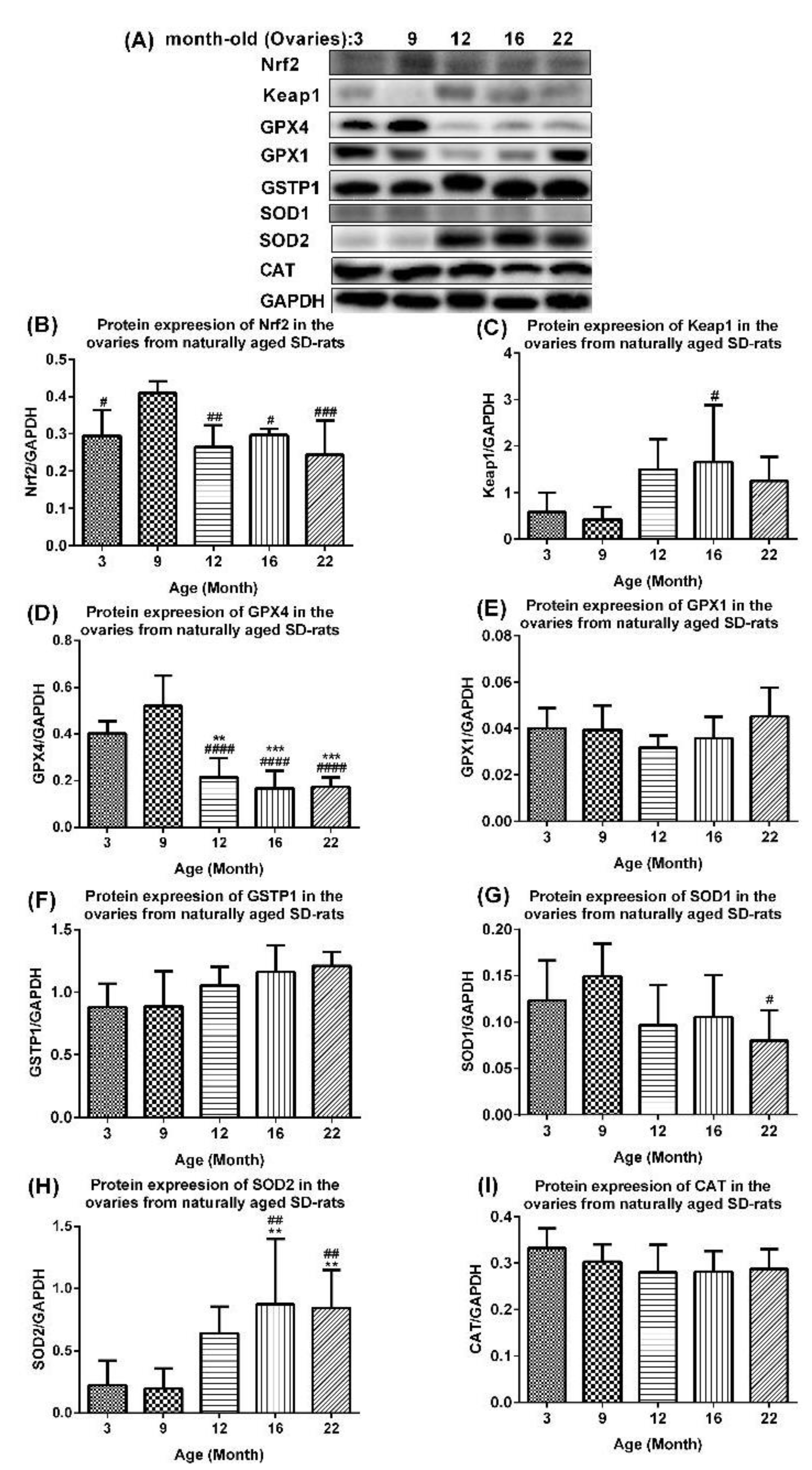
2.4. Age-Associated Aberrant Inhibition of Nrf2/GPX4 Pathway in Ovaries of Aging Rats

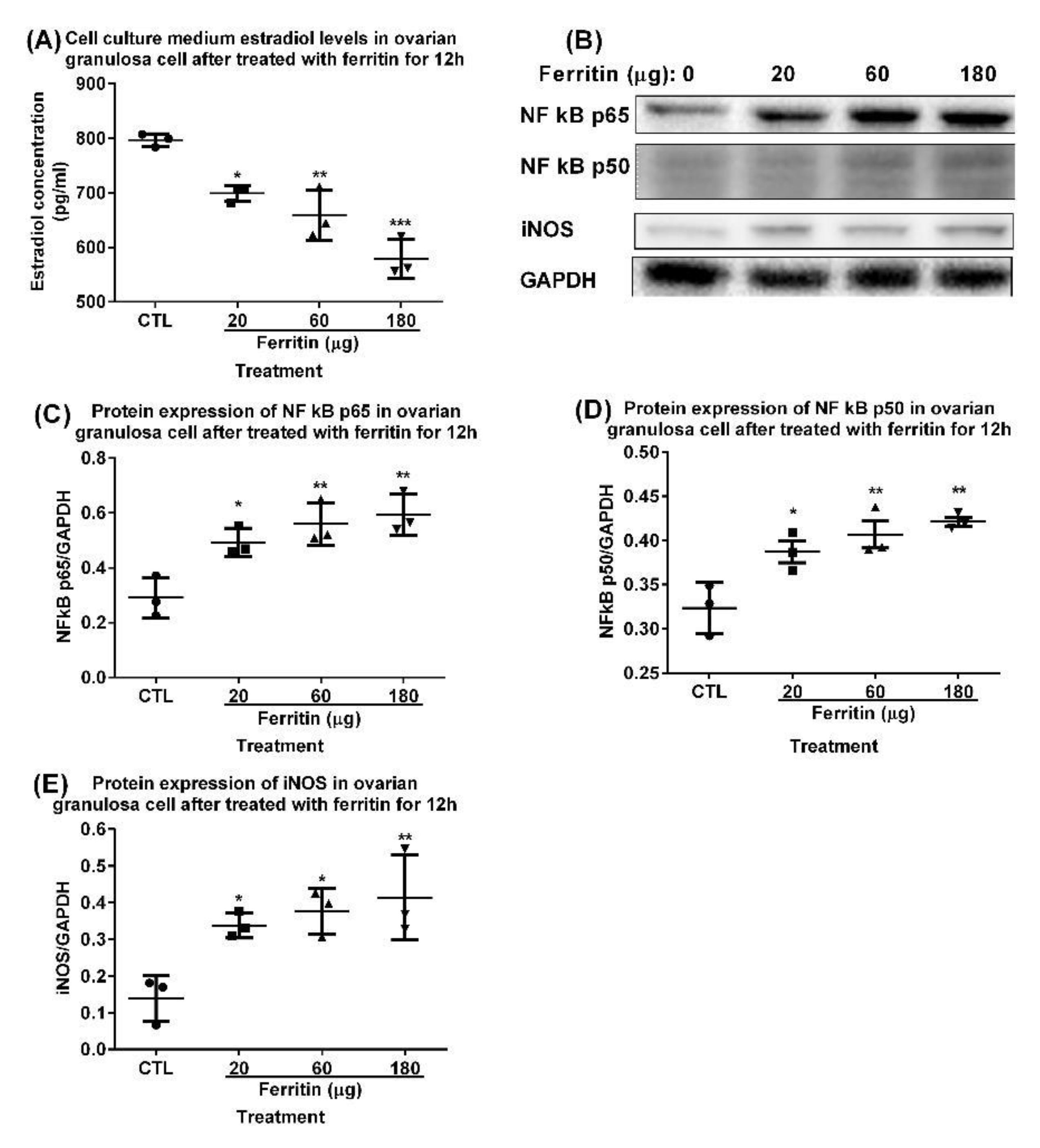
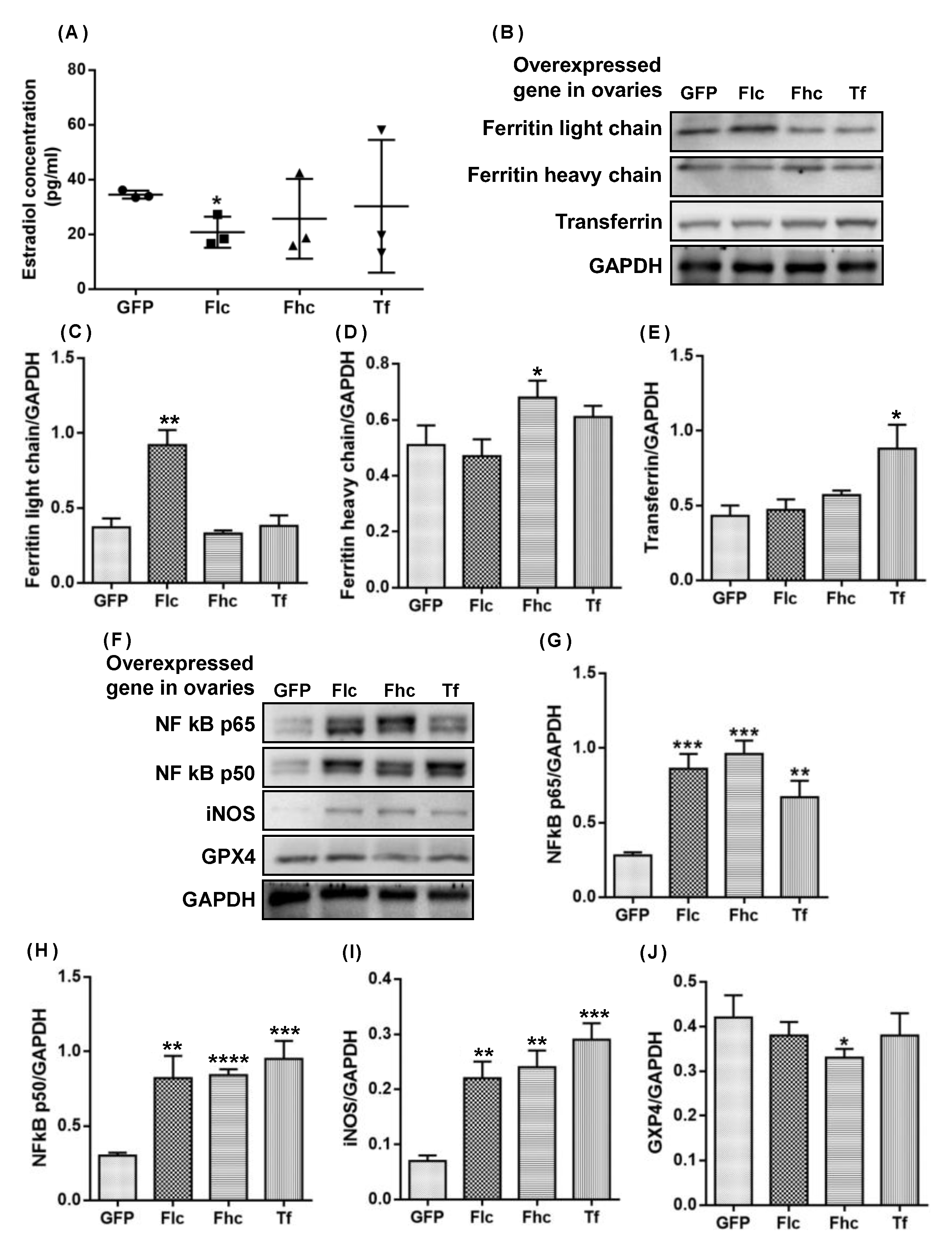
2.5. Intraovarian Injection of Adenovirus Overexpressing Ferritin Light Chain/Heavy Chain and Transferrin Induced Upregulation of NF Kb P65/P50 Mediated Inos and Downregulation of GPX4 in Ovaries of 3-Month-Old Rats


3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Selesniemi, K.; Lee, H.J.; Niikura, T.; Tilly, J.L. Young adult donor bone marrow infusions into female mice postpone age-related reproductive failure and improve offspring survival. Aging 2009, 1, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Singh, P. Andropause: Current concepts. Indian J. Endocrinol. Metab. 2013, 17 (Suppl. 3), S621–S629. [Google Scholar] [CrossRef] [PubMed]
- Perheentupa, A.; Huhtaniemi, I. Aging of the human ovary and testis. Mol. Cell Endocrinol. 2009, 299, 2–13. [Google Scholar] [CrossRef]
- Sharov, A.A.; Falco, G.; Piao, Y.; Poosala, S.; Becker, K.G.; Zonderman, A.B.; Longo, D.L.; Schlessinger, D.; Ko, M.S.H. Effects of aging and calorie restriction on the global gene expression profiles of mouse testis and ovary. BMC Biol. 2008, 6, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broekmans, F.J.; Soules, M.R.; Fauser, B.C. Ovarian Aging: Mechanisms and Clinical Consequences. Endocr. Rev. 2009, 30, 465–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hickey, M.; Elliott, J.; Davison, S.L. Hormone replacement therapy. BMJ 2012, 344, e763. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.; Nan, B.; Kong, S.; Harlow, S. Changes in iron measures over menopause and associations with insulin resistance. J. Women’s Health 2012, 21, 872–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jian, J.; Pelle, E.; Huang, X. Iron and menopause: Does increased iron affect the health of postmenopausal women? Antioxid. Redox Signal. 2009, 11, 2939–2943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asano, Y. Age-related accumulation of non-heme ferric and ferrous iron in mouse ovarian stroma visualized by sensitive non-heme iron histochemistry. J. Histochem. Cytochem. 2012, 60, 229–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemeth, E.; Ganz, T. Hepcidin-Ferroportin Interaction Controls Systemic Iron Homeostasis. Int. J. Mol. Sci. 2021, 22, 6493. [Google Scholar] [CrossRef]
- Wish, J.B. Assessing iron status: Beyond serum ferritin and transferrin saturation. Clin. J. Am. Soc. Nephrol. 2006, 1 (Suppl. 1), S4–S8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, N.; Pantopoulos, K. The IRP/IRE system in vivo: Insights from mouse models. Front. Pharmacol. 2014, 5, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Styś, A.; Galy, B.; Starzyński, R.R.; Smuda, E.; Drapier, J.C.; Lipiński, P.; Bouton, C. Iron regulatory protein 1 outcompetes iron regulatory protein 2 in regulating cellular iron homeostasis in response to nitric oxide. J. Biol. Chem. 2011, 286, 22846–22854. [Google Scholar] [CrossRef] [Green Version]
- Lieu, P.T.; Heiskala, M.; Peterson, P.A.; Yang, Y. The roles of iron in health and disease. Mol. Aspects Med. 2001, 22, 1–87. [Google Scholar] [CrossRef]
- Chenoufi, N.; Loreal, O.; Drenou, B.; Cariou, S.; Hubert, N.; Leroyer, P.; Brissot, P.; Lescoat, G. Iron may induce both DNA synthesis and repair in rat hepatocytes stimulated by EGF/pyruvate. J. Hepatol. 1997, 26, 650–658. [Google Scholar] [CrossRef]
- Masini, A.; Trenti, T.; Ceccarelli-Stanzani, D.; Ventura, E. The effect of ferric iron complex on isolated rat liver mitochondria. I. Respiratory and electrochemical responses. Biochim. Biophys. Acta 1985, 810, 20–26. [Google Scholar] [CrossRef]
- Valberg, L.S.; Sorbie, J.; Ludwig, J.; Pelletier, O. Serum ferritin and the iron status of Canadians. Can. Med. Assoc. J. 1976, 114, 417–421. [Google Scholar] [PubMed]
- De Domenico, I.; McVey Ward, D.; Kaplan, J. Regulation of iron acquisition and storage: Consequences for iron-linked disorders. Nat. Rev. Mol. Cell Biol. 2008, 9, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Das, S.K.; Patel, V.B.; Basu, R.; Wang, W.; DesAulniers, J.; Kassiri, Z.; Oudit, G.Y. Females Are Protected from Iron-Overload Cardiomyopathy Independent of Iron Metabolism: Key Role of Oxidative Stress. J. Am. Heart Assoc. 2017, 6, e003456. [Google Scholar] [CrossRef]
- Walter, P.B.; Fung, E.B.; Killilea, D.W.; Jiang, Q.; Hudes, M.; Madden, J.; Porter, J.; Evans, P.; Vichinsky, E.; Harmatz, P. Oxidative stress and inflammation in iron-overloaded patients with beta-thalassaemia or sickle cell disease. Br. J. Haematol. 2006, 135, 254–263. [Google Scholar] [CrossRef]
- Anderson, E.; Batten, B.E. Surface binding, uptake and fate of cationic ferritin in a steroid producing ovarian cell. Tissue Cell 1983, 15, 853–871. [Google Scholar] [CrossRef]
- Kim, O.Y.; Chae, J.S.; Paik, J.K.; Seo, H.S.; Jang, Y.; Cavaillon, J.M.; Lee, J.H. Effects of aging and menopause on serum interleukin-6 levels and peripheral blood mononuclear cell cytokine production in healthy nonobese women. Age 2012, 34, 415–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salminen, A.; Huuskonen, J.; Ojala, J.; Kauppinen, A.; Kaarniranta, K.; Suuronen, T. Activation of innate immunity system during aging: NF-kB signaling is the molecular culprit of inflamm-aging. Ageing Res. Rev. 2008, 7, 83–105. [Google Scholar] [CrossRef] [PubMed]
- Osorio, F.G.; Lopez-Otin, C.; Freije, J.M. NF-kB in premature aging. Aging 2012, 4, 726–727. [Google Scholar] [CrossRef] [PubMed]
- Li, W.G.; Khor, T.O.; Xu, C.J.; Shen, G.X.; Jeong, W.S.; Yu, S.; Kong, A.N. Activation of Nrf2-antioxidant signaling attenuates NF kappa B-inflammatory response and elicits apoptosis. Biochem. Pharmacol. 2008, 76, 1485–1489. [Google Scholar] [CrossRef] [Green Version]
- Brigelius-Flohe, R.; Friedrichs, B.; Maurer, S.; Schultz, M.; Streicher, R. Interleukin-1-induced nuclear factor kappa B activation is inhibited by overexpression of phospholipid hydroperoxide glutathione peroxidase in a human endothelial cell line. Biochem. J. 1997, 328 Pt 1, 199–203. [Google Scholar] [CrossRef] [Green Version]
- Zacharski, L.R.; Ornstein, D.L.; Woloshin, S.; Schwartz, L.M. Association of age, sex, and race with body iron stores in adults: Analysis of NHANES III data. Am. Heart J. 2000, 140, 98–104. [Google Scholar] [CrossRef]
- Ibanez, L. Early development and pubertal prevention of polycystic ovary syndrome. In Endocrine Abstracts; Bioscientifica: Bristol, UK, 2012. [Google Scholar]
- Lehmann, D.J.; Worwood, M.; Ellis, R.; Wimhurst, V.L.; Merryweather-Clarke, A.T.; Warden, D.R.; Smith, A.D.; Robson, K.J. Iron genes, iron load and risk of Alzheimer’s disease. J. Med. Genet. 2006, 43, e52. [Google Scholar] [CrossRef] [Green Version]
- Briggs, D.A.; Sharp, D.J.; Miller, D.; Gosden, R.G. Transferrin in the developing ovarian follicle: Evidence for de-novo expression by granulosa cells. Mol. Hum. Reprod. 1999, 5, 1107–1114. [Google Scholar] [CrossRef]
- Kawano, Y.; Narahara, H.; Miyamura, K.; Mifune, K.; Miyakawa, I. Inhibitory effect of transferrin on progesterone production in the granulosa cell of humans in vivo and porcine granulosa cell in vitro. Gynecol. Obstet. Investig. 1995, 40, 1–4. [Google Scholar] [CrossRef]
- Pantopoulos, K. Iron metabolism and the IRE/IRP regulatory system: An update. Ann. N. Y. Acad. Sci. 2004, 1012, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Muckenthaler, M.U.; Galy, B.; Hentze, M.W. Systemic iron homeostasis and the iron-responsive element/iron-regulatory protein (IRE/IRP) regulatory network. Annu. Rev. Nutr. 2008, 28, 197–213. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Ma, J.; Walden, W.E.; Merrick, W.C.; Theil, E.C.; Goss, D.J. Rapid kinetics of iron responsive element (IRE) RNA/iron regulatory protein 1 and IRE-RNA/eIF4F complexes respond differently to metal ions. Nucleic Acids Res. 2014, 42, 6567–6577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Shi, H.; Rajan, M.; Canarie, E.R.; Hong, S.; Simoneschi, D.; Pagano, M.; Bush, M.F.; Stoll, S.; Leibold, E.A.; et al. FBXL5 Regulates IRP2 Stability in Iron Homeostasis via an Oxygen-Responsive [2Fe2S] Cluster. Mol. Cell 2020, 78, 31–41.e5. [Google Scholar] [CrossRef]
- Zhang, D.L.; Ghosh, M.C.; Rouault, T.A. The physiological functions of iron regulatory proteins in iron homeostasis-an update. Front. Pharmacol. 2014, 5, 124. [Google Scholar] [CrossRef] [Green Version]
- Peters, H.; Aschheim, P.; Jacobs, H.S.; Rubens, R.; Talbert, G.B. The ageing ovary. In Female and Male Climacteric; Springer: Dordrecht, The Netherlands, 1979. [Google Scholar]
- Sengupta, P. A scientific review of age determination for a laboratory rat: How old is it in comparison with human age? Bio Med. Int. 2015, 2, 81–89. [Google Scholar]
- Sowers, M.R.; Randolph, J.F.; Zheng, H.; Jannausch, M.; McConnell, D.; Kardia, S.R.; Crandall, C.J.; Nan, B. Genetic polymorphisms and obesity influence estradiol decline during the menopause. Clin. Endocrinol. 2011, 74, 618–623. [Google Scholar] [CrossRef]
- Lim, J.; Luderer, U. Oxidative damage increases and antioxidant gene expression decreases with aging in the mouse ovary. Biol. Reprod. 2011, 84, 775–782. [Google Scholar] [CrossRef] [Green Version]
- Singer, S.T.; Vichinsky, E.P.; Gildengorin, G.; van Disseldorp, J.; Rosen, M.; Cedars, M.I. Reproductive capacity in iron overloaded women with thalassemia major. Blood 2011, 118, 2878–2881. [Google Scholar] [CrossRef] [Green Version]
- Siemons, L.J.; Mahler, C.H. Hypogonadotropic hypogonadism in hemochromatosis: Recovery of reproductive function after iron depletion. J. Clin. Endocrinol. Metab. 1987, 65, 585–587. [Google Scholar] [CrossRef]
- Reubinoff, B.E.; Har-El, R.; Kitrossky, N.; Friedler, S.; Levi, R.; Lewin, A.; Chevion, M. Increased levels of redox-active iron in follicular fluid: A possible cause of free radical-mediated infertility in beta-thalassemia major. Am. J. Obstet. Gynecol. 1996, 174, 914–918. [Google Scholar] [CrossRef]
- Bresgen, N.; Jaksch, H.; Lacher, H.; Ohlenschlager, I.; Uchida, K.; Eckl, P.M. Iron-mediated oxidative stress plays an essential role in ferritin-induced cell death. Free Radic. Biol. Med. 2010, 48, 1347–1357. [Google Scholar] [CrossRef] [PubMed]
- Kubrina, L.N.; Mikoyan, V.D.; Mordvintcev, P.I.; Vanin, A.F. Iron potentiates bacterial lipopolysaccharide-induced nitric oxide formation in animal organs. Biochim. Et Biophys. Acta 1993, 1176, 240–244. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, B.H.; Harris, D.C. Evidence suggesting that nitric oxide mediates iron-induced toxicity in cultured proximal tubule cells. Am. J. Physiol. 1998, 274, F18–F25. [Google Scholar] [CrossRef] [PubMed]
- Tobai, H.; Nishiya, I. Nitric oxide mediates inhibitory effect of interleukin-1beta on estrogen production in human granulosa-luteal cells. J. Obstet. Gynaecol. Res. 2001, 27, 53–59. [Google Scholar] [CrossRef]
- Rubbo, H.; Radi, R.; Trujillo, M.; Telleri, R.; Kalyanaraman, B.; Barnes, S.; Kirk, M.; Freeman, B.A. Nitric oxide regulation of superoxide and peroxynitrite-dependent lipid peroxidation. Formation of novel nitrogen-containing oxidized lipid derivatives. J. Biol. Chem. 1994, 269, 26066–26075. [Google Scholar] [CrossRef]
- Kolesarova, A.; Capcarova, M.; Medvedova, M.; Sirotkin, A.V.; Kovacik, J. In vitro assessment of iron effect on porcine ovarian granulosa cells: Secretory activity, markers of proliferation and apoptosis. Physiol. Res. 2011, 60, 503–510. [Google Scholar] [CrossRef]
- Pavlova, S.; Klucska, K.; Vasicek, D.; Kotwica, J.; Sirotkin, A.V. Transcription factor NF-kappaB (p50/p50, p65/p65) controls porcine ovarian cells functions. Anim. Reprod. Sci. 2011, 128, 73–84. [Google Scholar] [CrossRef]
- Xiong, S.; She, H.; Sung, C.K.; Tsukamoto, H. Iron-dependent activation of NF-kappaB in Kupffer cells: A priming mechanism for alcoholic liver disease. Alcohol 2003, 30, 107–113. [Google Scholar] [CrossRef]
- Ruddell, R.G.; Hoang-Le, D.; Barwood, J.M.; Rutherford, P.S.; Piva, T.J.; Watters, D.J.; Santambrogio, P.; Arosio, P.; Ramm, G.A. Ferritin Functions as a Proinflammatory Cytokine via Iron-Independent Protein Kinase C Zeta/Nuclear Factor KappaB-Regulated Signaling in Rat Hepatic Stellate Cells. Hepatology 2009, 49, 887–900. [Google Scholar] [CrossRef] [Green Version]
- Zoll, J.; Melchers, W.J.; Galama, J.M.; van Kuppeveld, F.J. The mengovirus leader protein suppresses alpha/beta interferon production by inhibition of the iron/ferritin-mediated activation of NF-kappa B. J. Virol. 2002, 76, 9664–9672. [Google Scholar] [CrossRef] [PubMed]
- Xiong, S.; She, H.; Takeuchi, H.; Han, B.; Engelhardt, J.F.; Barton, C.H.; Zandi, E.; Giulivi, C.; Tsukamoto, H. Signaling role of intracellular iron in NF-kappaB activation. J. Biol. Chem. 2003, 278, 17646–17654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimon, A.; Erat, A.; Von Wald, T.; Bissell, B.; Koulova, A.; Choi, C.H.; Bachvarov, D.; Reindollar, R.H.; Usheva, A. Genes invoked in the ovarian transition to menopause. Nucleic Acids Res. 2006, 34, 3279–3287. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb Perspect Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlos, K.B.F. The Biology of Nuclear Factor Kappa Beta (NF kB) in Health and Pathology. Res. World–Int. Refereed J. Arts Sci. Commer. Res. 2011, 2, 53–59. [Google Scholar]
- Khedr, N.F. Protective effect of mirtazapine and hesperidin on cyclophosphamide-induced oxidative damage and infertility in rat ovaries. Exp. Biol. Med. 2015, 240, 1682–1689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Assar, M.; Angulo, J.; Rodriguez-Manas, L. Oxidative stress and vascular inflammation in aging. Free Radic. Biol. Med. 2013, 65, 380–401. [Google Scholar] [CrossRef]
- Volonte, D.; Liu, Z.; Musille, P.M.; Stoppani, E.; Wakabayashi, N.; Di, Y.P.; Lisanti, M.P.; Kensler, T.W.; Galbiati, F. Inhibition of nuclear factor-erythroid 2-related factor (Nrf2) by caveolin-1 promotes stress-induced premature senescence. Mol. Biol. Cell 2013, 24, 1852–1862. [Google Scholar] [CrossRef]
- Cole-Ezea, P.; Swan, D.; Shanley, D.; Hesketh, J. Glutathione peroxidase 4 has a major role in protecting mitochondria from oxidative damage and maintaining oxidative phosphorylation complexes in gut epithelial cells. Free Radic. Biol. Med. 2012, 53, 488–497. [Google Scholar] [CrossRef]
- Aramaki, H. Recent advances in the mechanistic understanding of endocrine disruption by environmental chemicals. Foreword. Biol. Pharm. Bull. 2014, 37, 1429. [Google Scholar] [CrossRef] [Green Version]
- Yant, L.J.; Ran, Q.T.; Rao, L.; Van Remmen, H.; Shibatani, T.; Belter, J.G.; Motta, L.; Richardson, A.; Prolla, T.A. The selenoprotein GPX4 is essential for mouse development and protects from radiation and oxidative damage insults. Free. Radic. Biol. Med. 2003, 34, 496–502. [Google Scholar] [CrossRef]
- Muller, F.L.; Lustgarten, M.S.; Jang, Y.; Richardson, A.; Van Remmen, H. Trends in oxidative aging theories. Free. Radic. Biol. Med. 2007, 43, 477–503. [Google Scholar] [CrossRef] [PubMed]
- Viswanathan, V.S.; Cheah, J.H.; Clemons, P.A.; Shamji, A.F.; Clish, C.B.; Yang, W.S.; SriRamaratnam, R.; Welsch, M.E.; Shimada, K.; Skouta, R.; et al. Regulation of Ferroptotic Cancer Cell Death by GPX4. Cell 2014, 156, 317–331. [Google Scholar] [CrossRef] [Green Version]
- Angeli, J.P.F.; Schneider, M.; Proneth, B.; Tyurina, Y.Y.; Tyurin, V.A.; Hammond, V.J.; Herbach, N.; Aichler, M.; Walch, A.; Eggenhofer, E.; et al. Inactivation of the ferroptosis regulator Gpx4 triggers acute renal failure in mice. Nat. Cell Biol. 2014, 16, 1180–1191. [Google Scholar] [CrossRef] [Green Version]
- Lorincz, T.; Jemnitz, K.; Kardon, T.; Mandl, J.; Szarka, A. Ferroptosis is Involved in Acetaminophen Induced Cell Death. Pathol. Oncol. Res. 2015, 21, 1115–1121. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.J.; Hambright, W.S.; Na, R.; Ran, Q.T. Ablation of the Ferroptosis Inhibitor Glutathione Peroxidase 4 in Neurons Results in Rapid Motor Neuron Degeneration and Paralysis. J. Biol. Chem. 2015, 290, 28097–28106. [Google Scholar] [CrossRef] [Green Version]
- Carbone, M.C.; Tatone, C.; Delle Monache, S.; Marci, R.; Caserta, D.; Colonna, R.; Amicarelli, F. Antioxidant enzymatic defences in human follicular fluid: Characterization and age-dependent changes. Mol. Hum. Reprod. 2003, 9, 639–643. [Google Scholar] [CrossRef]
- Byrd, T.F.; Horwitz, M.A. Regulation of transferrin receptor expression and ferritin content in human mononuclear phagocytes. Coordinate upregulation by iron transferrin and downregulation by interferon gamma. J. Clin. Investig. 1993, 91, 969–976. [Google Scholar] [CrossRef] [Green Version]
- Graziadei, I.; Weiss, G.; Bohm, A.; Werner-Felmayer, G.; Vogel, W. Unidirectional upregulation of the synthesis of the major iron proteins, transferrin-receptor and ferritin, in HepG2 cells by the acute-phase protein alpha1-antitrypsin. J. Hepatol. 1997, 27, 716–725. [Google Scholar] [CrossRef]
- Kobak, K.A.; Franczuk, P.; Schubert, J.; Dzięgała, M.; Kasztura, M.; Tkaczyszyn, M.; Drozd, M.; Kosiorek, A.; Kiczak, L.; Bania, J.; et al. Primary Human Cardiomyocytes and Cardiofibroblasts Treated with Sera from Myocarditis Patients Exbit an Increased Iron Demand and Complex Changes in the Gene Expression. Cells 2021, 10, 818. [Google Scholar] [CrossRef]
- Rossi, E.M.; Marques, V.B.; Nunes Dde, O.; Carneiro, M.T.; Podratz, P.L.; Merlo, E.; dos Santos, L.; Graceli, J.B. Acute iron overload leads to hypothalamic-pituitary-gonadal axis abnormalities in female rats. Toxicol. Lett. 2016, 240, 196–213. [Google Scholar] [CrossRef]
- Li, A.; Ni, Z.; Zhang, J.; Cai, Z.; Kuang, Y.; Yu, C. Transferrin Insufficiency and Iron Overload in Follicular Fluid Contribute to Oocyte Dysmaturity in Infertile Women with Advanced Endometriosis. Front. Endocrinol. 2020, 11, 391. [Google Scholar] [CrossRef]
- Mihalas, B.P.; De Iuliis, G.N.; Redgrove, K.A.; McLaughlin, E.A.; Nixon, B. The lipid peroxidation product 4-hydroxynonenal contributes to oxidative stress-mediated deterioration of the ageing oocyte. Sci. Rep. 2017, 7, 6247. [Google Scholar] [CrossRef] [Green Version]
- Goud, A.P.; Goud, P.T.; Diamond, M.P.; Gonik, B.; Abu-Soud, H.M. Reactive oxygen species and oocyte aging: Role of superoxide, hydrogen peroxide, and hypochlorous acid. Free Radic. Biol. Med. 2008, 44, 1295–1304. [Google Scholar] [CrossRef] [Green Version]
- Park, S.U.; Walsh, L.; Berkowitz, K.M. Mechanisms of ovarian aging. Reproduction 2021, 162, R19–R33. [Google Scholar] [CrossRef]
- Urzua, U.; Chacon, C.; Espinoza, R.; Martinez, S.; Hernandez, N. Parity-Dependent Hemosiderin and Lipofuscin Accumulation in the Reproductively Aged Mouse Ovary. Anal. Cell. Pathol. 2018, 2018, 1289103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tesarik, J.; Galan-Lazaro, M.; Mendoza-Tesarik, R. Ovarian Aging: Molecular Mechanisms and Medical Management. Int. J. Mol. Sci. 2021, 22, 1371. [Google Scholar] [CrossRef]
- Milczarek, R.; Sokołowska, E.; Hallmann, A.; Kaleth, A.K.; Klimek, J. NADPH- and iron-dependent lipid peroxidation inhibit aromatase activity in human placental microsomes. J. Steroid Biochem. Mol. Biol. 2008, 110, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.J.; Chou, C.H.; Shun, C.T.; Mao, T.L.; Wen, W.F.; Chen, C.D.; Chen, S.U.; Yang, Y.S.; Ho, H.N. Iron suppresses ovarian granulosa cell proliferation and arrests cell cycle through regulating p38 mitogen-activated protein kinase/p53/p21 pathway. Biol. Reprod. 2017, 97, 438–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masaldan, S.; Clatworthy, S.A.S.; Gamell, C.; Meggyesy, P.M.; Rigopoulos, A.T.; Haupt, S.; Haupt, Y.; Denoyer, D.; Adlard, P.A.; Bush, A.I.; et al. Iron accumulation in senescent cells is coupled with impaired ferritinophagy and inhibition of ferroptosis. Redox Biol. 2018, 14, 100–115. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Marzetti, E.; Seo, A.Y.; Kim, J.S.; Prolla, T.A.; Leeuwenburgh, C. The emerging role of iron dyshomeostasis in the mitochondrial decay of aging. Mech. Ageing Dev. 2010, 131, 487–493. [Google Scholar] [CrossRef] [Green Version]
- Cornejo, P.; Varela, P.; Videla, L.A.; Fernández, V. Chronic iron overload enhances inducible nitric oxide synthase expression in rat liver. Nitric Oxide 2005, 13, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Aziza, S.A.; Mel-S, A.; El-Shall, S.K. Ameliorating role of rutin on oxidative stress induced by iron overload in hepatic tissue of rats. Pak. J. Biol. Sci. 2014, 17, 964–977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiskirchen, R.; Tacke, F. Cellular and molecular functions of hepatic stellate cells in inflammatory responses and liver immunology. Hepatobiliary Surg. Nutr. 2014, 3, 344–363. [Google Scholar] [CrossRef] [PubMed]
- Reid, D.; Snell, G.; Ward, C.; Krishnaswamy, R.; Ward, R.; Zheng, L.; Williams, T.; Walters, H. Iron overload and nitric oxide-derived oxidative stress following lung transplantation. J. Heart Lung Transplant. 2001, 20, 840–849. [Google Scholar] [CrossRef]
- König, J.; Ott, C.; Hugo, M.; Jung, T.; Bulteau, A.L.; Grune, T.; Höhn, A. Mitochondrial contribution to lipofuscin formation. Redox Biol. 2017, 11, 673–681. [Google Scholar] [CrossRef]
- Qin, X.; Liang, D.; Hu, M.; Lv, Z.; Duan, Z.; Liu, X.; Tang, L. Chronic overload of concentration-dependent iron exerts different effects on ovarian function in C57BL/6J mice. Biol. Reprod. 2021, 104, 1347–1359. [Google Scholar] [CrossRef]
- Foley, K.G.; Pritchard, M.T.; Duncan, F.E. Macrophage-derived multinucleated giant cells: Hallmarks of the aging ovary. Reproduction 2021, 161, V5–V9. [Google Scholar] [CrossRef]
- Huang, Y.; Hu, C.; Ye, H.; Luo, R.; Fu, X.; Li, X.; Huang, J.; Chen, W.; Zheng, Y. Inflamm-Aging: A New Mechanism Affecting Premature Ovarian Insufficiency. J. Immunol. Res. 2019, 2019, 8069898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, S.; Wen, J.; Cheng, J.; Shen, W.; Zhou, S.; Yan, W.; Shen, L.; Luo, A.; Wang, S. Age-associated up-regulation of EGR1 promotes granulosa cell apoptosis during follicle atresia in mice through the NF-κB pathway. Cell Cycle 2016, 15, 2895–2905. [Google Scholar] [CrossRef] [Green Version]
- Lliberos, C.; Liew, S.H.; Mansell, A.; Hutt, K.J. The Inflammasome Contributes to Depletion of the Ovarian Reserve During Aging in Mice. Front. Cell Dev. Biol. 2021, 8, 628473. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, F.; Li, D.; Yan, Y.; Wang, H. Transferrin receptor-mediated reactive oxygen species promotes ferroptosis of KGN cells via regulating NADPH oxidase 1/PTEN induced kinase 1/acyl-CoA synthetase long chain family member 4 signaling. Bioengineered 2021, 12, 4983–4994. [Google Scholar] [CrossRef] [PubMed]
- Jian, J.; Pelle, E.; Yang, Q.; Pernodet, N.; Maes, D.; Huang, X. Iron sensitizes keratinocytes and fibroblasts to UVA-mediated matrix metalloproteinase-1 through TNF-α and ERK activation. Exp. Dermatol. 2011, 20, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Ponka, P. Role of nitric oxide in cellular iron metabolism. Biometals 2003, 16, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Ponka, P. Nitrogen monoxide-mediated control of ferritin synthesis: Implications for macrophage iron homeostasis. Proc. Natl. Acad. Sci. USA 2002, 99, 12214–12219. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Chen, Y.; Liu, Y.; Xing, Y.; Miao, C.; Zhao, Y.; Chang, X.; Zhang, Q. The Role of Oxidative Stress and Natural Antioxidants in Ovarian Aging. Front. Pharmacol. 2021, 11, 617843. [Google Scholar] [CrossRef]
- Budani, M.C.; Tiboni, G.M. Novel Insights on the Role of Nitric Oxide in the Ovary: A Review of the Literature. Int. J. Environ. Res. Public Health 2021, 18, 980. [Google Scholar] [CrossRef]
- Meyron-Holtz, E.G.; Ghosh, M.C.; Iwai, K.; LaVaute, T.; Brazzolotto, X.; Berger, U.V.; Land, W.; Ollivierre-Wilson, H.; Grinberg, A.; Love, P.; et al. Genetic ablations of iron regulatory proteins 1 and 2 reveal why iron regulatory protein 2 dominates iron homeostasis. EMBO J. 2004, 23, 386–395. [Google Scholar] [CrossRef] [Green Version]
- Schalinske, K.L.; Blemings, K.P.; Steffen, D.W.; Chen, O.S.; Eisenstein, R.S. Iron regulatory protein 1 is not required for the modulation of ferritin and transferrin receptor expression by iron in a murine pro-B lymphocyte cell line. Proc. Natl. Acad. Sci. USA 1997, 94, 10681–10686. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Wang, M.Y.; Jiang, H.B.; Guo, C.R.; Zhu, X.D.; Yao, X.Q.; Zeng, W.W.; Zhao, Y.; Chi, L.K. Bisphenol A induces testicular oxidative stress in mice leading to ferroptosis. Asian J. Androl. 2022. [Google Scholar] [CrossRef]
- Hu, D.; Tian, L.; Li, X.; Chen, Y.; Xu, Z.; Ge, R.S.; Wang, Y. Tetramethyl bisphenol a inhibits leydig cell function in late puberty by inducing ferroptosis. Ecotoxicol. Environ. Saf. 2022, 236, 113515. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Liu, Z.; Gao, J.; Li, H.; Wang, X.; Li, Y.; Sun, F. Inhibition of ferroptosis attenuates busulfan-induced oligospermia in mice. Toxicology 2020, 440, 152489. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Schorpp, K.; Jin, J.; Yozwiak, C.E.; Hoffstrom, B.G.; Decker, A.M.; Rajbhandari, P.; Stokes, M.E.; Bender, H.G.; Csuka, J.M.; et al. Transferrin Receptor Is a Specific Ferroptosis Marker. Cell Rep. 2020, 30, 3411–3423.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, M.; Monian, P.; Quadri, N.; Ramasamy, R.; Jiang, X. Glutaminolysis and Transferrin Regulate Ferroptosis. Mol. Cell 2015, 59, 298–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, F.; Cui, X.; Zhang, Y.; Bei, Z.; Wang, H.; Zhao, D.; Wang, H.; Yang, Y. Iron regulatory protein 1 promotes ferroptosis by sustaining cellular iron homeostasis in melanoma. Oncol. Lett. 2021, 22, 657. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jin, C.; Shen, M.; Wang, Z.; Tan, S.; Chen, A.; Wang, S.; Shao, J.; Zhang, F.; Zhang, Z.; et al. Iron regulatory protein 2 is required for artemether-mediated anti-hepatic fibrosis through ferroptosis pathway. Free Radic. Biol. Med. 2020, 160, 845–859. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Accession No. (UniProt) | Protein Score | Protein Score C.I.% | Total Ion Score | Total Ion C.I.% | Protein MW | Protein pI | Pep. Count | Protein Function | Fold Changes (Compared to 3-Month-Old Group) | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 16-Month-Old | 22-Month-Old | Mean | ||||||||||
| 1. Ftl1 (Ferritin light chain 1) | P02793 | 610 | 100 | 463 | 100 | 20,793 | 5.99 | 13 | iron storage | 5 | 5.67 | 5.33 |
| 2. Cbr1 (Carbonyl reductase [NADPH] 1) | P47727 | 305 | 100 | 204 | 100 | 30,844 | 8.22 | 12 | Antioxidative | −3.29 | −3.83 | −3.56 |
| 3. Fabp3(Fatty acid-binding protein) | P07483 | 422 | 100 | 330 | 100 | 14,766 | 5.9 | 9 | lipid transport | 3.44 | 3.39 | 3.42 |
| 4. Ldhb (L-lactate dehydrogenase B chain) | P42123 | 190 | 100 | 131 | 100 | 36,874 | 5.7 | 9 | Oxidoreductase in the process of glycolysis | 3.71 | 2.86 | 3.29 |
| 5. Tf (Transferrin) | P12346 | 319 | 100 | 133 | 100 | 78,513 | 7.14 | 22 | iron transport | 3.43 | 3 | 3.21 |
| 6. Phb (Prohibitin) | P67779 | 93 | 100 | 48 | 99.975 | 29,859 | 5.57 | 7 | inhibition of cell proliferation | 3 | 3.4 | 3.2 |
| 7. Gstt3 (Glutathione S-transferase theta-3) | D3Z8I7 | 307 | 100 | 217 | 100 | 23,822 | 9.3 | 10 | combination of glutathione and oxidants | −3.09 | −3.09 | −3.09 |
| 8. Selenbp2 (Selenium- binding protein 2) | Q8VIF7 | 238 | 100 | 103 | 100 | 53,069 | 6.1 | 17 | sensing of reactive xenobiotics in the cytoplasm, intra-Golgi protein transport | 3.17 | 3 | 3.08 |
| 9. Hspa5 (78 kDa glucose-regulated protein) | P06761 | 466 | 100 | 332 | 100 | 72,474 | 5.07 | 20 | molecular chaperone for protein folding | 3.25 | 2.88 | 3.06 |
| 10. Fth1 (Ferritin heavy chain) | P19132 | 328 | 100 | 238 | 100 | 21,284 | 5.85 | 10 | iron storage | 3 | 3 | 3 |
| 11. Hba1 (Hemoglobin subunit alpha-1/2) | P01946 | 400 | 100 | 334 | 100 | 15,490 | 7.82 | 7 | oxygen transport | 3 | 3 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sze, S.C.W.; Zhang, L.; Zhang, S.; Lin, K.; Ng, T.B.; Ng, M.L.; Lee, K.-F.; Lam, J.K.W.; Zhang, Z.; Yung, K.K.L. Aberrant Transferrin and Ferritin Upregulation Elicits Iron Accumulation and Oxidative Inflammaging Causing Ferroptosis and Undermines Estradiol Biosynthesis in Aging Rat Ovaries by Upregulating NF-Κb-Activated Inducible Nitric Oxide Synthase: First Demonstration of an Intricate Mechanism. Int. J. Mol. Sci. 2022, 23, 12689. https://doi.org/10.3390/ijms232012689
Sze SCW, Zhang L, Zhang S, Lin K, Ng TB, Ng ML, Lee K-F, Lam JKW, Zhang Z, Yung KKL. Aberrant Transferrin and Ferritin Upregulation Elicits Iron Accumulation and Oxidative Inflammaging Causing Ferroptosis and Undermines Estradiol Biosynthesis in Aging Rat Ovaries by Upregulating NF-Κb-Activated Inducible Nitric Oxide Synthase: First Demonstration of an Intricate Mechanism. International Journal of Molecular Sciences. 2022; 23(20):12689. https://doi.org/10.3390/ijms232012689
Chicago/Turabian StyleSze, Stephen Cho Wing, Liang Zhang, Shiqing Zhang, Kaili Lin, Tzi Bun Ng, Man Ling Ng, Kai-Fai Lee, Jenny Ka Wing Lam, Zhang Zhang, and Ken Kin Lam Yung. 2022. "Aberrant Transferrin and Ferritin Upregulation Elicits Iron Accumulation and Oxidative Inflammaging Causing Ferroptosis and Undermines Estradiol Biosynthesis in Aging Rat Ovaries by Upregulating NF-Κb-Activated Inducible Nitric Oxide Synthase: First Demonstration of an Intricate Mechanism" International Journal of Molecular Sciences 23, no. 20: 12689. https://doi.org/10.3390/ijms232012689
APA StyleSze, S. C. W., Zhang, L., Zhang, S., Lin, K., Ng, T. B., Ng, M. L., Lee, K. -F., Lam, J. K. W., Zhang, Z., & Yung, K. K. L. (2022). Aberrant Transferrin and Ferritin Upregulation Elicits Iron Accumulation and Oxidative Inflammaging Causing Ferroptosis and Undermines Estradiol Biosynthesis in Aging Rat Ovaries by Upregulating NF-Κb-Activated Inducible Nitric Oxide Synthase: First Demonstration of an Intricate Mechanism. International Journal of Molecular Sciences, 23(20), 12689. https://doi.org/10.3390/ijms232012689