

Erythropoiesis and Malaria, a Multifaceted Interplay
{kind=link}
{kind=link}
Abstract
:1. Introduction
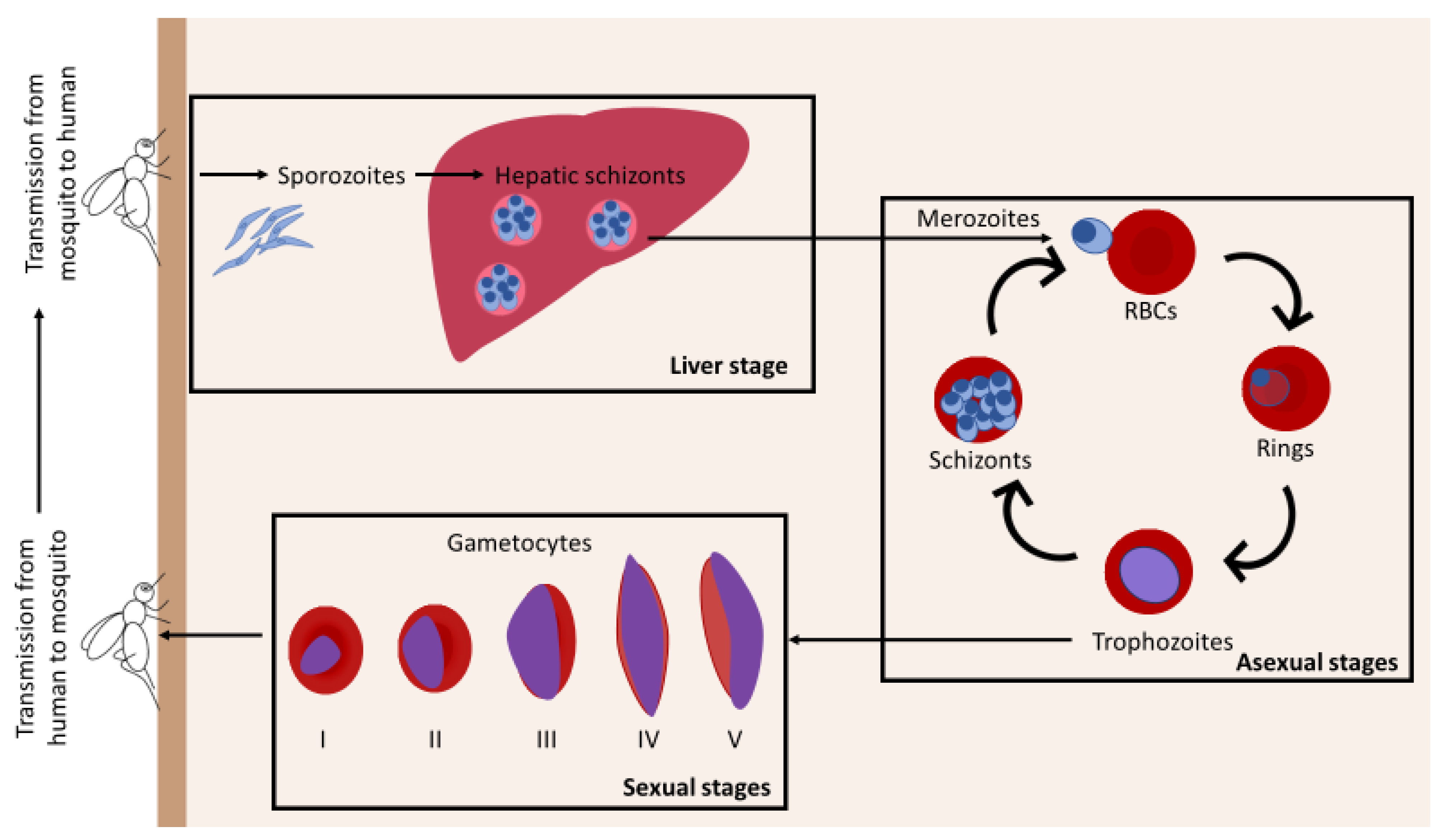
Plasmodium Parasite Sequestration in the Bone Marrow
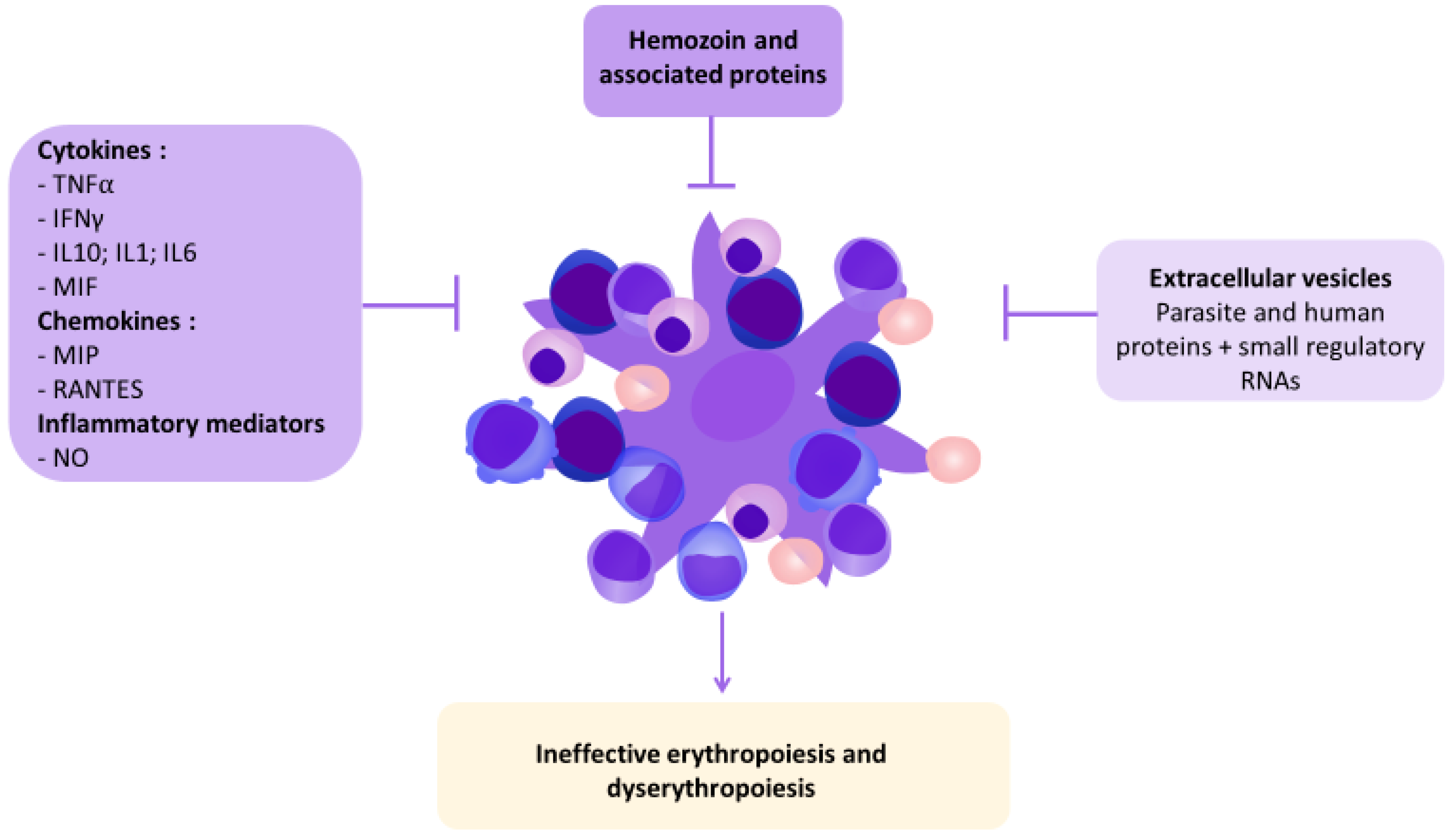
2. Ineffective Erythropoiesis and Dyserythropoiesis in Malaria
3. Human and Parasite Factors Affecting Erythropoiesis
3.1. Extracellular Vesicles
3.2. Hemozoin
3.3. Cytokines
4. Conclusions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Baso-E1 | Early basophilic erythroblast |
| Baso-E2 | Late basophilic erythroblast |
| BFU-E | Burst Forming Unit–Erythroid cells |
| BM | Bone Marrow |
| CFU-E | Colony Forming Unit–Erythroid cells |
| EPO | Erythropoietin |
| EV | Extracellular Vesicle |
| 4-HNE | 4-hydroxynonenal |
| Hz | Hemozoin |
| iEVs | EVs derived from iRBCs |
| IL | Interleukin |
| iRBC | Infected Red Blood Cell |
| IFNγ | Interferon γ |
| HSCs | Hematopoietic Stem Cells |
| HSPCs | Hematopoietic Stem and Progenitor Cells |
| MEP | Megakaryocyte Erythroid Progenitor |
| MIP | Macrophage inflammatory protein |
| MIF | macrophage migration inhibitory factor |
| NO | Nitric Oxide |
| Ortho-E | Orthochromatic erythroblast |
| P. falciparum | Plasmodium falciparum |
| P. vivax | Plasmodium vivax |
| Poly-E | Polychromatophilic erythroblast |
| Pro-E | Proerythroblast |
| RANTES | Regulated on activation, normal T-cell expressed and secreted |
| RBCs | Red Blood Cells |
| ROS | Reactive oxygen species |
| sHz | Synthetic hemozoin |
| TNFα | Tumor Necrosis Factor α |
References
- WHO. WHO Guidelines for Malaria; World Health Organization: Geneva, Switzerland, 2021. Available online: https://www.ncbi.nlm.nih.gov/books/NBK568489/ (accessed on 22 August 2022).
- Slutsker, L.; Taylor, T.E.; Wirima, J.J.; Steketee, R.W. In-hospital morbidity and mortality due to malaria-associated severe anaemia in two areas of Malawi with different patterns of malaria infection. Trans. R. Soc. Trop. Med. Hyg. 1994, 88, 548–555. [Google Scholar] [CrossRef]
- Laveran, C.L. Classics in infectious diseases: A newly discovered parasite in the blood of patients suffering from malaria. Parasitic etiology of attacks of malaria: Charles Louis Alphonse Laveran (1845–1922). Rev. Infect. Dis. 1982, 4, 908–911. [Google Scholar] [CrossRef] [PubMed]
- Marchiafava, E.; Bignami, A. On Summer-Autumn Malarial Fevers; Book on Demand Ltd.: Norderstedt, Germany, 1894. [Google Scholar]
- Garnham, P. Observations on Plasmodium falciparum with Special Reference to the Production of Crescents. Available online: https://www.semanticscholar.org/paper/Observations-on-Plasmodium-falciparum-with-Special-Garnham/5f3ef33ab0e3df1815a280293fc5cd9deae61dbc (accessed on 24 August 2022).
- Smalley, M.; Abdalla, S.; Brown, J. The distribution of Plasmodium falciparum in the peripheral blood and bone marrow of Gambian children. Trans. R. Soc. Trop. Med. Hyg. 1981, 75, 103–105. [Google Scholar] [CrossRef]
- Joice, R.; Nilsson, S.K.; Montgomery, J.; Dankwa, S.; Egan, E.; Morahan, B.; Seydel, K.B.; Bertuccini, L.; Alano, P.; Williamson, K.C.; et al. Plasmodium falciparum transmission stages accumulate in the human bone marrow. Sci. Transl. Med. 2014, 6, 244re5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilar, R.; Magallon-Tejada, A.; Achtman, A.H.; Moraleda, C.; Joice, R.; Cisteró, P.; Li Wai Suen, C.S.; Nhabomba, A.; Macete, E.; Mueller, I.; et al. Molecular evidence for the localization of Plasmodium falciparum immature gametocytes in bone marrow. Blood 2014, 123, 959–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, K.; Liu, J.; Heck, S.; Chasis, J.A.; An, X.; Mohandas, N. Resolving the distinct stages in erythroid differentiation based on dynamic changes in membrane protein expression during erythropoiesis. Proc. Natl. Acad. Sci. USA 2009, 106, 17413–17418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nandakumar, S.; Ulirsch, J.; Sankaran, V.G. Advances in understanding erythropoiesis: Evolving perspectives. Br. J. Haematol. 2016, 173, 206–218. [Google Scholar] [CrossRef] [Green Version]
- David, P.H.; Hommel, M.; Miller, L.H.; Udeinya, I.J.; Oligino, L.D. Parasite sequestration in Plasmodium falciparum malaria: Spleen and antibody modulation of cytoadherence of infected erythrocytes. Proc. Natl. Acad. Sci. USA 1983, 80, 5075–5079. [Google Scholar] [CrossRef] [Green Version]
- Looareesuwan, S.; Ho, M.; Wattanagoon, Y.; White, N.J.; Warrell, D.A.; Bunnag, D.; Harinasuta, T.; Wyler, D.J. Dynamic Alteration in Splenic Function during Acute falciparum Malaria. N. Engl. J. Med. 1987, 317, 675–679. [Google Scholar] [CrossRef]
- Chotivanich, K.; Udomsangpetch, R.; Mcgready, R.; Proux, S.; Newton, P.; Pukrittayakamee, S.; Looareesuwan, S.; White, N.J. Central Role of the Spleen in Malaria Parasite Clearance. J. Infect. Dis. 2002, 185, 1538–1541. [Google Scholar] [CrossRef]
- Buffet, A.P.; Safeukui, I.; Milon, G.; Mercereau-Puijalon, O.; David, P.H. Retention of erythrocytes in the spleen: A double-edged process in human malaria. Curr. Opin. Hematol. 2009, 16, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Henry, B.; Roussel, C.; Carucci, M.; Brousse, V.; Ndour, P.A.; Buffet, P. The Human Spleen in Malaria: Filter or Shelter? Trends Parasitol. 2020, 36, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Pathak, V.A.; Ghosh, K. Erythropoiesis in Malaria Infections and Factors Modifying the Erythropoietic Response. Anemia 2016, 2016, 9310905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, X.-Z.; Heatwole, V.M.; Wertheimer, S.P.; Guinet, F.; Herrfeldt, J.A.; Peterson, D.S.; Ravetch, J.A.; Wellems, T.E. The large diverse gene family var encodes proteins involved in cytoadherence and antigenic variation of plasmodium falciparum-infected erythrocytes. Cell 1995, 82, 89–100. [Google Scholar] [CrossRef] [Green Version]
- Turner, L.; Lavstsen, T.; Berger, S.S.; Wang, C.W.; Petersen, J.E.V.; Avril, M.; Brazier, A.J.; Freeth, J.; Jespersen, J.S.; Nielsen, M.A.; et al. Severe malaria is associated with parasite binding to endothelial protein C receptor. Nature 2013, 498, 502–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salanti, A.; Dahlbäck, M.; Turner, L.; Nielsen, M.A.; Barfod, L.; Magistrado, P.; Jensen, A.T.R.; Lavstsen, T.; Ofori, M.F.; Marsh, K.; et al. Evidence for the Involvement of VAR2CSA in Pregnancy-associated Malaria. J. Exp. Med. 2004, 200, 1197–1203. [Google Scholar] [CrossRef] [PubMed]
- Meibalan, E.; Marti, M. Biology of Malaria Transmission. Cold Spring Harb. Perspect. Med. 2017, 7, a025452. [Google Scholar] [CrossRef] [Green Version]
- Neveu, G.; Richard, C.; Dupuy, F.; Behera, P.K.; Volpe, F.; Subramani, P.A.; Marcel-Zerrougui, B.; Vallin, P.; Andrieu, M.; Minz, A.M.; et al. Plasmodium falciparum sexual parasites develop in human erythroblasts and affect erythropoiesis. Blood 2020, 136, 1381–1393. [Google Scholar] [CrossRef]
- Baro, B.; Deroost, K.; Raiol, T.; Brito, M.; Almeida, A.C.G.; Menezes-Neto, A.; Figueiredo, E.F.G.; Alencar, A.; Leitão, R.; Val, F.; et al. Plasmodium vivax gametocytes in the bone marrow of an acute malaria patient and changes in the erythroid miRNA profile. PLOS Negl. Trop. Dis. 2017, 11, e0005365. [Google Scholar] [CrossRef] [Green Version]
- Brito, M.A.M.; Baro, B.; Raiol, T.C.; Ayllon-Hermida, A.; Safe, I.P.; Deroost, K.; Figueiredo, E.F.G.; Costa, A.G.; Armengol, M.D.P.; Sumoy, L.; et al. Morphological and Transcriptional Changes in Human Bone Marrow During Natural Plasmodium vivax Malaria Infections. J. Infect. Dis. 2020, 225, 1274–1283. [Google Scholar] [CrossRef]
- Malleret, B.; Li, A.; Zhang, R.; Tan, K.S.W.; Suwanarusk, R.; Claser, C.; Cho, J.S.; Koh, E.G.L.; Chu, C.S.; Pukrittayakamee, S.; et al. Plasmodium vivax: Restricted tropism and rapid remodeling of CD71-positive reticulocytes. Blood 2015, 125, 1314–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kho, S.; Qotrunnada, L.; Leonardo, L.; Andries, B.; Wardani, P.A.I.; Fricot, A.; Henry, B.; Hardy, D.; Margyaningsih, N.I.; Apriyanti, D.; et al. Evaluation of splenic accumulation and colocalization of immature reticulocytes and Plasmodium vivax in asymptomatic malaria: A prospective human splenectomy study. PLOS Med. 2021, 18, e1003632. [Google Scholar] [CrossRef] [PubMed]
- Kho, S.; Qotrunnada, L.; Leonardo, L.; Andries, B.; Wardani, P.A.; Fricot, A.; Henry, B.; Hardy, D.; Margyaningsih, N.I.; Apriyanti, D.; et al. Hidden Biomass of Intact Malaria Parasites in the Human Spleen. N. Engl. J. Med. 2021, 384, 2067–2069. [Google Scholar] [CrossRef] [PubMed]
- De Niz, M.; Meibalan, E.; Mejia, P.; Ma, S.; Brancucci, N.M.B.; Agop-Nersesian, C.; Mandt, R.; Ngotho, P.; Hughes, K.R.; Waters, A.P.; et al. Plasmodium gametocytes display homing and vascular transmigration in the host bone marrow. Sci. Adv. 2018, 4, eaat3775. [Google Scholar] [CrossRef] [Green Version]
- Duffier, Y.; Lorthiois, A.; Cisteró, P.; Dupuy, F.; Jouvion, G.; Fiette, L.; Mazier, M.; Mayor, A.; Lavazec, C.; Sabater, A.M. A humanized mouse model for sequestration of Plasmodium falciparum sexual stages and in vivo evaluation of gametocytidal drugs. Sci. Rep. 2016, 6, 35025. [Google Scholar] [CrossRef] [Green Version]
- Obaldia, N.; Meibalan, E.; Sa, J.M.; Ma, S.; Clark, M.A.; Mejia, P.; Barros, R.R.M.; Otero, W.; Ferreira, M.U.; Mitchell, J.R.; et al. Bone Marrow Is a Major Parasite Reservoir in Plasmodium vivax Infection. mBio 2018, 9, e00625-18. [Google Scholar] [CrossRef] [Green Version]
- Farfour, E.; Charlotte, F.; Settegrana, C.; Miyara, M.; Buffet, P. The extravascular compartment of the bone marrow: A niche for Plasmodium falciparum gametocyte maturation? Malar. J. 2012, 11, 285. [Google Scholar] [CrossRef] [Green Version]
- Neveu, G.; Dupuy, F.; Ladli, M.; Barbieri, D.; Naissant, B.; Richard, C.; Martins, R.M.; Lopez-Rubio, J.-J.; Bachmann, A.; Verdier, F.; et al. Plasmodium falciparum gametocyte-infected erythrocytes do not adhere to human primary erythroblasts. Sci. Rep. 2018, 8, 17886. [Google Scholar] [CrossRef] [Green Version]
- Panichakul, T.; Sattabongkot, J.; Chotivanich, K.; Sirichaisinthop, J.; Cui, L.; Udomsangpetch, R. Production of erythropoietic cells in vitro for continuous culture of Plasmodium vivax. Int. J. Parasitol. 2007, 37, 1551–1555. [Google Scholar] [CrossRef]
- Tamez, P.A.; Liu, H.; Fernandez-Pol, S.; Haldar, K.; Wickrema, A. Stage-specific susceptibility of human erythroblasts to Plasmodium falciparum malaria infection. Blood 2009, 114, 3652–3655. [Google Scholar] [CrossRef]
- Peatey, C.L.; Watson, J.A.; Trenholme, K.R.; Brown, C.L.; Nielson, L.; Guenther, M.; Timmins, N.; Watson, G.S.; Gardiner, D.L. Enhanced Gametocyte Formation in Erythrocyte Progenitor Cells: A Site-Specific Adaptation by Plasmodium falciparum. J. Infect. Dis. 2013, 208, 1170–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furuya, T.; Sá, J.M.; Chitnis, C.E.; Wellems, T.E.; Stedman, T.T. Reticulocytes from cryopreserved erythroblasts support Plasmodium vivax infection in vitro. Parasitol. Int. 2014, 63, 278–284. [Google Scholar] [CrossRef] [Green Version]
- Scherf, A.; Carter, R.; Petersen, C.; Alano, P.; Nelson, R.; Aikawa, M.; Mattei, D.; Da Silva, L.P.; Leech, J. Gene inactivation of Pf11-1 of Plasmodium falciparum by chromosome breakage and healing: Identification of a gametocyte-specific protein with a potential role in gametogenesis. EMBO J. 1992, 11, 2293–2301. [Google Scholar] [CrossRef]
- Lee, R.S.; Waters, A.P.; Brewer, J.M. A cryptic cycle in haematopoietic niches promotes initiation of malaria transmission and evasion of chemotherapy. Nat. Commun. 2018, 9, 1689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hentzschel, F.; Gibbins, M.P.; Attipa, C.; Beraldi, D.; Moxon, C.A.; Otto, T.D.; Marti, M. Host cell maturation modulates parasite invasion and sexual differentiation in Plasmodium berghei. Sci. Adv. 2022, 8, abm7348. [Google Scholar] [CrossRef] [PubMed]
- Messina, V.; Valtieri, M.; Rubio, M.; Falchi, M.; Mancini, F.; Mayor, A.; Alano, P.; Silvestrini, F. Gametocytes of the Malaria Parasite Plasmodium falciparum Interact with and Stimulate Bone Marrow Mesenchymal Cells to Secrete Angiogenetic Factors. Front. Cell. Infect. Microbiol. 2018, 8, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aingaran, M.; Zhang, R.; Law, S.K.; Peng, Z.; Undisz, A.; Meyer, E.; Diez-Silva, M.; Burke, T.A.; Spielmann, T.; Lim, C.T.; et al. Host cell deformability is linked to transmission in the human malaria parasite Plasmodium falciparum. Cell Microbiol. 2012, 14, 983–993. [Google Scholar] [CrossRef] [Green Version]
- Dearnley, M.; Chu, T.; Zhang, Y.; Looker, O.; Huang, C.; Klonis, N.; Yeoman, J.; Kenny, S.; Arora, M.; Osborne, J.M.; et al. Reversible host cell remodeling underpins deformability changes in malaria parasite sexual blood stages. Proc. Natl. Acad. Sci. USA 2016, 113, 4800–4805. [Google Scholar] [CrossRef] [Green Version]
- Tibúrcio, M.; Niang, M.; Deplaine, G.; Perrot, S.; Bischoff, E.; Ndour, P.A.; Silvestrini, F.; Khattab, A.; Milon, G.; David, P.H.; et al. A switch in infected erythrocyte deformability at the maturation and blood circulation of Plasmodium falciparum transmission stages. Blood 2012, 119, e172–e180. [Google Scholar] [CrossRef]
- Maggio-Price, L.; Brookoff, D.; Weiss, L. Changes in hematopoietic stem cells in bone marrow of mice with Plasmodium berghei malaria. Blood 1985, 66, 1080–1085. [Google Scholar] [CrossRef]
- Jootar, S.; Chaisiripoomkere, W.; Pholvicha, P.; Leelasiri, A.; Prayoonwiwat, W.; Mongkonsvitragoon, W.; Srichaikul, T. Suppression of erythroid progenitor cells during malarial infection in Thai adults caused by serum inhibitor. Clin. Lab. Haematol. 1993, 15, 87–92. [Google Scholar] [CrossRef]
- Srichaikul, T.; Panikbutr, N.; Jeumtrakul, P. Bone-marrow changes in human malaria. Ann. Trop. Med. Parasitol. 1967, 61, 40–51. [Google Scholar] [CrossRef]
- Srichaikul, T.; Siriasawakul, T.; Poshyachinda, M.; Poshyachinda, V. Ferrokinetics in Patients with Malaria: Normoblasts and Iron Incorporation in Vitro. Am. J. Clin. Pathol. 1973, 60, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Dormer, P.; Dietrich, M.; Kern, P.; Horstmann, R.D. Ineffective erythropoiesis in acute human P. falciparum malaria. Blut 1983, 46, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.-H.; Tam, M.; Stevenson, M.M. Inappropriately low reticulocytosis in severe malarial anemia correlates with suppression in the development of late erythroid precursors. Blood 2004, 103, 3727–3735. [Google Scholar] [CrossRef]
- Hendrickse, R.G.; King, M.A.R. Anaemia of Uncertain Origin in Infancy. BMJ 1958, 2, 662–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickramasinghe, S.N.; Looareesuwan, S.; Nagachinta, B.; White, N.J. Dyserythropoiesis and ineffective erythropoiesis in Plasmodium vivax malaria. Br. J. Haematol. 1989, 72, 91–99. [Google Scholar] [CrossRef]
- Zuckerman, A. Blood Loss and Replacement in Plasmodial Infections I. Plasmodium Berghei in Untreated Rats of Varying Age and in Adult Rats with Erythropoietic Mechanisms Manipulated before Inoculation. J. Infect. Dis. 1957, 100, 172–206. [Google Scholar] [CrossRef]
- Rencricca, N.J.; Stout, J.P.; Coleman, R.M. Erythropoietin Production in Virulent Malaria. Infect. Immun. 1974, 10, 831–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thawani, N.; Tam, M.; Bellemare, M.J.; Bohle, D.S.; Olivier, M.; de Souza, J.B.; Stevenson, M.M. Plasmodium products contribute to severe malarial anemia by inhibiting erythropoietin-induced proliferation of erythroid precursors. J. Infect. Dis. 2014, 209, 140–149. [Google Scholar] [CrossRef]
- Egan, A.F.; Fabucci, M.E.; Saul, A.; Kaslow, D.C.; Miller, L.H. Aotus New World monkeys: Model for studying malaria-induced anemia. Blood 2002, 99, 3863–3866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panichakul, T.; Payuhakrit, W.; Panburana, P.; Wongborisuth, C.; Hongeng, S.; Udomsangpetch, R. Suppression of erythroid development in vitro by Plasmodium vivax. Malar. J. 2012, 11, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickramasinghe, S.N.; Abdalla, S.; Weatherall, D.J. Cell Cycle Distribution of Erythroblasts in P. falciparum Malaria. Scand. J. Haematol. 1982, 29, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Kurtzhals, J.A.; Rodrigues, O.; Addae, M.; Commey, J.O.; Nkrumah, F.K.; Hviid, L. Reversible suppression of bone marrow response to erythropoietin in Plasmodium falciparum malaria. Br. J. Haematol. 1997, 97, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Lakkavaram, A.; Lundie, R.J.; Do, H.; Ward, A.C.; De Koning-Ward, T.F. Acute Plasmodium berghei Mouse Infection Elicits Perturbed Erythropoiesis with Features That Overlap with Anemia of Chronic Disease. Front. Microbiol. 2020, 11, 702. [Google Scholar] [CrossRef] [Green Version]
- Abdalla, S.; Weatherall, D.J.; Wickramasinghe, S.N.; Hughes, M. The anaemia of P. falciparum malaria. Br. J. Haematol. 1980, 46, 171–183. [Google Scholar] [CrossRef]
- Abdalla, S.; Wickramasinghe, S.; Weatherall, D. The deoxyuridine suppression test in severe anaemia following Plasmodium falciparum malaria. Trans. R. Soc. Trop. Med. Hyg. 1984, 78, 60–63. [Google Scholar] [CrossRef]
- Phillips, R.; Warrell, D. The pathophysiology of severe falciparum malaria. Parasitol. Today 1986, 2, 271–282. [Google Scholar] [CrossRef]
- Sexton, A.C.; Good, R.T.; Hansen, D.S.; D’Ombrain, M.C.; Buckingham, L.; Simpson, K.; Schofield, L. Transcriptional Profiling Reveals Suppressed Erythropoiesis, Up-Regulated Glycolysis, and Interferon-Associated Responses in Murine Malaria. J. Infect. Dis. 2004, 189, 1245–1256. [Google Scholar] [CrossRef]
- Hattangadi, S.M.; Wong, P.; Zhang, L.; Flygare, J.; Lodish, H.F. From stem cell to red cell: Regulation of erythropoiesis at multiple levels by multiple proteins, RNAs, and chromatin modifications. Blood 2011, 118, 6258–6268. [Google Scholar] [CrossRef]
- Tamez, P.A.; Liu, H.; Wickrema, A.; Haldar, K.P. Falciparum Modulates Erythroblast Cell Gene Expression in Signaling and Erythrocyte Production Pathways. PLoS ONE 2011, 6, e19307. [Google Scholar] [CrossRef] [PubMed]
- Meldolesi, J. Exosomes and Ectosomes in Intercellular Communication. Curr. Biol. 2018, 28, R435–R444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robbins, P.D.; Morelli, A.E. Regulation of immune responses by extracellular vesicles. Nat. Rev. Immunol. 2014, 14, 195–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rank, A.; Nieuwland, R.; Köhler, A.; Franz, C.; Waidhauser, J.; Toth, B. Human bone marrow contains high levels of extracellular vesicles with a tissue-specific subtype distribution. PLoS ONE 2018, 13, e020795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidal, M. Exosomes in erythropoiesis. Transfus. Clin. Biol. 2010, 17, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Yáñez-Mó, M.; Siljander, P.R.-M.; Andreu, Z.; Bedina Zavec, A.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [Green Version]
- Doyle, L.; Wang, M. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef] [Green Version]
- Coakley, G.; Maizels, R.M.; Buck, A.H. Exosomes and Other Extracellular Vesicles: The New Communicators in Parasite Infections. Trends Parasitol. 2015, 31, 477–489. [Google Scholar] [CrossRef] [Green Version]
- Campos, F.M.; Franklin, B.S.; Teixeira-Carvalho, A.; Filho, A.L.; de Paula, S.C.; Fontes, C.J.; Brito, C.F.; Carvalho, L.H. Augmented plasma microparticles during acute Plasmodium vivax infection. Malar. J. 2010, 9, 327. [Google Scholar] [CrossRef] [Green Version]
- Nantakomol, D.; Dondorp, A.M.; Krudsood, S.; Udomsangpetch, R.; Pattanapanyasat, K.; Combes, V.; Grau, G.E.; White, N.J.; Viriyavejakul, P.; Day, N.P.; et al. Circulating Red Cell–derived Microparticles in Human Malaria. J. Infect. Dis. 2011, 203, 700–706. [Google Scholar] [CrossRef]
- El-Assaad, F.; Wheway, J.; Hunt, N.H.; Grau, G.; Combes, V. Production, Fate and Pathogenicity of Plasma Microparticles in Murine Cerebral Malaria. PLoS Pathog. 2014, 10, e1003839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Sousa, K.P.; Potriquet, J.; Mulvenna, J.; Sotillo, J.; Groves, P.L.; Loukas, A.; Apte, S.H.; Doolan, D.L. Proteomic identification of the contents of small extracellular vesicles from in vivo Plasmodium yoelii infection. Int. J. Parasitol. 2021, 52, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Demarta-Gatsi, C.; Rivkin, A.; Di Bartolo, V.; Peronet, R.; Ding, S.; Commere, P.; Guillonneau, F.; Bellalou, J.; Brûlé, S.; Karam, P.A.; et al. Histamine releasing factor and elongation factor 1 alpha secreted via malaria parasites extracellular vesicles promote immune evasion by inhibiting specific T cell responses. Cell. Microbiol. 2019, 21, e13021. [Google Scholar] [CrossRef]
- Mantel, P.Y.; Hoang, A.N.; Goldowitz, I.; Potashnikova, D.; Hamza, B.; Vorobjev, I.; Ghiran, I.; Toner, M.; Irimia, D.; Ivanov, A.R.; et al. Malaria-Infected Erythrocyte-Derived Microvesicles Mediate Cellular Communication within the Parasite Population and with the Host Immune System. Cell Host Microbe 2013, 13, 521–534. [Google Scholar] [CrossRef] [Green Version]
- Mantel, P.-Y.; Hjelmqvist, D.; Walch, M.; Kharoubi-Hess, S.; Nilsson, S.; Ravel, D.; Ribeiro, M.; Grüring, C.; Ma, S.; Padmanabhan, P.; et al. Infected erythrocyte-derived extracellular vesicles alter vascular function via regulatory Ago2-miRNA complexes in malaria. Nat. Commun. 2016, 7, 12727. [Google Scholar] [CrossRef] [Green Version]
- Regev-Rudzki, N.; Wilson, D.W.; Carvalho, T.G.; Sisquella, X.; Coleman, B.M.; Rug, M.; Bursac, D.; Angrisano, F.; Gee, M.; Hill, A.F.; et al. Cell-Cell Communication between Malaria-Infected Red Blood Cells via Exosome-like Vesicles. Cell 2013, 153, 1120–1133. [Google Scholar] [CrossRef] [Green Version]
- Dekel, E.; Karam, P.A.; Ohana-Daniel, Y.; Biton, M.; Regev-Rudzki, N.; Porat, Z. Antibody-Free Labeling of Malaria-Derived Extracellular Vesicles Using Flow Cytometry. Biomedicines 2020, 8, 98. [Google Scholar] [CrossRef]
- Opadokun, T.; Rohrbach, P. Extracellular vesicles in malaria: An agglomeration of two decades of research. Malar. J. 2021, 20, 442. [Google Scholar] [CrossRef]
- Correa, R.; Coronado, L.; Caballero, Z.; Faral-Tello, P.; Robello, C.; Spadafora, C. Extracellular vesicles carrying lactate dehydrogenase induce suicide in increased population density of Plasmodium falciparum in vitro. Sci. Rep. 2019, 9, 5042. [Google Scholar] [CrossRef]
- Karam, P.A.; Rosenhek-Goldian, I.; Ziv, T.; Pilo, H.B.A.; Azuri, I.; Rivkin, A.; Kiper, E.; Rotkopf, R.; Cohen, S.R.; Torrecilhas, A.C.; et al. Malaria parasites release vesicle subpopulations with signatures of different destinations. EMBO Rep. 2022, 23, e54755. [Google Scholar] [CrossRef] [PubMed]
- Antwi-Baffour, S.; Malibha-Pinchbeck, M.; Stratton, D.; Jorfi, S.; Lange, S.; Inal, J. Plasma mEV levels in Ghanain malaria patients with low parasitaemia are higher than those of healthy controls, raising the potential for parasite markers in mEVs as diagnostic targets. J. Extracell. Vesicles 2020, 9, 1697124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaular, L.M.; Nakayasu, E.; Ferrer, M.; Almeida, I.C.; Del Portillo, H.A. Exosomes from Plasmodium yoelii-Infected Reticulocytes Protect Mice from Lethal Infections. PLoS ONE 2011, 6, e26588. [Google Scholar]
- Couper, K.N.; Barnes, T.; Hafalla, J.C.; Combes, V.; Ryffel, B.; Secher, T.; Grau, G.E.; Riley, E.M.; de Souza, J.B. Parasite-derived plasma microparticles contribute significantly to malaria infection-induced inflammation through potent macrophage stimulation. PLoS Pathog. 2010, 6, e1000744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, W.; Chew, M.; Hou, J.; Lai, F.; Leopold, S.J.; Loo, H.L.; Ghose, A.; Dutta, A.K.; Chen, Q.; Ooi, E.E.; et al. Microvesicles from malaria-infected red blood cells activate natural killer cells via MDA5 pathway. PLOS Pathog. 2018, 14, e1007298. [Google Scholar] [CrossRef] [Green Version]
- Carrera-Bravo, C.; Koh, E.Y.; Tan, K.S.W. The roles of parasite-derived extracellular vesicles in disease and host-parasite communication. Parasitol. Int. 2021, 83, 102373. [Google Scholar] [CrossRef]
- Sisquella, X.; Ofir-Birin, Y.; Pimentel, M.A.; Cheng, L.; Karam, P.A.; Sampaio, N.G.; Penington, J.S.; Connolly, D.; Giladi, T.; Scicluna, B.J.; et al. Malaria parasite DNA-harbouring vesicles activate cytosolic immune sensors. Nat. Commun. 2017, 8, 1985. [Google Scholar] [CrossRef] [Green Version]
- Babatunde, K.A.; Mbagwu, S.; Hernández-Castañeda, M.A.; Adapa, S.R.; Walch, M.; Filgueira, L.; Falquet, L.; Jiang, R.H.Y.; Ghiran, I.; Mantel, P.-Y. Malaria infected red blood cells release small regulatory RNAs through extracellular vesicles. Sci. Rep. 2018, 8, 884. [Google Scholar] [CrossRef] [Green Version]
- Ketprasit, N.; Cheng, I.S.; Deutsch, F.; Tran, N.; Imwong, M.; Combes, V.; Palasuwan, D. The characterization of extracellular vesicles-derived microRNAs in Thai malaria patients. Malar. J. 2020, 19, 285. [Google Scholar] [CrossRef]
- Xue, X.; Zhang, Q.; Huang, Y.; Feng, L.; Pan, W. No miRNA were found in Plasmodium and the ones identified in erythrocytes could not be correlated with infection. Malar. J. 2008, 7, 47. [Google Scholar] [CrossRef] [Green Version]
- Baum, J.; Papenfuss, A.; Mair, G.; Janse, C.J.; Vlachou, D.; Waters, A.; Cowman, A.F.; Crabb, B.S.; De Koning-Ward, T.F. Molecular genetics and comparative genomics reveal RNAi is not functional in malaria parasites. Nucleic Acids Res. 2009, 37, 3788–3798. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Yu, Y.; Niu, B.; Wang, D. Red Blood Cells as Potential Repositories of MicroRNAs in the Circulatory System. Front. Genet. 2020, 11, 442. [Google Scholar] [CrossRef] [PubMed]
- Doss, J.F.; Corcoran, D.L.; Jima, D.D.; Telen, M.J.; Dave, S.S.; Chi, J.-T. A comprehensive joint analysis of the long and short RNA transcriptomes of human erythrocytes. BMC Genom. 2015, 16, 952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patrick, D.M.; Zhang, C.C.; Tao, Y.; Yao, H.; Qi, X.; Schwartz, R.J.; Huang, L.J.-S.; Olson, E.N. Defective erythroid differentiation in miR-451 mutant mice mediated by 14-3-3ζ. Genes Dev. 2010, 24, 1614–1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.-S.; Li, L.; Chu, S.; Shiang, K.-D.; Li, M.; Sun, H.-Y.; Xu, J.; Xiao, F.-J.; Sun, G.; Rossi, J.J.; et al. MicroRNA-486 regulates normal erythropoiesis and enhances growth and modulates drug response in CML progenitors. Blood 2015, 125, 1302–1313. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Xi, J.; Hao, X.; Deng, W.; Liu, J.; Wei, C.; Gao, Y.; Zhang, L.; Wang, H. Red blood cells release microparticles containing human argonaute 2 and miRNAs to target genes of Plasmodium falciparum. Emerg. Microbes Infect. 2017, 6, e75. [Google Scholar] [CrossRef] [Green Version]
- Xu, P.; Palmer, L.E.; Lechauve, C.; Zhao, G.; Yao, Y.; Luan, J.; Vourekas, A.; Tan, H.; Peng, J.; Schuetz, J.D.; et al. Regulation of gene expression by miR-144/451 during mouse erythropoiesis. Blood 2019, 133, 2518–2528. [Google Scholar] [CrossRef]
- Sangokoya, C.; Telen, M.J.; Chi, J.-T. microRNA miR-144 modulates oxidative stress tolerance and associates with anemia severity in sickle cell disease. Blood 2010, 116, 4338–4348. [Google Scholar] [CrossRef] [Green Version]
- Walzer, K.A.; Chi, J.-T. Trans-kingdom small RNA transfer during host-pathogen interactions: The case of P. falciparum and erythrocytes. RNA Biol. 2017, 14, 442–449. [Google Scholar] [CrossRef] [Green Version]
- Kataria, P.; Surela, N.; Chaudhary, A.; Das, J. MiRNA: Biological Regulator in Host-Parasite Interaction during Malaria Infection. Int. J. Environ. Res. Public Health 2022, 19, 2395. [Google Scholar] [CrossRef]
- Pagola, S.; Stephens, P.W.; Bohle, D.S.; Kosar, A.D.; Madsen, S.K. The structure of malaria pigment beta-haematin. Nature 2000, 404, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Francis, S.E.; Sullivan, D.J.; Goldberg, A.D.E. Hemoglobin Metabolism in the Malaria Parasite Plasmodium falciparum. Annu. Rev. Microbiol. 1997, 51, 97–112. [Google Scholar] [CrossRef] [PubMed]
- Papalexis, V.; Siomos, M.-A.; Campanale, N.; Guo, X.-G.; Kocak, G.; Foley, M.; Tilley, L. Histidine-rich protein 2 of the malaria parasite, Plasmodium falciparum, is involved in detoxification of the by-products of haemoglobin degradation. Mol. Biochem. Parasitol. 2001, 115, 77–86. [Google Scholar] [CrossRef]
- Shio, M.T.; Kassa, F.A.; Bellemare, M.-J.; Olivier, M. Innate inflammatory response to the malarial pigment hemozoin. Microbes Infect. 2010, 12, 889–899. [Google Scholar] [CrossRef]
- Casals-Pascual, C.; Kai, O.; Cheung, J.O.P.; Williams, S.; Lowe, B.; Nyanoti, M.; Williams, T.N.; Maitland, K.; Molyneux, M.; Newton, C.; et al. Suppression of erythropoiesis in malarial anemia is associated with hemozoin in vitro and in vivo. Blood 2006, 108, 2569–2577. [Google Scholar] [CrossRef] [Green Version]
- Griffith, J.W.; Sun, T.; McIntosh, M.T.; Bucala, R. Pure Hemozoin Is Inflammatory In Vivo and Activates the NALP3 Inflammasome via Release of Uric Acid. J. Immunol. 2009, 183, 5208–5220. [Google Scholar] [CrossRef] [Green Version]
- Jaramillo, M.; Plante, I.; Ouellet, N.; Vandal, K.; Tessier, P.A.; Olivier, M. Hemozoin-Inducible Proinflammatory Events In Vivo: Potential Role in Malaria Infection. J. Immunol. 2004, 172, 3101–3110. [Google Scholar] [CrossRef] [Green Version]
- Giribaldi, G.; Ulliers, D.; Schwarzer, E.; Roberts, I.; Piacibello, W.; Arese, P. Hemozoin- and 4-hydroxynonenal-mediated inhibition of erythropoiesis. Possible role in malarial dyserythropoiesis and anemia. Haematologica 2004, 89, 492–493. [Google Scholar]
- Skorokhod, O.; Caione, L.; Marrocco, T.; Migliardi, G.; Barrera, V.; Arese, P.; Piacibello, W.; Schwarzer, E. Inhibition of erythropoiesis in malaria anemia: Role of hemozoin and hemozoin-generated 4-hydroxynonenal. Blood 2010, 116, 4328–4337. [Google Scholar] [CrossRef] [Green Version]
- Lamikanra, A.A.; Theron, M.; Kooij, T.W.A.; Roberts, D.J. Hemozoin (Malarial Pigment) Directly Promotes Apoptosis of Erythroid Precursors. PLoS ONE 2009, 4, e8446. [Google Scholar] [CrossRef] [Green Version]
- Schwarzer, E.; Kühn, H.; Valente, E.; Arese, P. Malaria-parasitized erythrocytes and hemozoin nonenzymatically generate large amounts of hydroxy fatty acids that inhibit monocyte functions. Blood 2003, 101, 722–728. [Google Scholar] [CrossRef] [Green Version]
- Poli, G.; Schaur, R.J.; Siems, W.G.; Leonarduzzi, G.M. 4-Hydroxynonenal: A membrane lipid oxidation product of medicinal interest. Med. Res. Rev. 2008, 28, 569–631. [Google Scholar] [CrossRef]
- Boehm, D.; Healy, L.; Ring, S.; Bell, A. Inhibition of ex vivo erythropoiesis by secreted and haemozoin-associated Plasmodium falciparum products. Parasitology 2018, 145, 1865–1875. [Google Scholar] [CrossRef] [PubMed]
- Lamikanra, A.A.; Merryweather-Clarke, A.T.; Tipping, A.J.; Roberts, D.J. Distinct Mechanisms of Inadequate Erythropoiesis Induced by Tumor Necrosis Factor Alpha or Malarial Pigment. PLoS ONE 2015, 10, e0119836. [Google Scholar]
- Ochiel, D.O.; Awandare, G.A.; Keller, C.C.; Hittner, J.B.; Kremsner, P.G.; Weinberg, J.B.; Perkins, D.J. Differential Regulation of β-Chemokines in Children with Plasmodium falciparum Malaria. Infect. Immun. 2005, 73, 4190–4197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, C.C.; Kremsner, P.G.; Hittner, J.B.; Misukonis, M.A.; Weinberg, J.B.; Perkins, D.J. Elevated Nitric Oxide Production in Children with Malarial Anemia: Hemozoin-Induced Nitric Oxide Synthase Type 2 Transcripts and Nitric Oxide in Blood Mononuclear Cells. Infect. Immun. 2004, 72, 4868–4873. [Google Scholar] [CrossRef] [Green Version]
- Becker, K.; Tilley, L.; Vennerstrom, J.L.; Roberts, D.; Rogerson, S.; Ginsburg, H. Oxidative stress in malaria parasite-infected erythrocytes: Host–parasite interactions. Int. J. Parasitol. 2004, 34, 163–189. [Google Scholar] [CrossRef] [PubMed]
- Carney, C.K.; Schrimpe, A.C.; Halfpenny, K.; Harry, R.S.; Miller, C.M.; Broncel, M.; Sewell, S.L.; Schaff, J.E.; Deol, R.; Carter, M.D.; et al. The basis of the immunomodulatory activity of malaria pigment (hemozoin). J. Biol. Inorg. Chem. 2006, 11, 917–929. [Google Scholar] [CrossRef]
- Morceau, F.; Dicato, M.; Diederich, M. Pro-Inflammatory Cytokine-Mediated Anemia: Regarding Molecular Mechanisms of Erythropoiesis. Mediat. Inflamm. 2009, 2009, 405016. [Google Scholar] [CrossRef] [Green Version]
- Roodman, G.D. Mechanisms of erythroid suppression in the anemia of chronic disease. Blood Cells 1987, 13, 171–184. [Google Scholar]
- Felli, N.; Pedini, F.; Zeuner, A.; Petrucci, E.; Testa, U.; Conticello, C.; Biffoni, M.; Di Cataldo, A.; Winkles, J.A.; Peschle, C.; et al. Multiple members of the TNF superfamily contribute to IFN-gamma-mediated inhibition of erythropoiesis. J. Immunol. 2005, 175, 1464–1472. [Google Scholar] [CrossRef] [Green Version]
- Clark, I.A.; Chaudhri, G. Tumour necrosis factor may contribute to the anaemia of malaria by causing dyserythropoiesis and erythrophagocytosis. Br. J. Haematol. 1988, 70, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Torre, D.; Speranza, F.; Giola, M.; Matteelli, A.; Tambini, R.; Biondi, G. Role of Th1 and Th2 cytokines in immune response to uncomplicated Plasmodium falciparum malaria. Clin. Diagn. Lab. Immunol. 2002, 9, 348–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurtzhals, J.; Adabayeri, V.; Goka, B.Q.; Akanmori, B.D.; Oliver-Commey, O.J.; Nkrumah, F.K.; Behr, C.; Hviid, L. Low plasma concentrations of interleukin 10 in severe malarial anaemia compared with cerebral and uncomplicated malaria. Lancet 1998, 351, 1768–1772. [Google Scholar] [CrossRef]
- Othoro, C.; Lal, A.A.; Nahlen, B.; Koech, D.; Orago, A.S.S.; Udhayakumar, V. A Low Interleukin-10 Tumor Necrosis Factor-α Ratio Is Associated with Malaria Anemia in Children Residing in a Holoendemic Malaria Region in Western Kenya. J. Infect. Dis. 1999, 179, 279–282. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Q.; Udupa, K.B.; Lipschitz, D.A. Evidence suggesting a stimulatory role for interleukin-10 in erythropoiesis in vitro. J. Cell. Physiol. 1996, 166, 305–310. [Google Scholar] [CrossRef]
- Dybedal, I.; Larsen, S.; Jacobsen, S.E.W. IL-12 directly enhances in vitro murine erythropoiesis in combination with IL-4 and stem cell factor. J. Immunol. 1995, 154, 4950–4955. [Google Scholar]
- Mohan, K.; Stevenson, M.M. Interleukin-12 corrects severe anemia during blood-stage Plasmodium chabaudi AS in susceptible A/J mice. Exp. Hematol. 1998, 26, 45–52. [Google Scholar]
- Zhang, L.; Prather, D.; Eng, J.V.; Crawford, S.; Kariuki, S.; Ter Kuile, F.; Terlouw, D.; Nahlen, B.; Lal, A.A.; Slutsker, L.; et al. Polymorphisms in genes of interleukin 12 and its receptors and their association with protection against severe malarial anaemia in children in western Kenya. Malar. J. 2010, 9, 87. [Google Scholar] [CrossRef] [Green Version]
- Lyke, K.E.; Burges, R.; Cissoko, Y.; Sangare, L.; Dao, M.; Diarra, I.; Kone, A.; Harley, R.; Plowe, C.V.; Doumbo, O.K.; et al. Serum levels of the proinflammatory cytokines interleukin-1 beta (IL-1beta), IL-6, IL-8, IL-10, tumor necrosis factor alpha, and IL-12(p70) in Malian children with severe Plasmodium falciparum malaria and matched uncomplicated malaria or healthy controls. Infect. Immun. 2004, 72, 5630–5637. [Google Scholar] [CrossRef] [Green Version]
- Nemeth, E.; Tuttle, M.S.; Powelson, J.; Vaughn, M.B.; Donovan, A.; Ward, D.M.V.; Ganz, T.; Kaplan, J. Hepcidin Regulates Cellular Iron Efflux by Binding to Ferroportin and Inducing Its Internalization. Science 2004, 306, 2090–2093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haldar, K.; Mohandas, N. Malaria, erythrocytic infection, and anemia. Hematol. Am. Soc. Hematol. Educ. Program 2009, 1, 87–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaiyaroj, S.C.; Rutta, A.S.; Muenthaisong, K.; Watkins, P.; Na Ubol, M.; Looareesuwan, S. Reduced levels of transforming growth factor-β1, interleukin-12 and increased migration inhibitory factor are associated with severe malaria. Acta Trop. 2004, 89, 319–327. [Google Scholar] [CrossRef]
- McDevitt, M.A.; Xie, J.; Ganapathy-Kanniappan, S.; Griffith, J.; Liu, A.; McDonald, C.; Thuma, P.; Gordeuk, V.R.; Metz, C.N.; Mitchell, R.; et al. A critical role for the host mediator macrophage migration inhibitory factor in the pathogenesis of malarial anemia. J. Exp. Med. 2006, 203, 1185–1196. [Google Scholar] [CrossRef] [PubMed]
- Awandare, G.A.; Kempaiah, P.; Ochiel, D.O.; Piazza, P.; Keller, C.C.; Perkins, D.J. Mechanisms of erythropoiesis inhibition by malarial pigment and malaria-induced proinflammatory mediators in an in vitro model. Am. J. Hematol. 2010, 86, 155–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Were, T.; Hittner, J.B.; Ouma, C.; Otieno, O.R.; Orago, A.S.S.; Ong’Echa, J.M.; Vulule, J.M.; Keller, C.C.; Perkins, D.J. Suppression of RANTES in children with Plasmodium falciparum malaria. Haematologica 2006, 91, 4. [Google Scholar]
- Broxmeyer, H.E.; Sherry, B.; Lu, L.; Cooper, S.; Oh, K.O.; Tekamp-Olson, P.; Kwon, B.S.; Cerami, A. Enhancing and suppressing effects of recombinant murine macrophage inflammatory proteins on colony formation in vitro by bone marrow myeloid progenitor cells. Blood 1990, 76, 1110–1116. [Google Scholar] [CrossRef]
- Majka, M.; Janowska-Wieczorek, A.; Ratajczak, J.; Ehrenman, K.; Pietrzkowski, Z.; Kowalska, M.A.; Gewirtz, A.M.; Emerson, S.G.; Ratajczak, M.Z. Numerous growth factors, cytokines, and chemokines are secreted by human CD34(+) cells, myeloblasts, erythroblasts, and megakaryoblasts and regulate normal hematopoiesis in an autocrine/paracrine manner. Blood 2001, 97, 3075–3085. [Google Scholar] [CrossRef] [Green Version]
- Cokić, V.P.; Schechter, A.N. Effects of nitric oxide on red blood cell development and phenotype. Curr. Top. Dev. Biol. 2008, 82, 169–215. [Google Scholar]
- Shami, P.; Weinberg, J. Differential effects of nitric oxide on erythroid and myeloid colony growth from CD34+ human bone marrow cells. Blood 1996, 87, 977–982. [Google Scholar] [CrossRef] [Green Version]
- Perkins, D.J.; Kremsner, P.G.; Schmid, D.; Misukonis, M.A.; Kelly, M.A.; Weinberg, J.B. Blood Mononuclear Cell Nitric Oxide Production and Plasma Cytokine Levels in Healthy Gabonese Children with Prior Mild or Severe Malaria. Infect. Immun. 1999, 67, 4977–4981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awandare, G.A.; Goka, B.; Boeuf, P.; Tetteh, J.K.A.; Kurtzhals, J.A.L.; Behr, C.; Akanmori, B.D. Increased Levels of Inflammatory Mediators in Children with Severe Plasmodium falciparum Malaria with Respiratory Distress. J. Infect. Dis. 2006, 194, 1438–1446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbett, Y.; Parapini, S.; Perego, F.; Messina, V.; Delbue, S.; Misiano, P.; Falchi, M.; Silvestrini, F.; Taramelli, D.; Basilico, N.; et al. Phagocytosis and activation of bone marrow-derived macrophages by Plasmodium falciparum gametocytes. Malar. J. 2021, 20, 81. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dumarchey, A.; Lavazec, C.; Verdier, F. Erythropoiesis and Malaria, a Multifaceted Interplay. Int. J. Mol. Sci. 2022, 23, 12762. https://doi.org/10.3390/ijms232112762
Dumarchey A, Lavazec C, Verdier F. Erythropoiesis and Malaria, a Multifaceted Interplay. International Journal of Molecular Sciences. 2022; 23(21):12762. https://doi.org/10.3390/ijms232112762
Chicago/Turabian StyleDumarchey, Aurélie, Catherine Lavazec, and Frédérique Verdier. 2022. "Erythropoiesis and Malaria, a Multifaceted Interplay" International Journal of Molecular Sciences 23, no. 21: 12762. https://doi.org/10.3390/ijms232112762
APA StyleDumarchey, A., Lavazec, C., & Verdier, F. (2022). Erythropoiesis and Malaria, a Multifaceted Interplay. International Journal of Molecular Sciences, 23(21), 12762. https://doi.org/10.3390/ijms232112762