
Mucosal Adjuvants Delivered by a Mucoadhesive Patch for Sublingual Administration of Subunit Vaccines

Abstract
:
1. Introduction
2. Results and Discussion
2.1. Mucoadhesive SL Formulation for NP Delivery
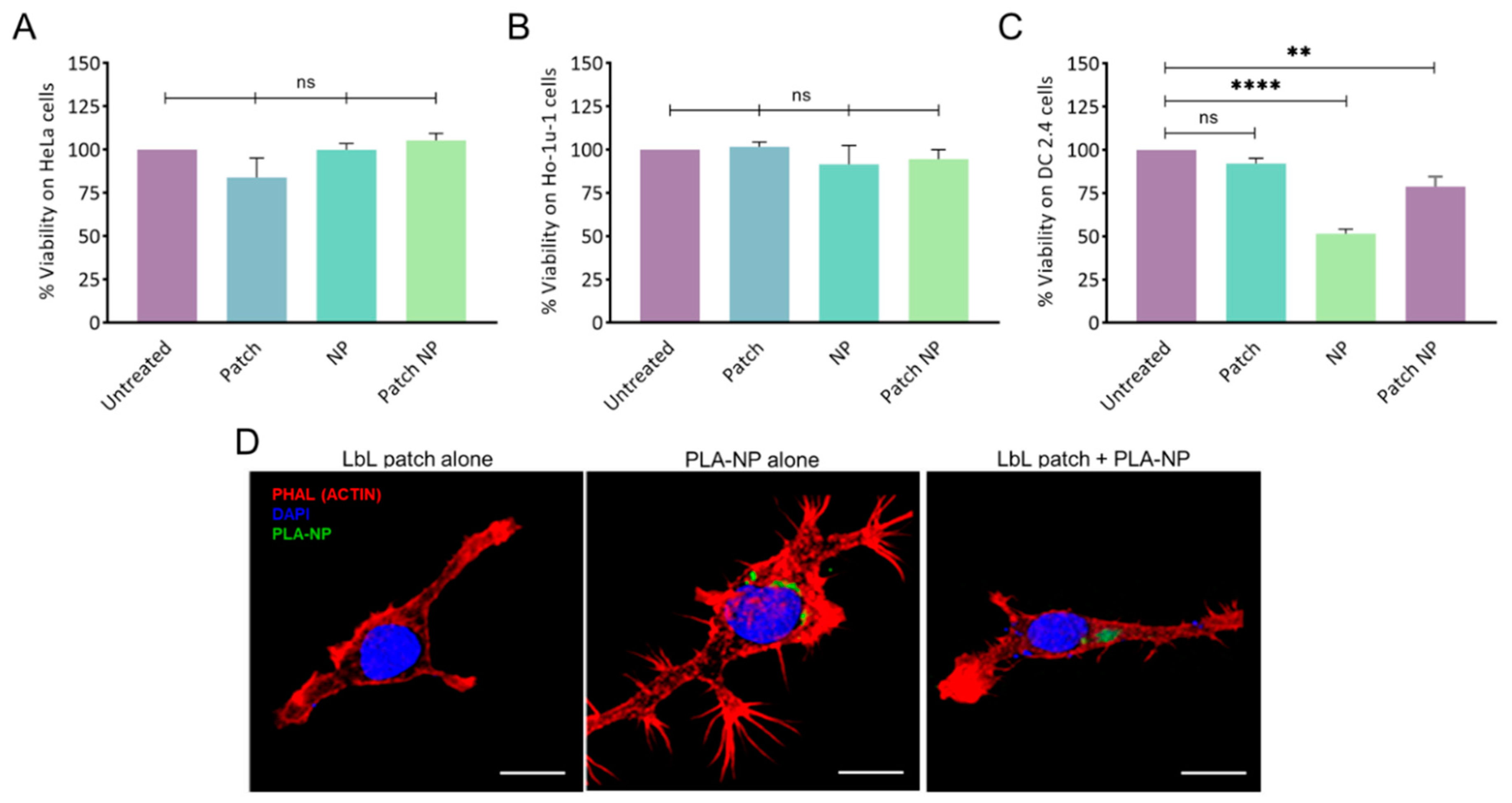
2.2. Cytotoxicity
2.3. NP Release from the LbL Patch and Uptake by Immune Cells
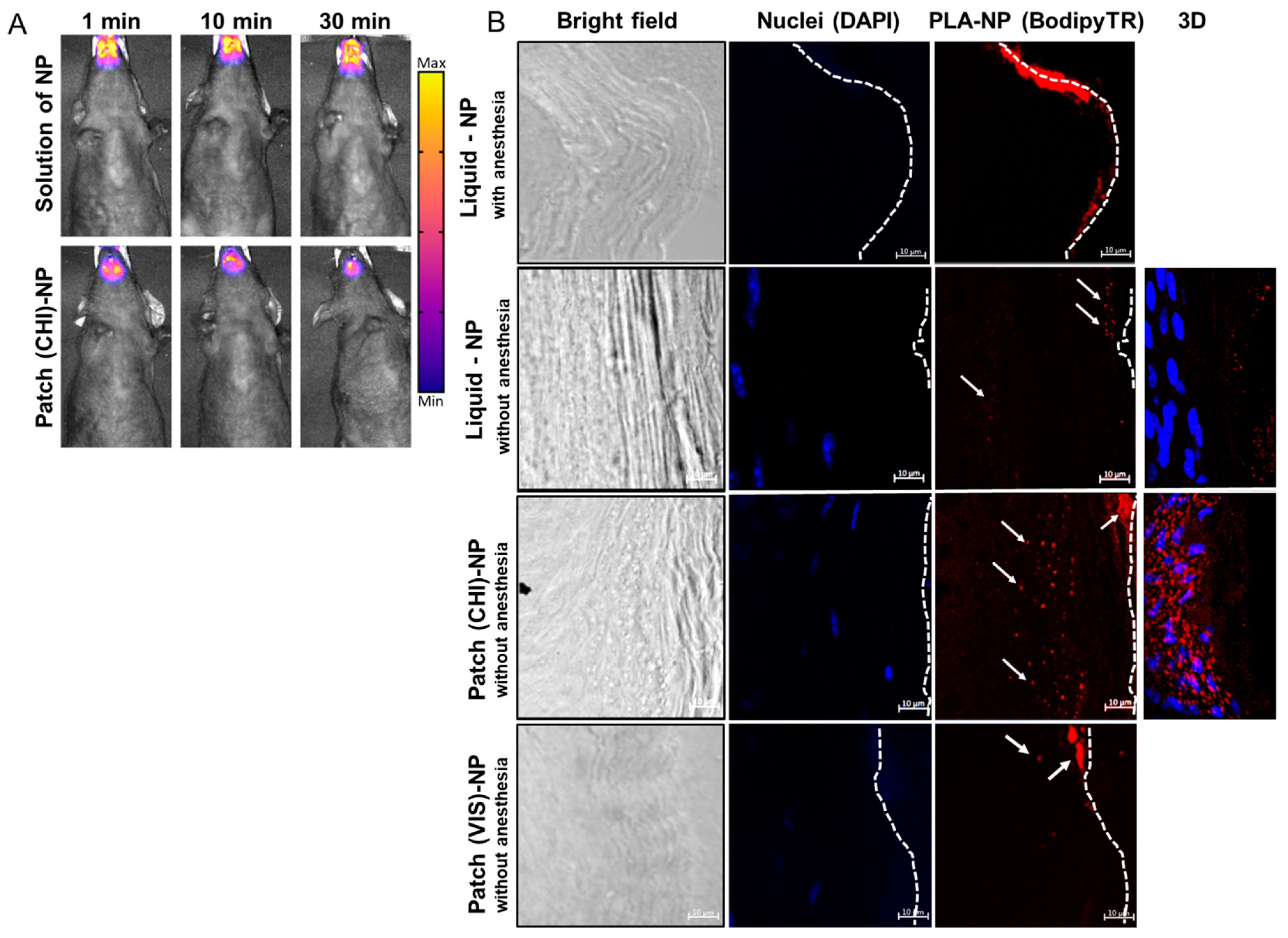
2.4. PLA-NP Transport across Sublingual Mucosa
2.5. Bioactivity of Adjuvants after Release from the LbL Patch
2.6. Systemic Inflammatory Response to Primary Sublingual Immunization with a Subunit Vaccine Formulation
3. Materials and Methods
3.1. Material
3.2. Adjuvant Formulation: Encapsulation into PLA-NPs
3.3. LbL Patch Production and PLA-NP Adsorption
3.4. Cell Culture
3.5. Cytotoxicity
3.6. Bioactivity of Cholera Toxin after Incorporation into the Mucoadhesive Patch
3.7. Activation of Innate Immune Receptors by Encapsulated Adjuvants
3.8. Animals
3.9. Nod2 Staining in SL Mucosa
3.10. In Vivo SL Administration of Formulations
3.11. Cytokine Quantification by Multiplex Assay
3.12. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lycke, N. Recent progress in mucosal vaccine development: Potential and limitations. Nat. Rev. Immunol. 2012, 12, 592–605. [Google Scholar] [CrossRef] [PubMed]
- Correa, V.A.; Portilho, A.I.; De Gaspari, E. Vaccines, adjuvants and key factors for mucosal immune response. Immunology 2022. [Google Scholar] [CrossRef] [PubMed]
- Bekri, S.; Bourdely, P.; Luci, C.; Dereuddre-Bosquet, N.; Su, B.; Martinon, F.; Braud, V.M.; Luque, I.; Mateo, P.L.; Crespillo, S.; et al. Sublingual Priming with a HIV gp41-Based Subunit Vaccine Elicits Mucosal Antibodies and Persistent B Memory Responses in Non-Human Primates. Front. Immunol. 2017, 8, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, A.T.; Shen, X.; Walter, K.L.; Labranche, C.C.; Wyatt, L.S.; Tomaras, G.D.; Montefiori, D.C.; Moss, B.; Barouch, D.H.; Clements, J.D.; et al. HIV-1 vaccination by needle-free oral injection induces strong mucosal immunity and protects against SHIV challenge. Nat. Commun. 2019, 10, 798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hameed, S.A.; Paul, S.; Dellosa, G.K.Y.; Jaraquemada, D.; Bello, M.B. Towards the future exploration of mucosal mRNA vaccines against emerging viral diseases; lessons from existing next-generation mucosal vaccine strategies. Npj Vaccines 2022, 7, 71. [Google Scholar] [CrossRef]
- Huang, M.; Zhang, M.; Zhu, H.; Du, X.; Wang, J. Mucosal vaccine delivery: A focus on the breakthrough of specific barriers. Acta Pharm. Sin. B 2022, 12, 3456–3474. [Google Scholar] [CrossRef]
- Paris, A.; Colomb, E.; Verrier, B.; Anjuère, F.; Monge, C. Sublingual vaccination and delivery systems. J. Control. Release 2021, 332, 553–562. [Google Scholar] [CrossRef]
- Trincado, V.; Gala, R.P.; Morales, J.O. Buccal and Sublingual Vaccines: A Review on Oral Mucosal Immunization and Delivery Systems. Vaccines 2021, 9, 1177. [Google Scholar] [CrossRef]
- Rio, L.G.; Diaz-Rodriguez, P.; Pedersen, G.K.; Christensen, D.; Landin, M. Sublingual Boosting with a Novel Mucoadhesive Thermogelling Hydrogel Following Parenteral CAF01 Priming as a Strategy against Chlamydia trachomatis. Adv. Health Mater. 2022, 11, 2102508. [Google Scholar] [CrossRef]
- Paris, A.-L.; Caridade, S.; Colomb, E.; Bellina, M.; Boucard, E.; Verrier, B.; Monge, C. Sublingual protein delivery by a mucoadhesive patch made of natural polymers. Acta Biomater. 2021, 128, 222–235. [Google Scholar] [CrossRef]
- Pulendran, B.; Arunachalam, P.S.; O’Hagan, D.T. Emerging concepts in the science of vaccine adjuvants. Nat. Rev. Drug Discov. 2021, 20, 454–475. [Google Scholar] [CrossRef] [PubMed]
- Bergamaschi, C.; Terpos, E.; Rosati, M.; Angel, M.; Bear, J.; Stellas, D.; Karaliota, S.; Apostolakou, F.; Bagratuni, T.; Patseas, D.; et al. Systemic IL-15, IFN-γ and IP-10/CXCL10 Signature Associated with Effective Immune Response to SARS-CoV-2 in BNT162b2 mRNA Vaccine Recipients. Cell Rep. 2021, 36, 109504. [Google Scholar] [CrossRef] [PubMed]
- Clements, J.D.; Norton, E.B. The Mucosal Vaccine Adjuvant LT(R192G/L211A) or dmLT. mSphere 2018, 3, e00215-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, J.A.; Blum, J.S.; A Hosken, N.; O Marshak, J.; Duncan, L.; Zhu, C.; Norton, E.B.; Clements, J.D.; Koelle, D.M.; Chen, D.; et al. Serum and mucosal antibody responses to inactivated polio vaccine after sublingual immunization using a thermoresponsive gel delivery system. Hum. Vaccines Immunother. 2014, 10, 3611–3621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebensen, T.; DeBarry, J.; Pedersen, G.; Blazejewska, P.; Weissmann, S.; Schulze, K.; McCullough, K.C.; Cox, R.J.; Guzmán, C.A. Mucosal Administration of Cycle-Di-Nucleotide-Adjuvanted Virosomes Efficiently Induces Protection against Influenza H5N1 in Mice. Front. Immunol. 2017, 8, 1223. [Google Scholar] [CrossRef]
- Singh, S.; Yang, G.; Byrareddy, S.N.; Barry, M.A.; Sastry, K.J. Natural killer T cell and TLR9 agonists as mucosal adjuvants for sublingual vaccination with clade C HIV-1 envelope protein. Vaccine 2014, 32, 6934–6940. [Google Scholar] [CrossRef] [Green Version]
- Jurk, M.; Heil, F.; Vollmer, J.; Schetter, C.; Krieg, A.M.; Wagner, H.; Lipford, G.B.; Bauer, S. Human TLR7 or TLR8 independently confer responsiveness to the antiviral compound R-848. Nat. Immunol. 2002, 3, 499. [Google Scholar] [CrossRef]
- Smirnov, D.; Schmidt, J.J.; Capecchi, J.T.; Wightman, P.D. Vaccine adjuvant activity of 3M-052: An imidazoquinoline designed for local activity without systemic cytokine induction. Vaccine 2011, 29, 5434–5442. [Google Scholar] [CrossRef]
- Guryanova, S.V.; Khaitov, R.M. Strategies for Using Muramyl Peptides—Modulators of Innate Immunity of Bacterial Origin—In Medicine. Front. Immunol. 2021, 12, 607178. [Google Scholar] [CrossRef]
- Pavot, V.; Climent, N.; Rochereau, N.; Garcia, F.; Genin, C.; Tiraby, G.; Vernejoul, F.; Perouzel, E.; Lioux, T.; Verrier, B.; et al. Directing vaccine immune responses to mucosa by nanosized particulate carriers encapsulating NOD ligands. Biomaterials 2016, 75, 327–339. [Google Scholar] [CrossRef]
- Kasturi, S.P.; Rasheed, M.A.U.; Havenar-Daughton, C.; Pham, M.; Legere, T.; Sher, Z.J.; Kovalenkov, Y.; Gumber, S.; Huang, J.Y.; Gottardo, R.; et al. 3M-052, a synthetic TLR-7/8 agonist, induces durable HIV-1 envelope–specific plasma cells and humoral immunity in nonhuman primates. Sci. Immunol. 2020, 5, eabb1025. [Google Scholar] [CrossRef]
- Pavot, V.; Berthet, M.; Resseguier, J.; Legaz, S.; Handké, N.; Gilbert, S.C.; Paul, S.; Verrier, B. Poly (lactic acid) and poly (lactic-co-glycolic acid) particles as versatile carrier platforms for vaccine delivery. Nanomedicine 2014, 9, 2703–2718. [Google Scholar] [CrossRef] [PubMed]
- Legaz, S.; Exposito, J.-Y.; Lethias, C.; Viginier, B.; Terzian, C.; Verrier, B. Evaluation of polylactic acid nanoparticles safety using Drosophila model. Nanotoxicology 2016, 10, 1136–1143. [Google Scholar] [CrossRef]
- Anjuère, F.; Bekri, S.; Bihl, F.; Braud, V.; Cuburu, N.; Czerkinsky, C.; Hervouet, C.; Luci, C. B cell and T cell immunity in the female genital tract: Potential of distinct mucosal routes of vaccination and role of tissue-associated dendritic cells and natural killer cells. Clin. Microbiol. Infect. 2012, 18, 117–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mašek, J.; Lubasová, D.; Lukáč, R.; Turánek-Knotigová, P.; Kulich, P.; Plocková, J.; Mašková, E.; Procházka, L.; Koudelka, Š.; Sasithorn, N.; et al. Multi-layered nanofibrous mucoadhesive films for buccal and sublingual administration of drug-delivery and vaccination nanoparticles—Important step towards effective mucosal vaccines. J. Control. Release 2017, 249, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.; Wood, E.; Dornish, M. Effect of Chitosan on Epithelial Cell Tight Junctions. Pharm. Res. 2004, 21, 43–49. [Google Scholar] [CrossRef]
- Le Borgne, M.; Etchart, N.; Goubier, A.; Lira, S.A.; Sirard, J.C.; van Rooijen, N.; Caux, C.; Aït-Yahia, S.; Vicari, A.; Kaiserlian, D.; et al. Dendritic Cells Rapidly Recruited into Epithelial Tissues via CCR6/CCL20 Are Responsible for CD8+ T Cell Crosspriming In Vivo. Immunity 2006, 24, 191–201. [Google Scholar] [CrossRef]
- Gutjahr, A.; Papagno, L.; Vernejoul, F.; Lioux, T.; Jospin, F.; Chanut, B.; Perouzel, E.; Rochereau, N.; Appay, V.; Verrier, B.; et al. New chimeric TLR7/NOD2 agonist is a potent adjuvant to induce mucosal immune responses. eBioMedicine 2020, 58, 102922. [Google Scholar] [CrossRef]
- Singh, S.; Anshita, D.; Ravichandiran, V. MCP-1: Function, regulation, and involvement in disease. Int. Immunopharmacol. 2021, 101, 107598. [Google Scholar] [CrossRef]
- McDonald, J.U.; Zhong, Z.; Groves, H.T.; Tregoning, J.S. Inflammatory responses to influenza vaccination at the extremes of age. Immunology 2017, 151, 451–463. [Google Scholar] [CrossRef]
- Datta, S.K.; Sabet, M.; Nguyen, K.P.L.; Valdez, P.A.; Gonzalez-Navajas, J.M.; Islam, S.; Mihajlov, I.; Fierer, J.; Insel, P.A.; Webster, N.J.; et al. Mucosal adjuvant activity of cholera toxin requires Th17 cells and protects against inhalation anthrax. Proc. Natl. Acad. Sci. USA 2010, 107, 10638–10643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattsson, J.; Schön, K.; Ekman, L.; Fahlén-Yrlid, L.; Yrlid, U.; Lycke, N.Y. Cholera toxin adjuvant promotes a balanced Th1/Th2/Th17 response independently of IL-12 and IL-17 by acting on Gsα in CD11b+ DCs. Mucosal Immunol. 2014, 8, 815–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.; Ivashkiv, L.B. Cross-regulation of Signaling Pathways by Interferon-γ: Implications for Immune Responses and Autoimmune Diseases. Immunity 2009, 31, 539–550. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Encapsulated Molecule +/− p24 Adsorption | Diameter (Z-Average, nm) | PDI | Zeta Potential (mV) |
|---|---|---|---|
| Bodipy TR | 176.3 ± 1.8 | 0.051 ± 0.022 | −59.1 ± 1.1 |
| Bodipy TR + p24 | 210.5 ± 5.5 | 0.111 ± 0.035 | −51.6 ± 1.1 |
| Telratolimod | 134.6 ± 0.1 | 0.127 ± 0.083 | −59.9 ± 2.4 |
| Telratolimod + p24 | 155.5 ± 0.8 | 0.175 ± 0.036 | −45.4 ± 0.8 |
| Mifamurtide | 158.7 ± 0.7 | 0.053 ± 0.007 | −60.0 ± 0.5 |
| Mifamurtide + p24 | 165.0 ± 0.4 | 0.057 ± 0.010 | −53.8 ± 1.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monge, C.; Ayad, C.; Paris, A.-L.; Rovera, R.; Colomb, E.; Verrier, B. Mucosal Adjuvants Delivered by a Mucoadhesive Patch for Sublingual Administration of Subunit Vaccines. Int. J. Mol. Sci. 2022, 23, 13440. https://doi.org/10.3390/ijms232113440
Monge C, Ayad C, Paris A-L, Rovera R, Colomb E, Verrier B. Mucosal Adjuvants Delivered by a Mucoadhesive Patch for Sublingual Administration of Subunit Vaccines. International Journal of Molecular Sciences. 2022; 23(21):13440. https://doi.org/10.3390/ijms232113440
Chicago/Turabian StyleMonge, Claire, Camille Ayad, Anne-Lise Paris, Renaud Rovera, Evelyne Colomb, and Bernard Verrier. 2022. "Mucosal Adjuvants Delivered by a Mucoadhesive Patch for Sublingual Administration of Subunit Vaccines" International Journal of Molecular Sciences 23, no. 21: 13440. https://doi.org/10.3390/ijms232113440
APA StyleMonge, C., Ayad, C., Paris, A. -L., Rovera, R., Colomb, E., & Verrier, B. (2022). Mucosal Adjuvants Delivered by a Mucoadhesive Patch for Sublingual Administration of Subunit Vaccines. International Journal of Molecular Sciences, 23(21), 13440. https://doi.org/10.3390/ijms232113440